NKCC1, an Elusive Molecular Target in Brain Development: Making Sense of the Existing Data



Abstract
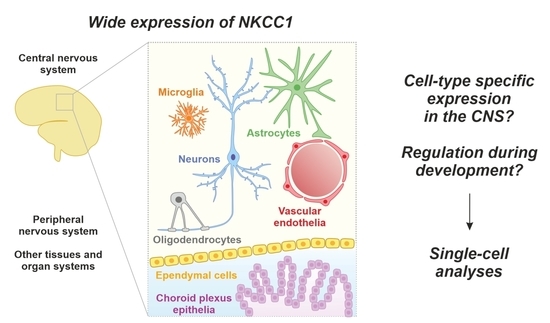
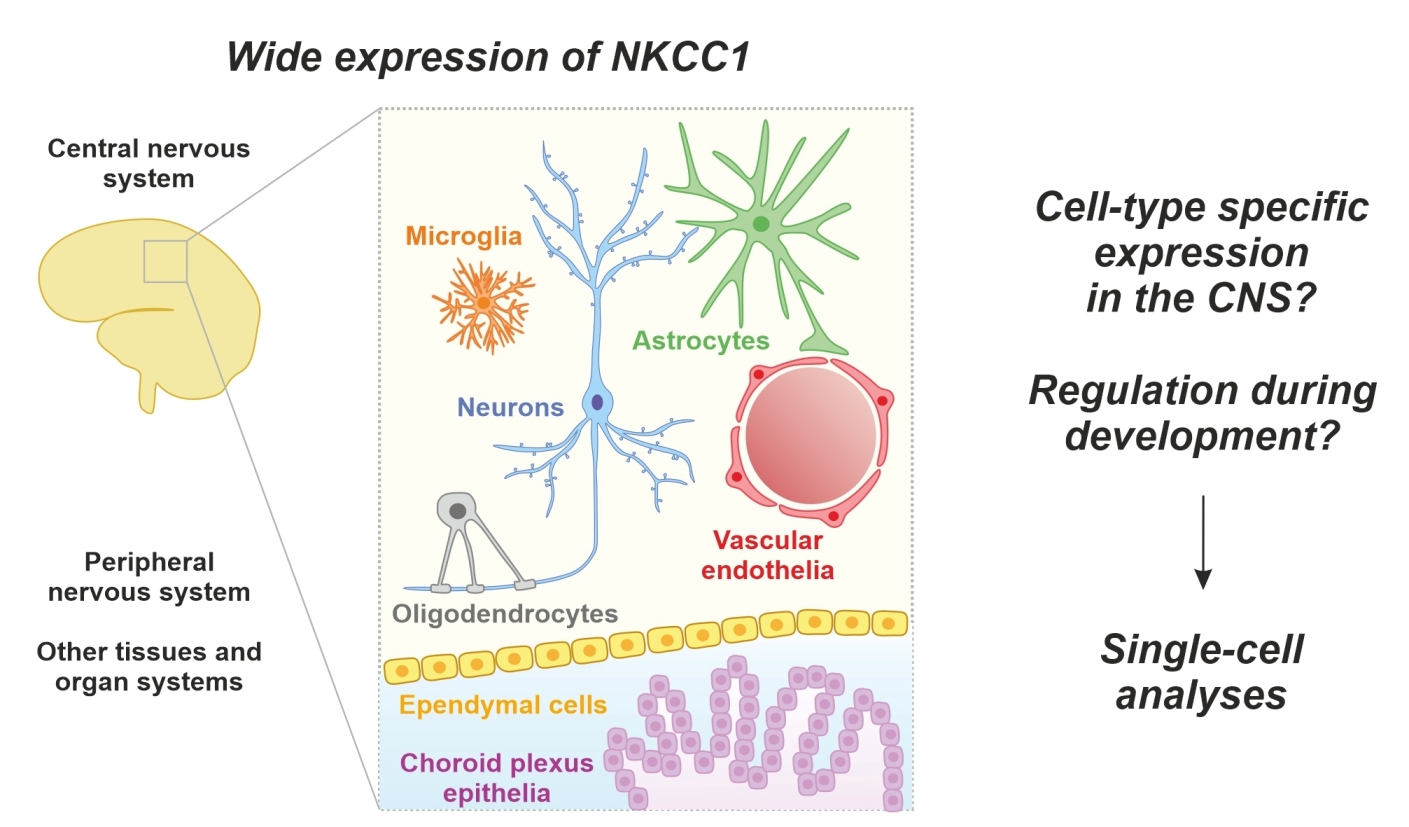
1. Introduction
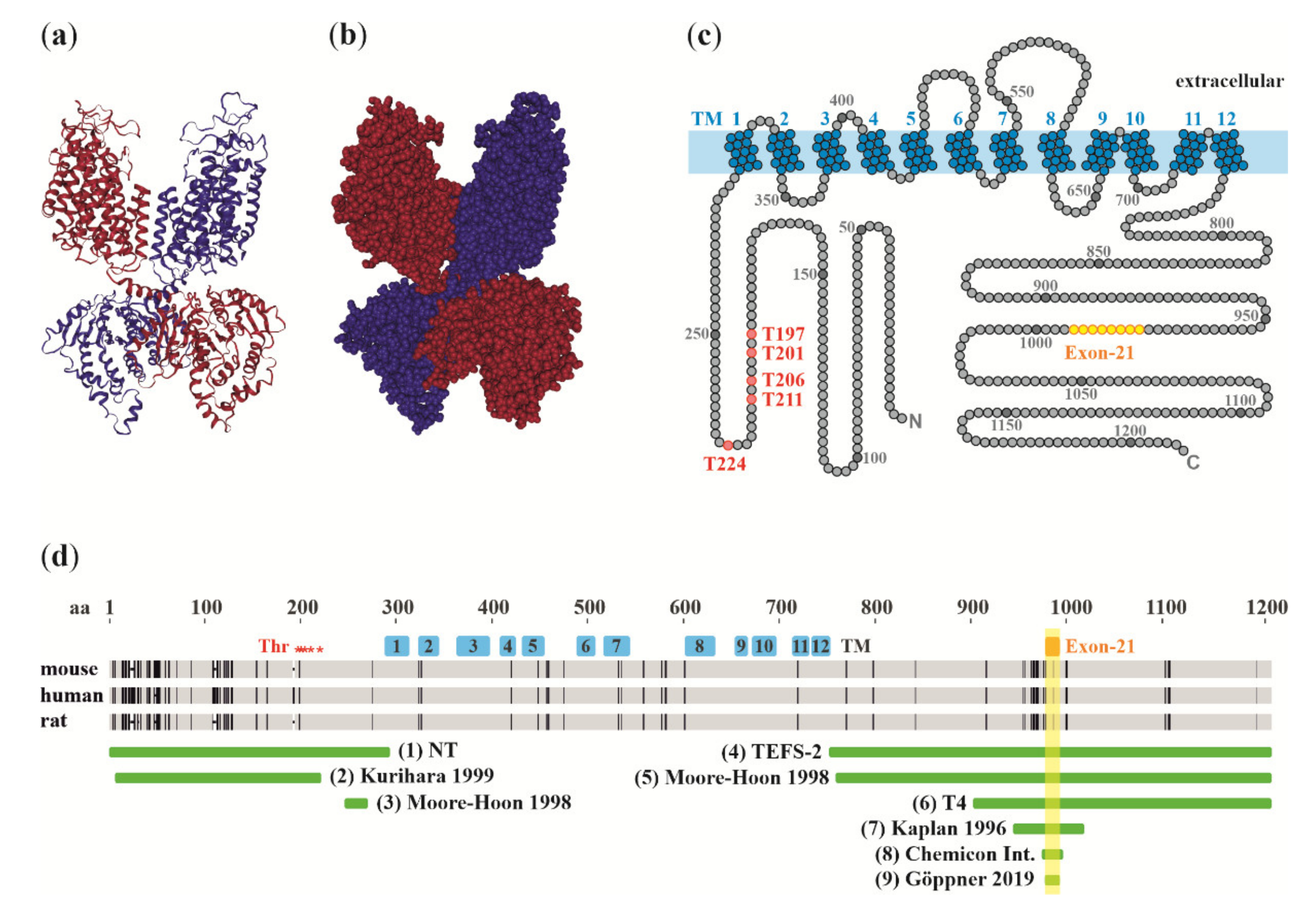
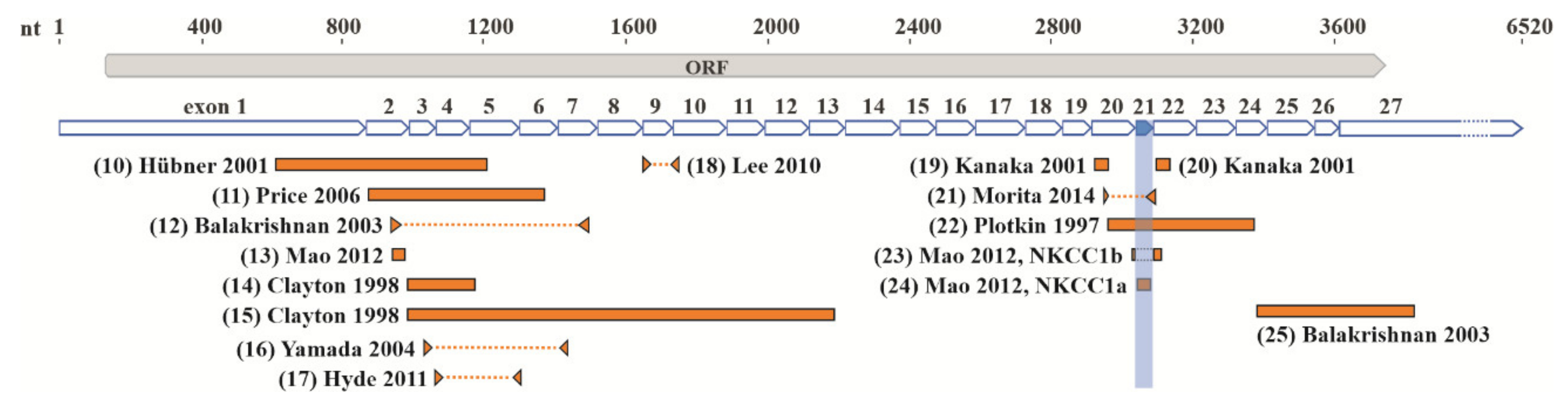
2. Molecular Structure of NKCC1
3. NKCC1 Functions outside the CNS
4. Functional Data Show Cl− Uptake by NKCC1 in CNS Neurons
5. NKCC1 Protein Levels and Localization in the CNS
6. NKCC1 mRNA Expression in the CNS
7. NKCC1 Expression during Neuronal Development
8. Implications for Future Work
Funding
Conflicts of Interest
References
- Blaesse, P.; Airaksinen, M.S.; Rivera, C.; Kaila, K. Cation-chloride cotransporters and neuronal function. Neuron 2009, 61, 820–838. [Google Scholar] [CrossRef] [PubMed]
- Gamba, G.; Miyanoshita, A.; Lombardi, M.; Lytton, J.; Lee, W.S.; Hediger, M.A.; Hebert, S.C. Molecular cloning, primary structure, and characterization of two members of the mammalian electroneutral sodium-(potassium)-chloride cotransporter family expressed in kidney. J. Biol. Chem. 1994, 269, 17713–17722. [Google Scholar]
- Payne, J.A.; Stevenson, T.J.; Donaldson, L.F. Molecular characterization of a putative K-Cl cotransporter in rat brain. A neuronal-specific isoform. J. Biol. Chem. 1996, 271, 16245–16252. [Google Scholar] [CrossRef] [PubMed]
- Rivera, C.; Voipio, J.; Payne, J.A.; Ruusuvuori, E.; Lahtinen, H.; Lamsa, K.; Pirvola, U.; Saarma, M.; Kaila, K. The K+/Cl− co-transporter KCC2 renders GABA hyperpolarizing during neuronal maturation. Nature 1999, 397, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Kaila, K.; Price, T.J.; Payne, J.A.; Puskarjov, M.; Voipio, J. Cation-chloride cotransporters in neuronal development, plasticity and disease. Nat. Rev. Neurosci. 2014, 15, 637–654. [Google Scholar] [CrossRef]
- Llano, O.; Rivera, C.; Ludwig, A. KCC2 regulates dendritic spine development. In Neuronal Chloride Transporters in Health and Disease; Tang, X., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 103–132. [Google Scholar]
- Chevy, Q.; Simonnet, C.; Al Awabdh, S.; Lévi, S.; Poncer, J.C. Transport-dependent and independent functions of KCC2 at excitatory synapses. In Neuronal Chloride Transporters in Health and Disease; Tang, X., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 133–158. [Google Scholar]
- Sedmak, G.; Jovanov-Milosevic, N.; Puskarjov, M.; Ulamec, M.; Kruslin, B.; Kaila, K.; Judas, M. Developmental Expression Patterns of KCC2 and Functionally Associated Molecules in the Human Brain. Cereb. Cortex 2016, 26, 4574–4589. [Google Scholar] [CrossRef] [PubMed]
- Vanhatalo, S.; Palva, J.M.; Andersson, S.; Rivera, C.; Voipio, J.; Kaila, K. Slow endogenous activity transients and developmental expression of K+-Cl- cotransporter 2 in the immature human cortex. Eur. J. Neurosci. 2005, 22, 2799–2804. [Google Scholar] [CrossRef]
- Ben-Ari, Y. NKCC1 Chloride Importer Antagonists Attenuate Many Neurological and Psychiatric Disorders. Trends Neurosci. 2017, 40, 536–554. [Google Scholar] [CrossRef]
- Savardi, A.; Borgogno, M.; Narducci, R.; La Sala, G.; Ortega, J.A.; Summa, M.; Armirotti, A.; Bertorelli, R.; Contestabile, A.; De Vivo, M.; et al. Discovery of a Small Molecule Drug Candidate for Selective NKCC1 Inhibition in Brain Disorders. Chem 2020, 6, 2073–2096. [Google Scholar] [CrossRef]
- Luo, L.; Wang, J.; Ding, D.; Hasan, M.N.; Yang, S.S.; Lin, S.H.; Schreppel, P.; Sun, B.; Yin, Y.; Erker, T.; et al. Role of NKCC1 Activity in Glioma K+ Homeostasis and Cell Growth: New Insights with the Bumetanide-Derivative STS66. Front. Physiol. 2020, 11, 911. [Google Scholar] [CrossRef]
- Brandt, C.; Seja, P.; Töllner, K.; Römermann, K.; Hampel, P.; Kalesse, M.; Kipper, A.; Feit, P.W.; Lykke, K.; Toft-Bertelsen, T.L.; et al. Bumepamine, a brain-permeant benzylamine derivative of bumetanide, does not inhibit NKCC1 but is more potent to enhance phenobarbital’s anti-seizure efficacy. Neuropharmacology 2018, 143, 186–204. [Google Scholar] [CrossRef] [PubMed]
- Henneberger, C.; Bard, L.; Panatier, A.; Reynolds, J.P.; Kopach, O.; Medvedev, N.I.; Minge, D.; Herde, M.K.; Anders, S.; Kraev, I.; et al. LTP Induction Boosts Glutamate Spillover by Driving Withdrawal of Perisynaptic Astroglia. Neuron 2020. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, M.E.; Tran, L.; Lam, T.I.; Liu, X.B.; Anderson, S.E. Bumetanide inhibition of the blood-brain barrier Na-K-Cl cotransporter reduces edema formation in the rat middle cerebral artery occlusion model of stroke. J. Cereb. Blood Flow Metab. 2004, 24, 1046–1056. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Dempsey, R.J.; Flemmer, A.; Forbush, B.; Sun, D. Inhibition of Na+-K+-Cl- cotransporter during focal cerebral ischemia decreases edema and neuronal damage. Brain Res. 2003, 961, 22–31. [Google Scholar] [CrossRef]
- Zhang, J.; Pu, H.; Zhang, H.; Wei, Z.; Jiang, X.; Xu, M.; Zhang, L.; Zhang, W.; Liu, J.; Meng, H.; et al. Inhibition of Na+-K+-2Cl− cotransporter attenuates blood-brain-barrier disruption in a mouse model of traumatic brain injury. Neurochem. Int. 2017, 111, 23–31. [Google Scholar] [CrossRef]
- Lenart, B.; Kintner, D.B.; Shull, G.E.; Sun, D. Na-K-Cl cotransporter-mediated intracellular Na+ accumulation affects Ca2+ signaling in astrocytes in an in vitro ischemic model. J. Neurosci. 2004, 24, 9585–9597. [Google Scholar] [CrossRef]
- Parvin, M.N.; Gerelsaikhan, T.; Turner, R.J. Regions in the cytosolic C-terminus of the secretory Na(+)-K(+)-2Cl(-) cotransporter NKCC1 are required for its homodimerization. Biochemistry 2007, 46, 9630–9637. [Google Scholar] [CrossRef]
- Xu, J.C.; Lytle, C.; Zhu, T.T.; Payne, J.A.; Benz, E.J.; Forbush, B. Molecular cloning and functional expression of the bumetanide-sensitive Na-K-Cl cotransporter. Proc. Natl. Acad. Sci. USA 1994, 91, 2201–2205. [Google Scholar] [CrossRef]
- Gerelsaikhan, T.; Parvin, M.N.; Turner, R.J. Biogenesis and topology of the secretory Na+-K+-2Cl- cotransporter (NKCC1) studied in intact mammalian cells. Biochemistry 2006, 45, 12060–12067. [Google Scholar] [CrossRef]
- Gerelsaikhan, T.; Turner, R.J. Transmembrane topology of the secretory Na+-K+-2Cl- cotransporter NKCC1 studied by in vitro translation. J. Biol. Chem. 2000, 275, 40471–40477. [Google Scholar] [CrossRef]
- Yang, X.; Wang, Q.; Cao, E. Structure of the human cation-chloride cotransporter NKCC1 determined by single-particle electron cryo-microscopy. Nat. Commun. 2020, 11, 1016. [Google Scholar] [CrossRef] [PubMed]
- Chew, T.A.; Orlando, B.J.; Zhang, J.; Latorraca, N.R.; Wang, A.; Hollingsworth, S.A.; Chen, D.H.; Dror, R.O.; Liao, M.; Feng, L. Structure and mechanism of the cation-chloride cotransporter NKCC1. Nature 2019, 572, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Rose, A.S.; Bradley, A.R.; Valasatava, Y.; Duarte, J.M.; Prlic, A.; Rose, P.W. NGL viewer: Web-based molecular graphics for large complexes. Bioinformatics 2018, 34, 3755–3758. [Google Scholar] [CrossRef]
- McDaniel, N.; Lytle, C. Parietal cells express high levels of Na-K-2Cl cotransporter on migrating into the gastric gland neck. Am. J. Physiol. 1999, 276, G1273–G1278. [Google Scholar] [CrossRef]
- Kurihara, K.; Moore-Hoon, M.L.; Saitoh, M.; Turner, R.J. Characterization of a phosphorylation event resulting in upregulation of the salivary Na+-K+-2Cl- cotransporter. Am. J. Physiol. 1999, 277, C1184–C1193. [Google Scholar] [CrossRef]
- Moore-Hoon, M.L.; Turner, R.J. Molecular and topological characterization of the rat parotid Na+-K+-2Cl- cotransporter1. Biochim. Biophys. Acta. 1998, 1373, 261–269. [Google Scholar] [CrossRef]
- Del Castillo, I.C.; Fedor-Chaiken, M.; Song, J.C.; Starlinger, V.; Yoo, J.; Matlin, K.S.; Matthews, J.B. Dynamic regulation of Na+-K+-2Cl- cotransporter surface expression by PKC-{epsilon} in Cl--secretory epithelia. Am. J. Physiol. Cell Physiol. 2005, 289, C1332–C1343. [Google Scholar] [CrossRef]
- Lytle, C.; Xu, J.C.; Biemesderfer, D.; Forbush, B. Distribution and diversity of Na-K-Cl cotransport proteins: A study with monoclonal antibodies. Am. J. Physiol. 1995, 269, C1496–C1505. [Google Scholar] [CrossRef]
- Kaplan, M.R.; Plotkin, M.D.; Brown, D.; Hebert, S.C.; Delpire, E. Expression of the mouse Na-K-2Cl cotransporter, mBSC2, in the terminal inner medullary collecting duct, the glomerular and extraglomerular mesangium, and the glomerular afferent arteriole. J. Clin. Investig. 1996, 98, 723–730. [Google Scholar] [CrossRef]
- He, X.; Tse, C.M.; Donowitz, M.; Alper, S.L.; Gabriel, S.E.; Baum, B.J. Polarized distribution of key membrane transport proteins in the rat submandibular gland. Pflugers Arch. 1997, 433, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Göppner, C.; Orozco, I.J.; Hoegg-Beiler, M.B.; Soria, A.H.; Hübner, C.A.; Fernandes-Rosa, F.L.; Boulkroun, S.; Zennaro, M.C.; Jentsch, T.J. Pathogenesis of hypertension in a mouse model for human CLCN2 related hyperaldosteronism. Nat. Commun. 2019, 10, 4678. [Google Scholar] [CrossRef] [PubMed]
- Randall, J.; Thorne, T.; Delpire, E. Partial cloning and characterization of Slc12a2: The gene encoding the secretory Na+-K+-2Cl- cotransporter. Am. J. Physiol. 1997, 273, C1267–C1277. [Google Scholar] [CrossRef] [PubMed]
- Carmosino, M.; Gimenez, I.; Caplan, M.; Forbush, B. Exon loss accounts for differential sorting of Na-K-Cl cotransporters in polarized epithelial cells. Mol. Biol Cell 2008, 19, 4341–4351. [Google Scholar] [CrossRef] [PubMed]
- Haas, M.; Forbush, B., 3rd. The Na-K-Cl cotransporter of secretory epithelia. Annu. Rev. Physiol. 2000, 62, 515–534. [Google Scholar] [CrossRef]
- Plotkin, M.D.; Kaplan, M.R.; Peterson, L.N.; Gullans, S.R.; Hebert, S.C.; Delpire, E. Expression of the Na(+)-K(+)-2Cl- cotransporter BSC2 in the nervous system. Am. J. Physiol. 1997, 272, C173–C183. [Google Scholar] [CrossRef]
- Gregoriades, J.M.C.; Madaris, A.; Alvarez, F.J.; Alvarez-Leefmans, F.J. Genetic and pharmacological inactivation of apical Na+-K+-2Cl- cotransporter 1 in choroid plexus epithelial cells reveals the physiological function of the cotransporter. Am. J. Physiol. Cell Physiol. 2019, 316, C525–c544. [Google Scholar] [CrossRef]
- Boettger, T.; Rust, M.B.; Maier, H.; Seidenbecher, T.; Schweizer, M.; Keating, D.J.; Faulhaber, J.; Ehmke, H.; Pfeffer, C.; Scheel, O.; et al. Loss of K-Cl co-transporter KCC3 causes deafness, neurodegeneration and reduced seizure threshold. EMBO J. 2003, 22, 5422–5434. [Google Scholar] [CrossRef]
- Howard, H.C.; Mount, D.B.; Rochefort, D.; Byun, N.; Dupre, N.; Lu, J.; Fan, X.; Song, L.; Riviere, J.B.; Prevost, C.; et al. The K-Cl cotransporter KCC3 is mutant in a severe peripheral neuropathy associated with agenesis of the corpus callosum. Nat. Genet. 2002, 32, 384–392. [Google Scholar] [CrossRef]
- Delpire, E.; Gagnon, K.B. Na+-K+-2Cl- Cotransporter (NKCC) Physiological Function in Nonpolarized Cells and Transporting Epithelia. Compr. Physiol. 2018, 8, 871–901. [Google Scholar] [CrossRef]
- Bulley, S.; Jaggar, J.H. Cl- channels in smooth muscle cells. Pflugers Arch. 2014, 466, 861–872. [Google Scholar] [CrossRef]
- Heinze, C.; Seniuk, A.; Sokolov, M.V.; Huebner, A.K.; Klementowicz, A.E.; Szijártó, I.A.; Schleifenbaum, J.; Vitzthum, H.; Gollasch, M.; Ehmke, H.; et al. Disruption of vascular Ca2+-activated chloride currents lowers blood pressure. J. Clin. Investig. 2014, 124, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Zdebik, A.A.; Wangemann, P.; Jentsch, T.J. Potassium ion movement in the inner ear: Insights from genetic disease and mouse models. Physiology 2009, 24, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, E.K.; Lambert, I.H.; Pedersen, S.F. Physiology of cell volume regulation in vertebrates. Physiol. Rev. 2009, 89, 193–277. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.M. Sodium-potassium-chloride cotransport. Physiol. Rev. 2000, 80, 211–276. [Google Scholar] [CrossRef] [PubMed]
- Demian, W.L.; Persaud, A.; Jiang, C.; Coyaud, É.; Liu, S.; Kapus, A.; Kafri, R.; Raught, B.; Rotin, D. The Ion Transporter NKCC1 Links Cell Volume to Cell Mass Regulation by Suppressing mTORC1. Cell Rep. 2019, 27, 1886–1896.e1886. [Google Scholar] [CrossRef]
- Delpire, E.; Lu, J.M.; England, R.; Dull, C.; Thorne, T. Deafness and imbalance associated with inactivation of the secretory Na-K-2Cl co-transporter. Nat. Genet. 1999, 22, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Flagella, M.; Clarke, L.L.; Miller, M.L.; Erway, L.C.; Giannella, R.A.; Andringa, A.; Gawenis, L.R.; Kramer, J.; Duffy, J.J.; Doetschman, T.; et al. Mice lacking the basolateral Na-K-2Cl cotransporter have impaired epithelial chloride secretion and are profoundly deaf. J. Biol. Chem. 1999, 274, 26946–26955. [Google Scholar] [CrossRef]
- Pace, A.J.; Lee, E.; Athirakul, K.; Coffman, T.M.; O’Brien, D.A.; Koller, B.H. Failure of spermatogenesis in mouse lines deficient in the Na+-K+-2Cl- cotransporter. J. Clin. Investig. 2000, 105, 441–450. [Google Scholar] [CrossRef]
- Antoine, M.W.; Hubner, C.A.; Arezzo, J.C.; Hebert, J.M. A causative link between inner ear defects and long-term striatal dysfunction. Science 2013, 341, 1120–1123. [Google Scholar] [CrossRef]
- Barbaric, I.; Miller, G.; Dear, T.N. Appearances can be deceiving: Phenotypes of knockout mice. Brief. Funct. Genomics Proteomics 2007, 6, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Dixon, M.J.; Gazzard, J.; Chaudhry, S.S.; Sampson, N.; Schulte, B.A.; Steel, K.P. Mutation of the Na-K-Cl co-transporter gene Slc12a2 results in deafness in mice. Hum. Mol. Genet. 1999, 8, 1579–1584. [Google Scholar] [CrossRef] [PubMed]
- Sung, K.W.; Kirby, M.; McDonald, M.P.; Lovinger, D.M.; Delpire, E. Abnormal GABAA receptor-mediated currents in dorsal root ganglion neurons isolated from Na-K-2Cl cotransporter null mice. J. Neurosci. 2000, 20, 7531–7538. [Google Scholar] [CrossRef] [PubMed]
- Laird, J.M.; Garcia-Nicas, E.; Delpire, E.J.; Cervero, F. Presynaptic inhibition and spinal pain processing in mice: A possible role of the NKCC1 cation-chloride co-transporter in hyperalgesia. Neurosci. Lett. 2004, 361, 200–203. [Google Scholar] [CrossRef]
- Johnson, K.R.; Cook, S.A.; Zheng, Q.Y. The original shaker-with-syndactylism mutation (sy) is a contiguous gene deletion syndrome. Mamm Genome 1998, 9, 889–892. [Google Scholar] [CrossRef][Green Version]
- Deol, M.S. The development of the inner ear in mice homozygous for shaker-with-syndactylism. J. Embryol. Exp. Morphol. 1963, 11, 493–512. [Google Scholar]
- Macnamara, E.F.; Koehler, A.E.; D’Souza, P.; Estwick, T.; Lee, P.; Vezina, G.; Fauni, H.; Braddock, S.R.; Torti, E.; Holt, J.M.; et al. Kilquist syndrome: A novel syndromic hearing loss disorder caused by homozygous deletion of SLC12A2. Hum. Mutat. 2019, 40, 532–538. [Google Scholar] [CrossRef]
- Delpire, E.; Wolfe, L.; Flores, B.; Koumangoye, R.; Schornak, C.C.; Omer, S.; Pusey, B.; Lau, C.; Markello, T.; Adams, D.R. A patient with multisystem dysfunction carries a truncation mutation in human SLC12A2, the gene encoding the Na-K-2Cl cotransporter, NKCC1. Cold Spring Harb. Mol. Case Stud. 2016, 2, a001289. [Google Scholar] [CrossRef]
- Mutai, H.; Wasano, K.; Momozawa, Y.; Kamatani, Y.; Miya, F.; Masuda, S.; Morimoto, N.; Nara, K.; Takahashi, S.; Tsunoda, T.; et al. Variants encoding a restricted carboxy-terminal domain of SLC12A2 cause hereditary hearing loss in humans. PLoS Genet. 2020, 16, e1008643. [Google Scholar] [CrossRef]
- Merner, N.D.; Mercado, A.; Khanna, A.R.; Hodgkinson, A.; Bruat, V.; Awadalla, P.; Gamba, G.; Rouleau, G.A.; Kahle, K.T. Gain-of-function missense variant in SLC12A2, encoding the bumetanide-sensitive NKCC1 cotransporter, identified in human schizophrenia. J. Psychiatr. Res. 2016, 77, 22–26. [Google Scholar] [CrossRef]
- Anazi, S.; Maddirevula, S.; Salpietro, V.; Asi, Y.T.; Alsahli, S.; Alhashem, A.; Shamseldin, H.E.; AlZahrani, F.; Patel, N.; Ibrahim, N.; et al. Expanding the genetic heterogeneity of intellectual disability. Hum. Genet. 2017, 136, 1419–1429. [Google Scholar] [CrossRef] [PubMed]
- Stödberg, T.; Magnusson, M.; Lesko, N.; Wredenberg, A.; Martin Munoz, D.; Stranneheim, H.; Wedell, A. SLC12A2 mutations cause NKCC1 deficiency with encephalopathy and impaired secretory epithelia. Neurol Genet. 2020, 6, e478. [Google Scholar] [CrossRef] [PubMed]
- Mao, S.; Garzon-Muvdi, T.; Di Fulvio, M.; Chen, Y.; Delpire, E.; Alvarez, F.J.; Alvarez-Leefmans, F.J. Molecular and functional expression of cation-chloride cotransporters in dorsal root ganglion neurons during postnatal maturation. J. Neurophysiol. 2012, 108, 834–852. [Google Scholar] [CrossRef] [PubMed]
- Price, T.J.; Hargreaves, K.M.; Cervero, F. Protein expression and mRNA cellular distribution of the NKCC1 cotransporter in the dorsal root and trigeminal ganglia of the rat. Brain Res. 2006, 1112, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Leefmans, F.J.; Leon-Olea, M.; Mendoza-Sotelo, J.; Alvarez, F.J.; Anton, B.; Garduno, R. Immunolocalization of the Na+-K+-2Cl- cotransporter in peripheral nervous tissue of vertebrates. Neuroscience 2001, 104, 569–582. [Google Scholar] [CrossRef]
- Hübner, C.A.; Lorke, D.E.; Hermans-Borgmeyer, I. Expression of the Na-K-2Cl-cotransporter NKCC1 during mouse development. Mech. Dev. 2001, 102, 267–269. [Google Scholar] [CrossRef]
- Balakrishnan, V.; Becker, M.; Löhrke, S.; Nothwang, H.G.; Güresir, E.; Friauf, E. Expression and function of chloride transporters during development of inhibitory neurotransmission in the auditory brainstem. J. Neurosci. 2003, 23, 4134–4145. [Google Scholar] [CrossRef]
- Clayton, G.H.; Owens, G.C.; Wolff, J.S.; Smith, R.L. Ontogeny of cation-Cl- cotransporter expression in rat neocortex. Brain Res. Dev. Brain Res. 1998, 109, 281–292. [Google Scholar] [CrossRef]
- Yamada, J.; Okabe, A.; Toyoda, H.; Kilb, W.; Luhmann, H.J.; Fukuda, A. Cl- uptake promoting depolarizing GABA actions in immature rat neocortical neurones is mediated by NKCC1. J. Physiol. 2004, 557, 829–841. [Google Scholar] [CrossRef]
- Hyde, T.M.; Lipska, B.K.; Ali, T.; Mathew, S.V.; Law, A.J.; Metitiri, O.E.; Straub, R.E.; Ye, T.; Colantuoni, C.; Herman, M.M.; et al. Expression of GABA signaling molecules KCC2, NKCC1, and GAD1 in cortical development and schizophrenia. J. Neurosci. 2011, 31, 11088–11095. [Google Scholar] [CrossRef]
- Lee, H.A.; Hong, S.H.; Kim, J.W.; Jang, I.S. Possible involvement of DNA methylation in NKCC1 gene expression during postnatal development and in response to ischemia. J. Neurochem. 2010, 114, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Kanaka, C.; Ohno, K.; Okabe, A.; Kuriyama, K.; Itoh, T.; Fukuda, A.; Sato, K. The differential expression patterns of messenger RNAs encoding K-Cl cotransporters (KCC1,2) and Na-K-2Cl cotransporter (NKCC1) in the rat nervous system. Neuroscience 2001, 104, 933–946. [Google Scholar] [CrossRef]
- Morita, Y.; Callicott, J.H.; Testa, L.R.; Mighdoll, M.I.; Dickinson, D.; Chen, Q.; Tao, R.; Lipska, B.K.; Kolachana, B.; Law, A.J.; et al. Characteristics of the cation cotransporter NKCC1 in human brain: Alternate transcripts, expression in development, and potential relationships to brain function and schizophrenia. J. Neurosci. 2014, 34, 4929–4940. [Google Scholar] [CrossRef] [PubMed]
- Crouch, J.J.; Sakaguchi, N.; Lytle, C.; Schulte, B.A. Immunohistochemical localization of the Na-K-Cl co-transporter (NKCC1) in the gerbil inner ear. J. Histochem. Cytochem. 1997, 45, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, A.C.; Rivera, C. NKCC1-Deficiency Results in Abnormal Proliferation of Neural Progenitor Cells of the Lateral Ganglionic Eminence. Front. Cell Neurosci. 2016, 10. [Google Scholar] [CrossRef]
- LoTurco, J.J.; Owens, D.F.; Heath, M.J.S.; Davis, M.B.E.; Kriegstein, A.R. GABA and glutamate depolarize cortical progenitor cells and inhibit DNA synthesis. Neuron 1995, 15, 1287–1298. [Google Scholar] [CrossRef]
- Haydar, T.F.; Wang, F.; Schwartz, M.L.; Rakic, P. Differential modulation of proliferation in the neocortical ventricular and subventricular zones. J. Neurosci. 2000, 20, 5764–5774. [Google Scholar] [CrossRef]
- Blanquie, O.; Liebmann, L.; Hübner, C.A.; Luhmann, H.J.; Sinning, A. NKCC1-Mediated GABAergic Signaling Promotes Postnatal Cell Death in Neocortical Cajal-Retzius Cells. Cereb Cortex 2017, 27, 1644–1659. [Google Scholar] [CrossRef]
- Achilles, K.; Okabe, A.; Ikeda, M.; Shimizu-Okabe, C.; Yamada, J.; Fukuda, A.; Luhmann, H.J.; Kilb, W. Kinetic properties of Cl uptake mediated by Na+-dependent K+-2Cl cotransport in immature rat neocortical neurons. J. Neurosci. 2007, 27, 8616–8627. [Google Scholar] [CrossRef]
- Rivera, C.; Voipio, J.; Kaila, K. Two developmental switches in GABAergic signalling: The K+-Cl- cotransporter KCC2 and carbonic anhydrase CAVII. J. Physiol. 2005, 564, 953. [Google Scholar] [CrossRef]
- Farrant, M.; Kaila, K. The cellular, molecular and ionic basis of GABA(A) receptor signalling. Prog. Brain Res. 2007, 160, 59–87. [Google Scholar] [CrossRef] [PubMed]
- Hentschke, M.; Wiemann, M.; Hentschke, S.; Kurth, I.; Hermans-Borgmeyer, I.; Seidenbecher, T.; Jentsch, T.J.; Gal, A.; Hübner, C.A. Mice with a targeted disruption of the Cl-/HCO3- exchanger AE3 display a reduced seizure threshold. Mol. Cell Biol. 2006, 26, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Rahmati, N.; Vinueza Veloz, M.F.; Xu, J.; Barone, S.; Rodolfo Ben Hamida, N.; Schonewille, M.; Hoebeek, F.E.; Soleimani, M.; De Zeeuw, C.I. SLC26A11 (KBAT) in Purkinje Cells Is Critical for Inhibitory Transmission and Contributes to Locomotor Coordination. eNeuro 2016, 3. [Google Scholar] [CrossRef] [PubMed]
- Rungta, R.L.; Choi, H.B.; Tyson, J.R.; Malik, A.; Dissing-Olesen, L.; Lin, P.J.C.; Cain, S.M.; Cullis, P.R.; Snutch, T.P.; MacVicar, B.A. The cellular mechanisms of neuronal swelling underlying cytotoxic edema. Cell 2015, 161, 610–621. [Google Scholar] [CrossRef]
- Chesler, M. Regulation and modulation of pH in the brain. Physiol. Rev. 2003, 83, 1183–1221. [Google Scholar] [CrossRef]
- Chesler, M.; Kaila, K. Modulation of pH by neuronal activity. Trends Neurosci. 1992, 15, 396–402. [Google Scholar] [CrossRef]
- Kaila, K.; Voipio, J.; Paalasmaa, P.; Pasternack, M.; Deisz, R.A. The role of bicarbonate in GABAA receptor-mediated IPSPs of rat neocortical neurones. J. Physiol. 1993, 464, 273–289. [Google Scholar] [CrossRef]
- Kaila, K.; Voipio, J. Postsynaptic fall in intracellular pH induced by GABA-activated bicarbonate conductance. Nature 1987, 330, 163–165. [Google Scholar] [CrossRef]
- Ruusuvuori, E.; Kaila, K. Carbonic anhydrases and brain pH in the control of neuronal excitability. Subcell Biochem. 2014, 75, 271–290. [Google Scholar] [CrossRef]
- Spoljaric, I.; Spoljaric, A.; Mavrovic, M.; Seja, P.; Puskarjov, M.; Kaila, K. KCC2-Mediated Cl- Extrusion Modulates Spontaneous Hippocampal Network Events in Perinatal Rats and Mice. Cell Rep. 2019, 26, 1073–1081. [Google Scholar] [CrossRef]
- Sulis Sato, S.; Artoni, P.; Landi, S.; Cozzolino, O.; Parra, R.; Pracucci, E.; Trovato, F.; Szczurkowska, J.; Luin, S.; Arosio, D.; et al. Simultaneous two-photon imaging of intracellular chloride concentration and pH in mouse pyramidal neurons in vivo. Proc. Natl. Acad. Sci. USA 2017, 114, e8770–e8779. [Google Scholar] [CrossRef] [PubMed]
- Sipilä, S.T.; Huttu, K.; Yamada, J.; Afzalov, R.; Voipio, J.; Blaesse, P.; Kaila, K. Compensatory enhancement of intrinsic spiking upon NKCC1 disruption in neonatal hippocampus. J. Neurosci. 2009, 29, 6982–6988. [Google Scholar] [CrossRef] [PubMed]
- Cleary, R.T.; Sun, H.; Huynh, T.; Manning, S.M.; Li, Y.; Rotenberg, A.; Talos, D.M.; Kahle, K.T.; Jackson, M.; Rakhade, S.N.; et al. Bumetanide enhances phenobarbital efficacy in a rat model of hypoxic neonatal seizures. PLoS ONE 2013, 8, e57148. [Google Scholar] [CrossRef]
- Walker, P.C.; Berry, N.S.; Edwards, D.J. Protein binding characteristics of bumetanide. Dev. Pharmacol. Ther. 1989, 12, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Turmen, T.; Thom, P.; Louridas, A.T.; LeMorvan, P.; Aranda, J.V. Protein binding and bilirubin displacing properties of bumetanide and furosemide. J. Clin. Pharmacol. 1982, 22, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Shim, H.J.; Lee, M.G.; Lee, M.H. Factors influencing the protein binding of bumetanide using an equilibrium dialysis technique. J. Clin. Pharm. Ther. 1991, 16, 467–476. [Google Scholar] [CrossRef]
- Puskarjov, M.; Kahle, K.T.; Ruusuvuori, E.; Kaila, K. Pharmacotherapeutic targeting of cation-chloride cotransporters in neonatal seizures. Epilepsia 2014, 55, 806–818. [Google Scholar] [CrossRef]
- Löscher, W.; Puskarjov, M.; Kaila, K. Cation-chloride cotransporters NKCC1 and KCC2 as potential targets for novel antiepileptic and antiepileptogenic treatments. Neuropharmacology 2013, 69, 62–74. [Google Scholar] [CrossRef]
- Murata, Y.; Colonnese, M.T. GABAergic interneurons excite neonatal hippocampus in vivo. Sci. Adv. 2020, 6, eaba1430. [Google Scholar] [CrossRef]
- Sun, L.; Yu, Z.; Wang, W.; Liu, X. Both NKCC1 and anion exchangers contribute to Cl⁻ accumulation in postnatal forebrain neuronal progenitors. Eur. J. Neurosci. 2012, 35, 661–672. [Google Scholar] [CrossRef]
- Mejia-Gervacio, S.; Murray, K.; Lledo, P.M. NKCC1 controls GABAergic signaling and neuroblast migration in the postnatal forebrain. Neural. Dev. 2011, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.; Goh, E.L.; Sailor, K.A.; Kitabatake, Y.; Ming, G.L.; Song, H. GABA regulates synaptic integration of newly generated neurons in the adult brain. Nature 2006, 439, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Grove, J.C.R.; Hirano, A.A.; de Los Santos, J.; McHugh, C.F.; Purohit, S.; Field, G.D.; Brecha, N.C.; Barnes, S. Novel hybrid action of GABA mediates inhibitory feedback in the mammalian retina. PLoS Biol. 2019, 17, e3000200. [Google Scholar] [CrossRef] [PubMed]
- Jang, I.S.; Nakamura, M.; Ito, Y.; Akaike, N. Presynaptic GABAA receptors facilitate spontaneous glutamate release from presynaptic terminals on mechanically dissociated rat CA3 pyramidal neurons. Neuroscience 2006, 138, 25–35. [Google Scholar] [CrossRef]
- Jang, I.S.; Jeong, H.J.; Akaike, N. Contribution of the Na-K-Cl cotransporter on GABAA receptor-mediated presynaptic depolarization in excitatory nerve terminals. J. Neurosci. 2001, 21, 5962–5972. [Google Scholar] [CrossRef]
- Stell, B.M.; Rostaing, P.; Triller, A.; Marty, A. Activation of presynaptic GABAA receptors induces glutamate release from parallel fiber synapses. J. Neurosci. 2007, 27, 9022–9031. [Google Scholar] [CrossRef]
- Khirug, S.; Yamada, J.; Afzalov, R.; Voipio, J.; Khiroug, L.; Kaila, K. GABAergic depolarization of the axon initial segment in cortical principal neurons is caused by the Na-K-2Cl cotransporter NKCC1. J. Neurosci. 2008, 28, 4635–4639. [Google Scholar] [CrossRef]
- Szabadics, J.; Varga, C.; Molnar, G.; Olah, S.; Barzo, P.; Tamas, G. Excitatory effect of GABAergic axo-axonic cells in cortical microcircuits. Science 2006, 311, 233–235. [Google Scholar] [CrossRef]
- Woodruff, A.; Xu, Q.; Anderson, S.A.; Yuste, R. Depolarizing effect of neocortical chandelier neurons. Front. Neural Circuits 2009, 3, 15. [Google Scholar] [CrossRef]
- Glickfeld, L.L.; Roberts, J.D.; Somogyi, P.; Scanziani, M. Interneurons hyperpolarize pyramidal cells along their entire somatodendritic axis. Nat. Neurosci. 2009, 12, 21–23. [Google Scholar] [CrossRef][Green Version]
- Wang, X.; Hooks, B.M.; Sun, Q.Q. Thorough GABAergic innervation of the entire axon initial segment revealed by an optogenetic ‘laserspritzer’. J. Physiol. 2014, 592, 4257–4276. [Google Scholar] [CrossRef] [PubMed]
- Rinetti-Vargas, G.; Phamluong, K.; Ron, D.; Bender, K.J. Periadolescent Maturation of GABAergic Hyperpolarization at the Axon Initial Segment. Cell Rep. 2017, 20, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Pan-Vazquez, A.; Wefelmeyer, W.; Gonzalez Sabater, V.; Neves, G.; Burrone, J. Activity-Dependent Plasticity of Axo-axonic Synapses at the Axon Initial Segment. Neuron 2020, 106, 265–276.e266. [Google Scholar] [CrossRef] [PubMed]
- Gavrikov, K.E.; Dmitriev, A.V.; Keyser, K.T.; Mangel, S.C. Cation-chloride cotransporters mediate neural computation in the retina. Proc. Natl. Acad. Sci. USA 2003, 100, 16047–16052. [Google Scholar] [CrossRef] [PubMed]
- Duebel, J.; Haverkamp, S.; Schleich, W.; Feng, G.; Augustine, G.J.; Kuner, T.; Euler, T. Two-photon imaging reveals somatodendritic chloride gradient in retinal ON-type bipolar cells expressing the biosensor clomeleon. Neuron 2006, 49, 81–94. [Google Scholar] [CrossRef]
- Romo-Parra, H.; Trevino, M.; Heinemann, U.; Gutierrez, R. GABA actions in hippocampal area CA3 during postnatal development: Differential shift from depolarizing to hyperpolarizing in somatic and dendritic compartments. J. Neurophysiol. 2008, 99, 1523–1534. [Google Scholar] [CrossRef]
- Boron, W.F. Regulation of intracellular pH. Adv. Physiol. Educ. 2004, 28, 160–179. [Google Scholar] [CrossRef]
- Pfeffer, C.K.; Stein, V.; Keating, D.J.; Maier, H.; Rinke, I.; Rudhard, Y.; Hentschke, M.; Rune, G.M.; Jentsch, T.J.; Hübner, C.A. NKCC1-dependent GABAergic excitation drives synaptic network maturation during early hippocampal development. J. Neurosci. 2009, 29, 3419–3430. [Google Scholar] [CrossRef]
- Brickley, S.G.; Revilla, V.; Cull-Candy, S.G.; Wisden, W.; Farrant, M. Adaptive regulation of neuronal excitability by a voltage-independent potassium conductance. Nature 2001, 409, 88–92. [Google Scholar] [CrossRef]
- Ortinski, P.I.; Turner, J.R.; Barberis, A.; Motamedi, G.; Yasuda, R.P.; Wolfe, B.B.; Kellar, K.J.; Vicini, S. Deletion of the GABAA receptor alpha1 subunit increases tonic GABAA receptor current: A role for GABA uptake transporters. J. Neurosci. 2006, 26, 9323–9331. [Google Scholar] [CrossRef]
- Ponomarev, I.; Maiya, R.; Harnett, M.T.; Schafer, G.L.; Ryabinin, A.E.; Blednov, Y.A.; Morikawa, H.; Boehm, S.L., 2nd; Homanics, G.E.; Berman, A.E.; et al. Transcriptional signatures of cellular plasticity in mice lacking the alpha1 subunit of GABAA receptors. J. Neurosci. 2006, 26, 5673–5683. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Shu, S.; Schwartz, L.C.; Sun, C.; Kapur, J.; Bayliss, D.A. Homeostatic regulation of synaptic excitability: Tonic GABAA receptor currents replace Ih in cortical pyramidal neurons of HCN1 knock-out mice. J. Neurosci. 2010, 30, 2611–2622. [Google Scholar] [CrossRef]
- Zonouzi, M.; Scafidi, J.; Li, P.; McEllin, B.; Edwards, J.; Dupree, J.L.; Harvey, L.; Sun, D.; Hubner, C.A.; Cull-Candy, S.G.; et al. GABAergic regulation of cerebellar NG2 cell development is altered in perinatal white matter injury. Nat. Neurosci. 2015, 18, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Watabe, T.; Xu, M.; Watanabe, M.; Nabekura, J.; Higuchi, T.; Hori, K.; Sato, M.P.; Nin, F.; Hibino, H.; Ogawa, K.; et al. Time-controllable Nkcc1 knockdown replicates reversible hearing loss in postnatal mice. Sci. Rep. 2017, 7, 13605. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Leefmans, F.J.; Gamiño, S.M.; Giraldez, F.; Noguerón, I. Intracellular chloride regulation in amphibian dorsal root ganglion neurones studied with ion-selective microelectrodes. J. Physiol. 1988, 406, 225–246. [Google Scholar] [CrossRef]
- Toyoda, H.; Yamada, J.; Ueno, S.; Okabe, A.; Kato, H.; Sato, K.; Hashimoto, K.; Fukuda, A. Differential functional expression of cation-Cl- cotransporter mRNAs (KCC1, KCC2, and NKCC1) in rat trigeminal nervous system. Brain Res. Mol. Brain Res. 2005, 133, 12–18. [Google Scholar] [CrossRef]
- Price, T.J.; Cervero, F.; De Koninck, Y. Role of cation-chloride-cotransporters (CCC) in pain and hyperalgesia. Curr. Top. Med. Chem. 2005, 5, 547–555. [Google Scholar] [CrossRef]
- Li, C.; Lei, Y.; Tian, Y.; Xu, S.; Shen, X.; Wu, H.; Bao, S.; Wang, F. The etiological contribution of GABAergic plasticity to the pathogenesis of neuropathic pain. Mol. Pain 2019, 15. [Google Scholar] [CrossRef]
- Hughes, A.J.; Spelke, D.P.; Xu, Z.; Kang, C.C.; Schaffer, D.V.; Herr, A.E. Single-cell western blotting. Nat. Methods 2014, 11, 749–755. [Google Scholar] [CrossRef]
- Marty, S.; Wehrle, R.; Alvarez-Leefmans, F.J.; Gasnier, B.; Sotelo, C. Postnatal maturation of Na+, K+, 2Cl- cotransporter expression and inhibitory synaptogenesis in the rat hippocampus: An immunocytochemical analysis. Eur. J. Neurosci. 2002, 15, 233–245. [Google Scholar] [CrossRef]
- Yan, Y.; Dempsey, R.J.; Sun, D. Expression of Na+-K+-Cl- cotransporter in rat brain during development and its localization in mature astrocytes. Brain Res. 2001, 911, 43–55. [Google Scholar] [CrossRef]
- Plotkin, M.D.; Snyder, E.Y.; Hebert, S.C.; Delpire, E. Expression of the Na-K-2Cl cotransporter is developmentally regulated in postnatal rat brains: A possible mechanism underlying GABA’s excitatory role in immature brain. J. Neurobiol. 1997, 33, 781–795. [Google Scholar] [CrossRef]
- Liu, Q.; Wong-Riley, M.T. Postnatal development of Na(+)-K(+)-2Cl(-) co-transporter 1 and K(+)-Cl(-) co-transporter 2 immunoreactivity in multiple brain stem respiratory nuclei of the rat. Neuroscience 2012, 210, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Delpire, E.; Vardi, N. NKCC1 does not accumulate chloride in developing retinal neurons. J. Neurophysiol. 2007, 98, 266–277. [Google Scholar] [CrossRef]
- Li, B.Q.; McKernan, K.; Shen, W. Spatial and temporal distribution patterns of Na-K-2C1 cotransporter in adult and developing mouse retinas. Vis. Neurosci. 2008, 25, 109–123. [Google Scholar] [CrossRef]
- Fritschy, J.M. Is my antibody-staining specific? How to deal with pitfalls of immunohistochemistry. Eur. J. Neurosci. 2008, 28, 2365–2370. [Google Scholar] [CrossRef]
- Saper, C.B.; Sawchenko, P.E. Magic peptides, magic antibodies: Guidelines for appropriate controls for immunohistochemistry. J. Comp. Neurol. 2003, 465, 161–163. [Google Scholar] [CrossRef]
- Hartmann, A.M.; Nothwang, H.G. Molecular and evolutionary insights into the structural organization of cation chloride cotransporters. Front. Cell Neurosci. 2014, 8, 470. [Google Scholar] [CrossRef]
- Delpire, E.; Austin, T.M. Kinase regulation of Na+-K+-2Cl- cotransport in primary afferent neurons. J. Physiol. 2010, 588, 3365–3373. [Google Scholar] [CrossRef]
- Schwanhäusser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global quantification of mammalian gene expression control. Nature 2011, 473, 337–342. [Google Scholar] [CrossRef]
- Azimifar, S.B.; Nagaraj, N.; Cox, J.; Mann, M. Cell-type-resolved quantitative proteomics of murine liver. Cell Metab. 2014, 20, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Holt, C.E.; Martin, K.C.; Schuman, E.M. Local translation in neurons: Visualization and function. Nat. Struct. Mol. Biol. 2019, 26, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Doyle, M.; Kiebler, M.A. Mechanisms of dendritic mRNA transport and its role in synaptic tagging. EMBO J. 2011, 30, 3540–3552. [Google Scholar] [CrossRef]
- Cioni, J.M.; Koppers, M.; Holt, C.E. Molecular control of local translation in axon development and maintenance. Curr. Opin. Neurobiol. 2018, 51, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Thelen, M.P.; Kye, M.J. The Role of RNA Binding Proteins for Local mRNA Translation: Implications in Neurological Disorders. Front. Mol. Biosci. 2019, 6, 161. [Google Scholar] [CrossRef]
- Kosik, K.S. Life at Low Copy Number: How Dendrites Manage with So Few mRNAs. Neuron 2016, 92, 1168–1180. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef]
- Zeisel, A.; Hochgerner, H.; Lonnerberg, P.; Johnsson, A.; Memic, F.; van der Zwan, J.; Haring, M.; Braun, E.; Borm, L.E.; La Manno, G.; et al. Molecular Architecture of the Mouse Nervous System. Cell 2018, 174, 999–1014.e1022. [Google Scholar] [CrossRef]
- Saunders, A.; Macosko, E.Z.; Wysoker, A.; Goldman, M.; Krienen, F.M.; de Rivera, H.; Bien, E.; Baum, M.; Bortolin, L.; Wang, S.; et al. Molecular Diversity and Specializations among the Cells of the Adult Mouse Brain. Cell 2018, 174, 1015–1030.e1016. [Google Scholar] [CrossRef]
- Loo, L.; Simon, J.M.; Xing, L.; McCoy, E.S.; Niehaus, J.K.; Guo, J.; Anton, E.S.; Zylka, M.J. Single-cell transcriptomic analysis of mouse neocortical development. Nat. Commun. 2019, 10, 134. [Google Scholar] [CrossRef]
- Brain RNA-Seq. Available online: https://www.brainrnaseq.org/ (accessed on 5 November 2020).
- Mouse Brain Atlas. Available online: http://mousebrain.org/ (accessed on 5 November 2020).
- Single-Cell Transcriptomic Analysis of Mouse Neocortical Development. Available online: http://zylkalab.org/datamousecortex (accessed on 5 November 2020).
- Dropviz. Available online: http://dropviz.org/ (accessed on 5 November 2020).
- Allen Brain Map Transcriptomics Explorer. Available online: https://celltypes.brain-map.org (accessed on 5 November 2020).
- Vibat, C.R.T.; Holland, M.J.; Kang, J.J.; Putney, L.K.; O’Donnell, M.E. Quantitation of Na+-K+-2Cl- cotransport splice variants in human tissues using kinetic polymerase chain reaction. Anal. Biochem. 2001, 298, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Fukuda, A. Development and regulation of chloride homeostasis in the central nervous system. Front. Cell Neurosci. 2015, 9, 371. [Google Scholar] [CrossRef] [PubMed]
- Medina, I.; Friedel, P.; Rivera, C.; Kahle, K.T.; Kourdougli, N.; Uvarov, P.; Pellegrino, C. Current view on the functional regulation of the neuronal K+-Cl- cotransporter KCC2. Front. Cell Neurosci. 2014, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Kahle, K.T.; Deeb, T.Z.; Puskarjov, M.; Silayeva, L.; Liang, B.; Kaila, K.; Moss, S.J. Modulation of neuronal activity by phosphorylation of the K-Cl cotransporter KCC2. Trends Neurosci. 2013, 36, 726–737. [Google Scholar] [CrossRef]
- Sun, D.; Murali, S.G. Na+-K+-2Cl- cotransporter in immature cortical neurons: A role in intracellular Cl- regulation. J. Neurophysiol. 1999, 81, 1939–1948. [Google Scholar] [CrossRef]
- Dzhala, V.I.; Talos, D.M.; Sdrulla, D.A.; Brumback, A.C.; Mathews, G.C.; Benke, T.A.; Delpire, E.; Jensen, F.E.; Staley, K.J. NKCC1 transporter facilitates seizures in the developing brain. Nat. Med. 2005, 11, 1205–1213. [Google Scholar] [CrossRef]
- Ikeda, M.; Toyoda, H.; Yamada, J.; Okabe, A.; Sato, K.; Hotta, Y.; Fukuda, A. Differential development of cation-chloride cotransporters and Cl- homeostasis contributes to differential GABAergic actions between developing rat visual cortex and dorsal lateral geniculate nucleus. Brain Res. 2003, 984, 149–159. [Google Scholar] [CrossRef]
- Shimizu-Okabe, C.; Yokokura, M.; Okabe, A.; Ikeda, M.; Sato, K.; Kilb, W.; Luhmann, H.J.; Fukuda, A. Layer-specific expression of Cl- transporters and differential [Cl-]i in newborn rat cortex. Neuroreport 2002, 13, 2433–2437. [Google Scholar] [CrossRef]
- Aronica, E.; Boer, K.; Redeker, S.; Spliet, W.G.; van Rijen, P.C.; Troost, D.; Gorter, J.A. Differential expression patterns of chloride transporters, Na+-K+-2Cl--cotransporter and K+-Cl--cotransporter, in epilepsy-associated malformations of cortical development. Neuroscience 2007, 145, 185–196. [Google Scholar] [CrossRef]
- Mikawa, S.; Wang, C.; Shu, F.; Wang, T.; Fukuda, A.; Sato, K. Developmental changes in KCC1, KCC2 and NKCC1 mRNAs in the rat cerebellum. Brain Res. Dev. Brain Res. 2002, 136, 93–100. [Google Scholar] [CrossRef]
- Wang, C.; Shimizu-Okabe, C.; Watanabe, K.; Okabe, A.; Matsuzaki, H.; Ogawa, T.; Mori, N.; Fukuda, A.; Sato, K. Developmental changes in KCC1, KCC2, and NKCC1 mRNA expressions in the rat brain. Brain Res. Dev. Brain Res. 2002, 139, 59–66. [Google Scholar] [CrossRef]
- Delpy, A.; Allain, A.E.; Meyrand, P.; Branchereau, P. NKCC1 cotransporter inactivation underlies embryonic development of chloride-mediated inhibition in mouse spinal motoneuron. J. Physiol. 2008, 586, 1059–1075. [Google Scholar] [CrossRef] [PubMed]
- Stil, A.; Liabeuf, S.; Jean-Xavier, C.; Brocard, C.; Viemari, J.C.; Vinay, L. Developmental up-regulation of the potassium-chloride cotransporter type 2 in the rat lumbar spinal cord. Neuroscience 2009, 164, 809–821. [Google Scholar] [CrossRef] [PubMed]
- Eng, C.L.; Lawson, M.; Zhu, Q.; Dries, R.; Koulena, N.; Takei, Y.; Yun, J.; Cronin, C.; Karp, C.; Yuan, G.C.; et al. Transcriptome-scale super-resolved imaging in tissues by RNA seqFISH. Nature 2019, 568, 235–239. [Google Scholar] [CrossRef]
- Chen, K.H.; Boettiger, A.N.; Moffitt, J.R.; Wang, S.; Zhuang, X. RNA imaging. Spatially resolved, highly multiplexed RNA profiling in single cells. Science 2015, 348, aaa6090. [Google Scholar] [CrossRef]
- Balapattabi, K.; Farmer, G.E.; Knapp, B.A.; Little, J.T.; Bachelor, M.; Yuan, J.P.; Cunningham, J.T. Effects of salt-loading on supraoptic vasopressin neurones assessed by ClopHensorN chloride imaging. J. Neuroendocrinol. 2019, 31, e12752. [Google Scholar] [CrossRef]
- Mikuni, T.; Nishiyama, J.; Sun, Y.; Kamasawa, N.; Yasuda, R. High-Throughput, High-Resolution Mapping of Protein Localization in Mammalian Brain by In Vivo Genome Editing. Cell 2016, 165, 1803–1817. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Study | Journal | Species | Structure | Age Range | Method | Target Region | Knock-Out Control 1 | Cell Types Expressing NKCC1 | |
|---|---|---|---|---|---|---|---|---|---|
| Earliest Age Point | Latest Age Point | ||||||||
| Studies Reporting an Upregulation of NKCC1 During Development: | |||||||||
| Sedmak, 2015 | Cereb Cortex | Human | Ctx, Hc, Th, Str, Cb | GW11–90Y | Microarray | Entire gene | - | - | - |
| Morita, 2014 | J Neurosci | Human | Ctx | GW14–78Y | PCR | (21) | - | - | - |
| Hyde, 2011 | J. Neurosci. | Human | Ctx, Hc | GW14–80Y | PCR | (17) | - | - | - |
| Li, 2008 | Vis. Neurosci. | Mouse | Ret | P0–P20 | IHC | (6) | Yes | Inner plexiform layer, neuroplastic cells | Horizontal cells, rod-bipolar cells, photoreceptors |
| Zhang, 2007 | J. Neurophysiol. | Mouse | Ret | P0–P90 | WB, IHC | (7) | Yes | Müller cells | Horizontal cells |
| Balakrishnan, 2003 | J. Neurosci. | Rat, Mouse | Bs | P3–P12 | ISH, PCR | (12) (25) | - | None | Lateral superior olive neurons |
| Mikawa, 2002 | Dev. Brain Res. | Rat | Cb | P1–P49 | ISH | (19) | - | None | Granule cells, glia |
| Wang, 2002 | Dev. Brain Res. | Rat | Hc, Th | E18–P40 | ISH | (19) | - | Neuroepithelium (Hc) | Neurons (Hc) |
| Yan, 2001 | Brain Res. | Rat | Ctx, Hc, Cb, Th, Str | P0–Adult | WB, IHC | (6) | - | Neurons | Neurons (Ctx, Hc, Cb, Th), astrocytes (Ctx, Hc, Cb) |
| Sun and Murali, 1999 | J. Neurophysiol. | Rat | Ctx | P0–P9 | WB | (6) | - | - | - |
| Studies Reporting a Downregulation of NKCC1 during Development: | |||||||||
| Liu and Wong-Riley, 2012 | Neuroscience | Rat | Bs | P0–P21 | IHC | (6) | Yes | Neurons | Neurons, glia |
| Lee, 2010 | J. Neurochem. | Rat | Ctx | P5–P90 | PCR | (18) | - | - | - |
| Stil, 2009 | Neuroscience | Rat | Sc | E17–P20 | IHC | (6) | - | - | - |
| Delpy, 2008 | J. Physiol. | Mouse | Sc | E11.5–P0 | IHC | (6) | - | Motoneurons | Motoneurons |
| Aronica, 2007 | Neuroscience | Human | Ctx | 2M–30Y | IHC | (8) | - | Neurons | Neurons, astrocytes |
| Dzhala, 2005 | Nat. Med. | Rat, Human | Ctx | P3–Adult, GW19–5Y2 | WB, IHC | (8) | - | Neurons, non-neuronal cells | Neurons, non-neuronal cells |
| Yamada, 2004 | J. Physiol. | Rat | Ctx | P1–P20 | PCR | (16) | - | Cortical plate neurons | - |
| Ikeda, 2003 | Brain Res. | Rat | Ctx | P0–P28 | ISH | (19) | - | Neurons | Neurons |
| Shimizu-Okabe, 2002 | Neuroreport | Rat | Ctx | P1–P28 | ISH | (19) (20) | - | - | - |
| Wang, 2002 | Dev. Brain Res. | Rat | Ctx | E18–P40 | ISH | (19) | - | Neuroepithelium, cortical plate | Neurons, glia |
| Hübner, 2001 | Mech. Dev. | Mouse | Ctx, Hc | E12.5–Adult | ISH | (10) | - | - | - |
| Plotkin, 1997 | J. Neurobiol. | Rat | Ctx, Hc, Cb | P0 Adult | WB, IHC, ISH | (7) | - | Neurons, transiently at P14 Oligodendrocytes | Neurons |
| Studies reporting stable expression of NKCC1 during development: | |||||||||
| Mao, 2012 | J. Neurophysiol. | Rat, Mouse | DRG | P1–Adult | PCR, IHC, ISH | (2) (5) (6) (7) (13) (23) (24) | Yes (IHC) | Neurons | Neurons |
| Ikeda, 2003 | Brain Res. | Rat | Th | P0–P28 | ISH | (19) | - | Neurons | Neurons |
| Marty, 2002 | Eur. J. Neurosci. | Rat | Hc | P1–P60 | IHC | (6) | - | Neurons | Pyramidal neurons, interneurons |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Virtanen, M.A.; Uvarov, P.; Hübner, C.A.; Kaila, K. NKCC1, an Elusive Molecular Target in Brain Development: Making Sense of the Existing Data. Cells 2020, 9, 2607. https://doi.org/10.3390/cells9122607
Virtanen MA, Uvarov P, Hübner CA, Kaila K. NKCC1, an Elusive Molecular Target in Brain Development: Making Sense of the Existing Data. Cells. 2020; 9(12):2607. https://doi.org/10.3390/cells9122607
Chicago/Turabian StyleVirtanen, Mari A., Pavel Uvarov, Christian A. Hübner, and Kai Kaila. 2020. "NKCC1, an Elusive Molecular Target in Brain Development: Making Sense of the Existing Data" Cells 9, no. 12: 2607. https://doi.org/10.3390/cells9122607
APA StyleVirtanen, M. A., Uvarov, P., Hübner, C. A., & Kaila, K. (2020). NKCC1, an Elusive Molecular Target in Brain Development: Making Sense of the Existing Data. Cells, 9(12), 2607. https://doi.org/10.3390/cells9122607