Natural Heart Regeneration in a Neonatal Rat Myocardial Infarction Model

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Care and Biosafety
2.2. Neonatal Rat Myocardial Infarction Model
2.3. Adult Rat Myocardial Infarction Model
2.4. Injection of 5-ethynyl-2′-deoxyuridine
2.5. Echocardiography
2.6. Heart Explant and Sample Preparation for Histology
2.7. Hematoxylin and Eosin Staining
2.8. Masson’s Trichrome and Picrosirius Red Staining
2.9. Immunohistochemistry for Troponin, Ki-67, Aurora B Kinase
2.10. EdU Processing
2.11. Image Acquisition and Cell Quantification
2.12. Statistical Analysis
3. Results
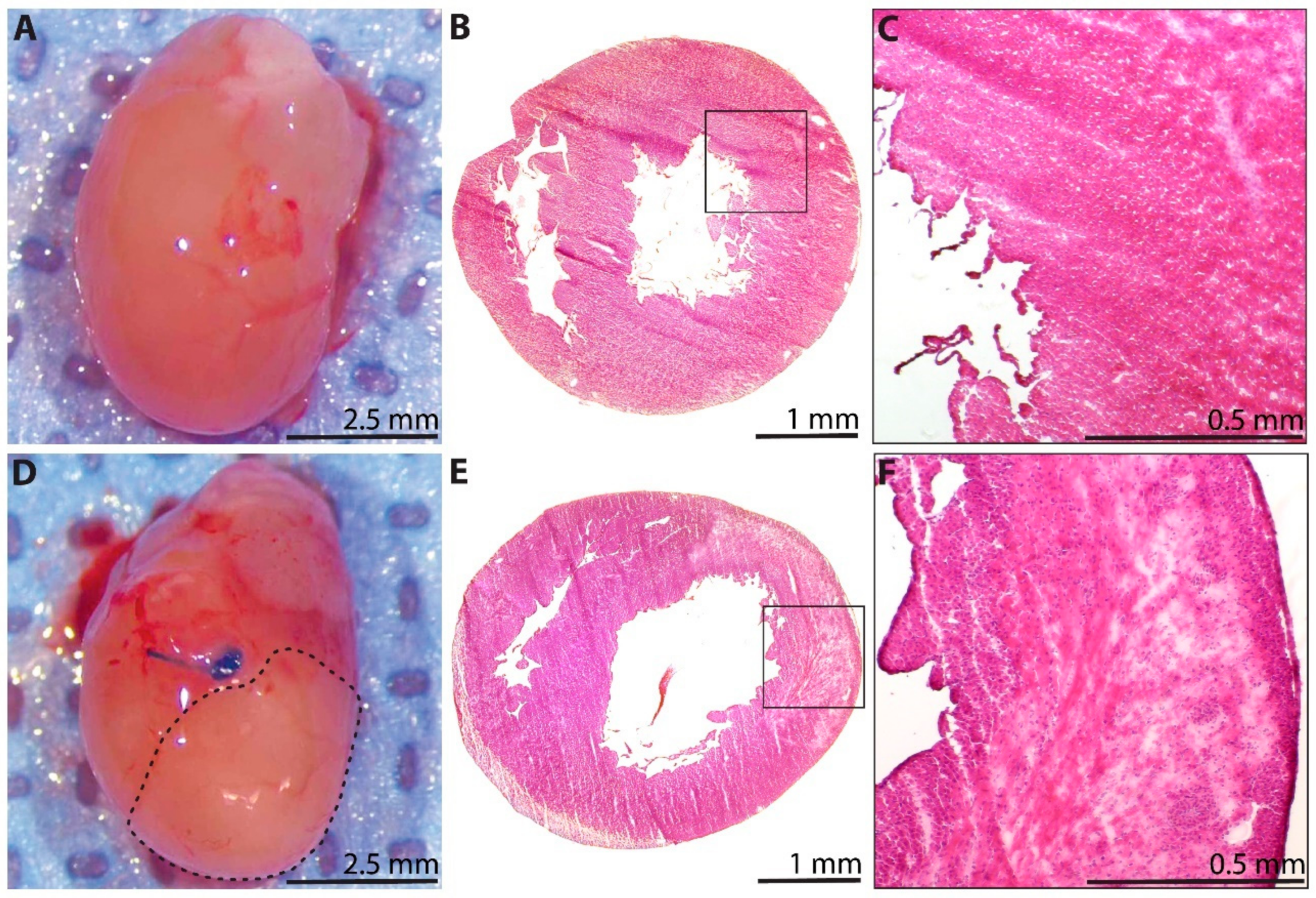
3.1. LAD Ligation Generates Acute Myocardial Infarcts in Neonatal Rats
3.2. Cardiac Troponin Is Recovered by 3 Weeks after Myocardial Infarction
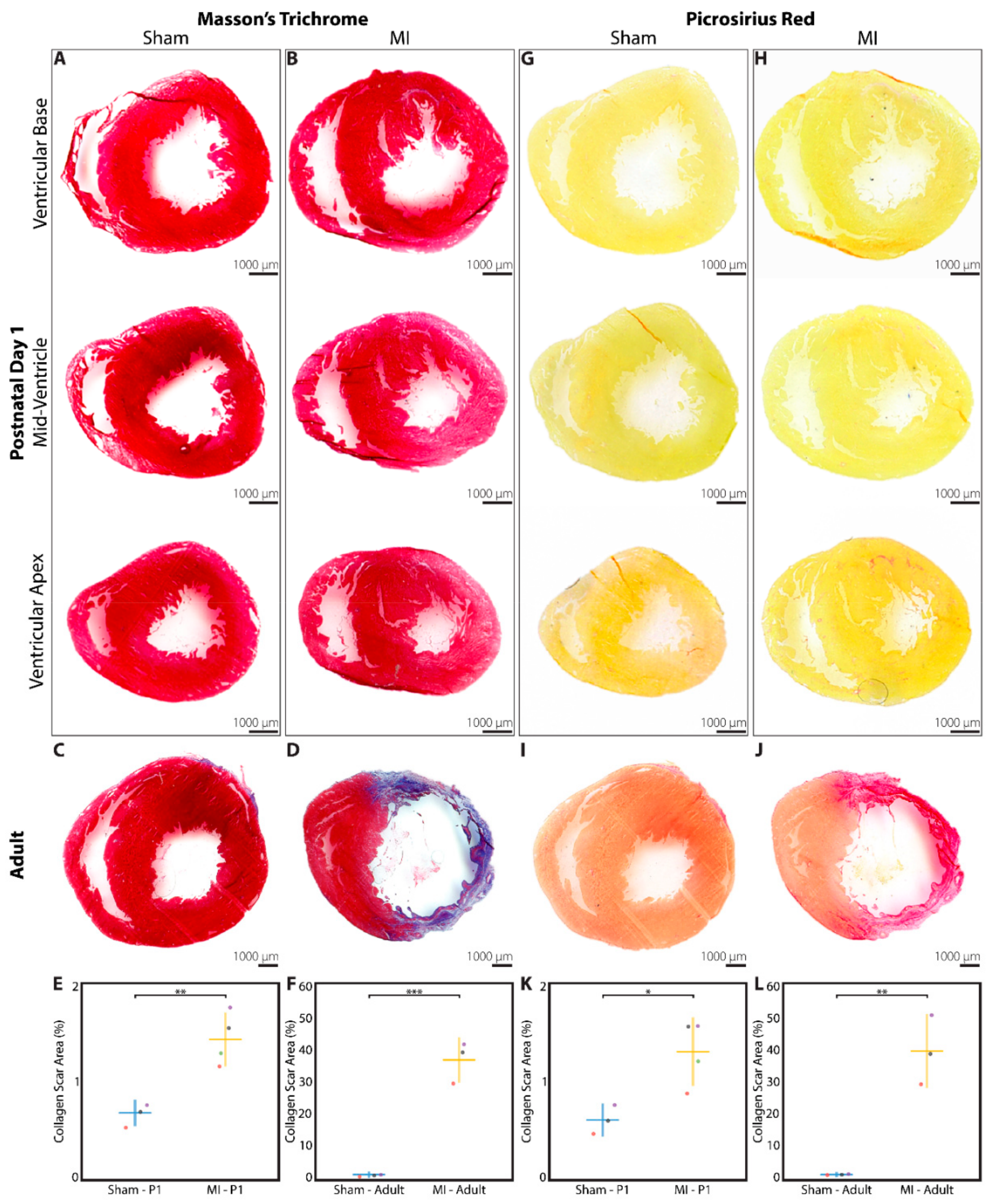
3.3. Minimal Fibrotic Scar Is Observed at 3 Weeks after Myocardial Infarction
3.4. Increased Cardiomyocyte Proliferation Is Observed after Myocardial Infarction
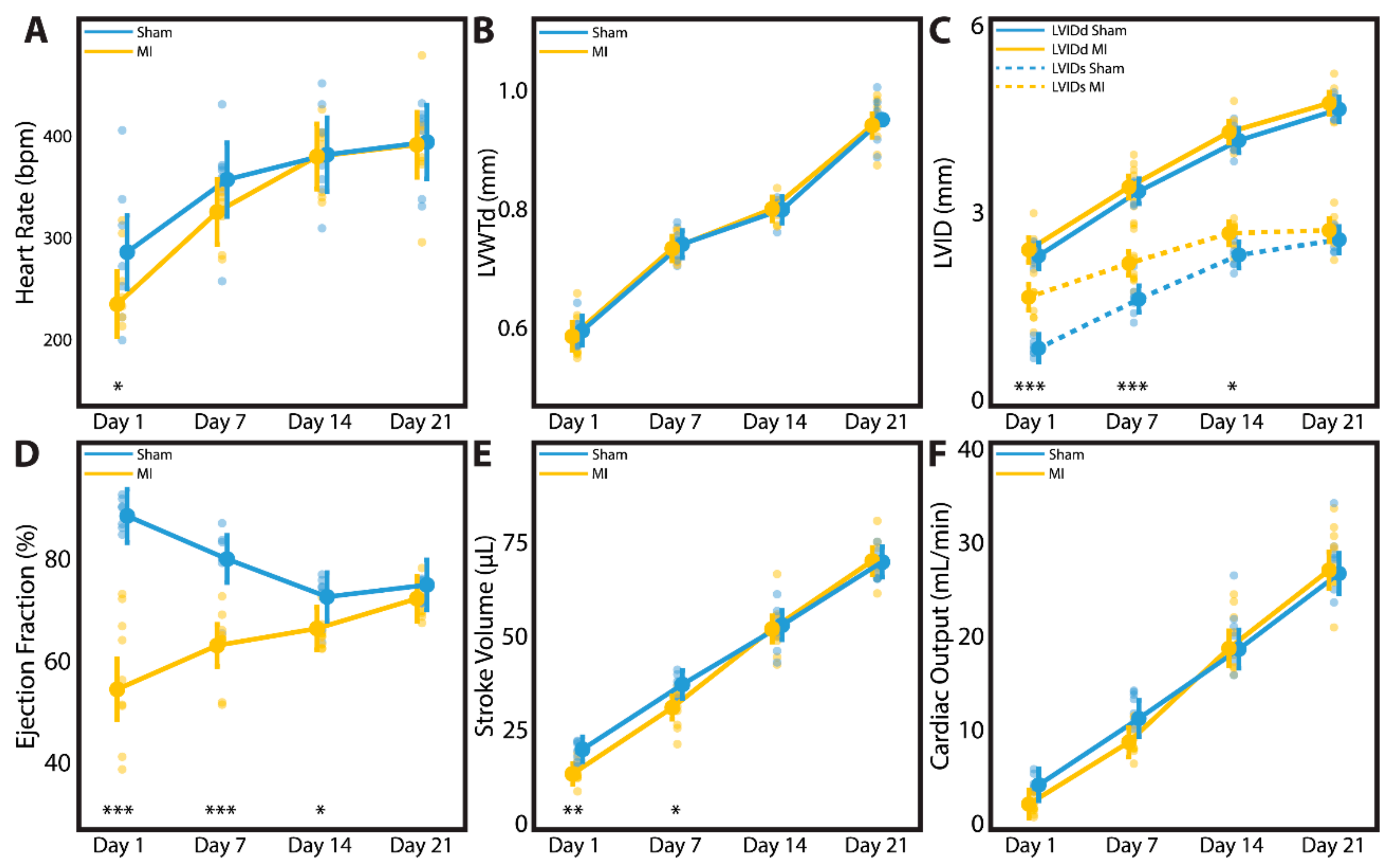
3.5. Cardiac Geometry and Function Fully Recover by 3 Weeks after Myocardial Infarction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benjamin, E.J.; Muntner, P.; Alonso, A.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Das, S.R.; et al. American Heart Association Council on Epidemiology and Prevention Statistics Committee and Stroke Statistics Subcommittee Heart Disease and Stroke Statistics-2019 Update: A Report From the American Heart Association. Circulation 2019, 139, e56–e528. [Google Scholar] [CrossRef]
- Heidenreich, P.A.; Trogdon, J.G.; Khavjou, O.A.; Butler, J.; Dracup, K.; Ezekowitz, M.D.; Finkelstein, E.A.; Hong, Y.; Johnston, S.C.; Khera, A.; et al. Forecasting the future of cardiovascular disease in the United States: A policy statement from the American Heart Association. Circulation 2011, 123, 933–944. [Google Scholar] [CrossRef]
- Li, S.; Jiao, Y.; Wang, H.; Shang, Q.; Lu, F.; Huang, L.; Liu, J.; Xu, H.; Chen, K. Sodium tanshinone IIA sulfate adjunct therapy reduces high-sensitivity C-reactive protein level in coronary artery disease patients: A randomized controlled trial. Sci. Rep. 2017, 7, 17451. [Google Scholar] [CrossRef]
- Goldstone, A.B.; Chiu, P.; Baiocchi, M.; Wang, H.; Lingala, B.; Boyd, J.H.; Woo, Y.J. Second arterial versus venous conduits for multivessel coronary artery bypass surgery in california. Circulation 2018, 137, 1698–1707. [Google Scholar] [CrossRef]
- Bolognese, L.; Neskovic, A.N.; Parodi, G.; Cerisano, G.; Buonamici, P.; Santoro, G.M.; Antoniucci, D. Left ventricular remodeling after primary coronary angioplasty: Patterns of left ventricular dilation and long-term prognostic implications. Circulation 2002, 106, 2351–2357. [Google Scholar] [CrossRef] [PubMed]
- Heallen, T.R.; Kadow, Z.A.; Kim, J.H.; Wang, J.; Martin, J.F. Stimulating cardiogenesis as a treatment for heart failure. Circ. Res. 2019, 124, 1647–1657. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, M.A.; Braunwald, E. Ventricular remodeling after myocardial infarction. Experimental observations and clinical implications. Circulation 1990, 81, 1161–1172. [Google Scholar] [CrossRef] [PubMed]
- Ascheim, D.D.; Gelijns, A.C.; Goldstein, D.; Moye, L.A.; Smedira, N.; Lee, S.; Klodell, C.T.; Szady, A.; Parides, M.K.; Jeffries, N.O.; et al. Mesenchymal precursor cells as adjunctive therapy in recipients of contemporary left ventricular assist devices. Circulation 2014, 129, 2287–2296. [Google Scholar] [CrossRef] [PubMed]
- Menasché, P. Cell therapy trials for heart regeneration-lessons learned and future directions. Nat. Rev. Cardiol. 2018, 15, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Shudo, Y.; Goldstone, A.B.; Cohen, J.E.; Patel, J.B.; Hopkins, M.S.; Steele, A.N.; Edwards, B.B.; Kawamura, M.; Miyagawa, S.; Sawa, Y.; et al. Layered smooth muscle cell-endothelial progenitor cell sheets derived from the bone marrow augment postinfarction ventricular function. J. Thorac. Cardiovasc. Surg. 2017, 154, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Shudo, Y.; Cohen, J.E.; Macarthur, J.W.; Atluri, P.; Hsiao, P.F.; Yang, E.C.; Fairman, A.S.; Trubelja, A.; Patel, J.; Miyagawa, S.; et al. Spatially oriented, temporally sequential smooth muscle cell-endothelial progenitor cell bi-level cell sheet neovascularizes ischemic myocardium. Circulation 2013, 128, S59–S68. [Google Scholar] [CrossRef] [PubMed]
- Wanjare, M.; Kawamura, M.; Hu, C.; Alcazar, C.; Wang, H.; Woo, Y.J.; Huang, N.F. Vascularization of Engineered Spatially Patterned Myocardial Tissue Derived From Human Pluripotent Stem Cells in vivo. Front. Bioeng. Biotechnol. 2019, 7, 208. [Google Scholar] [CrossRef]
- Von Bornstädt, D.; Wang, H.; Paulsen, M.J.; Goldstone, A.B.; Eskandari, A.; Thakore, A.; Stapleton, L.; Steele, A.N.; Truong, V.N.; Jaatinen, K.; et al. Rapid Self-Assembly of Bioengineered Cardiovascular Bypass Grafts From Scaffold-Stabilized, Tubular Bilevel Cell Sheets. Circulation 2018, 138, 2130–2144. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, M.; Paulsen, M.J.; Goldstone, A.B.; Shudo, Y.; Wang, H.; Steele, A.N.; Stapleton, L.M.; Edwards, B.B.; Eskandari, A.; Truong, V.N.; et al. Tissue-engineered smooth muscle cell and endothelial progenitor cell bi-level cell sheets prevent progression of cardiac dysfunction, microvascular dysfunction, and interstitial fibrosis in a rodent model of type 1 diabetes-induced cardiomyopathy. Cardiovasc. Diabetol. 2017, 16, 142. [Google Scholar] [CrossRef] [PubMed]
- Hiesinger, W.; Perez-Aguilar, J.M.; Atluri, P.; Marotta, N.A.; Frederick, J.R.; Fitzpatrick, J.R.; McCormick, R.C.; Muenzer, J.R.; Yang, E.C.; Levit, R.D.; et al. Computational protein design to reengineer stromal cell-derived factor-1α generates an effective and translatable angiogenic polypeptide analog. Circulation 2011, 124, S18–S26. [Google Scholar] [CrossRef] [PubMed]
- Macarthur, J.W.; Cohen, J.E.; McGarvey, J.R.; Shudo, Y.; Patel, J.B.; Trubelja, A.; Fairman, A.S.; Edwards, B.B.; Hung, G.; Hiesinger, W.; et al. Preclinical evaluation of the engineered stem cell chemokine stromal cell-derived factor 1α analog in a translational ovine myocardial infarction model. Circ. Res. 2014, 114, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wisneski, A.; Paulsen, M.J.; Imbrie-Moore, A.; Wang, Z.; Xuan, Y.; Hernandez, H.L.; Lucian, H.J.; Eskandari, A.; Thakore, A.D.; et al. Bioengineered analog of stromal cell-derived factor 1α preserves the biaxial mechanical properties of native myocardium after infarction. J. Mech. Behav. Biomed. Mater. 2019, 96, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.E.; Purcell, B.P.; MacArthur, J.W.; Mu, A.; Shudo, Y.; Patel, J.B.; Brusalis, C.M.; Trubelja, A.; Fairman, A.S.; Edwards, B.B.; et al. A bioengineered hydrogel system enables targeted and sustained intramyocardial delivery of neuregulin, activating the cardiomyocyte cell cycle and enhancing ventricular function in a murine model of ischemic cardiomyopathy. Circ. Heart Fail. 2014, 7, 619–626. [Google Scholar] [CrossRef][Green Version]
- Steele, A.N.; Cai, L.; Truong, V.N.; Edwards, B.B.; Goldstone, A.B.; Eskandari, A.; Mitchell, A.C.; Marquardt, L.M.; Foster, A.A.; Cochran, J.R.; et al. A novel protein-engineered hepatocyte growth factor analog released via a shear-thinning injectable hydrogel enhances post-infarction ventricular function. Biotechnol. Bioeng. 2017, 114, 2379–2389. [Google Scholar] [CrossRef]
- Oberpriller, J.O.; Oberpriller, J.C. Response of the adult newt ventricle to injury. J. Exp. Zool. 1974, 187, 249–253. [Google Scholar] [CrossRef]
- Poss, K.D.; Wilson, L.G.; Keating, M.T. Heart regeneration in zebrafish. Science 2002, 298, 2188–2190. [Google Scholar] [CrossRef] [PubMed]
- Porrello, E.R.; Mahmoud, A.I.; Simpson, E.; Hill, J.A.; Richardson, J.A.; Olson, E.N.; Sadek, H.A. Transient regenerative potential of the neonatal mouse heart. Science 2011, 331, 1078–1080. [Google Scholar] [CrossRef] [PubMed]
- Zogbi, C.; Saturi de Carvalho, A.E.T.; Nakamuta, J.S.; Caceres, V.M.; Prando, S.; Giorgi, M.C.P.; Rochitte, C.E.; Meneghetti, J.C.; Krieger, J.E. Early postnatal rat ventricle resection leads to long-term preserved cardiac function despite tissue hypoperfusion. Physiol. Rep. 2014, 2. [Google Scholar] [CrossRef]
- Haubner, B.J.; Adamowicz-Brice, M.; Khadayate, S.; Tiefenthaler, V.; Metzler, B.; Aitman, T.; Penninger, J.M. Complete cardiac regeneration in a mouse model of myocardial infarction. Aging 2012, 4, 966–977. [Google Scholar] [CrossRef] [PubMed]
- Porrello, E.R.; Mahmoud, A.I.; Simpson, E.; Johnson, B.A.; Grinsfelder, D.; Canseco, D.; Mammen, P.P.; Rothermel, B.A.; Olson, E.N.; Sadek, H.A. Regulation of neonatal and adult mammalian heart regeneration by the miR-15 family. Proc. Natl. Acad. Sci. USA 2013, 110, 187–192. [Google Scholar] [CrossRef]
- Das, S.; Goldstone, A.B.; Wang, H.; Farry, J.; D’Amato, G.; Paulsen, M.J.; Eskandari, A.; Hironaka, C.E.; Phansalkar, R.; Sharma, B.; et al. A unique collateral artery development program promotes neonatal heart regeneration. Cell 2019, 176, 1128–1142. [Google Scholar] [CrossRef]
- Zhu, W.; Zhang, E.; Zhao, M.; Chong, Z.; Fan, C.; Tang, Y.; Hunter, J.D.; Borovjagin, A.V.; Walcott, G.P.; Chen, J.Y.; et al. Regenerative potential of neonatal porcine hearts. Circulation 2018, 138, 2809–2816. [Google Scholar] [CrossRef]
- Ye, L.; D’Agostino, G.; Loo, S.J.; Wang, C.X.; Su, L.P.; Tan, S.H.; Tee, G.Z.; Pua, C.J.; Pena, E.M.; Cheng, R.B.; et al. Early regenerative capacity in the porcine heart. Circulation 2018, 138, 2798–2808. [Google Scholar] [CrossRef]
- Haubner, B.J.; Schneider, J.; Schweigmann, U.; Schuetz, T.; Dichtl, W.; Velik-Salchner, C.; Stein, J.-I.; Penninger, J.M. Functional recovery of a human neonatal heart after severe myocardial infarction. Circ. Res. 2016, 118, 216–221. [Google Scholar] [CrossRef]
- Phifer, C.B.; Terry, L.M. Use of hypothermia for general anesthesia in preweanling rodents. Physiol. Behav. 1986, 38, 887–890. [Google Scholar] [CrossRef]
- Mahmoud, A.I.; Porrello, E.R.; Kimura, W.; Olson, E.N.; Sadek, H.A. Surgical models for cardiac regeneration in neonatal mice. Nat. Protoc. 2014, 9, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, L.M.; Steele, A.N.; Wang, H.; Lopez Hernandez, H.; Yu, A.C.; Paulsen, M.J.; Smith, A.A.A.; Roth, G.A.; Thakore, A.D.; Lucian, H.J.; et al. Use of a supramolecular polymeric hydrogel as an effective post-operative pericardial adhesion barrier. Nat. Biomed. Eng. 2019, 3, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Rothman, K.J. No adjustments are needed for multiple comparisons. Epidemiology 1990, 1, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Tong, S.; Ren, X.; Xia, H. Cardiac cell proliferation assessed by EdU, a novel analysis of cardiac regeneration. Cytotechnology 2016, 68, 763–770. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gerdes, J.; Lemke, H.; Baisch, H.; Wacker, H.H.; Schwab, U.; Stein, H. Cell cycle analysis of a cell proliferation-associated human nuclear antigen defined by the monoclonal antibody Ki-67. J. Immunol. 1984, 133, 1710–1715. [Google Scholar] [PubMed]
- Murata-Hori, M.; Tatsuka, M.; Wang, Y.-L. Probing the dynamics and functions of aurora B kinase in living cells during mitosis and cytokinesis. Mol. Biol. Cell 2002, 13, 1099–1108. [Google Scholar] [CrossRef]
- Lam, N.T.; Sadek, H.A. Neonatal Heart Regeneration. Circulation 2018, 138, 412–423. [Google Scholar] [CrossRef]
- Liu, Z.-J.; Hoffmeister, K.M.; Hu, Z.; Mager, D.E.; Ait-Oudhia, S.; Debrincat, M.A.; Pleines, I.; Josefsson, E.C.; Kile, B.T.; Italiano, J.; et al. Expansion of the neonatal platelet mass is achieved via an extension of platelet lifespan. Blood 2014, 123, 3381–3389. [Google Scholar] [CrossRef]
- Ingason, A.B.; Goldstone, A.B.; Paulsen, M.J.; Thakore, A.D.; Truong, V.N.; Edwards, B.B.; Eskandari, A.; Bollig, T.; Steele, A.N.; Woo, Y.J. Angiogenesis precedes cardiomyocyte migration in regenerating mammalian hearts. J. Thorac. Cardiovasc. Surg. 2018, 155, 1118–1127. [Google Scholar] [CrossRef]
- Li, F.; Wang, X.; Capasso, J.M.; Gerdes, A.M. Rapid transition of cardiac myocytes from hyperplasia to hypertrophy during postnatal development. J. Mol. Cell Cardiol. 1996, 28, 1737–1746. [Google Scholar] [CrossRef]
- Marín-Juez, R.; El-Sammak, H.; Helker, C.S.M.; Kamezaki, A.; Mullapuli, S.T.; Bibli, S.-I.; Foglia, M.J.; Fleming, I.; Poss, K.D.; Stainier, D.Y.R. Coronary revascularization during heart regeneration is regulated by epicardial and endocardial cues and forms a scaffold for cardiomyocyte repopulation. Dev. Cell 2019, 51, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Aurora, A.B.; Porrello, E.R.; Tan, W.; Mahmoud, A.I.; Hill, J.A.; Bassel-Duby, R.; Sadek, H.A.; Olson, E.N. Macrophages are required for neonatal heart regeneration. J. Clin. Investig. 2014, 124, 1382–1392. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, K.Y.; Tam, R.C.Y.; Chan, V.W.; Lan, H.Y.; Hori, S.; Zhou, B.; Lui, K.O. Regulatory T-cells regulate neonatal heart regeneration by potentiating cardiomyocyte proliferation in a paracrine manner. Theranostics 2019, 9, 4324–4341. [Google Scholar] [CrossRef] [PubMed]
- Hirose, K.; Payumo, A.Y.; Cutie, S.; Hoang, A.; Zhang, H.; Guyot, R.; Lunn, D.; Bigley, R.B.; Yu, H.; Wang, J.; et al. Evidence for hormonal control of heart regenerative capacity during endothermy acquisition. Science 2019, 364, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.I.; O’Meara, C.C.; Gemberling, M.; Zhao, L.; Bryant, D.M.; Zheng, R.; Gannon, J.B.; Cai, L.; Choi, W.-Y.; Egnaczyk, G.F.; et al. Nerves regulate cardiomyocyte proliferation and heart regeneration. Dev. Cell 2015, 34, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Sano, Y.; Ito, S.; Yoneda, M.; Nagasawa, K.; Matsuura, N.; Yamada, Y.; Uchinaka, A.; Bando, Y.K.; Murohara, T.; Nagata, K. Effects of various types of anesthesia on hemodynamics, cardiac function, and glucose and lipid metabolism in rats. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H1360–H1366. [Google Scholar] [CrossRef]
- Konfino, T.; Landa, N.; Ben-Mordechai, T.; Leor, J. The type of injury dictates the mode of repair in neonatal and adult heart. J. Am. Heart Assoc. 2015, 4. [Google Scholar] [CrossRef]
- Bryant, D.M.; O’Meara, C.C.; Ho, N.N.; Gannon, J.; Cai, L.; Lee, R.T. A systematic analysis of neonatal mouse heart regeneration after apical resection. J. Mol. Cell Cardiol. 2015, 79, 315–318. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P1 Rats: Day 1 After Surgery | P1 Sham (n = 15) | P1 MI (n = 20) | p-Value |
|---|---|---|---|
| Heart Rate (bpm) | 290.2 ± 57.0 | 238.2 ± 43.3 | 0.0042 |
| LVWTd (mm) | 0.59 ± 0.02 | 0.58 ± 0.03 | 0.5343 |
| LVIDs (mm) | 1.10 ± 0.21 | 1.86 ± 0.33 | <0.0001 |
| LVIDd (mm) | 2.44 ± 0.21 | 2.51 ± 0.27 | 0.4551 |
| Ejection Fraction (%) | 87.1 ± 4.5 | 53.0 ± 10.4 | <0.0001 |
| Stroke Volume (μL) | 18.5 ± 3.6 | 11.8 ± 2.4 | <0.0001 |
| Cardiac Output (mL/min) | 5.3 ± 1.2 | 2.8 ± 0.8 | <0.0001 |
| P1 Rats: Week 3 After Surgery | P1 Sham (n = 22) | P1 MI (n = 25) | p-Value |
|---|---|---|---|
| Heart Rate (bpm) | 399.4 ± 51.8 | 408.7 ± 43.8 | 0.5060 |
| Female | 399.7 ± 43.7 | 394.8 ± 41.9 | 0.8181 |
| Male | 399.3 ± 55.9 | 423.8 ± 42.3 | 0.2147 |
| LVWTd (mm) | 0.91 ± 0.05 | 0.91 ± 0.06 | 0.9458 |
| Female | 0.92 ± 0.05 | 0.92 ± 0.06 | 0.9327 |
| Male | 0.90 ± 0.06 | 0.90 ± 0.05 | 0.8508 |
| LVIDs (mm) | 2.68 ± 0.26 | 2.76 ± 0.29 | 0.3592 |
| Female | 2.66 ± 0.29 | 2.76 ± 0.30 | 0.5114 |
| Male | 2.69 ± 0.26 | 2.75 ± 0.29 | 0.5462 |
| LVIDd (mm) | 4.50 ± 0.29 | 4.52 ± 0.26 | 0.7801 |
| Female | 4.43 ± 0.30 | 4.52 ± 0.32 | 0.5546 |
| Male | 4.53 ± 0.29 | 4.52 ± 0.20 | 0.9619 |
| Ejection Fraction (%) | 71.1 ± 4.3 | 69.2 ± 6.3 | 0.2511 |
| Female | 70.7 ± 4.2 | 69.1 ± 7.4 | 0.6275 |
| Male | 71.3 ± 4.5 | 69.4 ± 5.3 | 0.3271 |
| Stroke Volume (μL) | 66.2 ± 13.7 | 65.0 ± 10.5 | 0.7259 |
| Female | 62.7 ± 13.1 | 66.4 ± 12.8 | 0.5609 |
| Male | 67.5 ± 14.1 | 63.4 ± 7.5 | 0.3622 |
| Cardiac Output (mL/min) | 26.3 ± 4.6 | 26.7 ± 4.0 | 0.7652 |
| Female | 25.1 ± 3.0 | 26.0 ± 4.7 | 0.6750 |
| Male | 26.8 ± 5.1 | 27.5 ± 3.2 | 0.6743 |
| Adult Rats: Week 3 After Surgery | Adult Sham (n = 3) | Adult MI (n = 3) | p-value |
| Heart Rate (bpm) | 384.5 ± 15.8 | 360 ± 41.9 | 0.3969 |
| LVWTd (mm) | 1.52 ± 0.11 | 0.82 ± 0.07 | 0.0008 |
| LVIDs (mm) | 4.93 ± 0.09 | 8.46 ± 0.18 | <0.0001 |
| LVIDd (mm) | 8.74 ± 0.30 | 10.26 ± 0.44 | 0.0078 |
| Ejection Fraction (%) | 72.5 ± 2.2 | 34.4 ± 3.2 | 0.0001 |
| Stroke Volume (μL) | 290.0 ± 58.9 | 197.1 ± 54.5 | 0.1158 |
| Cardiac Output (mL/min) | 117.1 ± 13.3 | 73.6 ± 7.3 | 0.0078 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Paulsen, M.J.; Hironaka, C.E.; Shin, H.S.; Farry, J.M.; Thakore, A.D.; Jung, J.; Lucian, H.J.; Eskandari, A.; Anilkumar, S.; et al. Natural Heart Regeneration in a Neonatal Rat Myocardial Infarction Model. Cells 2020, 9, 229. https://doi.org/10.3390/cells9010229
Wang H, Paulsen MJ, Hironaka CE, Shin HS, Farry JM, Thakore AD, Jung J, Lucian HJ, Eskandari A, Anilkumar S, et al. Natural Heart Regeneration in a Neonatal Rat Myocardial Infarction Model. Cells. 2020; 9(1):229. https://doi.org/10.3390/cells9010229
Chicago/Turabian StyleWang, Hanjay, Michael J. Paulsen, Camille E. Hironaka, Hye Sook Shin, Justin M. Farry, Akshara D. Thakore, Jinsuh Jung, Haley J. Lucian, Anahita Eskandari, Shreya Anilkumar, and et al. 2020. "Natural Heart Regeneration in a Neonatal Rat Myocardial Infarction Model" Cells 9, no. 1: 229. https://doi.org/10.3390/cells9010229
APA StyleWang, H., Paulsen, M. J., Hironaka, C. E., Shin, H. S., Farry, J. M., Thakore, A. D., Jung, J., Lucian, H. J., Eskandari, A., Anilkumar, S., Wu, M. A., Cabatu, M. C., Steele, A. N., Stapleton, L. M., Zhu, Y., & Woo, Y. J. (2020). Natural Heart Regeneration in a Neonatal Rat Myocardial Infarction Model. Cells, 9(1), 229. https://doi.org/10.3390/cells9010229