Osteopontin-Enhanced Autophagy Attenuates Early Brain Injury via FAK–ERK Pathway and Improves Long-Term Outcome after Subarachnoid Hemorrhage in Rats

 , ,
, ,  ,
,
Abstract
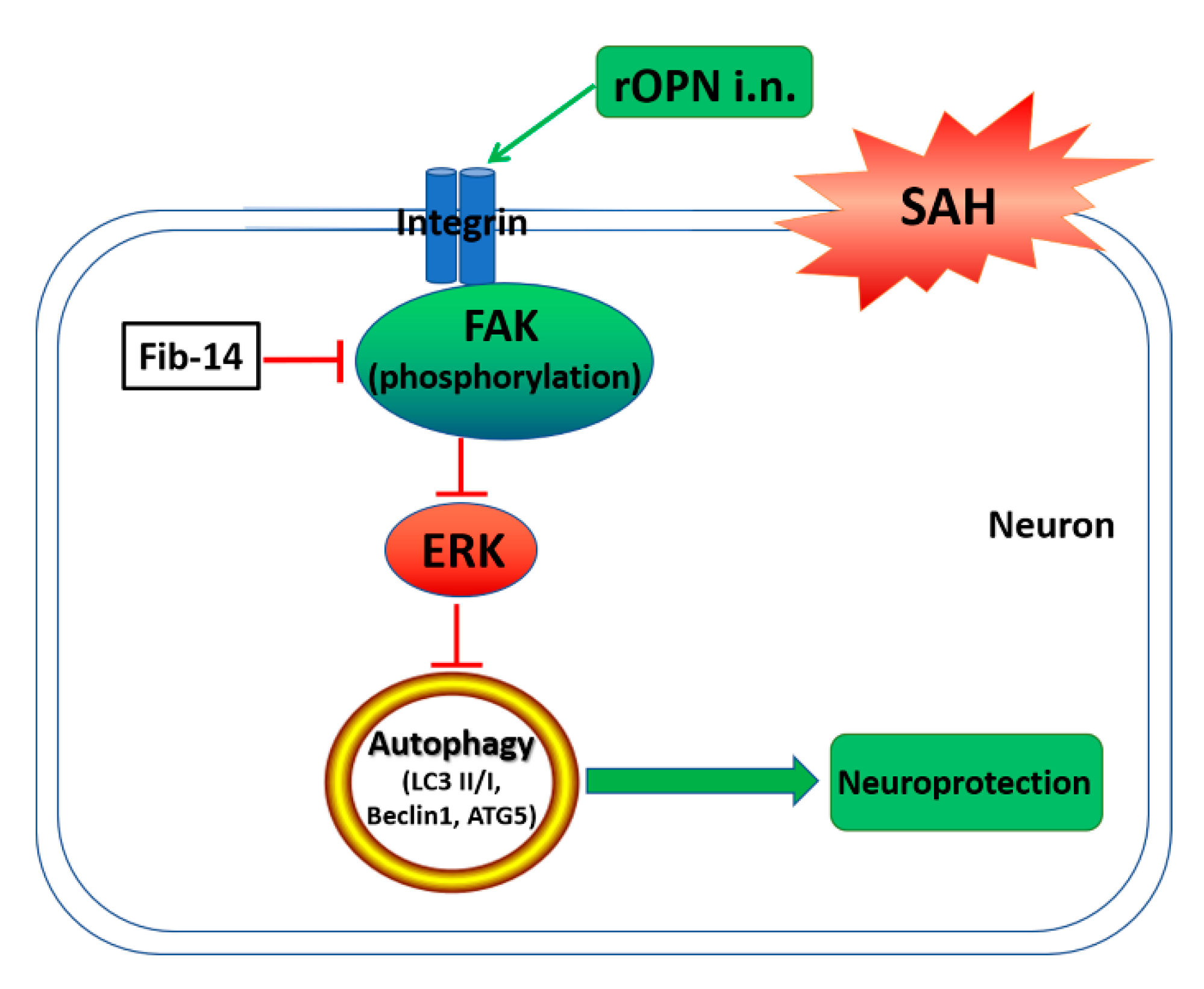
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. SAH Model
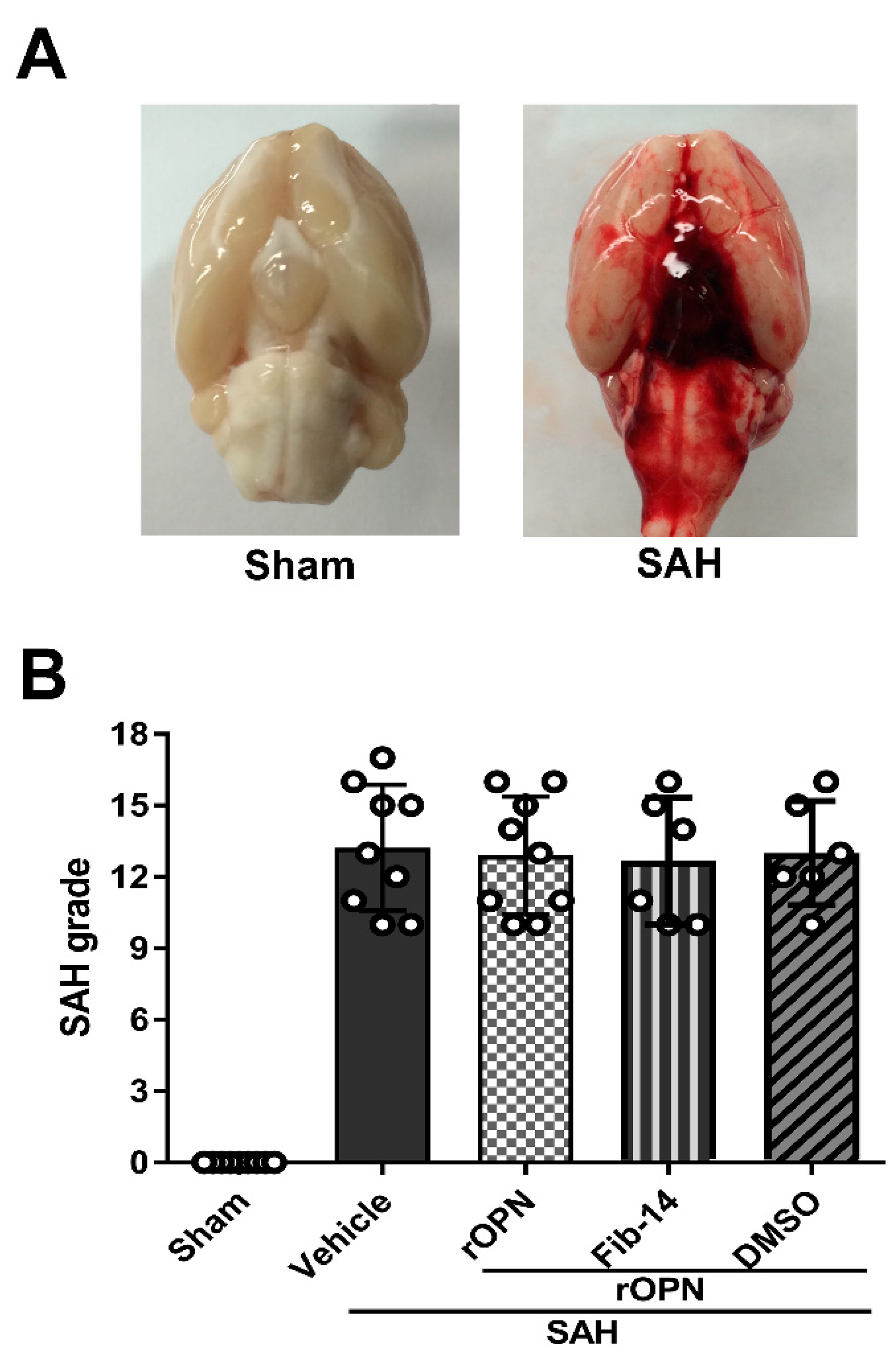
2.3. SAH Grading
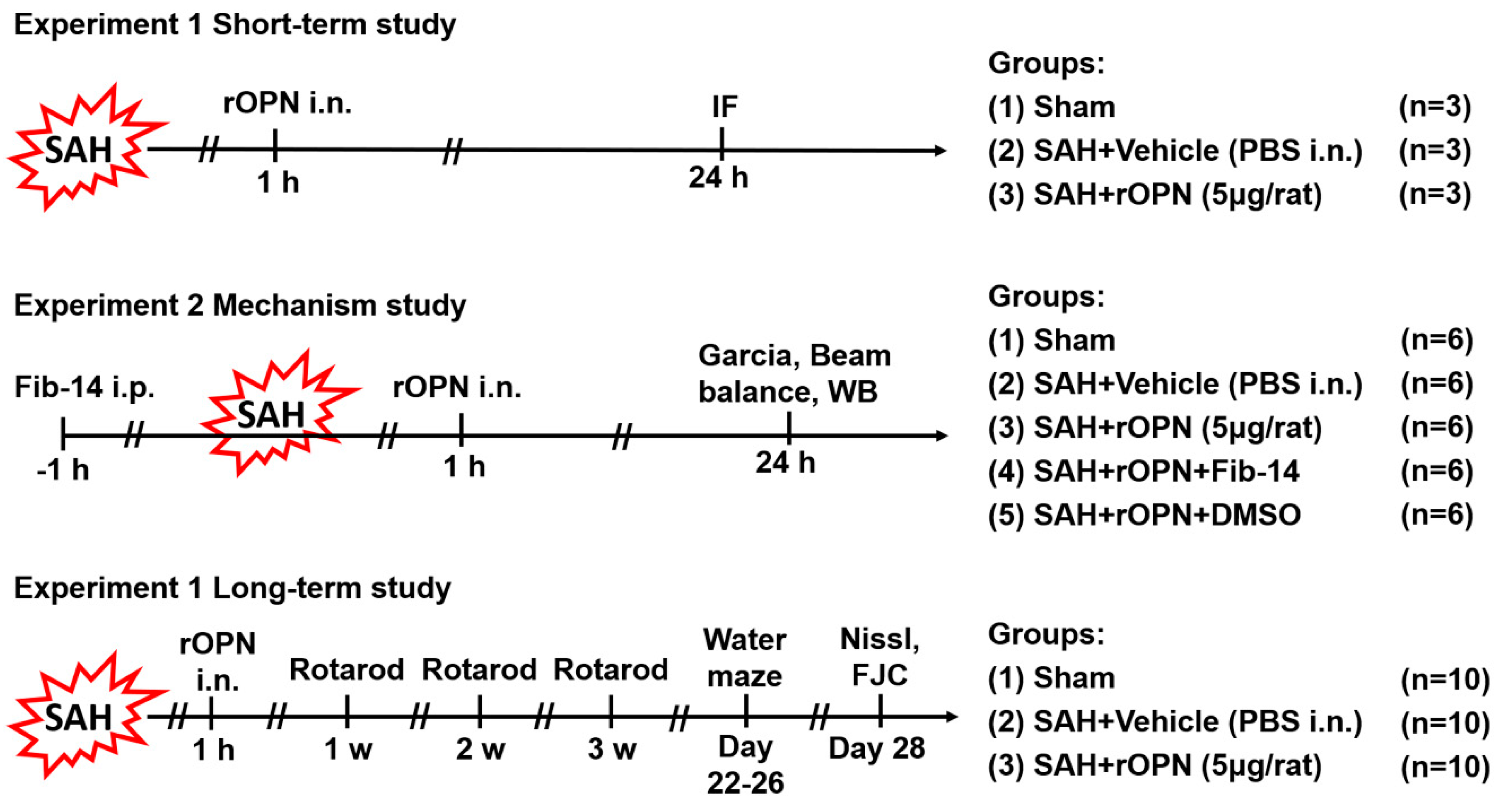
2.4. Experimental Design
2.4.1. Experiment 1
2.4.2. Experiment 2
2.4.3. Experiment 3
2.5. Intranasal Administration
2.6. Short-Term Neurobehavior Assessment
2.7. Long-Term Neurobehavior Assessment
2.8. Western Blot Analysis
2.9. Double Immunofluorescence Staining
2.10. Nissl Staining
2.11. Fluoro-Jade C Staining
2.12. Statistical Analysis
3. Results
3.1. Mortality and SAH Grades
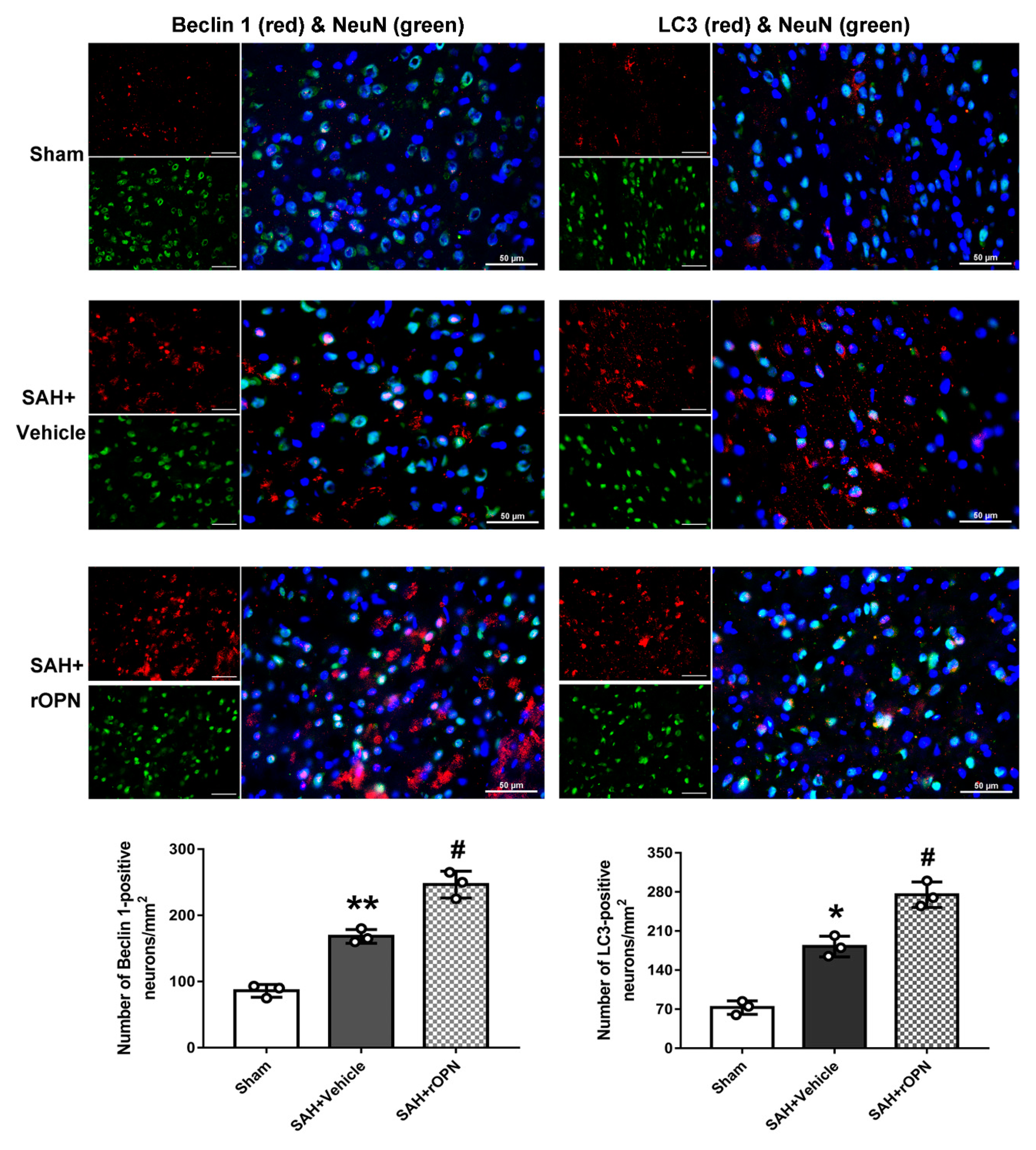
3.2. Nasal Administration of rOPN Increased the Expression of Beclin 1 and LC3 in Neurons 24 h after SAH
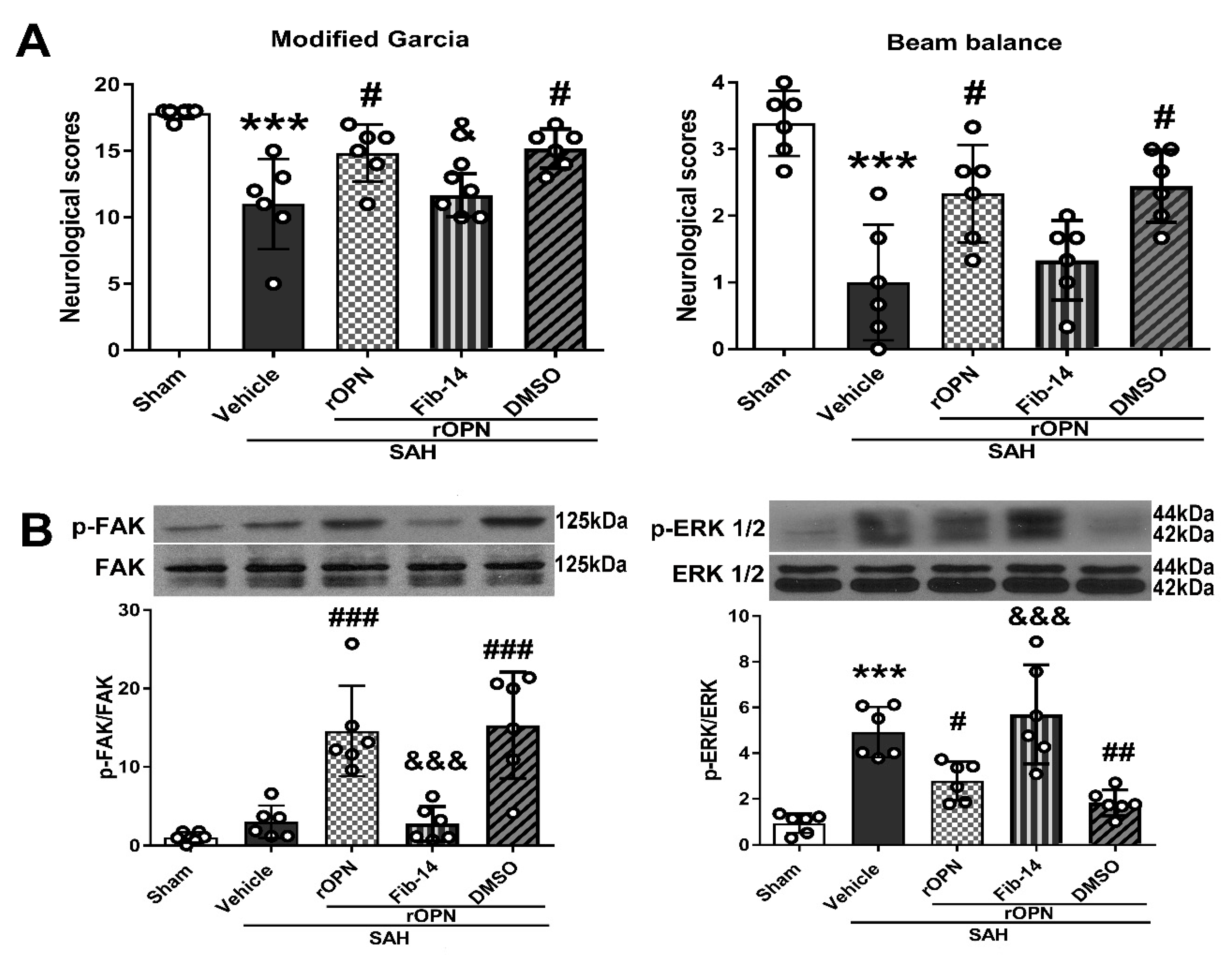
3.3. Administration of FAK Inhibitor Abolished Neurobehavioral Improvement Induced by rOPN
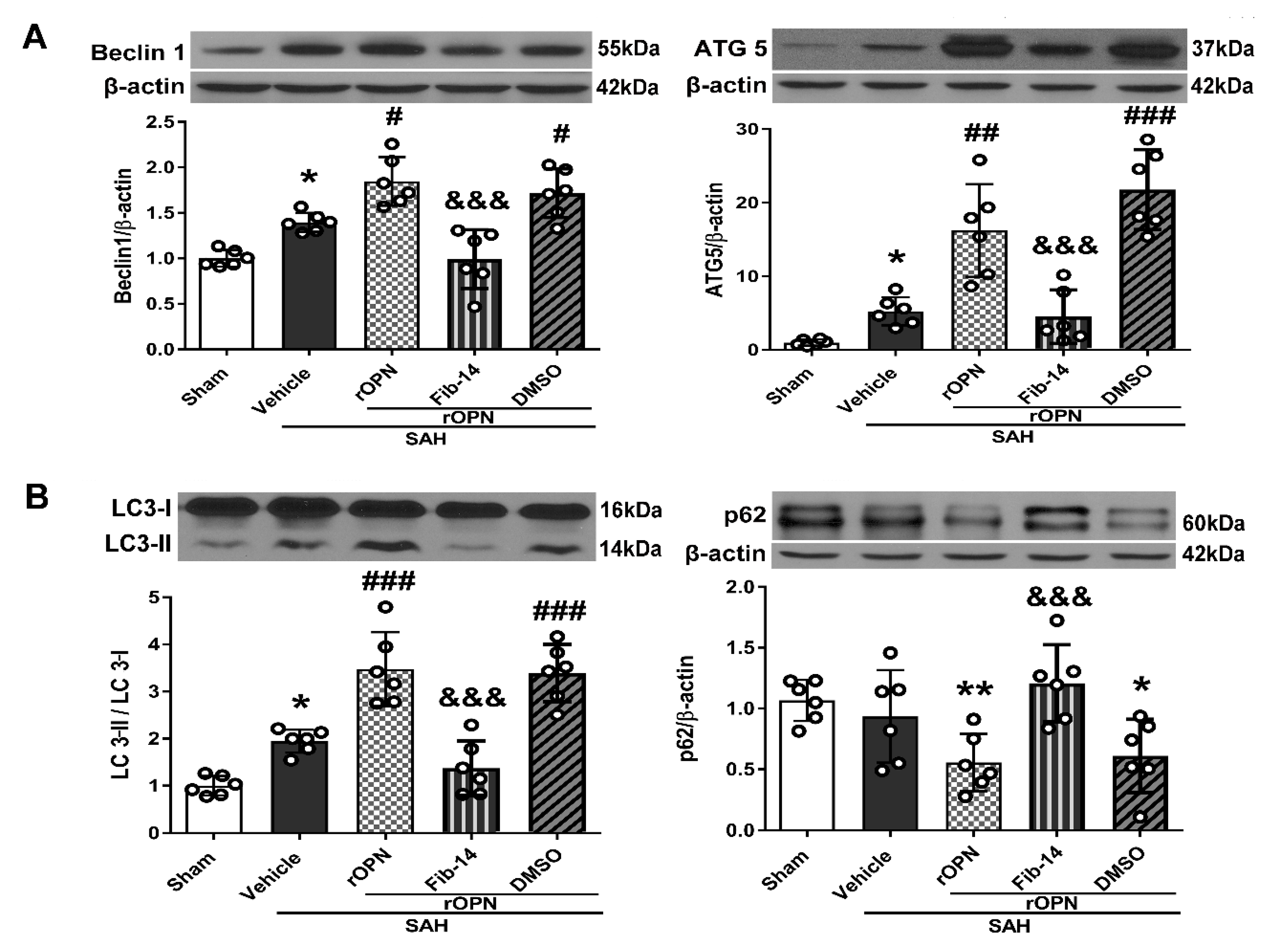
3.4. FAK Inhibitor Partially Reversed rOPN-Enhanced Autophagy Level in the Brain and Blocked Autophagy Flux after SAH
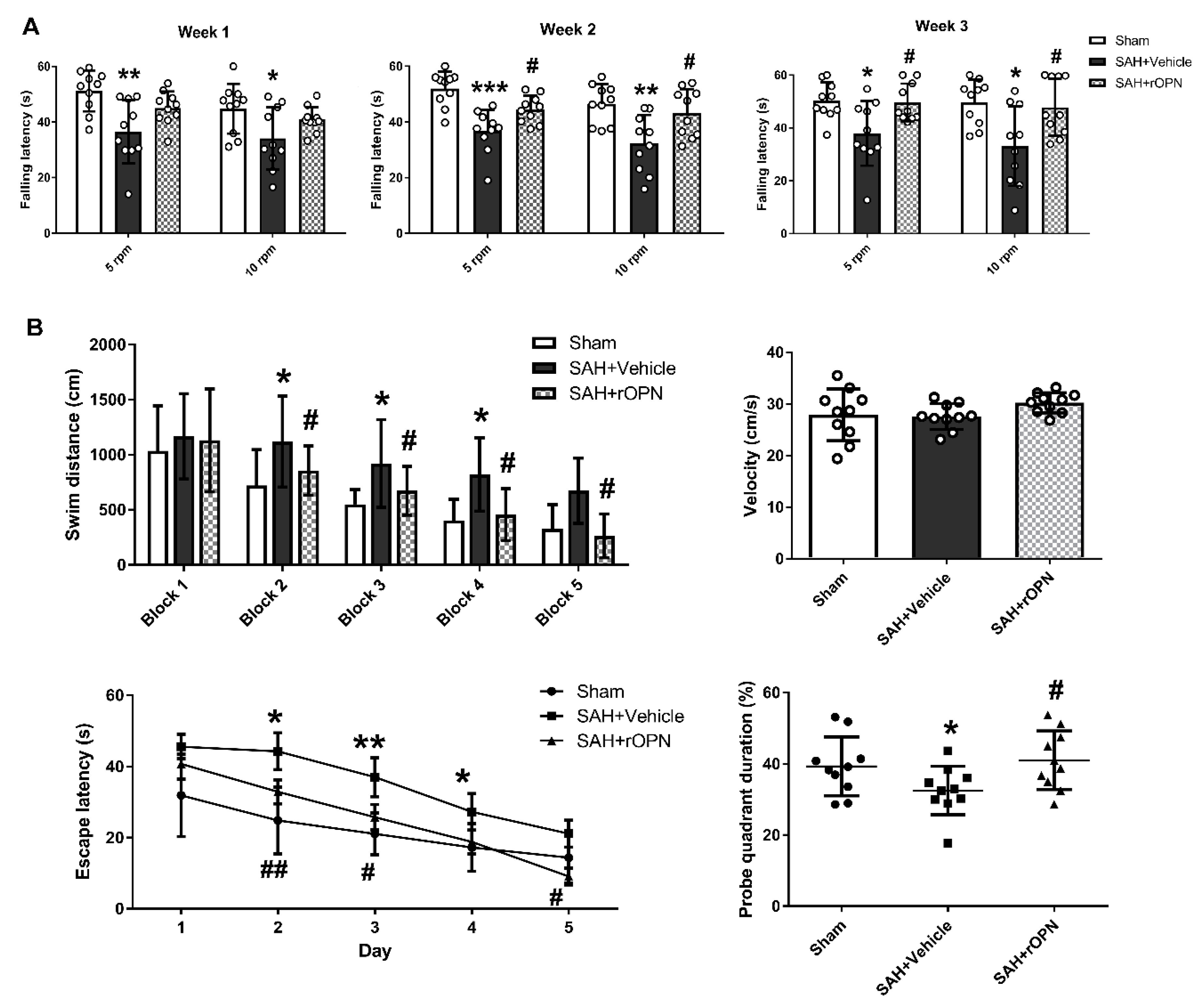
3.5. An Early Nasal Administration of rOPN Improved Long-Term Neurobehavior after SAH
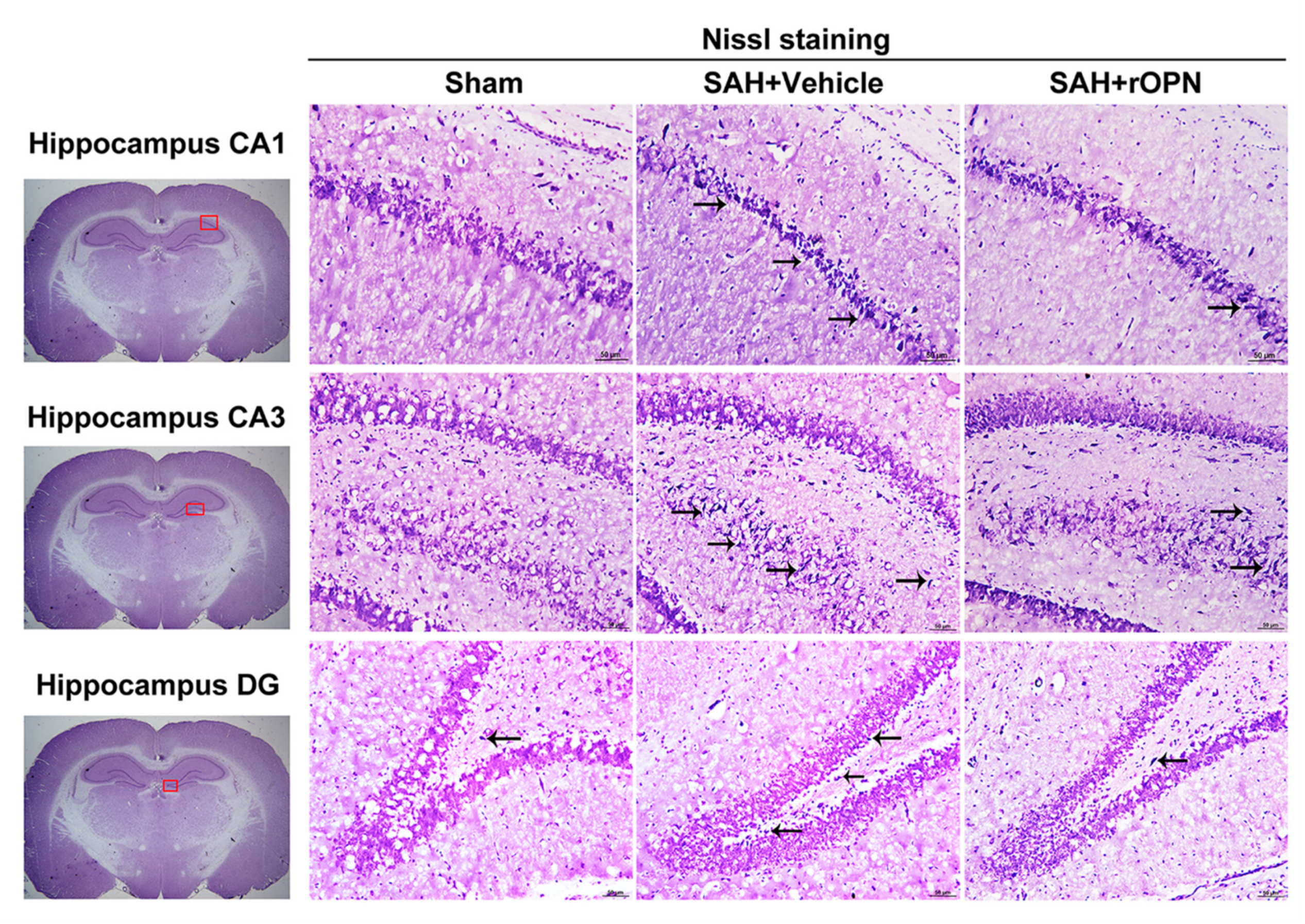
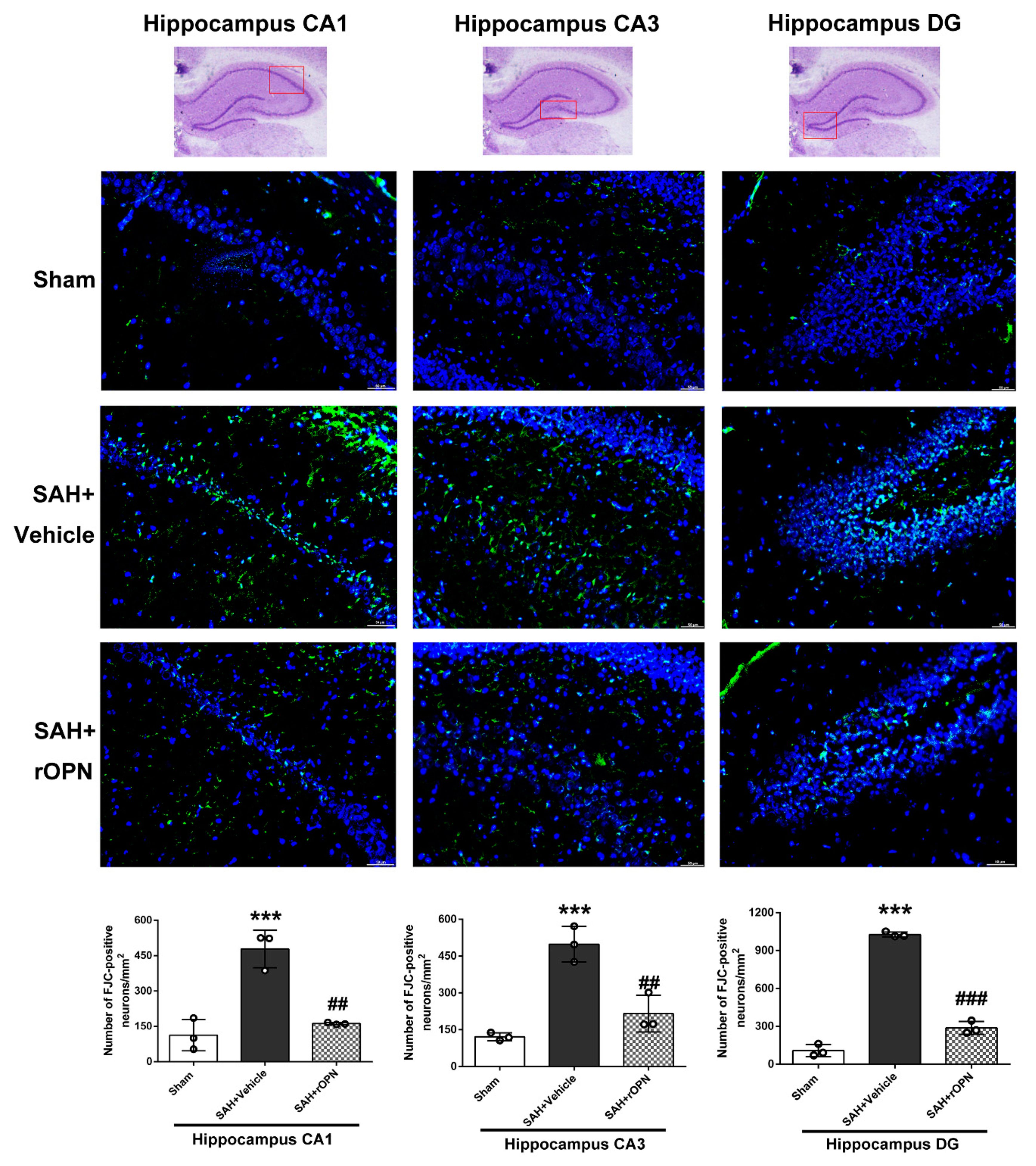
3.6. Nasal Administration of rOPN Decreased Neuronal Deaths in the Hippocampus Region and Attenuated Delayed Brain Injury after SAH
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhu, Q.; Enkhjargal, B.; Huang, L.; Zhang, T.; Sun, C.; Xie, Z.; Wu, P.; Mo, J.; Tang, J.; Xie, Z.; et al. Aggf1 attenuates neuroinflammation and BBB disruption via PI3K/Akt/NF-kappaB pathway after subarachnoid hemorrhage in rats. J. Neuroinflammation 2018, 15, 178. [Google Scholar] [CrossRef]
- Adil, S.M.; Liu, B.; Charalambous, L.T.; Kiyani, M.; Gramer, R.; Swisher, C.B.; Verbick, L.Z.; McCabe, A.; Parente, B.A.; Pagadala, P.; et al. Healthcare Economics of Hydrocephalus After Aneurysmal Subarachnoid Hemorrhage in the United States. Transl. Stroke Res. 2019. [Google Scholar] [CrossRef]
- Cahill, J.; Calvert, J.W.; Zhang, J.H. Mechanisms of early brain injury after subarachnoid hemorrhage. J. Cereb. Blood Flow Metab. 2006, 26, 1341–1353. [Google Scholar] [CrossRef]
- Mo, J.; Enkhjargal, B.; Travis, Z.D.; Zhou, K.; Wu, P.; Zhang, G.; Zhu, Q.; Zhang, T.; Peng, J.; Xu, W.; et al. AVE 0991 attenuates oxidative stress and neuronal apoptosis via Mas/PKA/CREB/UCP-2 pathway after subarachnoid hemorrhage in rats. Redox Biol. 2019, 20, 75–86. [Google Scholar] [CrossRef]
- Li, J.R.; Xu, H.Z.; Nie, S.; Peng, Y.C.; Fan, L.F.; Wang, Z.J.; Wu, C.; Yan, F.; Chen, J.Y.; Gu, C.; et al. Fluoxetine-enhanced autophagy ameliorates early brain injury via inhibition of NLRP3 inflammasome activation following subrachnoid hemorrhage in rats. J. Neuroinflammation 2017, 14, 186. [Google Scholar] [CrossRef]
- Chen, S.; Mei, S.; Luo, Y.; Wu, H.; Zhang, J.; Zhu, J. Gasdermin Family: A Promising Therapeutic Target for Stroke. Transl. Stroke Res. 2018, 9, 555–563. [Google Scholar] [CrossRef]
- Topkoru, B.C.; Altay, O.; Duris, K.; Krafft, P.R.; Yan, J.; Zhang, J.H. Nasal administration of recombinant osteopontin attenuates early brain injury after subarachnoid hemorrhage. Stroke 2013, 44, 3189–3194. [Google Scholar] [CrossRef]
- Yu, L.; Chen, Y.; Tooze, S.A. Autophagy pathway: Cellular and molecular mechanisms. Autophagy 2018, 14, 207–215. [Google Scholar] [CrossRef]
- Jing, C.H.; Wang, L.; Liu, P.P.; Wu, C.; Ruan, D.; Chen, G. Autophagy activation is associated with neuroprotection against apoptosis via a mitochondrial pathway in a rat model of subarachnoid hemorrhage. Neuroscience 2012, 213, 144–153. [Google Scholar] [CrossRef]
- Wang, Z.; Shi, X.Y.; Yin, J.; Zuo, G.; Zhang, J.; Chen, G. Role of autophagy in early brain injury after experimental subarachnoid hemorrhage. J. Mol. Neurosci. 2012, 46, 192–202. [Google Scholar] [CrossRef]
- Chen, J.; Wang, L.; Wu, C.; Hu, Q.; Gu, C.; Yan, F.; Li, J.; Yan, W.; Chen, G. Melatonin-enhanced autophagy protects against neural apoptosis via a mitochondrial pathway in early brain injury following a subarachnoid hemorrhage. J. Pineal Res. 2014, 56, 12–19. [Google Scholar] [CrossRef]
- Meng, N.; Mu, X.; Lv, X.; Wang, L.; Li, N.; Gong, Y. Autophagy represses fascaplysin-induced apoptosis and angiogenesis inhibition via ROS and p8 in vascular endothelia cells. Biomed. Pharmacother. 2019, 114, 108866. [Google Scholar] [CrossRef]
- Galluzzi, L.; Bravo-San Pedro, J.M.; Blomgren, K.; Kroemer, G. Autophagy in acute brain injury. Nat. Rev. Neurosci. 2016, 17, 467–484. [Google Scholar] [CrossRef]
- Shao, A.; Wang, Z.; Wu, H.; Dong, X.; Li, Y.; Tu, S.; Tang, J.; Zhao, M.; Zhang, J.; Hong, Y. Enhancement of Autophagy by Histone Deacetylase Inhibitor Trichostatin A Ameliorates Neuronal Apoptosis After Subarachnoid Hemorrhage in Rats. Mol. Neurobiol. 2016, 53, 18–27. [Google Scholar] [CrossRef]
- Kaleta, B.; Krata, N.; Zagozdzon, R.; Mucha, K. Osteopontin Gene Polymorphism and Urinary OPN Excretion in Patients with Immunoglobulin A Nephropathy. Cells 2019, 8, 524. [Google Scholar] [CrossRef]
- Denhardt, D.T.; Guo, X. Osteopontin: A protein with diverse functions. FASEB J. 1993, 7, 1475–1482. [Google Scholar] [CrossRef]
- Fisher, L.W.; Torchia, D.A.; Fohr, B.; Young, M.F.; Fedarko, N.S. Flexible structures of SIBLING proteins, bone sialoprotein, and osteopontin. Biochem. Biophys. Res. Commun. 2001, 280, 460–465. [Google Scholar] [CrossRef]
- Suzuki, H.; Hasegawa, Y.; Chen, W.; Kanamaru, K.; Zhang, J.H. Recombinant osteopontin in cerebral vasospasm after subarachnoid hemorrhage. Ann. Neurol. 2010, 68, 650–660. [Google Scholar] [CrossRef]
- Zheng, Y.H.; Tian, C.; Meng, Y.; Qin, Y.W.; Du, Y.H.; Du, J.; Li, H.H. Osteopontin stimulates autophagy via integrin/CD44 and p38 MAPK signaling pathways in vascular smooth muscle cells. J. Cell Physiol. 2012, 227, 127–135. [Google Scholar] [CrossRef]
- Liu, G.; Fan, X.; Tang, M.; Chen, R.; Wang, H.; Jia, R.; Zhou, X.; Jing, W.; Wang, H.; Yang, Y.; et al. Osteopontin induces autophagy to promote chemo-resistance in human hepatocellular carcinoma cells. Cancer Lett. 2016, 383, 171–182. [Google Scholar] [CrossRef]
- Sun, C.; Enkhjargal, B.; Reis, C.; Zhou, K.; Xie, Z.; Wu, L.; Zhang, T.; Zhu, Q.; Tang, J.; Jiang, X.; et al. Osteopontin attenuates early brain injury through regulating autophagy-apoptosis interaction after subarachnoid hemorrhage in rats. CNS Neurosci. Ther. in press. [CrossRef]
- Visavadiya, N.P.; Keasey, M.P.; Razskazovskiy, V.; Banerjee, K.; Jia, C.; Lovins, C.; Wright, G.L.; Hagg, T. Integrin-FAK signaling rapidly and potently promotes mitochondrial function through STAT3. Cell Commun. Signal. 2016, 14, 32. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, Y.; Yang, P.; Enkhjargal, B.; Manaenko, A.; Tang, J.; Pearce, W.J.; Hartman, R.; Obenaus, A.; Chen, G.; et al. Recombinant Osteopontin Stabilizes Smooth Muscle Cell Phenotype via Integrin Receptor/Integrin-Linked Kinase/Rac-1 Pathway After Subarachnoid Hemorrhage in Rats. Stroke 2016, 47, 1319–1327. [Google Scholar] [CrossRef]
- Gong, L.; Manaenko, A.; Fan, R.; Huang, L.; Enkhjargal, B.; McBride, D.; Ding, Y.; Tang, J.; Xiao, X.; Zhang, J.H. Osteopontin attenuates inflammation via JAK2/STAT1 pathway in hyperglycemic rats after intracerebral hemorrhage. Neuropharmacology 2018, 138, 160–169. [Google Scholar] [CrossRef]
- Peng, J.; Pang, J.; Huang, L.; Enkhjargal, B.; Zhang, T.; Mo, J.; Wu, P.; Xu, W.; Zuo, Y.; Peng, J.; et al. LRP1 activation attenuates white matter injury by modulating microglial polarization through Shc1/PI3K/Akt pathway after subarachnoid hemorrhage in rats. Redox Biol. 2019, 21, 101121. [Google Scholar] [CrossRef]
- Xie, Z.; Enkhjargal, B.; Reis, C.; Huang, L.; Wan, W.; Tang, J.; Cheng, Y.; Zhang, J.H. Netrin-1 Preserves Blood-Brain Barrier Integrity Through Deleted in Colorectal Cancer/Focal Adhesion Kinase/RhoA Signaling Pathway Following Subarachnoid Hemorrhage in Rats. J. Am. Heart Assoc. 2017, 6. [Google Scholar] [CrossRef]
- Garcia, J.H.; Wagner, S.; Liu, K.-F.; Hu, X.-j. Neurological Deficit and Extent of Neuronal Necrosis Attributable to Middle Cerebral Artery Occlusion in Rats. Stroke 1995, 26, 627–634. [Google Scholar] [CrossRef]
- Chen, H.; Burris, M.; Fajilan, A.; Spagnoli, F.; Tang, J.; Zhang, J.H. Prolonged Exposure to Isoflurane Ameliorates Infarction Severity in the Rat Pup Model of Neonatal Hypoxia-Ischemia. Transl. Stroke Res. 2011, 2, 382–390. [Google Scholar] [CrossRef]
- Sherchan, P.; Lekic, T.; Suzuki, H.; Hasegawa, Y.; Rolland, W.; Duris, K.; Zhan, Y.; Tang, J.; Zhang, J.H. Minocycline improves functional outcomes, memory deficits, and histopathology after endovascular perforation-induced subarachnoid hemorrhage in rats. J. Neurotrauma 2011, 28, 2503–2512. [Google Scholar] [CrossRef]
- Sozen, T.; Tsuchiyama, R.; Hasegawa, Y.; Suzuki, H.; Jadhav, V.; Nishizawa, S.; Zhang, J.H. Role of Interleukin-1β in Early Brain Injury After Subarachnoid Hemorrhage in Mice. Stroke 2009, 40, 2519–2525. [Google Scholar] [CrossRef]
- Li, L.; Tao, Y.; Tang, J.; Chen, Q.; Yang, Y.; Feng, Z.; Chen, Y.; Yang, L.; Yang, Y.; Zhu, G.; et al. A Cannabinoid Receptor 2 Agonist Prevents Thrombin-Induced Blood-Brain Barrier Damage via the Inhibition of Microglial Activation and Matrix Metalloproteinase Expression in Rats. Transl. Stroke Res. 2015, 6, 467–477. [Google Scholar] [CrossRef]
- Gittins, R.; Harrison, P.J. Neuronal density, size and shape in the human anterior cingulate cortex: A comparison of Nissl and NeuN staining. Brain Res. Bull. 2004, 63, 155–160. [Google Scholar] [CrossRef]
- Shi, X.; Xu, L.; Doycheva, D.M.; Tang, J.; Yan, M.; Zhang, J.H. Sestrin2, as a negative feedback regulator of mTOR, provides neuroprotection by activation AMPK phosphorylation in neonatal hypoxic-ischemic encephalopathy in rat pups. J. Cereb. Blood Flow Metab. 2016, 37, 1447–1460. [Google Scholar] [CrossRef]
- Neulen, A.; Meyer, S.; Kramer, A.; Pantel, T.; Kosterhon, M.; Kunzelmann, S.; Goetz, H.; Thal, S.C. Large Vessel Vasospasm Is Not Associated with Cerebral Cortical Hypoperfusion in a Murine Model of Subarachnoid Hemorrhage. Transl. Stroke Res. 2018, 10, 319–326. [Google Scholar] [CrossRef]
- Enkhjargal, B.; McBride, D.W.; Manaenko, A.; Reis, C.; Sakai, Y.; Tang, J.; Zhang, J.H. Intranasal administration of vitamin D attenuates blood-brain barrier disruption through endogenous upregulation of osteopontin and activation of CD44/P-gp glycosylation signaling after subarachnoid hemorrhage in rats. J. Cereb. Blood Flow Metab. 2017, 37, 2555–2566. [Google Scholar] [CrossRef]
- Suzuki, H.; Hasegawa, Y.; Kanamaru, K.; Zhang, J.H. Mechanisms of osteopontin-induced stabilization of blood-brain barrier disruption after subarachnoid hemorrhage in rats. Stroke 2010, 41, 1783–1790. [Google Scholar] [CrossRef]
- Agnihotri, R.; Crawford, H.C.; Haro, H.; Matrisian, L.M.; Havrda, M.C.; Liaw, L. Osteopontin, a Novel Substrate for Matrix Metalloproteinase-3 (Stromelysin-1) and Matrix Metalloproteinase-7 (Matrilysin). J. Biol. Chem. 2001, 276, 28261–28267. [Google Scholar] [CrossRef]
- Ku, M.J.; Kim, J.H.; Lee, J.; Cho, J.Y.; Chun, T.; Lee, S.Y. Maclurin suppresses migration and invasion of human non-small-cell lung cancer cells via anti-oxidative activity and inhibition of the Src/FAK–ERK–β-catenin pathway. Mol. Cell. Biochem. 2015, 402, 243–252. [Google Scholar] [CrossRef]
- Viale-Bouroncle, S.; Gosau, M.; Morsczeck, C. Laminin regulates the osteogenic differentiation of dental follicle cells via integrin-α2/-β1 and the activation of the FAK/ERK signaling pathway. Cell Tissue Res. 2014, 357, 345–354. [Google Scholar] [CrossRef]
- Lu, Z.; Yang, H.; Sutton, M.N.; Yang, M.; Clarke, C.H.; Liao, W.S.; Bast, R.C., Jr. ARHI (DIRAS3) induces autophagy in ovarian cancer cells by downregulating the epidermal growth factor receptor, inhibiting PI3K and Ras/MAP signaling and activating the FOXo3a-mediated induction of Rab7. Cell Death Differ. 2014, 21, 1275–1289. [Google Scholar] [CrossRef]
- Ko, C.P.; Lin, C.W.; Chen, M.K.; Yang, S.F.; Chiou, H.L.; Hsieh, M.J. Pterostilbene induce autophagy on human oral cancer cells through modulation of Akt and mitogen-activated protein kinase pathway. Oral Oncol. 2015, 51, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Chen, Y.; Wang, X.-y.; Lu, R.-f.; Zhang, S.-y.; Tian, M.; Xie, T.; Liu, B.; He, G. Polygonatum odoratum lectin induces apoptosis and autophagy via targeting EGFR-mediated Ras-Raf-MEK-ERK pathway in human MCF-7 breast cancer cells. Phytomedicine 2014, 21, 1658–1665. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chen, H.; Shang, Z.; Jiao, B.; Yuan, B.; Sun, W.; Wang, B.; Miao, M.; Huang, C. SD118-xanthocillin X (1), a novel marine agent extracted from Penicillium commune, induces autophagy through the inhibition of the MEK/ERK pathway. Mar. Drugs 2012, 10, 1345–1359. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wu, P.; Budbazar, E.; Zhu, Q.; Sun, C.; Mo, J.; Peng, J.; Gospodarev, V.; Tang, J.; Shi, H.; et al. Mitophagy Reduces Oxidative Stress Via Keap1 (Kelch-Like Epichlorohydrin-Associated Protein 1)/Nrf2 (Nuclear Factor-E2-Related Factor 2)/PHB2 (Prohibitin 2) Pathway After Subarachnoid Hemorrhage in Rats. Stroke 2019, 50, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.; Ai, J.; Sabri, M.; Tariq, A.; Macdonald, R.L. Learning deficits after experimental subarachnoid hemorrhage in rats. Neuroscience 2010, 169, 1805–1814. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Li, Y.; Feng, D.; Wang, J.; Wen, H.; Liu, D.; Zhao, D.; Liu, H.; Gao, G.; Yin, Z.; et al. Protective effect of HIF-1alpha against hippocampal apoptosis and cognitive dysfunction in an experimental rat model of subarachnoid hemorrhage. Brain Res. 2013, 1517, 114–121. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Ostrowski, R.P.; Sun, X.; Ma, Q.; Huang, B.; Zhan, Y.; Zhang, J.H. CHOP silencing reduces acute brain injury in the rat model of subarachnoid hemorrhage. Stroke 2012, 43, 484–490. [Google Scholar] [CrossRef]
- Suzuki, H. Inflammation: A Good Research Target to Improve Outcomes of Poor-Grade Subarachnoid Hemorrhage. Transl. Stroke Res. 2019. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Mortality | Exclusion |
|---|---|---|
| Experiment 1 Short-term study | ||
| Sham | 0 (0/3) | 0 |
| SAH + Vehicle (PBS) | 25% (1/4) | 1 |
| SAH + rOPN (5 µg/rat) | 25% (1/4) | 0 |
| Experiment 2 Mechanism study | ||
| Sham * | 0 (0/6) | 0 |
| SAH + Vehicle (PBS) * | 14.29 (1/7) | 2 |
| SAH + rOPN (5 µg/rat) * | 25% (2/8) | 0 |
| SAH + rOPN + Fib-14 | 25% (2/8) | 1 |
| SAH + rOPN + DMSO | 14.29% (1/7) | 1 |
| Experiment 3 Long-term study | ||
| Sham | 0 (0/10) | 0 |
| SAH + Vehicle (PBS) | 23.08% (3/13) | 0 |
| SAH + rOPN (5 µg/rat) | 16.67% (2/12) | 0 |
| Total | ||
| Sham | 0 (0/19) | 0 |
| SAH | 20.63% (13/63) | 5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, C.; Enkhjargal, B.; Reis, C.; Zhang, T.; Zhu, Q.; Zhou, K.; Xie, Z.; Wu, L.; Tang, J.; Jiang, X.; et al. Osteopontin-Enhanced Autophagy Attenuates Early Brain Injury via FAK–ERK Pathway and Improves Long-Term Outcome after Subarachnoid Hemorrhage in Rats. Cells 2019, 8, 980. https://doi.org/10.3390/cells8090980
Sun C, Enkhjargal B, Reis C, Zhang T, Zhu Q, Zhou K, Xie Z, Wu L, Tang J, Jiang X, et al. Osteopontin-Enhanced Autophagy Attenuates Early Brain Injury via FAK–ERK Pathway and Improves Long-Term Outcome after Subarachnoid Hemorrhage in Rats. Cells. 2019; 8(9):980. https://doi.org/10.3390/cells8090980
Chicago/Turabian StyleSun, Chengmei, Budbazar Enkhjargal, Cesar Reis, Tongyu Zhang, Qiquan Zhu, Keren Zhou, Zhiyi Xie, Lingyun Wu, Jiping Tang, Xiaodan Jiang, and et al. 2019. "Osteopontin-Enhanced Autophagy Attenuates Early Brain Injury via FAK–ERK Pathway and Improves Long-Term Outcome after Subarachnoid Hemorrhage in Rats" Cells 8, no. 9: 980. https://doi.org/10.3390/cells8090980
APA StyleSun, C., Enkhjargal, B., Reis, C., Zhang, T., Zhu, Q., Zhou, K., Xie, Z., Wu, L., Tang, J., Jiang, X., & Zhang, J. H. (2019). Osteopontin-Enhanced Autophagy Attenuates Early Brain Injury via FAK–ERK Pathway and Improves Long-Term Outcome after Subarachnoid Hemorrhage in Rats. Cells, 8(9), 980. https://doi.org/10.3390/cells8090980