Mutant Huntingtin Affects Diabetes and Alzheimer’s Markers in Human and Cell Models of Huntington’s Disease


Abstract

1. Introduction
1.1. Biology of Huntingtin and Identification of DNA Polymorphism Causing HD
1.2. Neurodegeneration and Metabolic Diseases
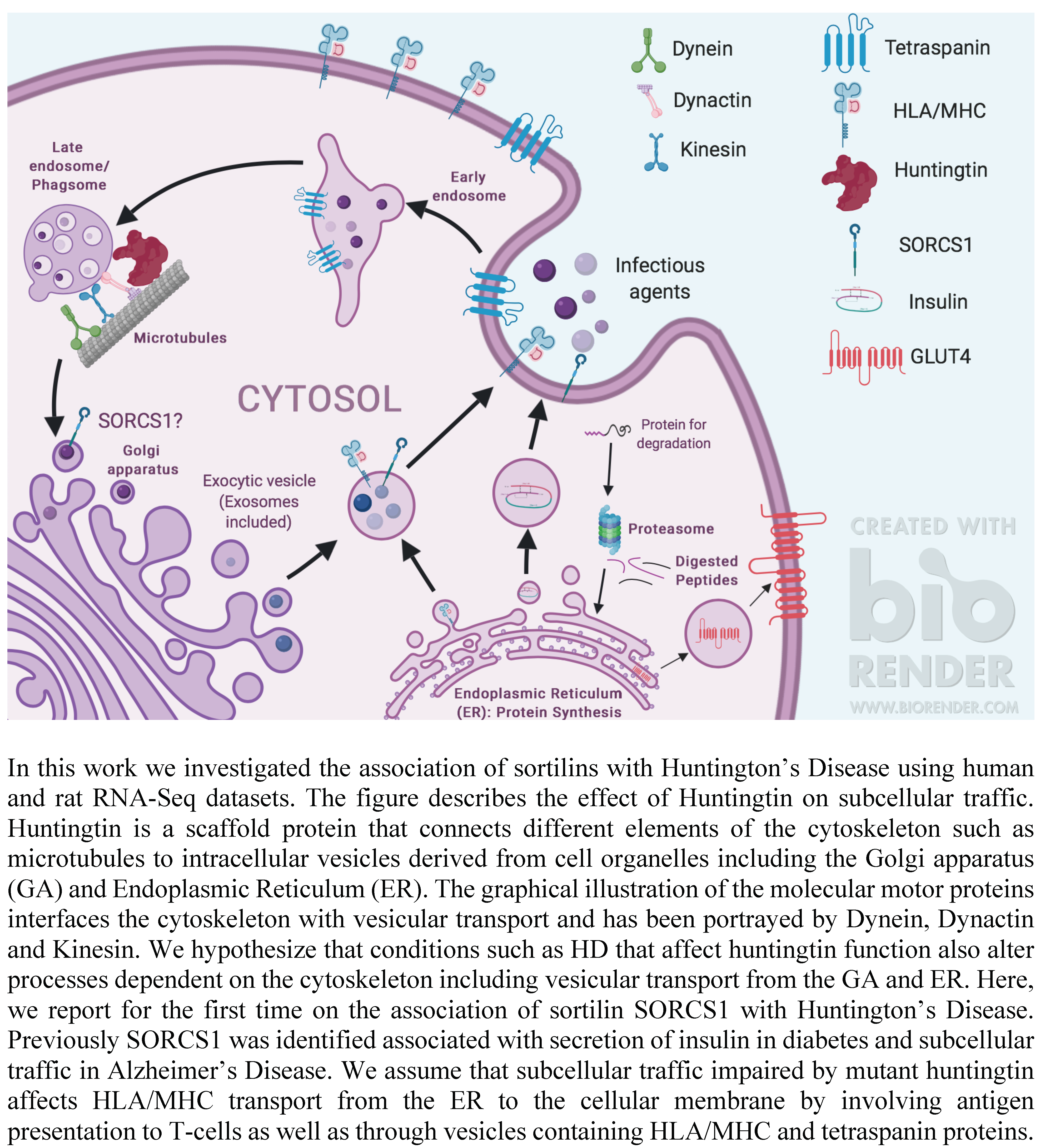
1.3. VPS10P-Domain Receptors or Sortilins: Regulators of Subcellular Protein Trafficking and Markers of Diabetes and Neurodegeneration
2. Materials and Methods
2.1. Library Preparation and Sequencing
2.2. Human and Rat GENOMES
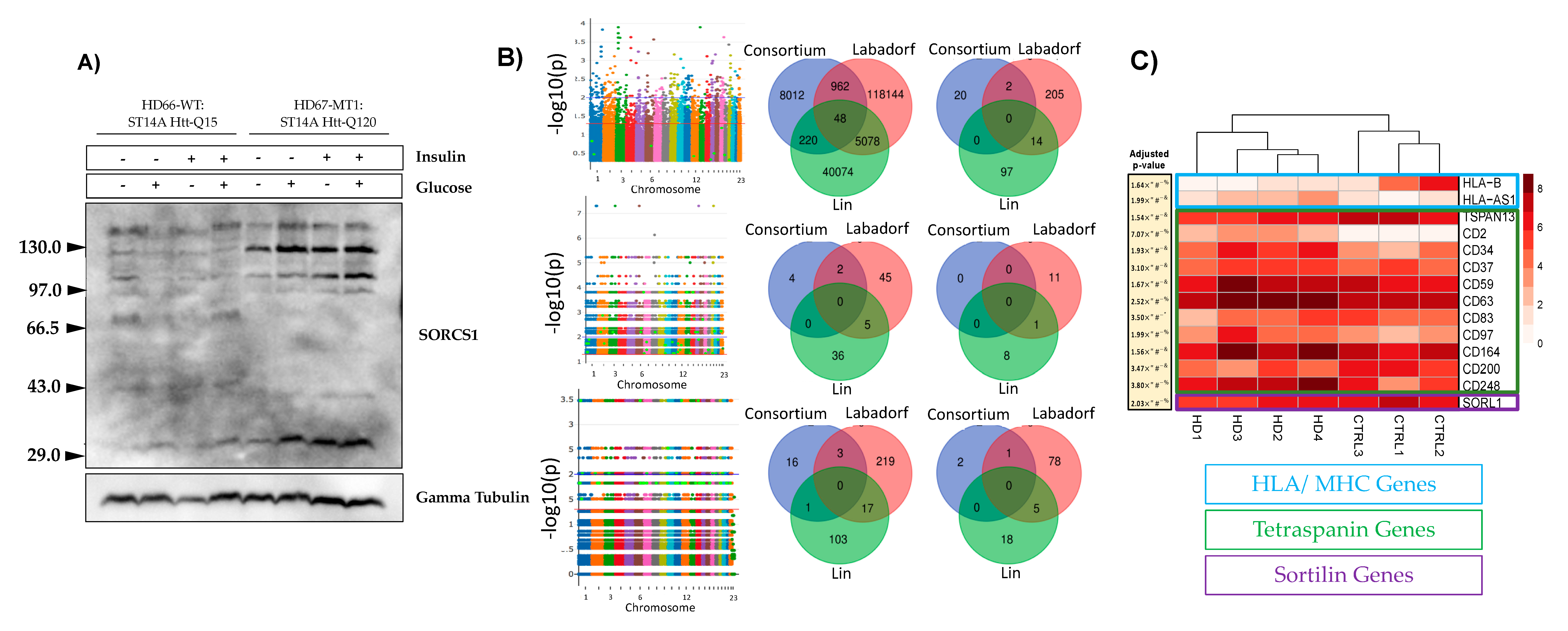
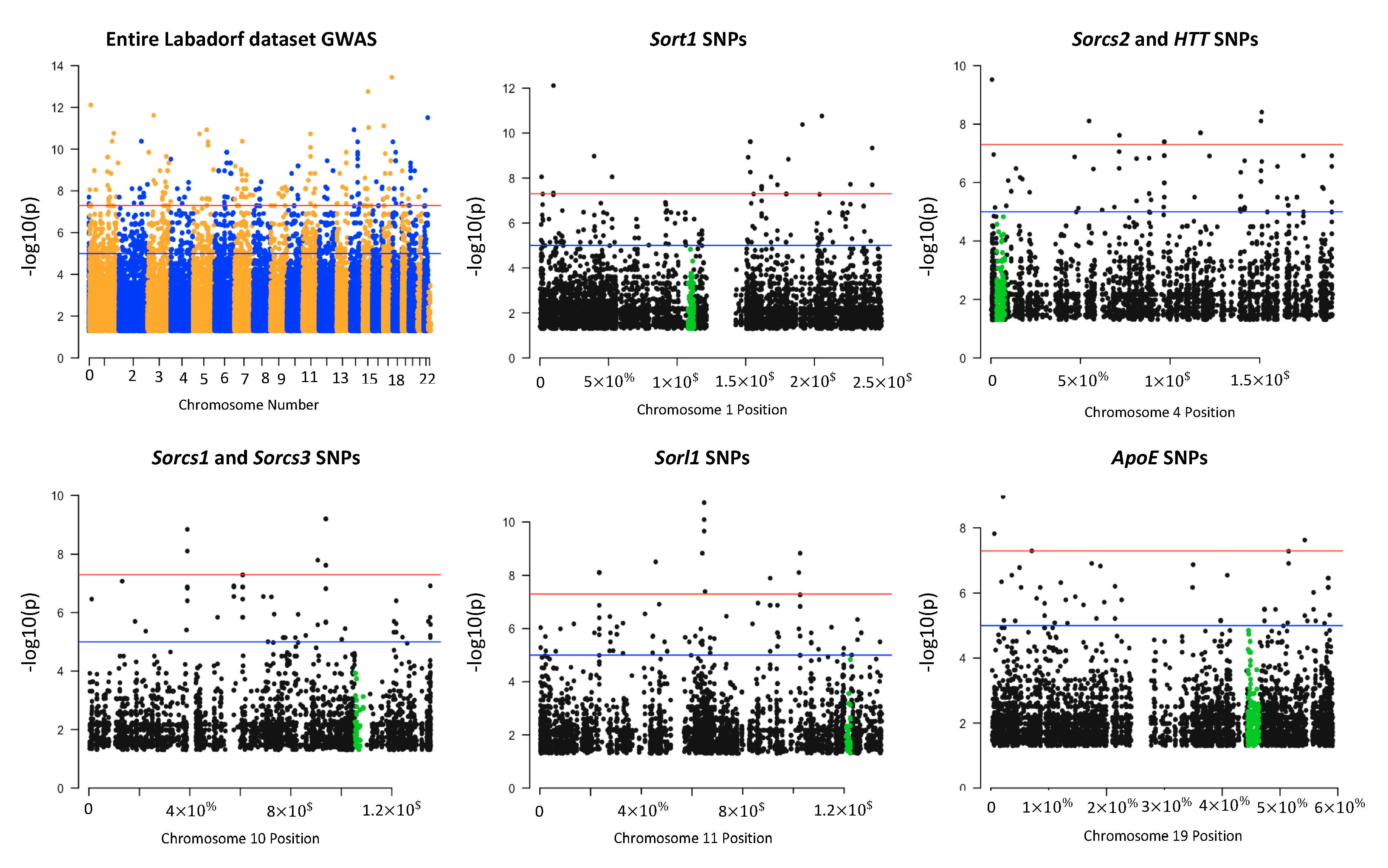
2.3. Genome-Wide Association Study (GWAS)
2.4. Gene Expression Omnibus (GEO) RNA-Seq Datasets
2.5. Post-GATK/GWAS Processing
2.6. Statistical Analysis
2.7. Manhattan Plots
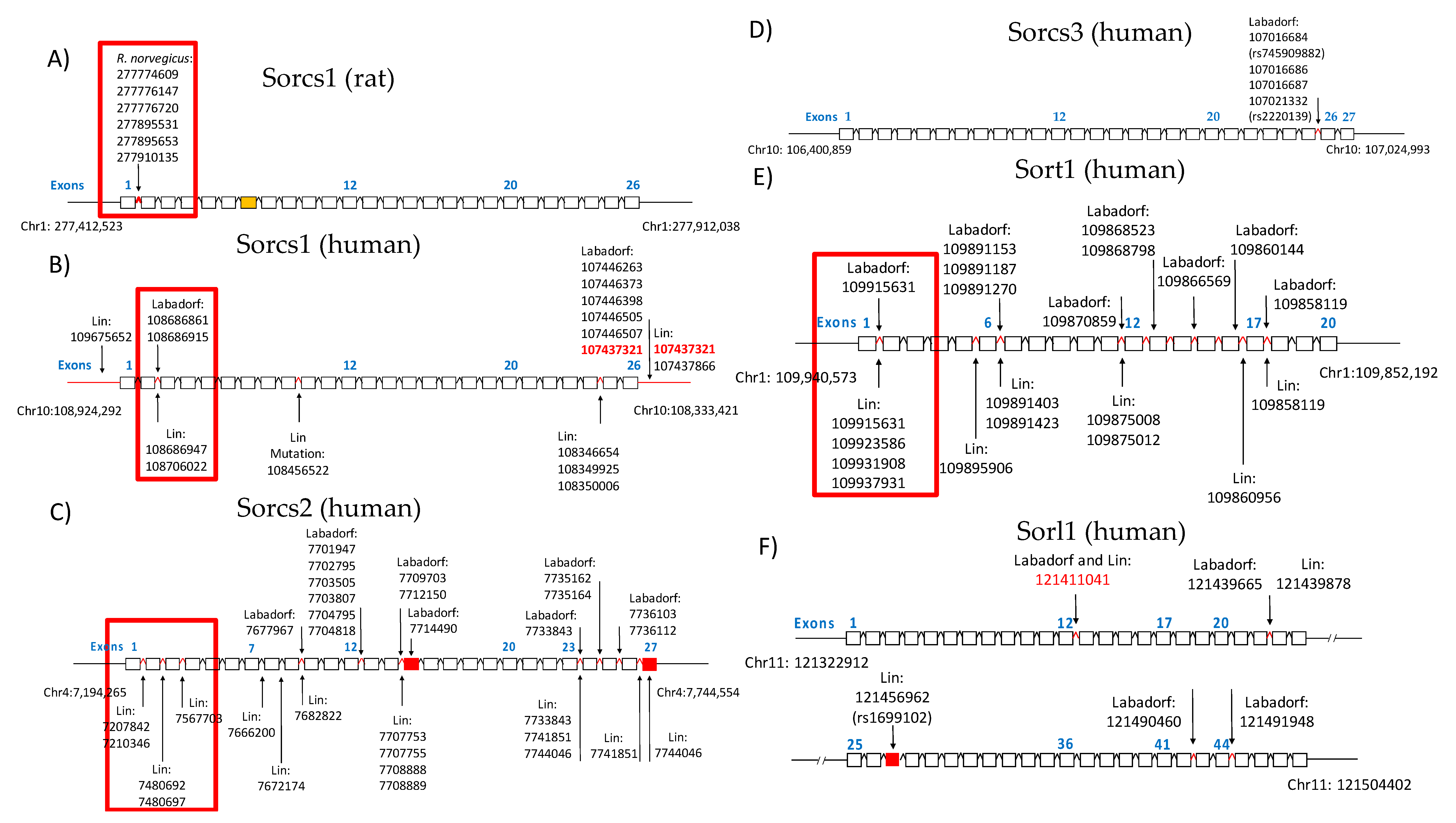
2.8. Linkage Visualization Using Haploview
2.9. Validation of GATK/GWAS DNA Variants Identification by Pyrosequencing
3. Results
3.1. mHTT Is Associated with SORCS1 Protein Up-Regulation and Sortilins SNPs
3.2. mHTT Affects Pathways Important for Immunological Function
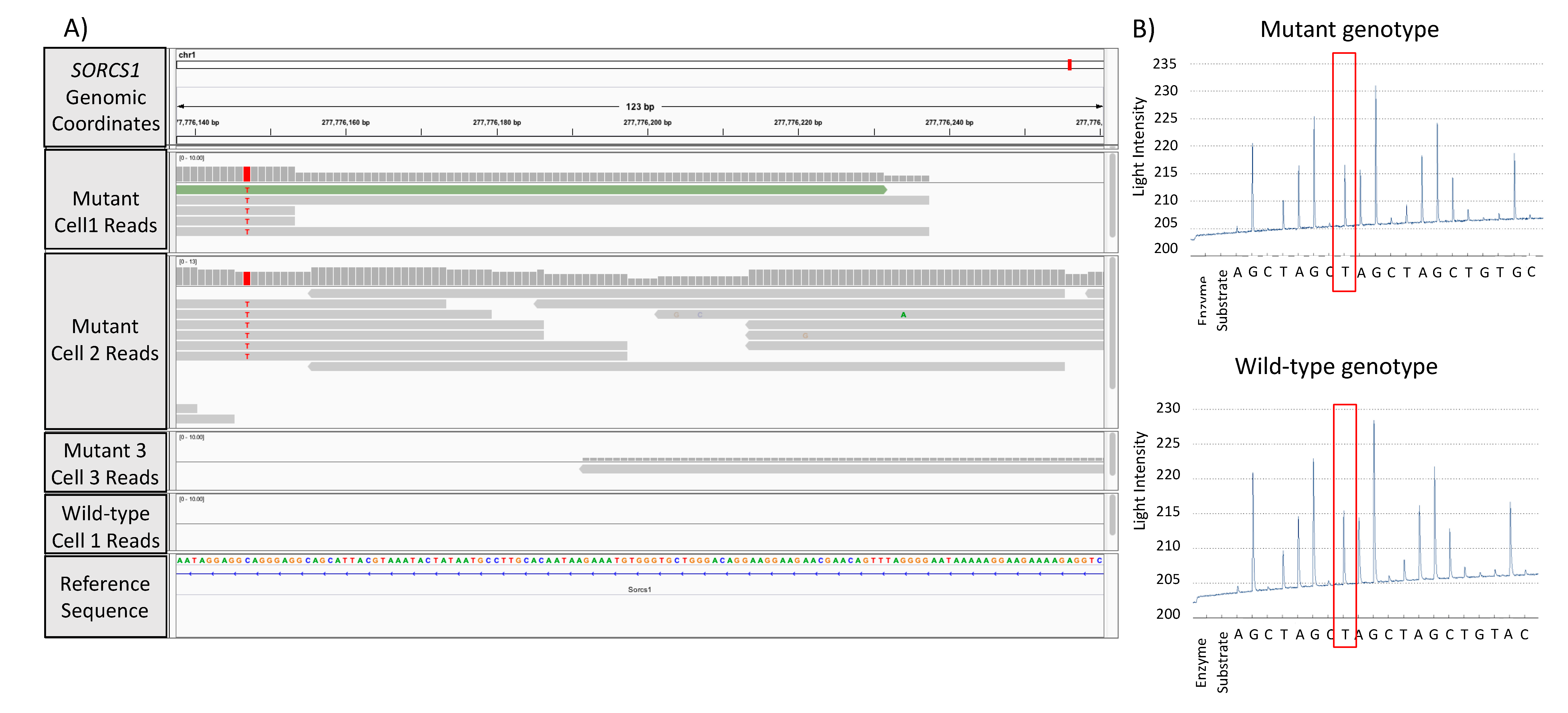
3.3. Validation of GATK/GWAS DNA Variants Identification by Pyrosequencing Methodology
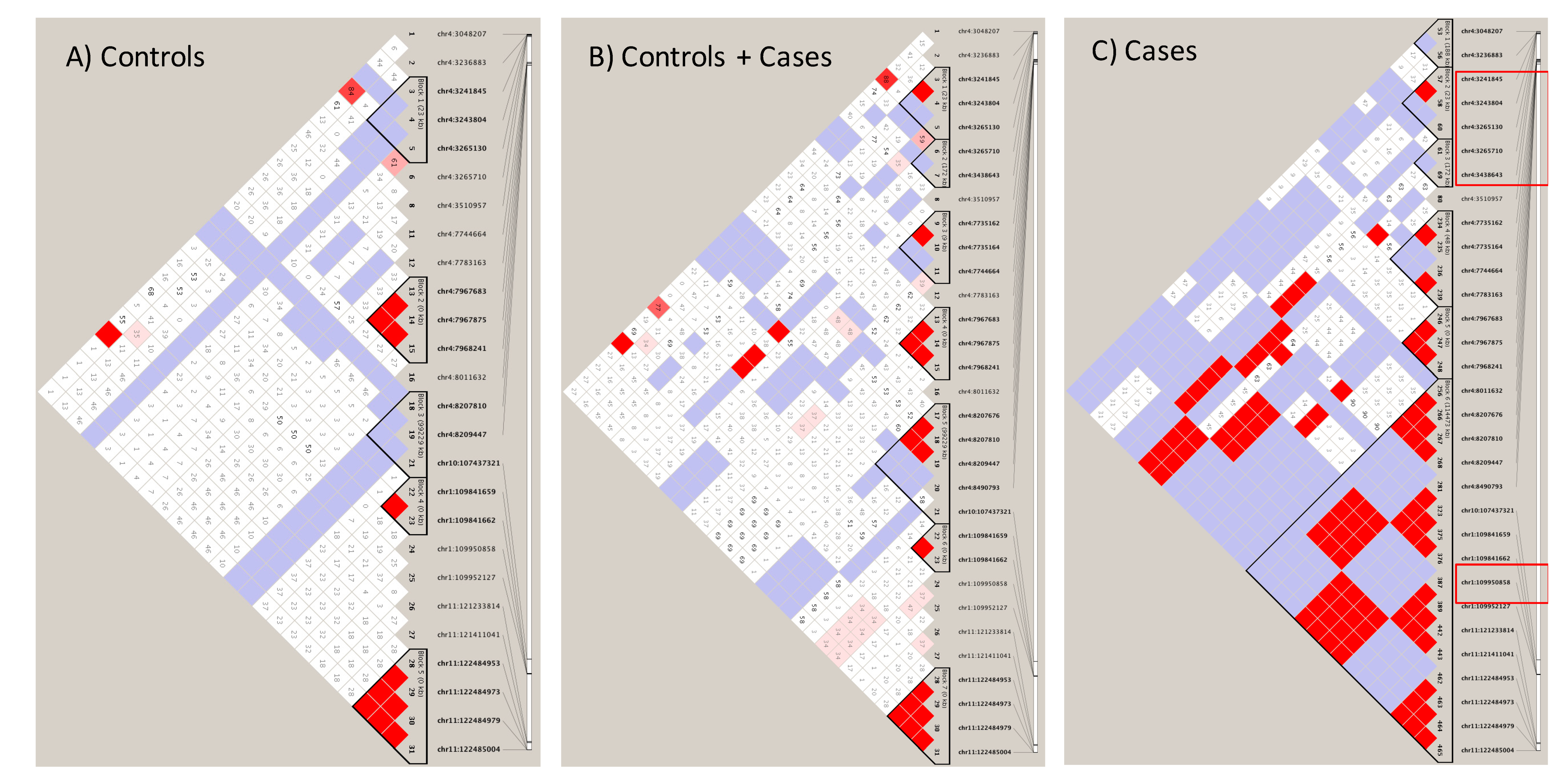
3.4. Sortilin and HTT SNPs Identified by GATK/GWAS Pipeline of RNA-Seq Datasets of Human Origin Are in Linkage Disequilibrium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Montojo, M.; Aganzo, M.; González, N. Huntington’s Disease and Diabetes: Chronological Sequence of its Association. J. Huntington’s Dis. 2017, 6, 179–188. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, M.E.; Ambrose, C.M.; Duyao, M.P.; Myers, R.H.; Lin, C.; Srinidhi, L.; Barnes, G.; Taylor, S.A.; James, M.; Groot, N.; et al. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. Cell 1993, 72, 971–983. [Google Scholar] [CrossRef]
- Palidwor, G.A.; Shcherbinin, S.; Huska, M.R.; Rasko, T.; Stelzl, U.; Arumughan, A.; Foulle, R.; Porras, P.; Sanchez-Pulido, L.; Wanker, E.E.; et al. Detection of Alpha-Rod Protein Repeats Using a Neural Network and Application to Huntingtin. Plos Comput. Biol. 2009, 5, e1000304. [Google Scholar] [CrossRef] [PubMed]
- Gusella, J.F.; Wexler, N.S.; Conneally, M.P.; Naylor, S.L.; Anderson, M.; Tanzi, R.E.; Watkins, P.C.; Ottina, K.; Wallace, M.R.; Sakaguchi, A.Y.; et al. A polymorphic DNA marker genetically linked to Huntington’s disease. Nature 1983, 306, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Warby, S.C.; Montpetit, A.; Hayden, A.R.; Carroll, J.B.; Butland, S.L.; Visscher, H.; Collins, J.A.; Semaka, A.; Hudson, T.J.; Hayden, M.R. CAG Expansion in the Huntington Disease Gene Is Associated with a Specific and Targetable Predisposing Haplogroup. Am. J. Hum. Genet. 2009, 84, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Podolsky, S.; Leopold, N.; Sax, D. Increased frequency of diabetes mellitus in patients with huntington’s chorea. Lancet 1972, 299, 1356–1359. [Google Scholar] [CrossRef]
- Farrer, L.A. Diabetes mellitus in Huntington disease. Clin. Genet. 1985, 27, 62–67. [Google Scholar] [CrossRef]
- Saudou, F.; Humbert, S. The Biology of Huntingtin. Neuron 2016, 89, 910–926. [Google Scholar] [CrossRef]
- Zala, D.; Hinckelmann, M.-V.; Yu, H.; Lyra da Cunha, M.; Liot, G.; Cordelières, F.P.; Marco, S.; Saudou, F. Vesicular Glycolysis Provides On-Board Energy for Fast Axonal Transport. Cell 2013, 152, 479–491. [Google Scholar] [CrossRef]
- McLaughlin, K.A.; Richardson, C.C.; Ravishankar, A.; Brigatti, C.; Liberati, D.; Lampasona, V.; Piemonti, L.; Morgan, D.; Feltbower, R.G.; Christie, M.R. Identification of Tetraspanin-7 as a Target of Autoantibodies in Type 1 Diabetes. Diabetes 2016, 65, 1690–1698. [Google Scholar] [CrossRef]
- Hou, J.; Pessin, J.E. Ins (endocytosis) and outs (exocytosis) of GLUT4 trafficking. Curr. Opin. Cell Biol. 2007, 19, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Kebede, M.A.; Oler, A.T.; Gregg, T.; Balloon, A.J.; Johnson, A.; Mitok, K.; Rabaglia, M.; Schueler, K.; Stapleton, D.; Thorstenson, C.; et al. SORCS1 is necessary for normal insulin secretory granule biogenesis in metabolically stressed β cells. J. Clin. Investig. 2014, 124, 4240–4256. [Google Scholar] [CrossRef] [PubMed]
- Marcusson, E.G.; Horazdovsky, B.F.; Cereghino, J.; Gharakhanian, E.; Emr, S.D. The sorting receptor for yeast vacuolar carboxypeptidase Y is encoded by the VPS10 gene. Cell 1994, 77, 579–586. [Google Scholar] [CrossRef]
- Reitz, C. The role of the retromer complex in aging-related neurodegeneration: A molecular and genomic review. Mol. Genet. Genomics 2015, 290, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Reitz, C.; Tosto, G.; Vardarajan, B.; Rogaeva, E.; Ghani, M.; Rogers, R.S.; Conrad, C.; Haines, J.L.; Pericak-Vance, M.A.; Fallin, M.D.; et al. Independent and epistatic effects of variants in VPS10-d receptors on Alzheimer disease risk and processing of the amyloid precursor protein (APP). Transl. Psychiatry 2013, 3, e256. [Google Scholar] [CrossRef] [PubMed]
- Reitz, C.; Tokuhiro, S.; Clark, L.N.; Conrad, C.; Vonsattel, J.; Hazrati, L.; Palotás, A.; Lantigua, R.; Medrano, M.; Jiménez-Velázquez, I.Z.; et al. SORCS1 alters amyloid precursor protein processing and variants may increase Alzheimer’s disease risk. Ann. Neurol. 2011, 69, 47–64. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, C.L.; Rodgers, A.; Herbst, S.; Coade, S.; Gronow, A.; Guzman, C.A.; Wilson, M.S.; Kanzaki, M.; Nykjaer, A.; Gutierrez, M.G. The proneurotrophin receptor sortilin is required for Mycobacterium tuberculosis control by macrophages. Sci. Rep. 2016, 6, 29332. [Google Scholar] [CrossRef]
- Reuter, E.; Weber, J.; Paterka, M.; Ploen, R.; Breiderhoff, T.; van Horssen, J.; Willnow, T.E.; Siffrin, V.; Zipp, F. Role of Sortilin in Models of Autoimmune Neuroinflammation. J. Immunol. 2015, 195, 5762–5769. [Google Scholar] [CrossRef]
- Chaves, G.; Özel, R.; Rao, N.V.; Hadiprodjo, H.; Costa, Y.; Tokuno, Z.; Pourmand, N. Metabolic and transcriptomic analysis of Huntington’s disease model reveal changes in intracellular glucose levels and related genes. Heliyon 2017, 3, e00381. [Google Scholar] [CrossRef]
- Picelli, S.; Faridani, O.R.; Björklund, Å.K.; Winberg, G.; Sagasser, S.; Sandberg, R. Full-length RNA-seq from single cells using Smart-seq. Nat. Protoc. 2014, 9, 171. [Google Scholar] [CrossRef]
- Nvigen. Available online: www.nvigen.com (accessed on 1 June 2019).
- Ensembl Genome Browser. Available online: https://grch37.ensembl.org/Homo_sapiens/Info/Index; https://mar2015.archive.ensembl.org/Rattus_norvegicus/Info/Index (accessed on 1 June 2019).
- Haploview. Available online: https://learn.gencore.bio.nyu.edu/variant-calling/; broadinstitute.org/haploview/haploview (accessed on 1 June 2019).
- DePristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491. [Google Scholar] [CrossRef] [PubMed]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. Current Protocols in Bioinformatics; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2013. [Google Scholar]
- Visscher, P.M.; Wray, N.R.; Zhang, Q.; Sklar, P.; McCarthy, M.I.; Brown, M.A.; Yang, J. 10 Years of GWAS Discovery: Biology, Function, and Translation. Am. J. Hum. Genetics 2017, 101, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Labadorf, A.; Hoss, A.G.; Lagomarsino, V.; Latourelle, J.C.; Hadzi, T.C.; Bregu, J.; MacDonald, M.E.; Gusella, J.F.; Chen, J.-F.; Akbarian, S.; et al. RNA Sequence Analysis of Human Huntington Disease Brain Reveals an Extensive Increase in Inflammatory and Developmental Gene Expression. PLoS ONE 2015, 10, e0143563. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Park, J.; Ramachandran, S.; Zhang, Y.; Tseng, Y.-T.; Shen, S.; Waldvogel, H.J.; Curtis, M.A.; Faull, R.L.; Troncoso, J.C.; et al. Transcriptome sequencing reveals aberrant alternative splicing in Huntington’s disease. Hum. Mol. Genet. 2016, 25, 3454–3466. [Google Scholar] [CrossRef] [PubMed]
- Consortium, T.H.; Lim, R.G.; Salazar, L.L.; Wilton, D.K.; King, A.R.; Stocksdale, J.T.; Sharifabad, D.; Lau, A.L.; Stevens, B.; Reidling, J.C.; et al. Developmental alterations in Huntington’s disease neural cells and pharmacological rescue in cells and mice. Nat. Neurosci. 2017, 20, 648. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; The Sequence Alignment/Map format and SAMtools. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Turner, S.D. qqman: An R package for visualizing GWAS results using Q-Q and manhattan plots. Biorxiv 2014, 005165. [Google Scholar] [CrossRef]
- Barrett, J.; Fry, B.; Maller, J.; Daly, M. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Pourmand, N.; Elahi, E.; Davis, R.W.; Ronaghi, M. Multiplex Pyrosequencing. Nucleic Acids Res. 2002, 30, e31. [Google Scholar] [CrossRef] [PubMed]
- Royo, J.; Hidalgo, M.; Ruiz, A. Pyrosequencing protocol using a universal biotinylated primer for mutation detection and SNP genotyping. Nat. Protoc. 2007, 2, 1734–1739. [Google Scholar] [CrossRef] [PubMed]
- Schächter, F.; Faure-Delanef, L.; Guénot, F.; Rouger, H.; Froguel, P.; Lesueur-Ginot, L.; Cohen, D. Genetic associations with human longevity at the APOE and ACE loci. Nat. Genet. 1994, 6, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Nejentsev, S.; Howson, J.M.; Walker, N.M.; Szeszko, J.; Field, S.F.; Stevens, H.E.; Reynolds, P.; Hardy, M.; King, E.; Masters, J.; et al. Localization of type 1 diabetes susceptibility to the MHC class I genes HLA-B and HLA-A. Nature 2007, 450, 887. [Google Scholar] [CrossRef]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Cellular and Molecular Immunology, 6th ed.; WB Saunders: St. Louis, MO, USA, 2010. [Google Scholar]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Bu, G. Apolipoprotein E and its receptors in Alzheimer’s disease: Pathways, pathogenesis and therapy. Nat. Rev. Neurosci. 2009, 10, 333–344. [Google Scholar] [CrossRef]
- Scherzer, C.R.; Offe, K.; Gearing, M.; Rees, H.D.; Fang, G.; Heilman, C.J.; Schaller, C.; Bujo, H.; Levey, A.I.; Lah, J.J. Loss of Apolipoprotein E Receptor LR11 in Alzheimer Disease. Arch. Neurol-chicago 2004, 61, 1200–1205. [Google Scholar] [CrossRef]
- Leloup, N.; Lössl, P.; Meijer, D.H.; Brennich, M.; Heck, A.J.; Thies-Weesie, D.M.; Janssen, B.J. Low pH-induced conformational change and dimerization of sortilin triggers endocytosed ligand release. Nat. Commun. 2017, 8, 1708. [Google Scholar] [CrossRef]
- Östensson, M.; Montén, C.; Bacelis, J.; Gudjonsdottir, A.H.; Adamovic, S.; Ek, J.; Ascher, H.; Pollak, E.; Arnell, H.; Browaldh, L.; et al. A Possible Mechanism behind Autoimmune Disorders Discovered By Genome-Wide Linkage and Association Analysis in Celiac Disease. PLoS ONE 2013, 8, e70174. [Google Scholar] [CrossRef]
- Foerster, K.; Freudenberg, J. HLA antigen frequencies in patients with huntington’s chorea and their relatives. J. Neurol. 1980, 223, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Madsen, H.; Nielsen, L.; Sørensen, S. An association study of Huntington’s disease and HLA. J. Med. Genet. 1982, 19, 452–454. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | Total Number of SNPs Identified | Significant SNPs (p < 0.05) | Sortilin Gene Body Size (bp) | SNPs in Gene Body (p < 0.05) | SNPs in Vicinity (±106 bp Away from Gene Body) | |
|---|---|---|---|---|---|---|
| Rattus norvegicus (ST14A cells) | 222997 | 8464 (3.80%) | SORCS1 | 499515 | 6 | 13 |
| SORCS2 | 371129 | 0 | 8 | |||
| SORCS3 | 641740 | 0 | 2 | |||
| SORT1 | 89539 | 0 | 11 | |||
| SORL1 | 170117 | 0 | 5 | |||
| Labadorf | 2658838 | 84349 (3.17%) | SORCS1 | 590871 | 2 | 8 |
| SORCS2 | 550289 | 15 | 171 | |||
| SORCS3 | 624134 | 4 | 53 | |||
| SORT1 | 88381 | 9 | 153 | |||
| SORL1 | 181490 | 4 | 38 | |||
| Lin | 3172675 | 44669 (1.4%) | SORCS1 | 590871 | 6 | 9 |
| SORCS2 | 550289 | 10 | 121 | |||
| SORCS3 | 624134 | 0 | 23 | |||
| SORT1 | 88381 | 11 | 111 | |||
| SORL1 | 181490 | 4 | 41 | |||
| HD iPSC Consortium (iPSC Cells) | 238013 | 8979 (3.38%) | SORCS1 | 590871 | 0 | 0 |
| SORCS2 | 550289 | 0 | 20 | |||
| SORCS3 | 624134 | 0 | 3 | |||
| SORT1 | 88381 | 0 | 22 | |||
| SORL1 | 181490 | 3 | 6 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaves, G.; Stanley, J.; Pourmand, N. Mutant Huntingtin Affects Diabetes and Alzheimer’s Markers in Human and Cell Models of Huntington’s Disease. Cells 2019, 8, 962. https://doi.org/10.3390/cells8090962
Chaves G, Stanley J, Pourmand N. Mutant Huntingtin Affects Diabetes and Alzheimer’s Markers in Human and Cell Models of Huntington’s Disease. Cells. 2019; 8(9):962. https://doi.org/10.3390/cells8090962
Chicago/Turabian StyleChaves, Gepoliano, John Stanley, and Nader Pourmand. 2019. "Mutant Huntingtin Affects Diabetes and Alzheimer’s Markers in Human and Cell Models of Huntington’s Disease" Cells 8, no. 9: 962. https://doi.org/10.3390/cells8090962
APA StyleChaves, G., Stanley, J., & Pourmand, N. (2019). Mutant Huntingtin Affects Diabetes and Alzheimer’s Markers in Human and Cell Models of Huntington’s Disease. Cells, 8(9), 962. https://doi.org/10.3390/cells8090962