gga-miRNA-18b-3p Inhibits Intramuscular Adipocytes Differentiation in Chicken by Targeting the ACOT13 Gene

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cell Isolation, Culture, and Differentiation of Primary Preadipocytes
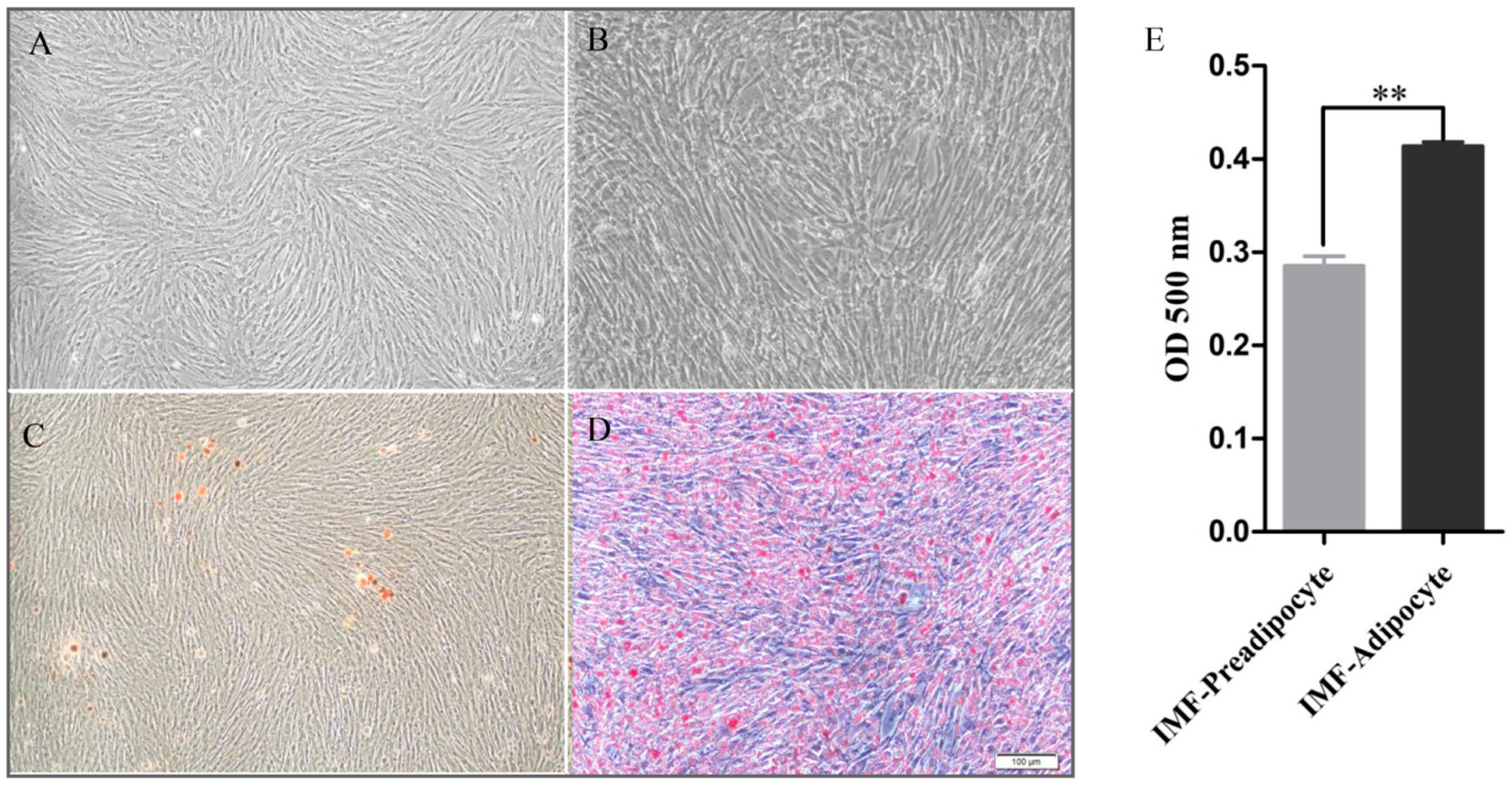
2.3. Oil Red O Staining and Cellular TG Content Measurement
2.4. Small RNAs Library Construction and Deep Sequencing
2.5. Identification and Prediction of miRNAs
2.6. Prediction of miRNA Target Genes and GO and Pathway Enrichment Analysis
2.7. Functional Assays
2.7.1. Vector Construction
2.7.2. Luciferase Assays
2.7.3. Overexpression and Knockdown Assay in Preadipocytes
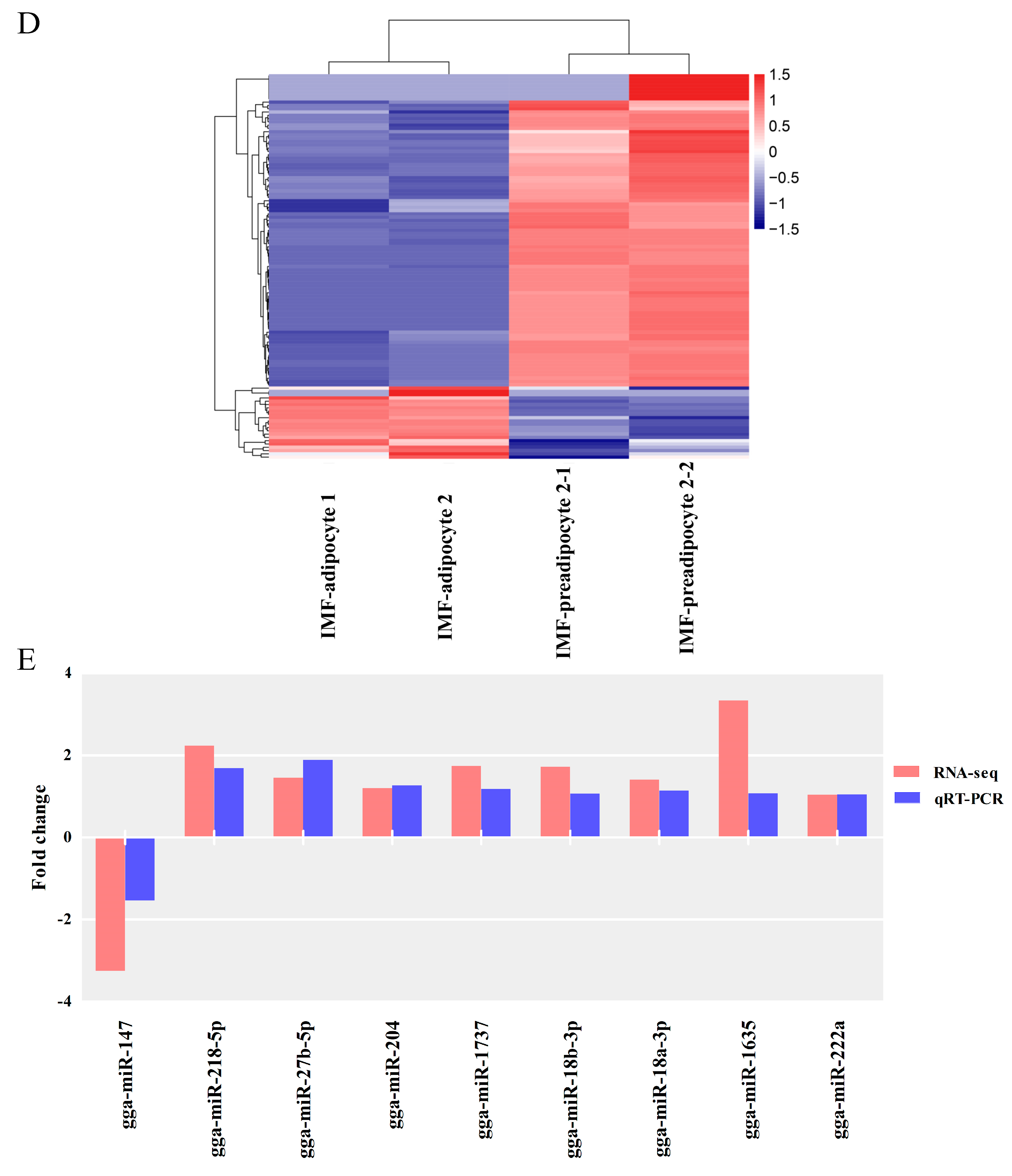
2.7.4. Quantitative PCR
2.8. Statistical Analysis
3. Results
3.1. Phenotypic Variations between IMF-Preadipocytes and IMF-Adipocytes
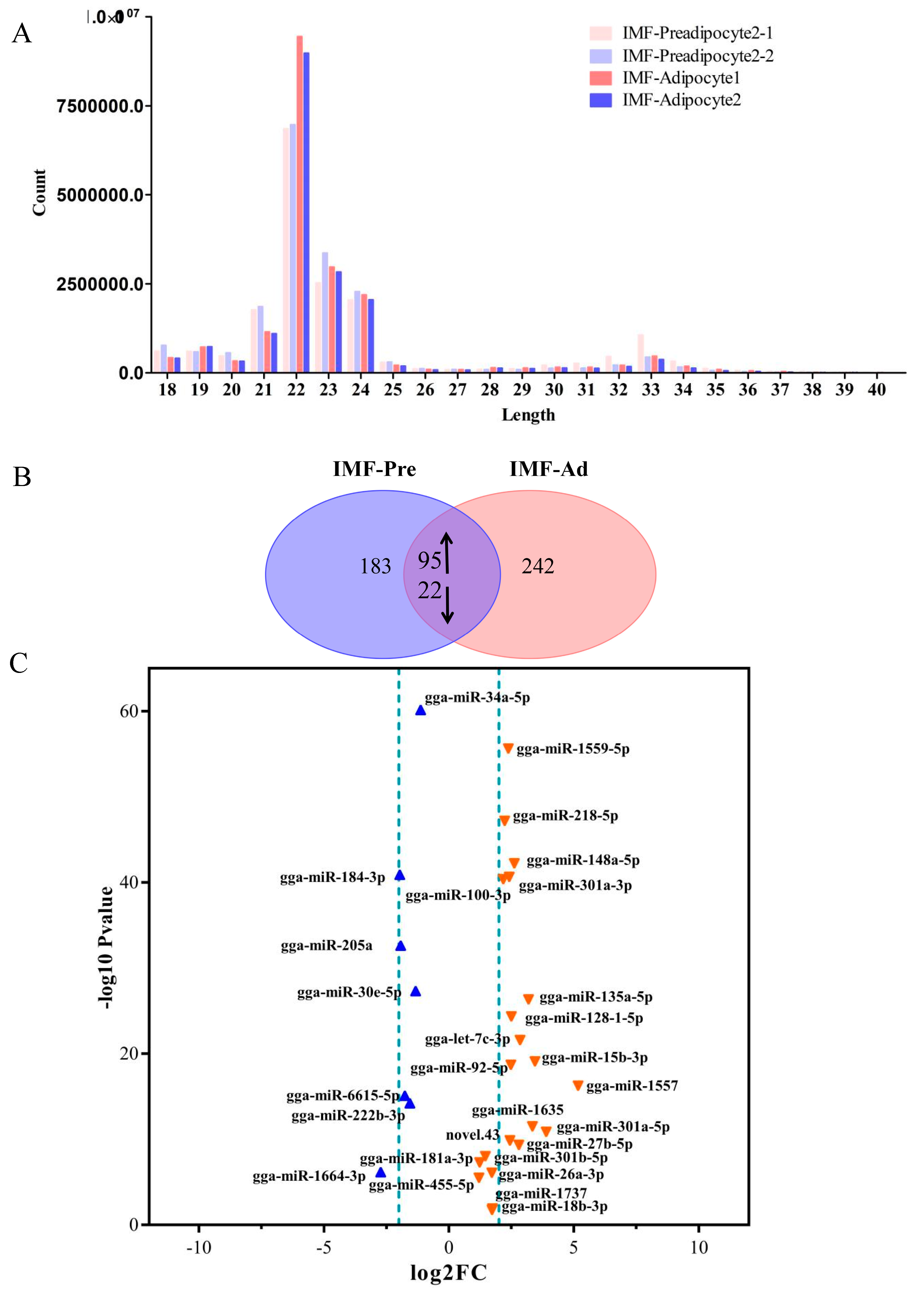
3.2. Summary of Sequencing Data
3.3. Screening for Differentially Expressed miRNAs
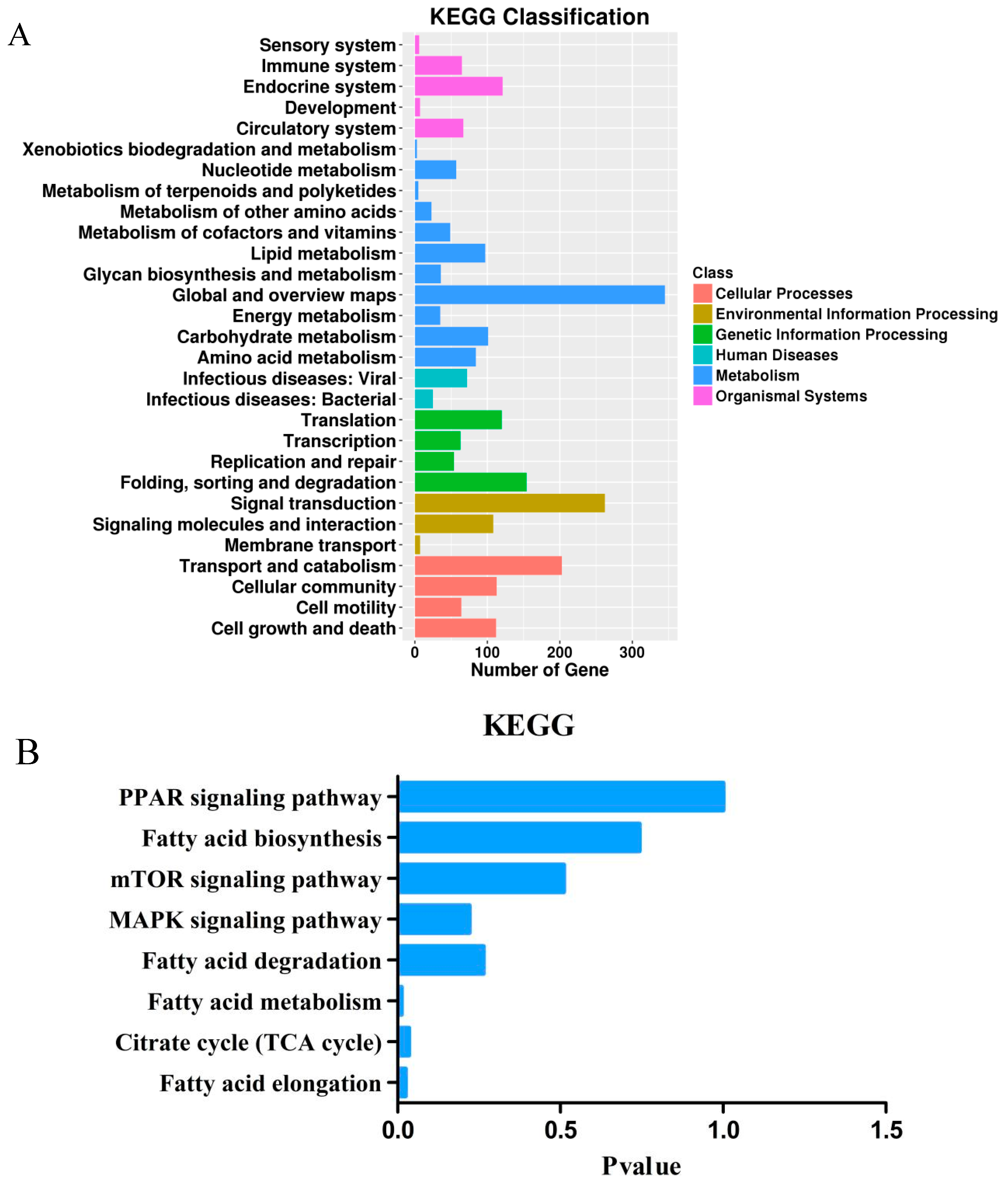
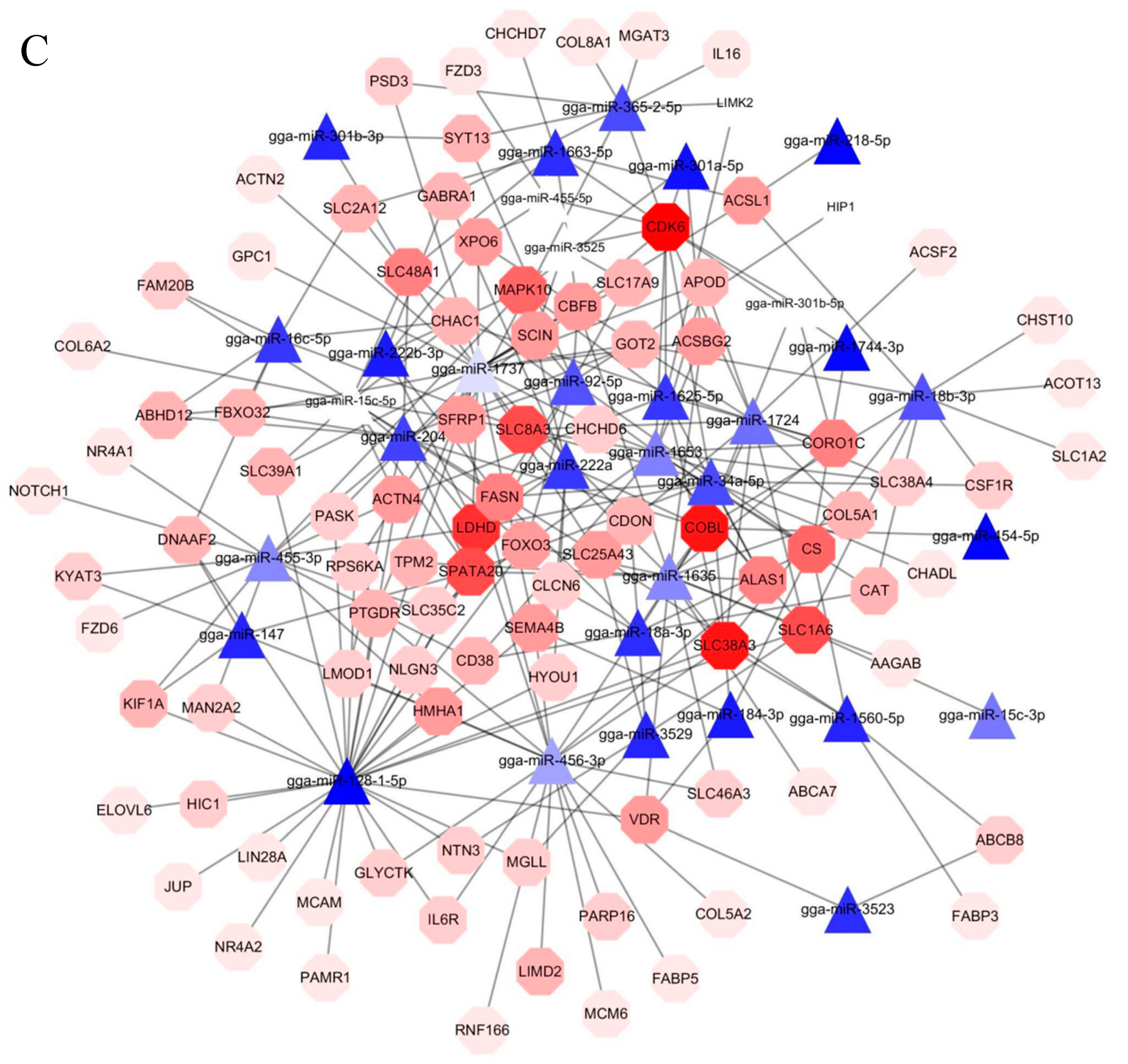
3.4. Target Gene Prediction, Functional Enrichment Analysis, and Candidate Genes Identification
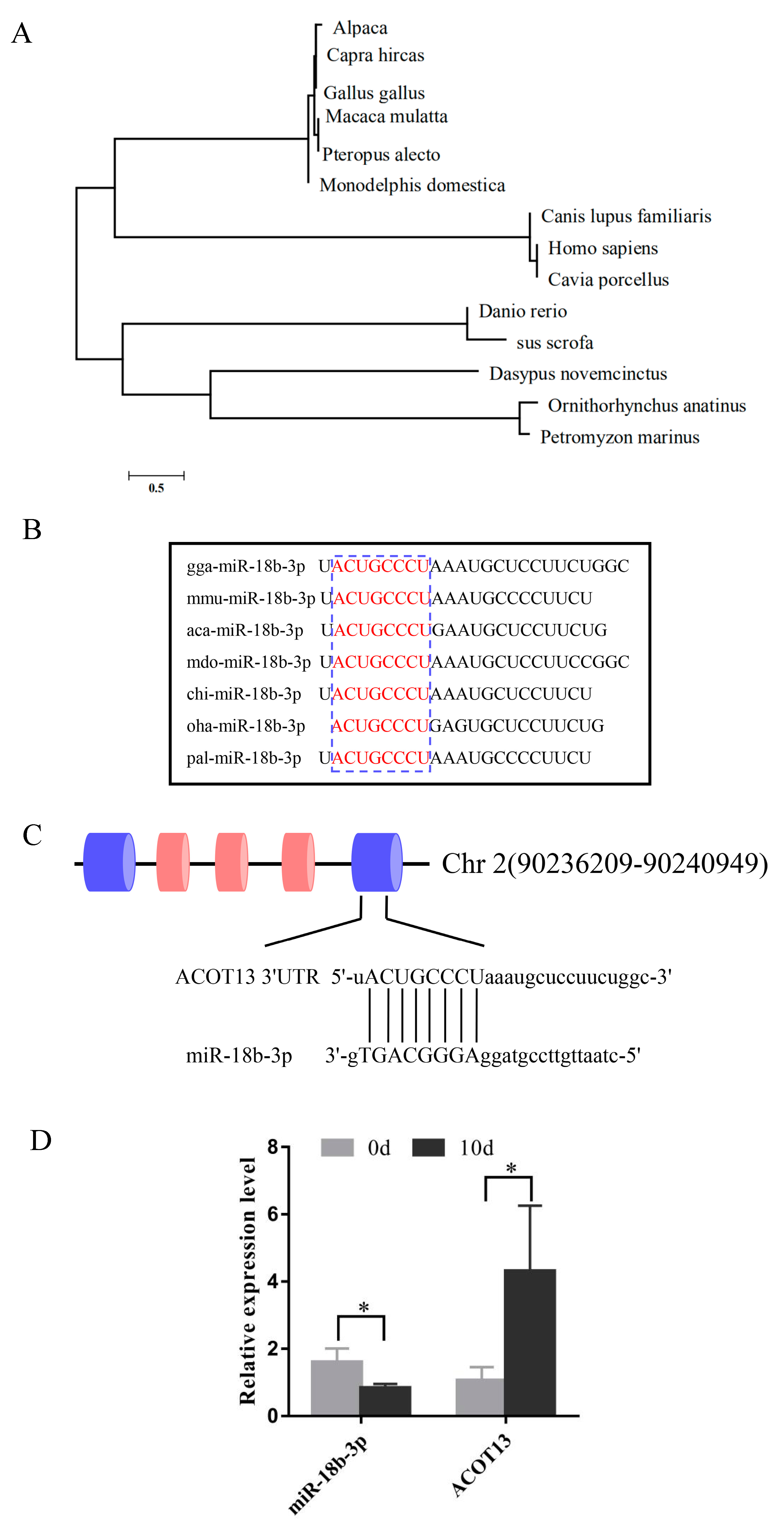
3.5. Expression of gga-miR-18b-3p and ACOT13 in Preadipocytes and Adipocytes
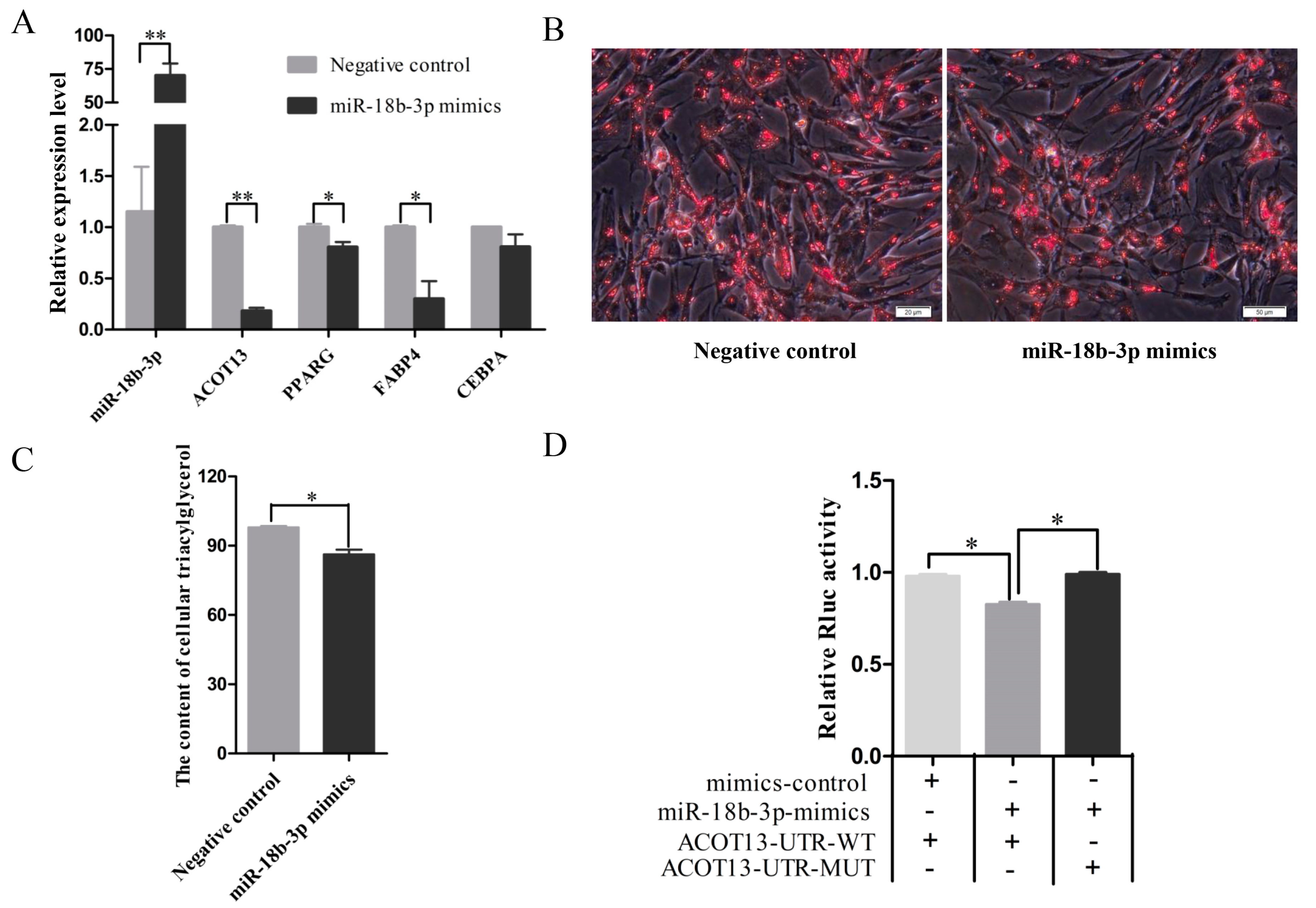
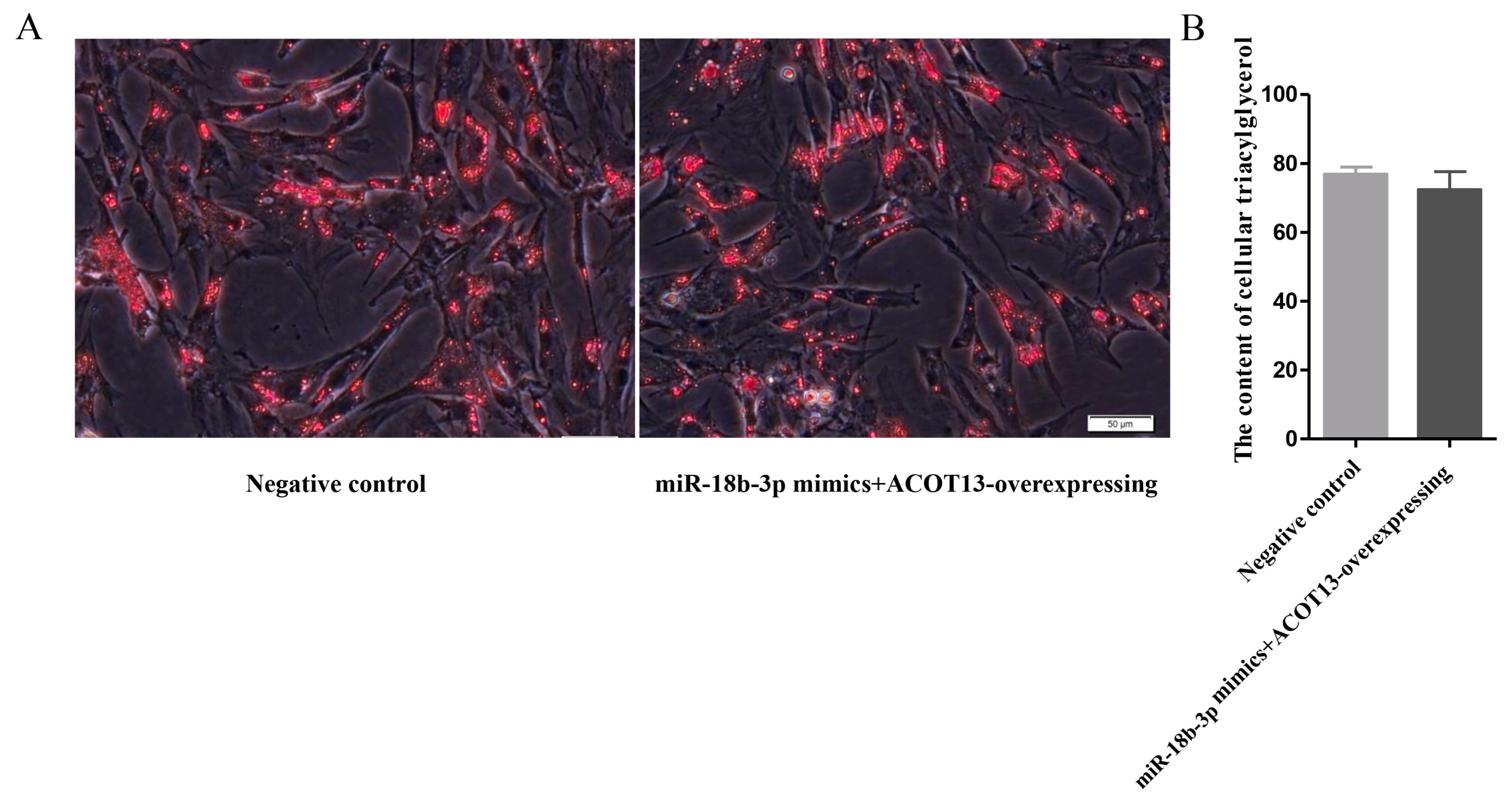
3.6. miR-18b-3p Functioned as an Inhibitor during Preadipocytes Differentiation by Targeting the ACOT13 Gene
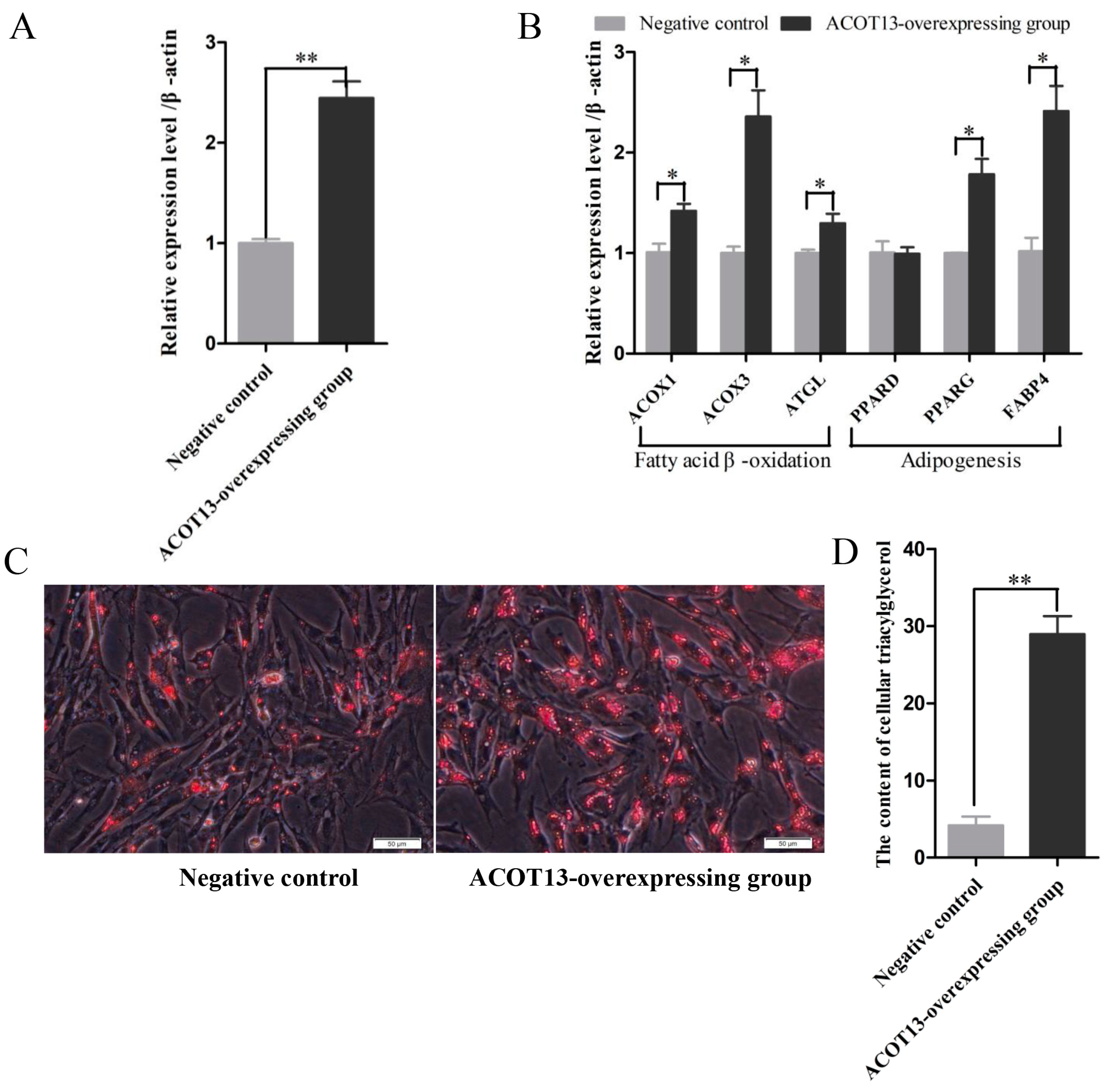
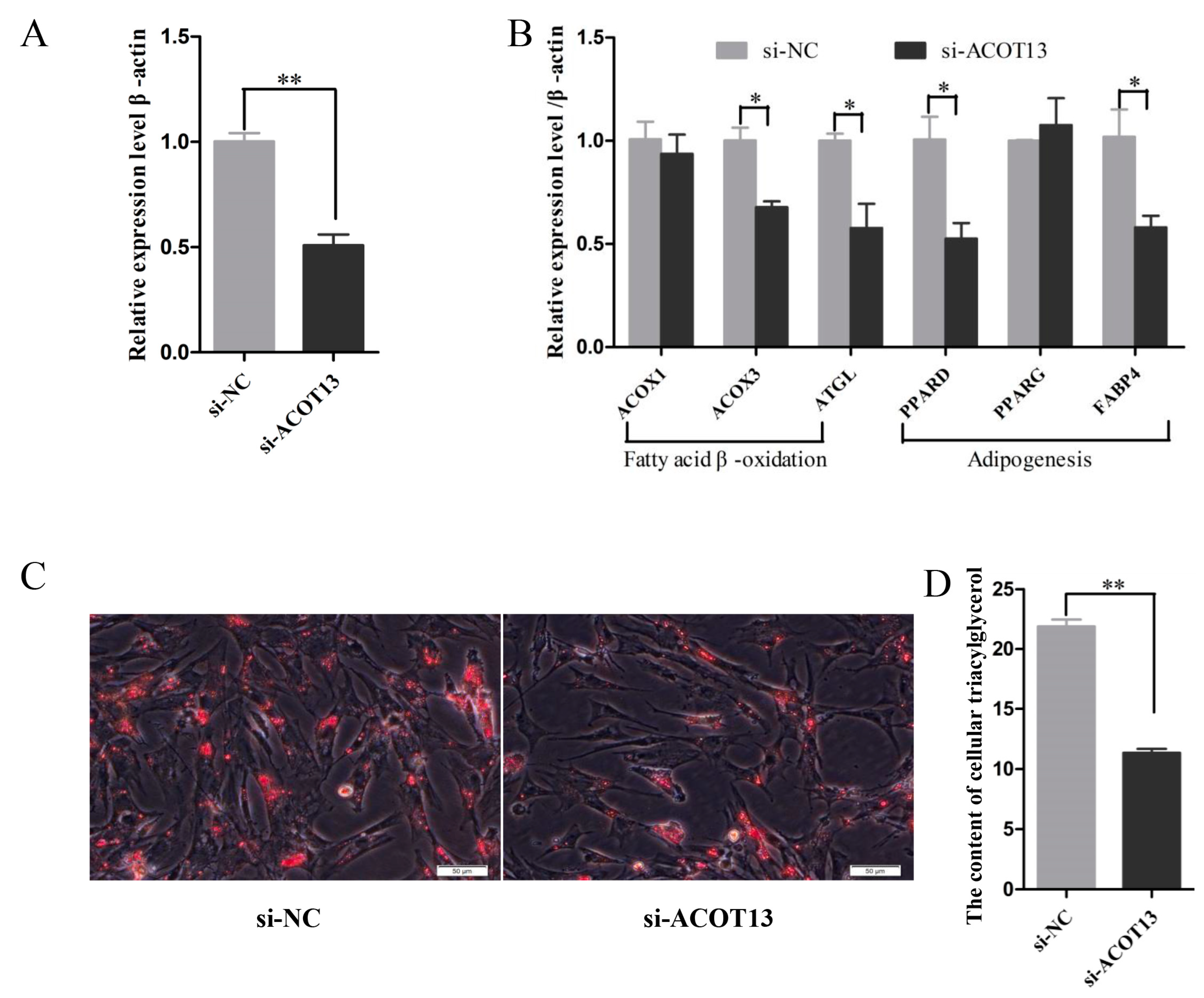
3.7. ACOT13 Promoted Preadipocyte Differentiation by Fatty Acid β-Oxidation and Lipogenic Transcription
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Accession Numbers
Abbreviations
| ACOX1 | acyl coenzyme A oxidase 1: palmitoyl |
| ACOX3 | acyl coenzyme A oxidase 3, palmitoyl |
| ATGL | adipose triglyceride lipase |
| PPARA | peroxisome proliferator-activated receptor α |
| PPARD | peroxisome proliferator-activated receptor β |
| CEBPA | CCAAT-enhancer binding protein-alpha |
| FABP4 | fatty acid binding protein 4 |
| ACOT13 | acyl-CoA thioesterase 13 |
| IMF | Intramuscular fat |
| PBS | phosphate buffered saline |
| TG | triacylglycerol |
| GO | gene ontology |
| KEGG | Kyoto encyclopedia of genes and genomes |
References
- Du, M.; Yin, J.; Zhu, M.J. Cellular signaling pathways regulating the initial stage of adipogenesis and marbling of skeletal muscle. Meat Sci. 2010, 86, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Massimiliano, P.; Claudio, C. Muscle growth and poultry meat quality issues. Nutrients 2012, 4, 1–12. [Google Scholar]
- Hocquette, J.F.; Gondret, F.; Baéza, E.; Médale, F.; Jurie, C.; Pethick, D.W. Intramuscular fat content in meat-producing animals: Development, genetic and nutritional control, and identification of putative markers. Animal 2010, 4, 303–319. [Google Scholar] [CrossRef]
- Zhang, M.; Li, D.-H.; Li, F.; Sun, J.-W.; Jiang, R.-R.; Li, Z.-J.; Han, R.-L.; Li, G.-X.; Liu, X.-J.; Kang, X.-T.; et al. Integrated Analysis of MiRNA and Genes Associated with Meat Quality Reveals that Gga-MiR-140-5p Affects Intramuscular Fat Deposition in Chickens. Cell. Physiol. Biochem. 2018, 46, 2421–2433. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Y.; Yuan, X.; Pan, R.; Yao, W.; Zhong, L.; Song, Q.; Zheng, S.; Wang, Z.; Xu, Q.; et al. Identification of differentially expressed microRNAs during preadipocyte differentiation in Chinese crested duck. Gene 2018, 661, 126–132. [Google Scholar] [CrossRef]
- Gan, L.; Yan, J.; Liu, Z.; Feng, M.; Sun, C. Adiponectin Prevents Reduction of Lipid-Induced Mitochondrial Biogenesis via AMPK/ACC2 Pathway in Chicken Adipocyte. J. Cell. Biochem. 2015, 116, 1090–1100. [Google Scholar] [CrossRef]
- Song, J.H.; Yang, J.; Pan, F.; Jin, B. Differential expression of microRNAs may regulate pollen development in Brassica oleracea. Genet. Mol. Res. 2015, 14, 15024–15034. [Google Scholar] [CrossRef] [PubMed]
- Fonseca-Sanchéz, M.A.; Pérez-Plasencia, C.; Fernández-Retana, J.; Arechaga-Ocampo, E.; Marchat, L.A.; Rodríguez-Cuevas, S.; Bautista-Piña, V.; Arellano-Anaya, Z.E.; Flores-Pérez, A.; Diaz-Chávez, J. MicroRNA-18b is upregulated in breast cancer and modulates genes involved in cell migration. Oncol. Rep. 2013, 30, 2399–2410. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.; Dong, M.J.; Wang, M.; Yin, B.Q.; Luo, H.; Lin-Yun, H.E.; Fang, H.U. The roles of miRNAs in the regulation of brown adipocyte function and white adipocyte browning. Chin. Bull. Life Sci. 2014, 26, 1051–1056. [Google Scholar]
- Chen, L.; Song, J.; Cui, J.; Hou, J.; Zheng, X.; Li, C.; Liu, L. MicroRNAs regulate adipocyte differentiation. Cell Boil. Int. 2013, 37, 533–546. [Google Scholar] [CrossRef]
- Esau, C.; Kang, X.; Peralta, E.; Hanson, E.; Marcusson, E.G.; Ravichandran, L.V.; Sun, Y.; Koo, S.; Perera, R.J.; Jain, R.; et al. MicroRNA-143 Regulates Adipocyte Differentiation. J. Boil. Chem. 2004, 279, 52361–52365. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Gao, Z.; Alarcon, R.M.; Ye, J.; Yun, Z. A role of miR-27 in the regulation of adipogenesis. Febs J. 2010, 276, 2348–2358. [Google Scholar] [CrossRef]
- Leivonen, S.K.; Mäkelä, R.; Östling, P.; Kohonen, P.; Haapapaananen, S.; Kleivi, K.; Enerly, E.; Aakula, A.; Hellström, K.; Sahlberg, N. Protein lysate microarray analysis to identify microRNAs regulating estrogen receptor signaling in breast cancer cell lines. Oncogene 2009, 28, 3926. [Google Scholar] [CrossRef] [PubMed]
- Morselli, E.; Santos, R.D.S.; Gao, S.; Y, Á.; Criollo, A.; Palmer, B.F.; Clegg, D.J. Impact of Estrogens and Estrogen Receptor Alpha (ESR1) in Brain Lipid Metabolism. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E7–E14. [Google Scholar] [CrossRef] [PubMed]
- Heine, P.A.; Taylor, J.A.; Iwamoto, G.A.; Lubahn, D.B.; Cooke, P.S. Increased adipose tissue in male and female estrogen receptor-alpha knockout mice. Proc. Natl. Acad. Sci. USA 2000, 97, 12729–12734. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Robertson, K.M.; Jones, M.E.E.; Simpson, E.R. Effect of estrogen deficiency in the male: the ArKO mouse model. Mol. Cell. Endocrinol. 2002, 193, 7–12. [Google Scholar] [CrossRef]
- Grodstein, F. Response to comments on ‘A Prospective, Observational Study of Postmenopausal Hormone Therapy and Primary Prevention of Cardiovascular Disease’. Maturitas 2001, 38, 239–241. [Google Scholar] [CrossRef]
- Willembergink, E.; Wallace, R.A.; Berg, J.A.V.D. Estrogen-Induced Synthesis of Yolk Proteins in Roosters. Am. Zool. 1974, 14, 1177–1193. [Google Scholar]
- Kirchgessner, T.G.; Heinzmann, C.; Svenson, K.L.; Gordon, D.A.; Nicosia, M.; Lebherz, H.G.; Lusis, A.J.; Williams, D.L. Regulation of chicken apolipoprotein B: Cloning, tissue distribution, and estrogen induction of mRNA. Gene 1987, 59, 241–251. [Google Scholar] [CrossRef]
- Schneider, W.J.; Carroll, R.; Severson, D.L.; Nimpf, J. Apolipoprotein VLDL-II inhibits lipolysis of triglyceride-rich lipoproteins in the laying hen. J. Lipid Res. 1990, 31, 507–513. [Google Scholar]
- Clegg, D.J. Minireview: The year in review of estrogen regulation of metabolism. Mol. Endocrinol. 1957, 26, 1957–1960. [Google Scholar] [CrossRef] [PubMed]
- Cooke, P.S.; Naaz, A. Role of estrogens in adipocyte development and function. Exp. Biol. Med. 2004, 229, 1127–1135. [Google Scholar] [CrossRef]
- Wu, J.-H.; Wang, Y.-H.; Wang, W.; Shen, W.; Sang, Y.-Z.; Liu, L.; Chen, C.-M. MiR-18b suppresses high-glucose-induced proliferation in HRECs by targeting IGF-1/IGF1R signaling pathways. Int. J. Biochem. Cell Boil. 2016, 73, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.J.; Wise, L.S.; Berkowitz, R. Insulin-like growth factor-I is an essential regulator of the differentiation of 3T3-L1 adipocytes. J. Biol. Chem. 1988, 263, 9402. [Google Scholar] [PubMed]
- Jin, S.; Zhai, B.; Qiu, Z.; Wu, J.; Lane, M.D.; Liao, K. c-Crk, a substrate of the insulin-like growth factor-1 receptor tyrosine kinase, functions as an early signal mediator in the adipocyte differentiation process. J. Biol. Chem. 2000, 275, 34344. [Google Scholar] [CrossRef] [PubMed]
- Hairong, H.; Xuemin, G.; Shangyu, H.; Manrong, J.; Xinyuan, L.; Kan, L. Lipid rafts/caveolae are essential for insulin-like growth factor-1 receptor signaling during 3T3-L1 preadipocyte differentiation induction. J. Boil. Chem. 2003, 278, 11561–11569. [Google Scholar]
- Langmead, B. Aligning short sequencing reads with Bowtie. Curr. Protoc. Bioinform. 2010, 32, 11.7.1–11.7.14. [Google Scholar]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68. [Google Scholar] [CrossRef]
- Moxon, S.; Schwach, F.; Dalmay, T.; Maclean, D.; Studholme, D.J.; Moulton, V. A toolkit for analysing large-scale plant small RNA datasets. Bioinformatics 2008, 24, 2252–2253. [Google Scholar] [CrossRef]
- Chang, T.-H.; Huang, H.-Y.; Hsu, J.B.-K.; Weng, S.-L.; Horng, J.-T.; Huang, H.-D. An enhanced computational platform for investigating the roles of regulatory RNA and for identifying functional RNA motifs. BMC Bioinform. 2013, 14, S4. [Google Scholar]
- Kohl, M.; Wiese, S.; Warscheid, B. Cytoscape: Software for visualization and analysis of biological networks. Methods Mol. Biol. 2011, 696, 291–303. [Google Scholar] [PubMed]
- Li, F.; Li, D.; Zhang, M.; Sun, J.; Li, W.; Jiang, R.; Han, R.; Wang, Y.; Tian, Y.; Kang, X. miRNA-223 targets the GPAM gene and regulates the differentiation of intramuscular adipocytes. Gene 2019, 685, 106–113. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Stocks, M.B.; Moxon, S.; Mapleson, D.; Woolfenden, H.C.; Mohorianu, I.; Folkes, L.; Schwach, F.; Dalmay, T.; Moulton, V. The UEA sRNA workbench: A suite of tools for analysing and visualizing next generation sequencing microRNA and small RNA datasets. Bioinformatics 2012, 28, 2059–2061. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Kanao, Y.; Yamanaka, M.; Sakuraba, H.; Mizutani, Y.; Igarashi, Y.; Kihara, A. Characterization of four mammalian 3-hydroxyacyl-CoA dehydratases involved in very long-chain fatty acid synthesis. FEBS Lett. 2008, 582, 2435–2440. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.A.; Hammer, R.E.; Horton, J.D. Deletion of ELOVL5 leads to fatty liver through activation of SREBP-1c in mice. J. Lipid Res. 2009, 50, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.W.; Niepel, M.W.; Han, S.; Kawano, Y.; Cohen, D.E. Thioesterase superfamily member 2/acyl-CoA thioesterase 13 (Them2/Acot13) regulates hepatic lipid and glucose metabolism. FASEB J. 2012, 26, 2209–2221. [Google Scholar] [CrossRef]
- Zhang, B.W.; Cai, H.F.; Wei, X.F.; Sun, J.J.; Lan, X.Y.; Lei, C.Z.; Lin, F.P.; Qi, X.L.; Martin, P.; Hong, C. miR-30-5p Regulates Muscle Differentiation and Alternative Splicing of Muscle-Related Genes by Targeting MBNL. Int. J. Mol. Sci. 2016, 17, 182. [Google Scholar] [CrossRef]
- Roos, J.; Enlund, E.; Funcke, J.B.; Tews, D.; Holzmann, K.; Debatin, K.M.; Wabitsch, M.; Fischerposovszky, P. miR-146a-mediated suppression of the inflammatory response in human adipocytes. Sci. Rep. 2016, 6, 38339. [Google Scholar] [CrossRef]
- Li, H.; Xue, M.; Xu, J.; Qin, X. MiR-301a is involved in adipocyte dysfunction during obesity-related inflammation via suppression of PPARγ. Die Pharm. 2016, 71, 84. [Google Scholar]
- Kersten, S.; Desvergne, B.; Wahli, W. Roles of PPARs in health and disease. Nature 2000, 405, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-N.; Choi, H.Y.; Lee, W.; Park, G.M.; Shin, W.S.; Kim, Y.K. Sargaquinoic acid and sargahydroquinoic acid from Sargassum yezoense stimulate adipocyte differentiation through PPARα/γ activation in 3T3-L1 cells. FEBS Lett. 2008, 582, 3465–3472. [Google Scholar] [CrossRef] [PubMed]
- Bai, N.; He, K.; Ibarra, A.; Bily, A.; Roller, M.; Chen, X.; Rühl, R. Iridoids from Fraxinus excelsior with Adipocyte Differentiation-Inhibitory and PPARα Activation Activity. J. Nat. Prod. 2015, 73, 2. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Hashizaki, H.; Goto, T.; Sakamoto, T.; Takahashi, N.; Kawada, T. Activation of peroxisome proliferator-activated receptor-α enhances fatty acid oxidation in human adipocytes. Biochem. Biophys. Res. Commun. 2011, 407, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Lee, J.O.; Kim, J.H.; Kim, N.; You, G.Y.; Moon, J.W.; Sha, J.; Kim, S.J.; Lee, Y.W.; Kang, H.J.; et al. Coenzyme Q10 increases the fatty acid oxidation through AMPK-mediated PPARα induction in 3T3-L1 preadipocytes. Cell Signal. 2012, 24, 2329–2336. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, Y.; Xu, Q.; Yuan, X.; Dai, W.; Shen, X.; Wang, Z.; Chang, G.; Wang, Z.; Chen, G. The differentiation of preadipocytes and gene expression related to adipogenesis in ducks (Anas platyrhynchos). PLoS ONE 2018, 13, e0196371. [Google Scholar] [CrossRef] [PubMed]
- Starčević, K.; Filipović, N.; Šperanda, M.; Đidara, M.; Mašek, T. The influence of sex and gonadectomy on hepatic and brain fatty acid composition, lipogenesis and β-oxidation. J. Anim. Physiol. Anim. Nutr. 2017, 101, 649–657. [Google Scholar] [CrossRef]
- Wang, B.; Wang, H.; Xiang, M.A.; Guang, P.U.; Niu, P.; Zhang, Z.; Zhou, J.; Pinghua, L.I.; Huang, R. Analysis of intramuscular fat content in Suhuai pigs and its correlation with polymorphisms of IGF2 and SCD genes. Anim. Husb. Vet. Med. 2018, 50, 20–25. [Google Scholar]
- Shi, H.B.; Du, Y.; Zhang, C.H.; Sun, C.; He, Y.L.; Wu, Y.H.; Liu, J.X.; Luo, J.; Loor, J.J. Fatty acid elongase 5 (ELOVL5) alters the synthesis of long-chain unsaturated fatty acids in goat mammary epithelial cells. J. Dairy Sci. 2018, 101, 4586–4594. [Google Scholar] [CrossRef]
- Murakami, Y.; Tamori, A.; Itami, S.; Tanahashi, T.; Toyoda, H.; Tanaka, M.; Wu, W.; Brojigin, N.; Kaneoka, Y.; Maeda, A.; et al. The expression level of miR-18b in hepatocellular carcinoma is associated with the grade of malignancy and prognosis. BMC Cancer 2013, 13, 99. [Google Scholar] [CrossRef]
- Namløs, H.M.; Meza-Zepeda, L.A.; Tale, B.Y.; ?Stensen, I.H.G.; Kresse, S.H.; Kuijjer, M.L.; Massimo, S.; Horst, B.; Anne-Marie, C.J.; Ola, M. Modulation of the osteosarcoma expression phenotype by microRNAs. PLoS ONE 2012, 7, e48086. [Google Scholar] [CrossRef] [PubMed]
- Cantu, D.C.; Ardèvol, A.; Rovira, C.; Reilly, P.J. Molecular Mechanism of a Hotdog-Fold Acyl-CoA Thioesterase. Chem. A Eur. J. 2014, 20, 9045–9051. [Google Scholar] [CrossRef] [PubMed]
- Kawano, Y.; Ersoy, B.A.; Li, Y.; Nishiumi, S.; Yoshida, M.; Cohen, D.E. Thioesterase Superfamily Member 2 (Them2) and Phosphatidylcholine Transfer Protein (PC-TP) Interact To Promote Fatty Acid Oxidation and Control Glucose Utilization. Mol. Cell. Boil. 2014, 34, 2396–2408. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.W.; Ozdemir, C.; Kawano, Y.; LeClair, K.B.; Vernochet, C.; Kahn, C.R.; Hagen, S.J.; Cohen, D.E. Thioesterase Superfamily Member 2/Acyl-CoA Thioesterase 13 (Them2/Acot13) Regulates Adaptive Thermogenesis in Mice. J. Boil. Chem. 2013, 288, 33376–33386. [Google Scholar] [CrossRef] [PubMed]
- Li, L.O.; Klett, E.L.; Coleman, R.A. Acyl-CoA synthesis, lipid metabolism and lipotoxicity. Biochim. Biophys. Acta (BBA) Mol. Cell Boil. Lipids 2010, 1801, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.M.; Lei, O.; Li Pei-Chi, W.; Koves, T.R.; Olga, I.; Stevens, R.D.; Watkins, S.M.; Muoio, D.M.; Coleman, R.A. Adipose acyl-CoA synthetase-1 directs fatty acids toward beta-oxidation and is required for cold thermogenesis. Cell Metab. 2010, 12, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Hung, J.-Y.; Chiang, S.-R.; Liu, K.-T.; Tsai, M.-J.; Huang, M.-S.; Shieh, J.-M.; Yen, M.-C.; Hsu, Y.-L. Overexpression and proliferation dependence of acyl-CoA thioesterase 11 and 13 in lung adenocarcinoma. Oncol. Lett. 2017, 14, 3647–3656. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, H.T.; Hornick, J.L.; Cohen, D.E. Phosphatidylcholine transfer protein/StarD2 promotes microvesicular steatosis and liver injury in murine experimental steatohepatitis. Am. J. Physiol. Liver Physiol. 2017, 313, G50–G61. [Google Scholar] [CrossRef]
- Bushati, N.; Cohen, S.M. microRNA Functions. Annu. Rev. Cell Dev. Biol. 2007, 23, 175–205. [Google Scholar] [CrossRef]
- Wienholds, E.; Plasterk, R.H.A. MicroRNA function in animal development. FEBS Lett. 2005, 579, 5911–5922. [Google Scholar] [CrossRef]
- Tripathi, G.; Salih, D.A.M.; Drozd, A.C.; Cosgrove, R.A.; Cobb, L.J.; Pell, J.M. IGF-independent effects of insulin-like growth factor binding protein-5 (Igfbp5) in vivo. FASEB J. 2009, 23, 2616. [Google Scholar] [CrossRef] [PubMed]
- Cave, M.C.; Clair, H.B.; Hardesty, J.E.; Falkner, K.C.; Feng, W.; Clark, B.J.; Sidey, J.; Shi, H.; Aqel, B.A.; Mcclain, C.J.; et al. Nuclear receptors and nonalcoholic fatty liver disease. Biochim. Biophys. Acta 2016, 1859, 1083–1099. [Google Scholar] [CrossRef] [PubMed]
- Lollar, P.; Parker, E.T.; Fay, P.J. Coagulant properties of hybrid human/porcine factor VIII molecules. J. Boil. Chem. 1992, 267, 23652. [Google Scholar]
- Petersen, B.; Carnwath, J.W.; Niemann, H. The perspectives for porcine-to-human xenografts. Comp. Immunol. Microbiol. Infect. Dis. 2009, 32, 91–105. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Acc. # | Forward Primer Sequence 5’-3′ | Reverse Primer Sequence 5′-3′ |
|---|---|---|---|
| ACOT13 | XM_015282414.2 | CTTTGGAACAGCAGCACAGTT | TTACAGGAACCCTTCACTGCC |
| β-actin | NM_205518.1 | CAGCCAGCCATGGATGATGA | ACCAACCATCACACCCTGAT |
| siRNA | XM_015282414.2 | GCACGUUACACGGAGGUUUTTAAACCUCCGUGUAACGUGCTT | |
| ACOX1 | NM_001006205.1 | AAGGAGATCGAGGCCTTAGTG | GCCGTCCACGATGAACAAAG |
| ACOX3 | XM_420814.6 | AAAGAAGACAGTGGCAACCG | TCACCATCGAGAAACACTGC |
| ATGL | NM_001113291.1 | CACTGCCATGATGGTCCCCTA | CCACAAGGAGATGCTGAAGAA |
| PPARA | XM_015289937.2 | AGTAAGCTCTCAGAAACTTTGTTG | GTCATTTCACTTCACGCAGCA |
| PPARD | XM_015298893.2 | GCAAGCCCTTCAACGAGATCA | GGGACGATCTCCACACAGG |
| CEBPA | NM_001031459.1 | TTCTACGAGGTCGATTCCCG | AGCCTCTCTGTAGCCGTAG |
| FABP4 | NM_204290.1 | ATGTGCGACCAGTTTGT | TCACCATTGATGCTGATAG |
| PPARG | XM_015292931.2 | GTGCAATCAAAATGGAGCC | CTTACAACCTTCACATGCAT |
| ACOT13(CDS) | XM_015282414.2 | CCCAAGCTTGCCACCATGGGCAGCATGCGCTTCAC | CCGGAATTCCTGTCCTATGTACTTTGTAT |
| WT-ACOT13 | ENSGALT00000086884.1 | CCGCTCGAGAAGCAACTCACCCTTCAGGC | ATTTGCGGCCGCAGCTTGCTTCACACTTCCCAT |
| MUT-ACOT13 | ENSGALT00000086884.1 | CTTACTAATTGTTCCGTAGGTCCCGTCAGAAGGGTTCCTGTAAGCAAC | GTTGCTTACAGGAACCCTTCTGACGGGACCTACGGAACAATTAGTAAG |
| Pathway ID | Pathway Term | Target Gene List |
|---|---|---|
| gga00062 | Fatty acid elongation | ELOVL5, HACD3, HADHA, ACAA2, ELOVL4, HACD1, ECHS1, ELOVL1, HSD17B12, MECR, PPT1, ACOT7, HACD2, ELOVL6 |
| gga00020 | Citrate cycle | DLD, IDH3B, IDH3A, SDHA, ACLY, SUCLG2, SUCLG1, ACO2, MDH1, IDH1, ACO1, OGDH, SDHB, IDH3G, IDH2, CS |
| gga01212 | Fatty acid metabolism | ELOVL5, HACD3, ACADS, EHHADH, HADHA, ACAA2, SCD5, FASN, ACSBG2, ACOX1, HACD1, ACSBG1, ECHS1, SCD, HSD17B12, MECR, PPT1, ACSL1, PECR, CPT2, HACD2, |
| ACADSB, ACADL, ELOVL6 | ||
| gga00071 | Fatty acid degradation | ACADS, EHHADH, HADHA, ACAA2, ACSBG2, ACOX1, ECI1, ACSBG1, ECHS1, ACSL1, CPT2, ALDH7A1, ECI2, ACADSB, ACADL |
| gga04010 | MAPK (mitogen-activated protein kinase) signaling pathway | RELA, CHUK, TAOK3, BDNF, EGFR, TNFSF6, PAK1, ChALK5, TAB1, TGFB3, ARR3, RAC2, MAP2K1, NF1, FOS, MAP3K13, PRKCA, CRKL, DUSP6, SRF, CACNG5, GADD45A, FGFR4, MAP3K14, HSPA2, NRAS, cRac1B, PDGF-A,CACNA1D, CASP3, CDC42, MAPK10, STK4, PTPN7, TGFBR2, RPS6KA5, FGF1, DUSP4, GNG12, DUSP7, MAP3K1, NR4A1, HSPA8, LAMTOR3, PPP3CB, TAOK1, MRAS, CACNA1S, MAP2K5, MAPK11, PPP3R1, NFATC3, DUSP10, CACNA1G, RPS6KA, DUSP3, cRac1A, FLNB, CACNA2D1, H-RAS, GADD45B, PPM1B, MAPKAPK3, CACNA1B, TNFRSF1A, BRAF, FGFR1, NFKB2, FGFR3, MAP3K5, CACNG4, FGF8, MAPK9, CACNG3, MAP2K3 |
| gga04150 | mTOR (mammalian target of rapamycin) signaling pathway | CHUK, ATP6V1G1, PDPK1, SLC7A5, STRADA, RNF152, PIK3R5, FZD5, MAP2K1, CAB39L, RPS6KB1, WDR24, SKP2,PRKCA, ATP6V1G3, NPRL2, WNT6, STK11, FZD6, FZD9, SLC38A9, ATP6V1B2, RPS6, FZD4, NRAS, WDR59, MLST8, Wnt8c, RRAGC, PIK3CB, PIK3CD, RRAGD, LAMTOR3, FZD3, ATP6V1A, RPTOR, ULK3, ATP6V1D, CLIP1, WNT11, RPS6KA, PRKAA1,HRAS, EIF4E2, PRKAA2, TNFRSF1A, BRAF, RHOA, TBC1D |
| gga00061 | Fatty acid biosynthesis | FASN, ACSBG2, ACSBG1, ACSL1, ACOT13 |
| gga03320 | PPAR (peroxisome proliferator-activated receptors) signaling pathway | FABP5, PDPK1, EHHADH, FABP3, LXR, PLIN2, SCD5, ACSBG2, LPL, ACOX1, SLC27A2, ACSBG1, SCD, ACSL1, CPT2, CYP8B1, PLIN1, ACADL |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, G.; Li, F.; Ma, X.; Sun, J.; Jiang, R.; Tian, Y.; Han, R.; Li, G.; Wang, Y.; Li, Z.; et al. gga-miRNA-18b-3p Inhibits Intramuscular Adipocytes Differentiation in Chicken by Targeting the ACOT13 Gene. Cells 2019, 8, 556. https://doi.org/10.3390/cells8060556
Sun G, Li F, Ma X, Sun J, Jiang R, Tian Y, Han R, Li G, Wang Y, Li Z, et al. gga-miRNA-18b-3p Inhibits Intramuscular Adipocytes Differentiation in Chicken by Targeting the ACOT13 Gene. Cells. 2019; 8(6):556. https://doi.org/10.3390/cells8060556
Chicago/Turabian StyleSun, Guirong, Fang Li, Xiangfei Ma, Junwei Sun, Ruirui Jiang, Yadong Tian, Ruili Han, Guoxi Li, Yanbin Wang, Zhuanjian Li, and et al. 2019. "gga-miRNA-18b-3p Inhibits Intramuscular Adipocytes Differentiation in Chicken by Targeting the ACOT13 Gene" Cells 8, no. 6: 556. https://doi.org/10.3390/cells8060556
APA StyleSun, G., Li, F., Ma, X., Sun, J., Jiang, R., Tian, Y., Han, R., Li, G., Wang, Y., Li, Z., Kang, X., & Li, W. (2019). gga-miRNA-18b-3p Inhibits Intramuscular Adipocytes Differentiation in Chicken by Targeting the ACOT13 Gene. Cells, 8(6), 556. https://doi.org/10.3390/cells8060556