Characterisation of Osteopontin in an In Vitro Model of Embryo Implantation

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Immunoprecipitation
2.3. SDS PAGE and Western Blotting
2.4. Mouse Embryo Collection
2.5. In Vitro Implantation Assay
2.6. Immunofluorescence Staining
2.7. Fluorescence Microscopy
2.8. Blastocyst RNA Extraction and Quantitative PCR
2.9. Statistical Analysis
3. Results
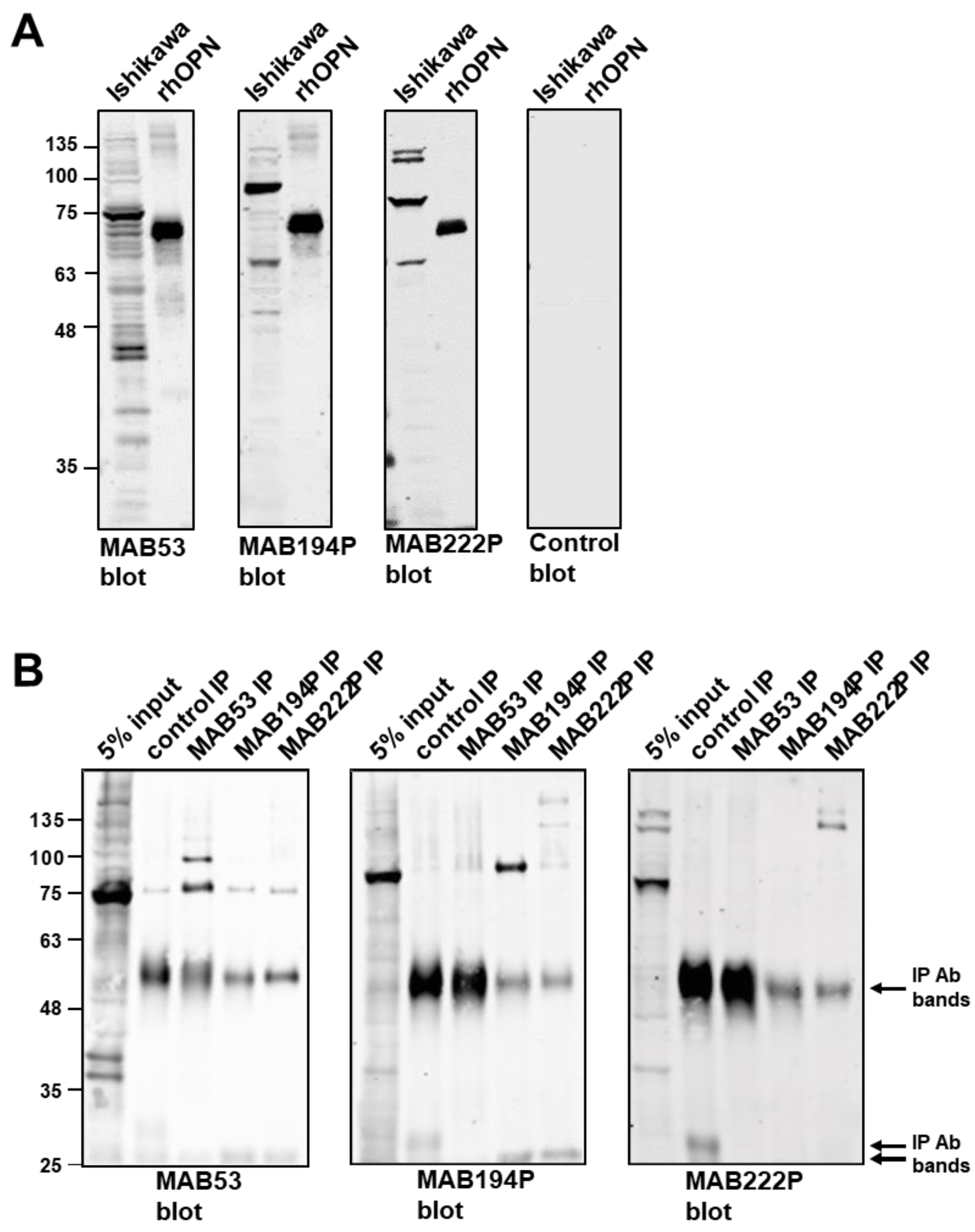
3.1. Biochemical Characterisation of OPN from Ishikawa Endometrial Epithelial Cells using Three Different Antibodies
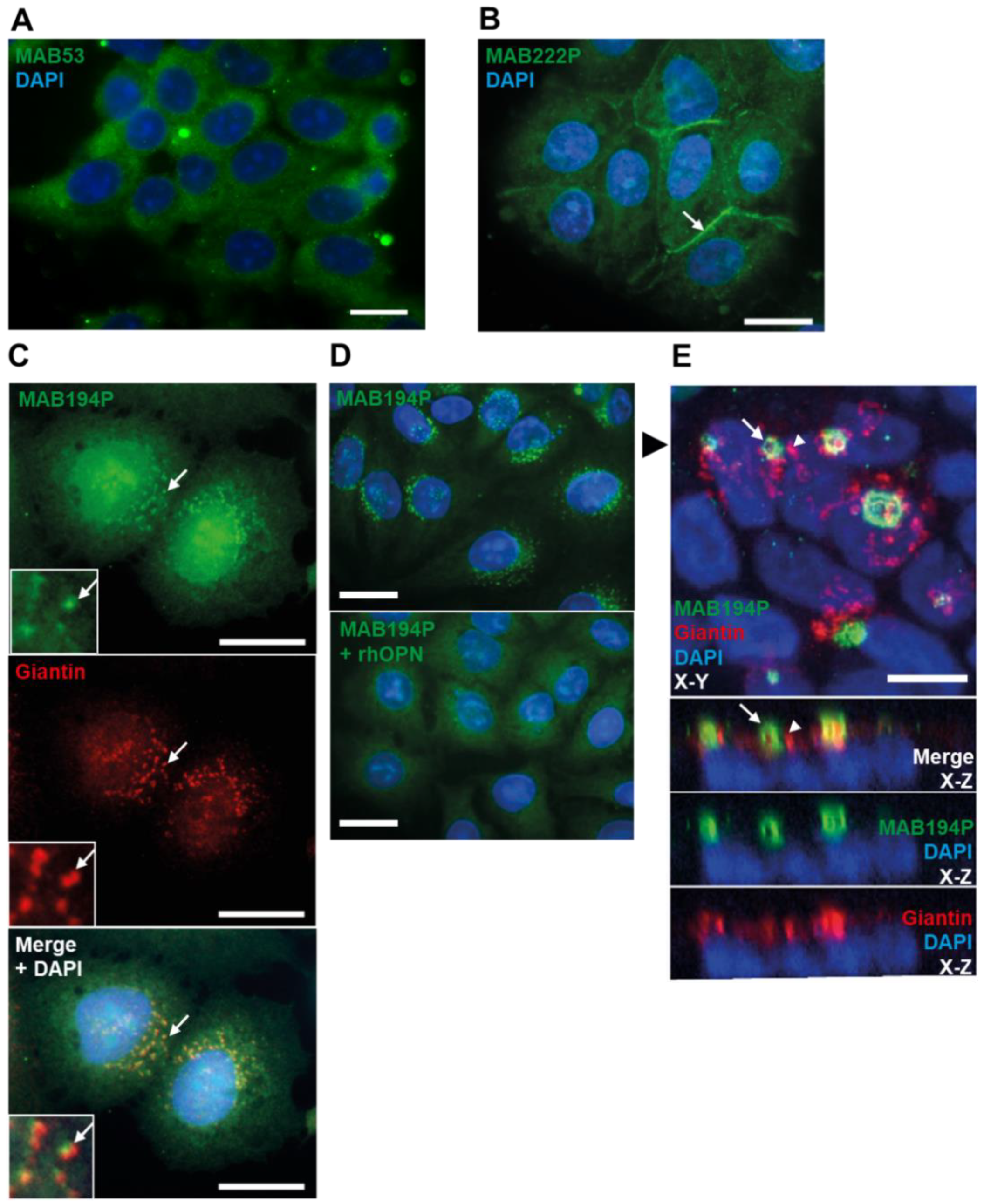
3.2. Distinct OPN Localisation in Ishikawa Cells Revealed by Three Different Antibodies
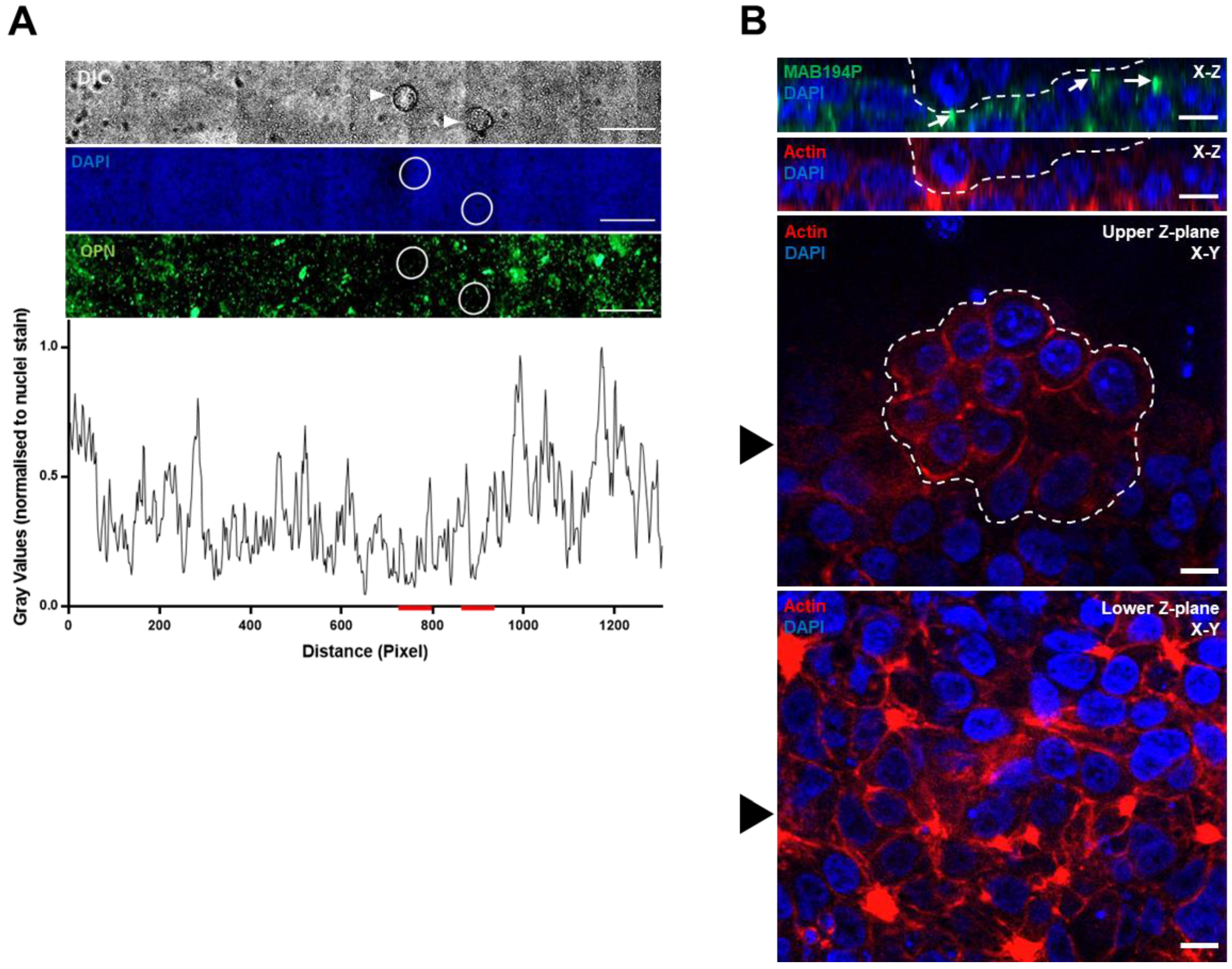
3.3. Ishikawa OPN Localisation is not Changed during Interaction with Mouse Blastocysts
3.4. Exogenous OPN Inhibits Mouse Blastocyst Invasion of Ishikawa Cells
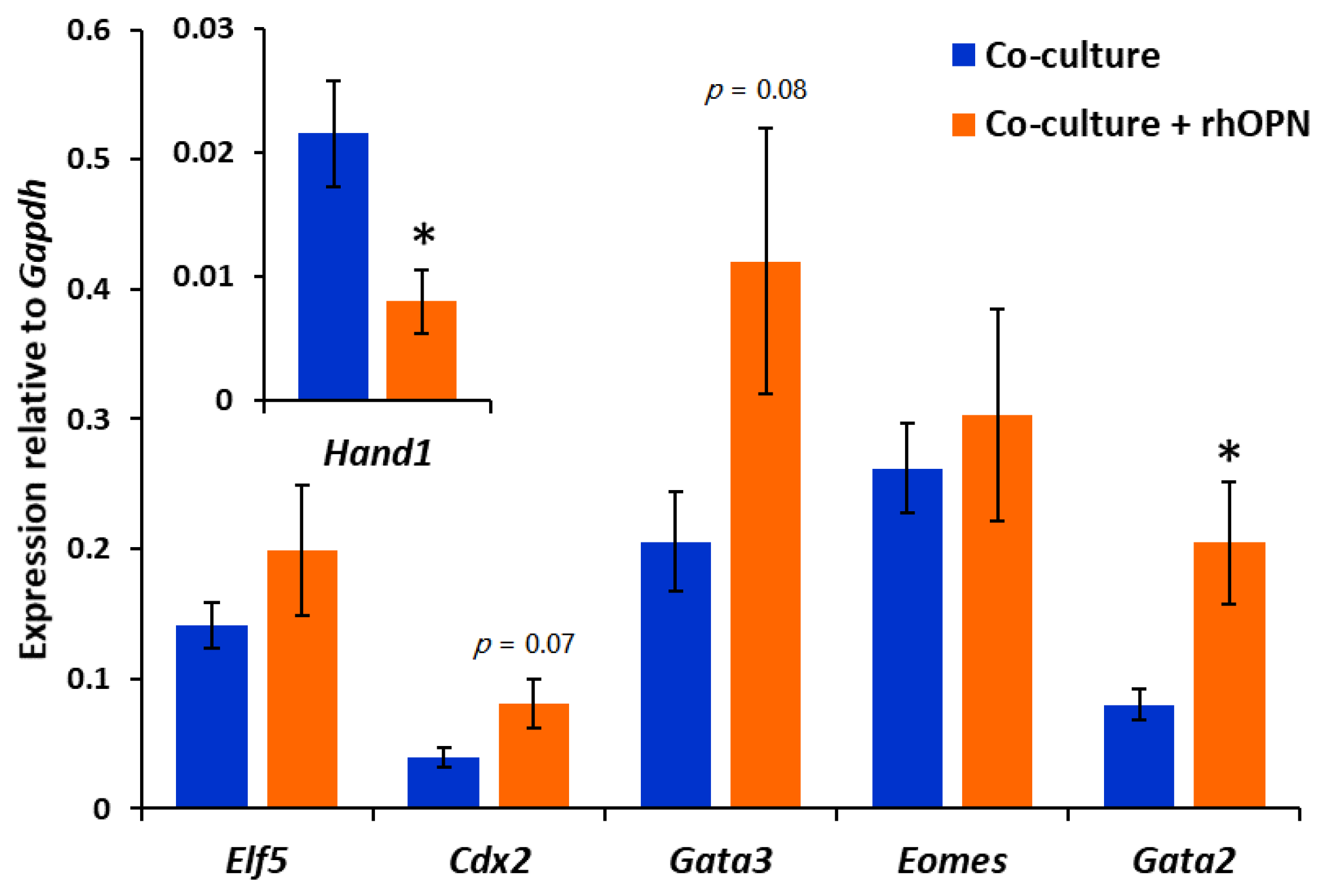
3.5. Exogenous OPN Regulates Mouse Blastocyst Gene Expression during Apposition with Ishikawa Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Koot, Y.E.; Boomsma, C.M.; Eijkemans, M.J.; Lentjes, E.G.; Macklon, N.S. Recurrent pre-clinical pregnancy loss is unlikely to be a ‘cause’ of unexplained infertility. Hum. Reprod. 2011, 26, 2636–2641. [Google Scholar] [CrossRef]
- De Geyter, C.; Calhaz-Jorge, C.; Kupka, M.S.; Wyns, C.; Mocanu, E.; Motrenko, T.; Scaravelli, G.; Smeenk, J.; Vidakovic, S.; Goossens, V. ART in Europe, 2014: results generated from European registries by ESHRE: The European IVF-monitoring Consortium (EIM) for the European Society of Human Reproduction and Embryology (ESHRE). Hum. Reprod. 2018, 33, 1586–1601. [Google Scholar] [CrossRef] [PubMed]
- Vasquez, Y.; DeMayo, F. Role of nuclear receptors in blastocyst implantation. Semin. Cell Dev. Biol. 2013, 24, 724–735. [Google Scholar] [CrossRef] [PubMed]
- Aplin, J.D.; Ruane, P.T. Embryo–epithelium interactions during implantation at a glance. J. Cell Sci. 2017, 130, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Craciunas, L.; Gallos, I.; Chu, J.; Bourne, T.; Quenby, S.; Brosens, J.J.; Coomarasamy, A. Conventional and modern markers of endometrial receptivity: a systematic review and meta-analysis. Hum. Reprod. Updat. 2019, 25, 202–223. [Google Scholar] [CrossRef]
- Kao, L.C.; Tulac, S.; Lobo, S.; Imani, B.; Yang, J.P.; Germeyer, A.; Osteen, K.; Taylor, R.N.; Lessey, B.A.; Giudice, L.C. Global Gene Profiling in Human Endometrium during the Window of Implantation. Endocrinology 2002, 143, 2119–2138. [Google Scholar] [CrossRef]
- Garlow, J.E.; Ka, H.; Johnson, G.A.; Burghardt, R.C.; Jaeger, L.A.; Bazer, F.W. Analysis of Osteopontin at the Maternal-Placental Interface in Pigs1. Boil. Reprod. 2002, 66, 718–725. [Google Scholar] [CrossRef]
- Apparao, K.; Illera, M.J.; Beyler, S.A.; Olson, G.E.; Osteen, K.G.; Corjay, M.H.; Boggess, K.; Lessey, B.A. Regulated Expression of Osteopontin in the Peri-Implantation Rabbit Uterus1. Boil. Reprod. 2003, 68, 1484–1490. [Google Scholar] [CrossRef] [PubMed]
- Nomura, S.; Wills, A.; Edwards, D.; Heath, J.; Hogan, B. Developmental expression of 2ar (osteopontin) and SPARC (osteonectin) RNA as revealed by in situ hybridization. J. Cell Boil. 1988, 106, 441–450. [Google Scholar] [CrossRef]
- Johnson, G.A.; Burghardt, R.C.; Spencer, T.E.; Newton, G.R.; Ott, T.L.; Bazer, F.W. Ovine osteopontin: II. Osteopontin and alpha(v)beta(3) integrin expression in the uterus and conceptus during the periimplantation period. Boil. Reprod. 1999, 61, 892–899. [Google Scholar] [CrossRef]
- Griffith, O.W.; Chavan, A.R.; Protopapas, S.; Maziarz, J.; Romero, R.; Wagner, G.P. Embryo implantation evolved from an ancestral inflammatory attachment reaction. Proc. Natl. Acad. Sci. USA 2017, 114, E6566–E6575. [Google Scholar] [CrossRef] [PubMed]
- Quenby, S.; Anim-Somuah, M.; Kalumbi, C.; Farquharson, R.; Aplin, J.D. Different types of recurrent miscarriage are associated with varying patterns of adhesion molecule expression in endometrium. Reprod. Biomed. Online 2007, 14, 224–234. [Google Scholar] [CrossRef]
- Apparao, K.B.; Murray, M.J.; Fritz, M.A.; Meyer, W.R.; Chambers, A.F.; Truong, P.R.; Lessey, B.A. Osteopontin and its receptor alphavbeta(3) integrin are coexpressed in the human endometrium during the menstrual cycle but regulated differentially. J. Clin. Endocrinol. Metab. 2001, 86, 4991–5000. [Google Scholar] [PubMed]
- Carson, D.D.; Lagow, E.; Thathiah, A.; Al-Shami, R.; Vernon, M.; Yuan, L.; Fritz, M.A.; Lessey, B.; Farach-Carson, M.C. Changes in gene expression during the early to mid-luteal (receptive phase) transition in human endometrium detected by high-density microarray screening. Mol. Hum. Reprod. 2002, 8, 871–879. [Google Scholar] [CrossRef]
- Johnson, G.A.; Burghardt, R.C.; Bazer, F.W.; Spencer, T.E. Osteopontin: Roles in Implantation and Placentation1. Boil. Reprod. 2003, 69, 1458–1471. [Google Scholar] [CrossRef]
- Castello, L.M.; Raineri, D.; Salmi, L.; Clemente, N.; Vaschetto, R.; Quaglia, M.; Garzaro, M.; Gentilli, S.; Navalesi, P.; Cantaluppi, V.; et al. Osteopontin at the Crossroads of Inflammation and Tumor Progression. Mediat. Inflamm. 2017, 2017, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Kazanecki, C.C.; Uzwiak, D.J.; Denhardt, D.T. Control of osteopontin signaling and function by post-translational phosphorylation and protein folding. J. Cell. Biochem. 2007, 102, 912–924. [Google Scholar] [CrossRef] [PubMed]
- Takada, Y.; Takada, Y.K.; Fujita, M. Crosstalk between insulin-like growth factor (IGF) receptor and integrins through direct integrin binding to IGF1. Cytokine Growth Factor Rev. 2017, 34, 67–72. [Google Scholar] [CrossRef]
- Carson, D.D.; Bagchi, I.; Dey, S.K.; Enders, A.C.; Fazleabas, A.T.; Lessey, B.A.; Yoshinaga, K. Embryo implantation. Dev. Biol. 2000, 223, 217–237. [Google Scholar] [CrossRef]
- Frank, J.W.; Seo, H.; Burghardt, R.C.; Bayless, K.J.; Johnson, G.A. ITGAV (alpha v integrins) bind SPP1 (osteopontin) to support trophoblast cell adhesion. Reproduction 2017, 153, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, H.L.; Labrosse, J.M.; McIntosh, F.A.; Mäenpää, P.H.; Kaartinen, M.T.; McKee, M.D. Homotypic Interactions of Soluble and Immobilized Osteopontin. Ann. Biomed. Eng. 2002, 30, 840–850. [Google Scholar] [CrossRef]
- Johnson, G.A.; Burghardt, R.C.; Bazer, F.W. Osteopontin: a leading candidate adhesion molecule for implantation in pigs and sheep. J. Sci. Biotechnol. 2014, 5, 56. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Zhou, C.; Chen, Y.; Zhao, J. The involvement of osteopontin and beta3 integrin in implantation and endometrial receptivity in an early mouse pregnancy model. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 170, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.J.; Forbes, K.; Carver, J.; Aplin, J.D. The role of the osteopontin-integrin alphavbeta3 interaction at implantation: functional analysis using three different in vitro models. Hum. Reprod. 2014, 29, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Chaen, T.; Konno, T.; Egashira, M.; Bai, R.; Nomura, N.; Nomura, S.; Hirota, Y.; Sakurai, T.; Imakawa, K. Estrogen-Dependent Uterine Secretion of Osteopontin Activates Blastocyst Adhesion Competence. PLOS ONE 2012, 7, e48933. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.-Z.; Qi, Q.-R.; Chen, Y.-X.; Xu, W.-M.; Liu, Q.; Yang, J. Uterine Micro-Environment and Estrogen-Dependent Regulation of Osteopontin Expression in Mouse Blastocyst. Int. J. Mol. Sci. 2013, 14, 14504–14517. [Google Scholar] [CrossRef]
- Liaw, L.; Birk, D.E.; Ballas, C.B.; Whitsitt, J.S.; Davidson, J.M.; Hogan, B.L. Altered wound healing in mice lacking a functional osteopontin gene (spp1). J. Clin. Investig. 1998, 101, 1468–1478. [Google Scholar] [CrossRef] [PubMed]
- Aplin, J.; Aplin, J. Embryo implantation: the molecular mechanism remains elusive. Reprod. Biomed. Online 2006, 13, 833–839. [Google Scholar] [CrossRef]
- Bentin-Ley, U.; Horn, T.; Sjögren, A.; Sörensen, S.; Larsen, J.F.; Hamberger, L. Ultrastructure of human blastocyst-endometrial interactions in vitro. Reproduction 2000, 120, 337–350. [Google Scholar] [CrossRef]
- Ruane, P.T.; Berneau, S.C.; Koeck, R.; Watts, J.; Kimber, S.J.; Brison, D.R.; Westwood, M.; Aplin, J.D. Apposition to endometrial epithelial cells activates mouse blastocysts for implantation. Mol. Hum. Reprod. 2017, 23, 617–627. [Google Scholar] [CrossRef]
- Berneau, S.C.; Ruane, P.T.; Brison, D.R.; Kimber, S.J.; Westwood, M.; Aplin, J.D. Investigating the role of CD44 and hyaluronate in embryo-epithelial interaction using an in vitro model. Mol. Hum. Reprod. 2019, 25, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Bontekoe, S.; Heineman, M.J.; Johnson, N.; Blake, D. Adherence compounds in embryo transfer media for assisted reproductive technologies. Cochrane Database Syst. Rev. 2014, 25, CD007421. [Google Scholar] [CrossRef]
- Verhulst, A.; Persy, V.P.; Van Rompay, A.R.; Verstrepen, W.A.; Helbert, M.F.; De Broe, M.E. Osteopontin synthesis and localization along the human nephron. J. Am. Soc. Nephrol. 2002, 13, 1210–1218. [Google Scholar]
- Riew, T.-R.; Kim, H.L.; Jin, X.; Choi, J.-H.; Shin, Y.-J.; Kim, J.S.; Lee, M.-Y. Spatiotemporal expression of osteopontin in the striatum of rats subjected to the mitochondrial toxin 3-nitropropionic acid correlates with microcalcification. Sci. Rep. 2017, 7, 45173. [Google Scholar] [CrossRef] [PubMed]
- Rentsendorj, A.; Sheyn, J.; Fuchs, D.T.; Daley, D.; Salumbides, B.C.; Schubloom, H.E.; Hart, N.J.; Li, S.; Hayden, E.Y.; Teplow, D.B.; et al. A novel role for osteopontin in macrophage-mediated amyloid-beta clearance in Alzheimer’s models. Brain Behav. Immun. 2018, 67, 163–180. [Google Scholar] [CrossRef] [PubMed]
- Dockery, P.; Li, T.; Rogers, A.; Cooke, I.; Lenton, E. The ultrastructure of the glandular epithelium in the timed endometrial biopsy. Hum. Reprod. 1988, 3, 826–834. [Google Scholar] [CrossRef]
- Singh, H.; Aplin, J.D. Endometrial apical glycoproteomic analysis reveals roles for cadherin 6, desmoglein-2 and plexin b2 in epithelial integrity. Mol. Hum. Reprod. 2014, 21, 81–94. [Google Scholar] [CrossRef]
- Johnson, G.A.; Burghardt, R.C.; Joyce, M.M.; Spencer, T.E.; Bazer, F.W.; Gray, C.A.; Pfarrer, C. Osteopontin Is Synthesized by Uterine Glands and a 45-kDa Cleavage Fragment Is Localized at the Uterine-Placental Interface Throughout Ovine Pregnancy1. Boil. Reprod. 2003, 69, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Kon, S.; Yokosaki, Y.; Maeda, M.; Segawa, T.; Horikoshi, Y.; Tsukagoshi, H.; Rashid, M.M.; Morimoto, J.; Inobe, M.; Shijubo, N.; et al. Mapping of functional epitopes of osteopontin by monoclonal antibodies raised against defined internal sequences. J. Cell. Biochem. 2002, 84, 420–432. [Google Scholar] [CrossRef]
- Campbell, S.; Swann, H.R.; Aplin, J.D.; Seif, M.W.; Kimber, S.J.; Elstein, M. CD44 is expressed throughout pre-implantation human embryo development. Hum. Reprod. 1995, 10, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Campbell, S.; Swann, H.; Seif, M.; Kimber, S.; Aplin, J. Cell adhesion molecules on the oocyte and preimplantation human embryo. Mol. Hum. Reprod. 1995, 1, 171–178. [Google Scholar] [CrossRef]
- Albers, A.; Thie, M.; Hohn, H.-P.; Denker, H.-W. Differential Expression and Localization of Integrins and CD44 in the Membrane Domains of Human Uterine Epithelial Cells During the Menstrual Cycle. Cells Tissues Organs 1995, 153, 12–19. [Google Scholar] [CrossRef]
- Bloor, D.; Metcalfe, A.; Brison, D.; Rutherford, A.; Kimber, S. Expression of cell adhesion molecules during human preimplantation embryo development. Mol. Hum. Reprod. 2002, 8, 237–245. [Google Scholar] [CrossRef]
- Ralston, A.; Cox, B.J.; Nishioka, N.; Sasaki, H.; Chea, E.; Rugg-Gunn, P.; Guo, G.; Robson, P.; Draper, J.S.; Rossant, J. Gata3 regulates trophoblast development downstream of Tead4 and in parallel to Cdx2. Development 2010, 137, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.; Dobric, N.; Scott, I.C.; Su, L.; Starovic, M.; St-Pierre, B.; Egan, S.E.; Kingdom, J.C.; Cross, J.C. The Hand1, Stra13 and Gcm1 transcription factors override FGF signaling to promote terminal differentiation of trophoblast stem cells. Dev. Boil. 2004, 271, 26–37. [Google Scholar] [CrossRef]
- Ruane, P.T.; Koeck, R.; Berneau, S.C.; Kimber, S.J.; Westwood, M.; Brison, D.R.; Aplin, J.D. Osmotic stress induces JNK-dependent embryo invasion in a model of implantation. Reproduction 2018, 156, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Bolnick, A.; Shamir, A.; Abdulhasan, M.; Li, Q.; Parker, G.C.; Puscheck, E.E.; Rappolee, D.A. Blastocyst-Derived Stem Cell Populations under Stress: Impact of Nutrition and Metabolism on Stem Cell Potency Loss and Miscarriage. Stem Cell Rev. Rep. 2017, 8, 333–464. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Erikson, D.W.; Burghardt, R.C.; Spencer, T.E.; Wu, G.; Bayless, K.J.; Johnson, G.A.; Bazer, F.W. Secreted phosphoprotein 1 binds integrins to initiate multiple cell signaling pathways, including FRAP1/mTOR, to support attachment and force-generated migration of trophectoderm cells. Matrix Biol. 2010, 29, 369–382. [Google Scholar] [CrossRef]
- Wang, X.; Johnson, G.A.; Burghardt, R.; Wu, G.; Bazer, F.W. Uterine Histotroph and Conceptus Development. I. Cooperative Effects of Arginine and Secreted Phosphoprotein 1 on Proliferation of Ovine Trophectoderm Cells via Activation of the PDK1-Akt/PKB-TSC2-MTORC1 Signaling Cascade. Boil. Reprod. 2015, 92, 51. [Google Scholar] [CrossRef]
- Wang, X.; Johnson, G.A.; Burghardt, R.C.; Wu, G.; Bazer, F.W. Uterine Histotroph and Conceptus Development. II. Arginine and Secreted Phosphoprotein 1 Cooperatively Stimulate Migration and Adhesion of Ovine Trophectoderm Cells via Focal Adhesion-MTORC2 Mediated Cytoskeleton Reorganization. Biol. Reprod. 2016, 95, 71. [Google Scholar] [CrossRef]
- Lichtner, B.; Knaus, P.; Lehrach, H.; Adjaye, J. BMP10 as a potent inducer of trophoblast differentiation in human embryonic and induced pluripotent stem cells. Biomaterials 2013, 34, 9789–9802. [Google Scholar] [CrossRef]
- Gonzalez, I.M.; Martin, P.M.; Burdsal, C.; Sloan, J.L.; Mager, S.; Harris, T.; Sutherland, A.E. Leucine and arginine regulate trophoblast motility through mTOR-dependent and independent pathways in the preimplantation mouse embryo. Dev. Biol. 2012, 361, 286–300. [Google Scholar] [CrossRef]
- Martin, P.M.; Sutherland, A.E. Exogenous Amino Acids Regulate Trophectoderm Differentiation in the Mouse Blastocyst through an mTOR-Dependent Pathway. Dev. Boil. 2001, 240, 182–193. [Google Scholar] [CrossRef]
- Green, C.J.; Fraser, S.T.; Day, M.L. Insulin-like growth factor 1 increases apical fibronectin in blastocysts to increase blastocyst attachment to endometrial epithelial cells in vitro. Hum. Reprod. 2015, 30, 284–298. [Google Scholar] [CrossRef]
- Kang, Y.-J.; Lees, M.; Matthews, L.C.; Kimber, S.J.; Forbes, K.; Aplin, J.D. miR-145 suppresses embryo-epithelial juxtacrine communication at implantation by modulating maternal IGF1R. Development 2015, 128, 804–814. [Google Scholar]
- Clemmons, D.R.; Maile, L.A. Interaction between insulin-like growth factor-I receptor and alphaVbeta3 integrin linked signaling pathways: cellular responses to changes in multiple signaling inputs. Mol. Endocrinol. 2005, 19, 1–11. [Google Scholar] [CrossRef] [PubMed][Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody (Clone/Catalogue Number) | Source |
|---|---|
| OPN (MAB53) | Assay Design |
| OPN (MAB194P) | Maine Biotechnologies |
| OPN (MAB222P) | Maine Biotechnologies |
| Giantin (24586) | Abcam |
| β-Actin (4967) | Cell Signalling Technologies |
| Mouse serum IgG (I8765) | Sigma |
| Gene | Primers (5′–3′) |
|---|---|
| Cdx2 | CAAGGACGTGAGCATGTATCC GTAACCACCGTAGTCCGGGTA |
| Gata3 | CTCGGCCATTCGTACATGGAA GGATACCTCTGCACCGTAGC |
| Eomes | GCGCATGTTTCCTTTCTTGAG GGTCGGCCAGAACCACTTC |
| Elf5 | ACCGATCTGTTCAGCAATGAAG CGCTTGGTCCAGTATTCAGG |
| Gata2 | CACCCCGCCGTATTGAATG CCTGCGAGTCGAGATGGTTG |
| Hand1 | CTACCAGTTACATCGCCTACTTG ACCACCATCCGTCTTTTTGAG |
| Gapdh | AGGTCGGTGTGAACGGATTTG GGGGTCGTTGATGGCAACA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berneau, S.C.; Ruane, P.T.; Brison, D.R.; Kimber, S.J.; Westwood, M.; Aplin, J.D. Characterisation of Osteopontin in an In Vitro Model of Embryo Implantation. Cells 2019, 8, 432. https://doi.org/10.3390/cells8050432
Berneau SC, Ruane PT, Brison DR, Kimber SJ, Westwood M, Aplin JD. Characterisation of Osteopontin in an In Vitro Model of Embryo Implantation. Cells. 2019; 8(5):432. https://doi.org/10.3390/cells8050432
Chicago/Turabian StyleBerneau, Stéphane C, Peter T Ruane, Daniel R Brison, Susan J Kimber, Melissa Westwood, and John D Aplin. 2019. "Characterisation of Osteopontin in an In Vitro Model of Embryo Implantation" Cells 8, no. 5: 432. https://doi.org/10.3390/cells8050432
APA StyleBerneau, S. C., Ruane, P. T., Brison, D. R., Kimber, S. J., Westwood, M., & Aplin, J. D. (2019). Characterisation of Osteopontin in an In Vitro Model of Embryo Implantation. Cells, 8(5), 432. https://doi.org/10.3390/cells8050432