A New Targeted Lipidomics Approach Reveals Lipid Droplets in Liver, Muscle and Heart as a Repository for Diacylglycerol and Ceramide Species in Non-Alcoholic Fatty Liver

Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals and Reagents
2.2. Animals
2.3. Human Muscle Samples
2.4. Sample Preparation
2.5. Lipid Analysis by LC-MS/MS
Calibration and Quantification of Lipid Species
2.6. Method Validation
Recovery
3. Statistical Analysis
4. Results
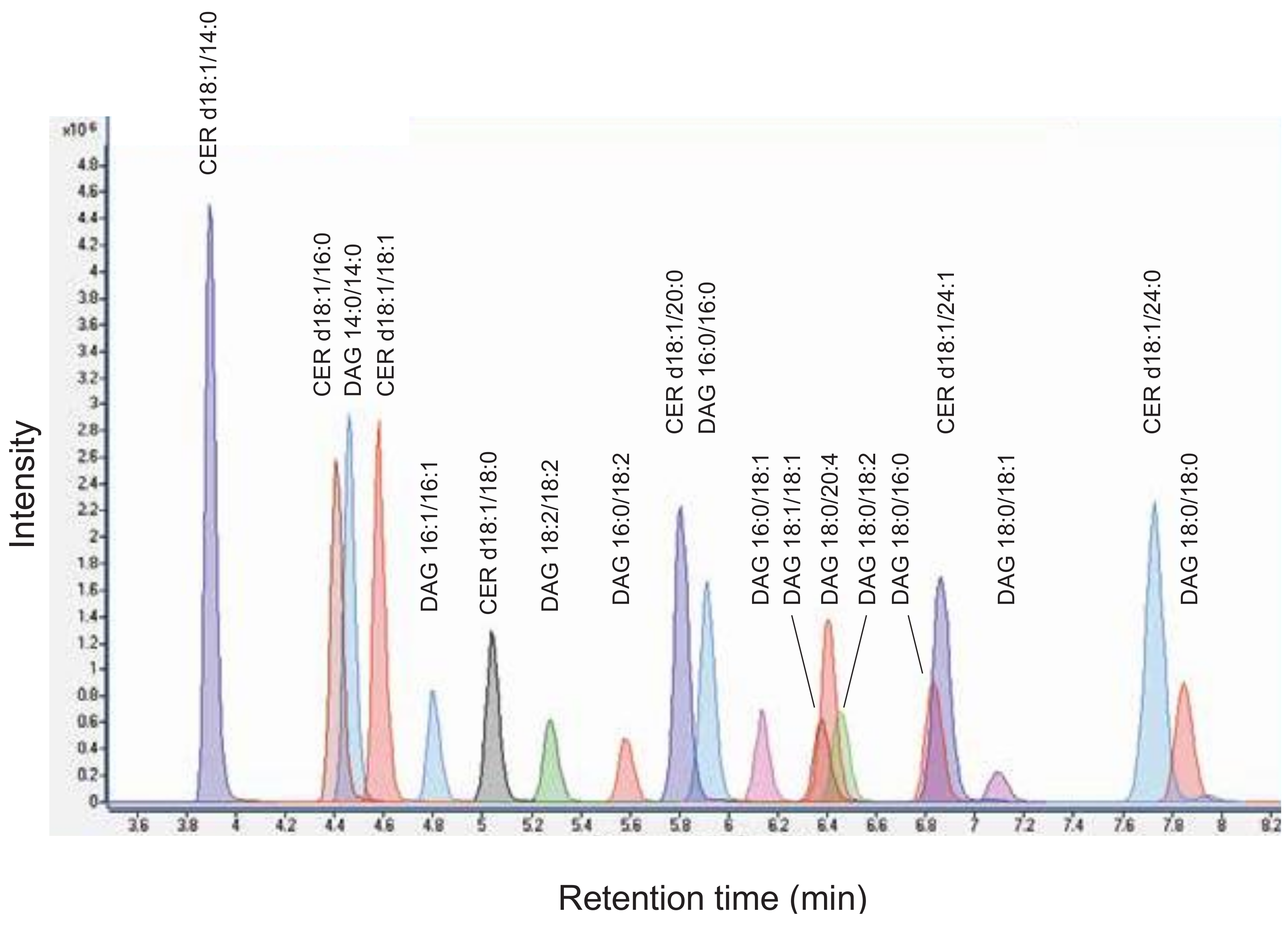
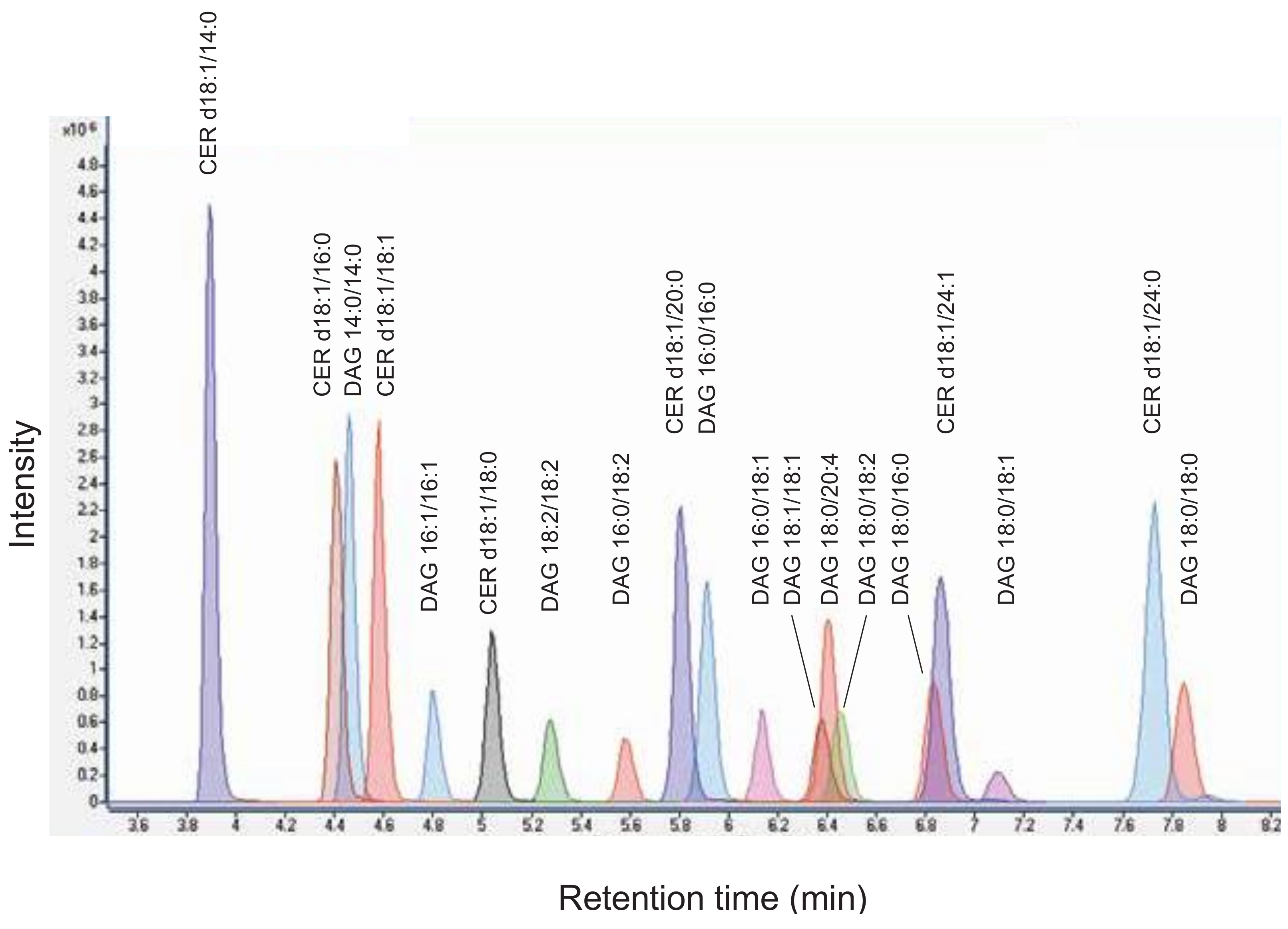
4.1. Analysis of Diacylglycerols and Ceramides by LC-MS/MS
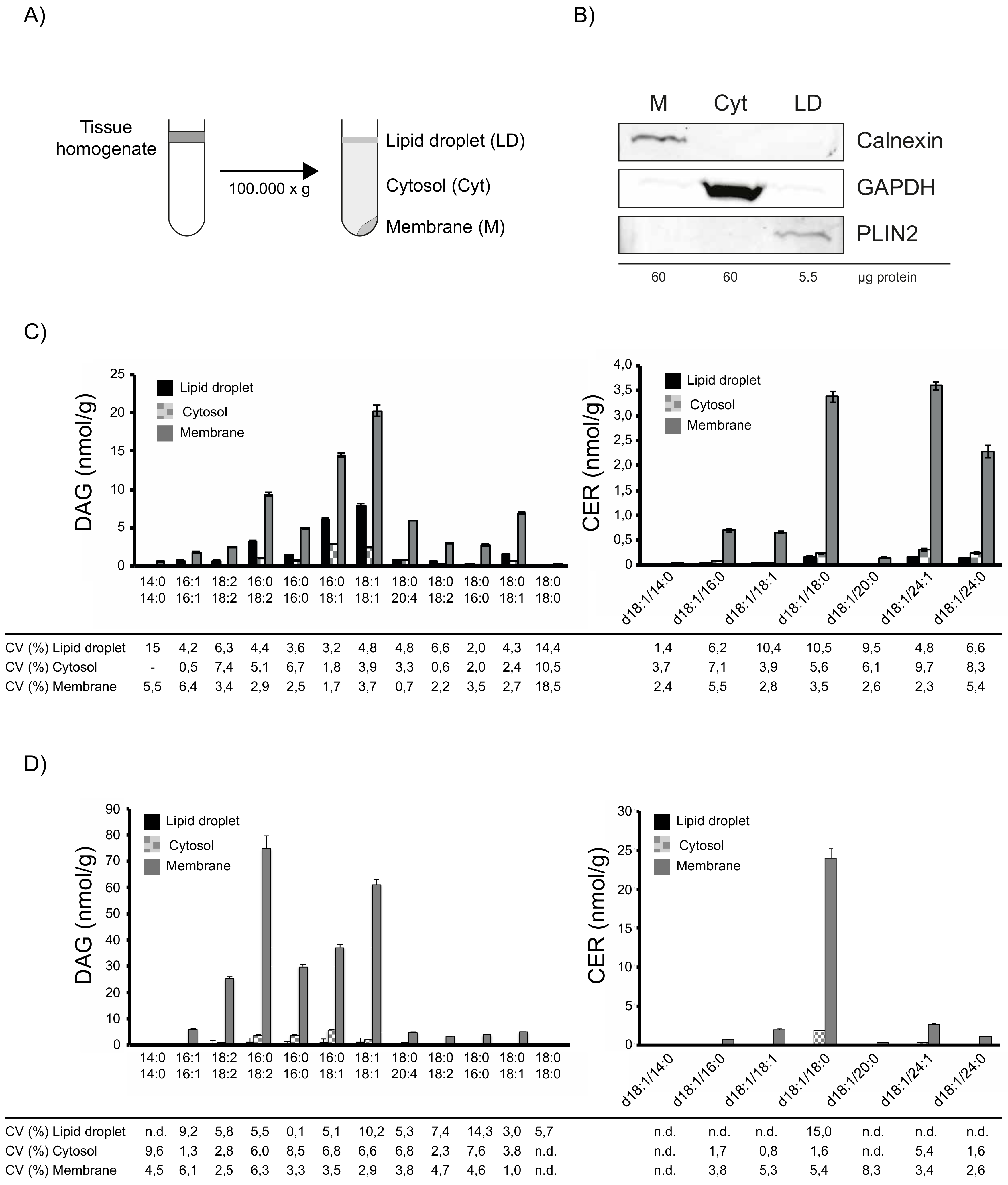
4.2. Tissue Fractionation and LC-MS/MS Analysis of DAGs and CERs
4.3. Multi-Tissue Lipid Analysis in a Mouse Model of NAFLD
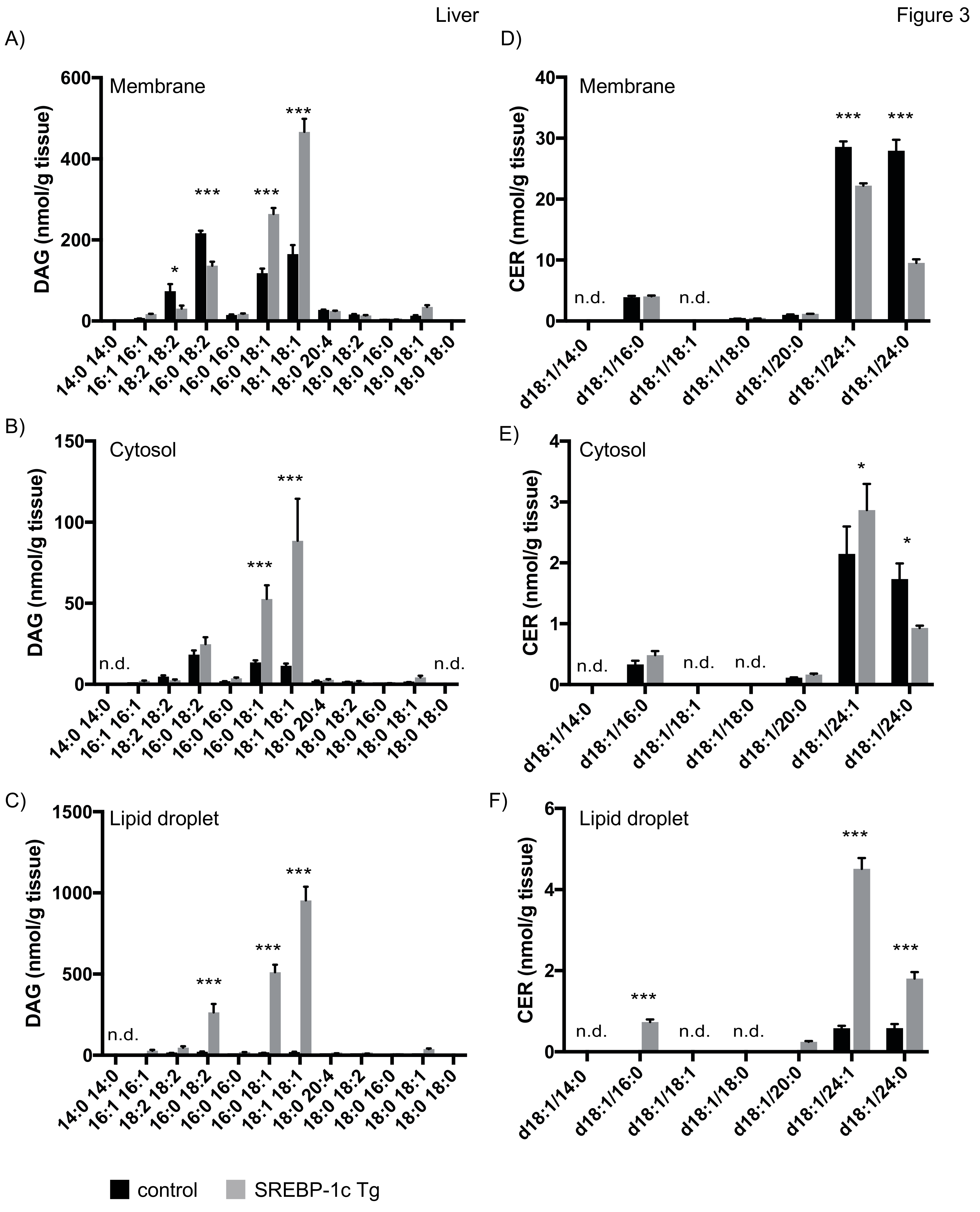
4.3.1. Lipid Analysis of Liver
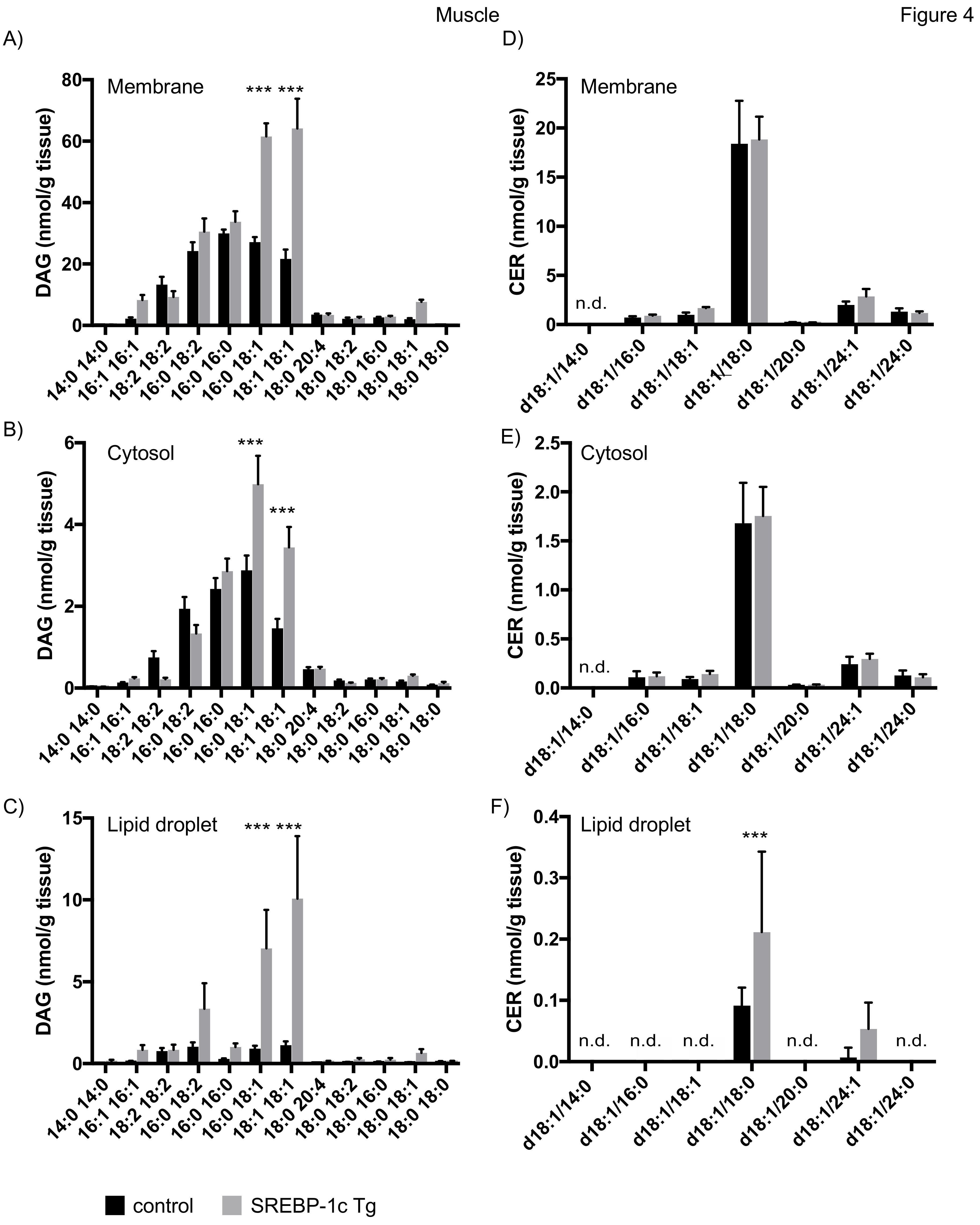
4.3.2. Lipid Analysis of Skeletal Muscle
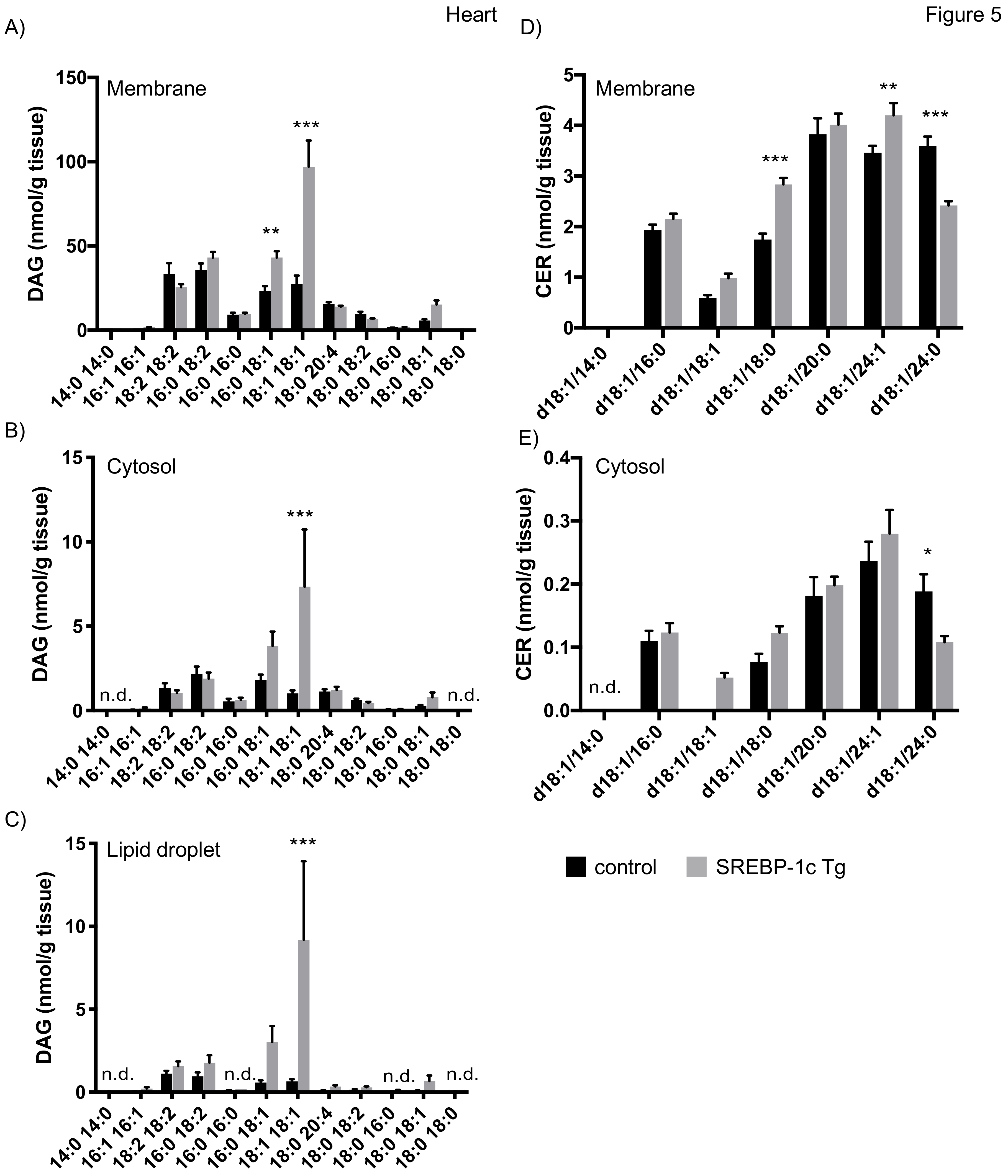
4.3.3. Lipid Analysis of Heart
5. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fahy, E.; Cotter, D.; Sud, M.; Subramaniam, S. Lipid classification, structures and tools. Biochim. Biophys. Acta 2011, 1811, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Perreault, L.; Starling, A.P.; Glueck, D.; Brozinick, J.T.; Sanders, P.; Siddall, P.; Kuo, M.S.; Dabelea, D.; Bergman, B.C. Biomarkers of Ectopic Fat Deposition: The Next Frontier in Serum Lipidomics. J. Clin. Endocrinol. Metab. 2016, 101, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Krssak, M.; Falk Petersen, K.; Dresner, A.; DiPietro, L.; Vogel, S.M.; Rothman, D.L.; Roden, M.; Shulman, G.I. Intramyocellular lipid concentrations are correlated with insulin sensitivity in humans: A 1H NMR spectroscopy study. Diabetologia 1999, 42, 113–116. [Google Scholar] [CrossRef]
- Apostolopoulou, M.; Gordillo, R.; Koliaki, C.; Gancheva, S.; Jelenik, T.; De Filippo, E.; Herder, C.; Markgraf, D.; Jankowiak, F.; Esposito, I.; et al. Specific Hepatic Sphingolipids Relate to Insulin Resistance, Oxidative Stress and Inflammation in Nonalcoholic Steatohepatitis. Diabetes Care 2018, 41, 1235–1243. [Google Scholar] [CrossRef]
- Samuel, V.T.; Shulman, G.I. Nonalcoholic Fatty Liver Disease as a Nexus of Metabolic and Hepatic Diseases. Cell Metab. 2018, 27, 22–41. [Google Scholar] [CrossRef]
- Roden, M. Mechanisms of Disease: Hepatic steatosis in type 2 diabetes—Pathogenesis and clinical relevance. Nat. Clin. Pract. Endocrinol. Metab. 2006, 2, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Szendroedi, J.; Roden, M. Ectopic lipids and organ function. Curr. Opin. Lipidol. 2009, 20, 50–56. [Google Scholar] [CrossRef]
- Buhman, K.K.; Chen, H.C.; Farese, R.V., Jr. The enzymes of neutral lipid synthesis. J. Biol. Chem. 2001, 276, 40369–40372. [Google Scholar] [CrossRef]
- Brasaemle, D.L.; Wolins, N.E. Packaging of fat: An evolving model of lipid droplet assembly and expansion. J. Biol. Chem. 2012, 287, 2273–2279. [Google Scholar] [CrossRef]
- Wilfling, F.; Wang, H.; Haas, J.T.; Krahmer, N.; Gould, T.J.; Uchida, A.; Cheng, J.X.; Graham, M.; Christiano, R.; Frohlich, F.; et al. Triacylglycerol synthesis enzymes mediate lipid droplet growth by relocalizing from the ER to lipid droplets. Dev. Cell 2013, 24, 384–399. [Google Scholar] [CrossRef] [PubMed]
- Kuerschner, L.; Moessinger, C.; Thiele, C. Imaging of lipid biosynthesis: How a neutral lipid enters lipid droplets. Traffic 2008, 9, 338–352. [Google Scholar] [CrossRef]
- Duncan, R.E.; Ahmadian, M.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Regulation of lipolysis in adipocytes. Annu. Rev. Nutr. 2007, 27, 79–101. [Google Scholar] [CrossRef] [PubMed]
- Lankester, D.L.; Brown, A.M.; Zammit, V.A. Use of cytosolic triacylglycerol hydrolysis products and of exogenous fatty acid for the synthesis of triacylglycerol secreted by cultured rat hepatocytes. J. Lipid. Res. 1998, 39, 1889–1895. [Google Scholar]
- Markgraf, D.F.; Klemm, R.W.; Junker, M.; Hannibal-Bach, H.K.; Ejsing, C.S.; Rapoport, T.A. An ER protein functionally couples neutral lipid metabolism on lipid droplets to membrane lipid synthesis in the ER. Cell Rep. 2014, 6, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Larsen, P.J.; Tennagels, N. On ceramides, other sphingolipids and impaired glucose homeostasis. Mol. Metab. 2014, 3, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Obeid, L.M. Many ceramides. J. Biol. Chem. 2011, 286, 27855–27862. [Google Scholar] [CrossRef]
- Samuel, V.T.; Shulman, G.I. Mechanisms for insulin resistance: Common threads and missing links. Cell 2012, 148, 852–871. [Google Scholar] [CrossRef] [PubMed]
- Chavez, J.A.; Summers, S.A. A ceramide-centric view of insulin resistance. Cell Metab. 2012, 15, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Blouin, C.M.; Prado, C.; Takane, K.K.; Lasnier, F.; Garcia-Ocana, A.; Ferre, P.; Dugail, I.; Hajduch, E. Plasma membrane subdomain compartmentalization contributes to distinct mechanisms of ceramide action on insulin signaling. Diabetes 2010, 59, 600–610. [Google Scholar] [CrossRef]
- Chavez, J.A.; Knotts, T.A.; Wang, L.P.; Li, G.; Dobrowsky, R.T.; Florant, G.L.; Summers, S.A. A role for ceramide but not diacylglycerol, in the antagonism of insulin signal transduction by saturated fatty acids. J. Biol. Chem. 2003, 278, 10297–10303. [Google Scholar] [CrossRef]
- Bikman, B.T.; Summers, S.A. Ceramides as modulators of cellular and whole-body metabolism. J. Clin. Investig. 2011, 121, 4222–4230. [Google Scholar] [CrossRef] [PubMed]
- Holland, W.L.; Bikman, B.T.; Wang, L.P.; Yuguang, G.; Sargent, K.M.; Bulchand, S.; Knotts, T.A.; Shui, G.; Clegg, D.J.; Wenk, M.R.; et al. Lipid-induced insulin resistance mediated by the proinflammatory receptor TLR4 requires saturated fatty acid-induced ceramide biosynthesis in mice. J. Clin. Investig. 2011, 121, 1858–1870. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Soos, T.J.; Li, X.; Wu, J.; Degennaro, M.; Sun, X.; Littman, D.R.; Birnbaum, M.J.; Polakiewicz, R.D. Protein kinase C Theta inhibits insulin signaling by phosphorylating IRS1 at Ser(1101). J. Biol. Chem. 2004, 279, 45304–45307. [Google Scholar] [CrossRef]
- Szendroedi, J.; Yoshimura, T.; Phielix, E.; Koliaki, C.; Marcucci, M.; Zhang, D.; Jelenik, T.; Muller, J.; Herder, C.; Nowotny, P.; et al. Role of diacylglycerol activation of PKCtheta in lipid-induced muscle insulin resistance in humans. Proc. Natl. Acad. Sci. USA 2014, 111, 9597–9602. [Google Scholar] [CrossRef]
- Nowotny, B.; Zahiragic, L.; Krog, D.; Nowotny, P.J.; Herder, C.; Carstensen, M.; Yoshimura, T.; Szendroedi, J.; Phielix, E.; Schadewaldt, P.; et al. Mechanisms underlying the onset of oral lipid-induced skeletal muscle insulin resistance in humans. Diabetes 2013, 62, 2240–2248. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.J.; Samuel, V.T.; Petersen, K.F.; Shulman, G.I. The role of hepatic lipids in hepatic insulin resistance and type 2 diabetes. Nature 2014, 510, 84–91. [Google Scholar] [CrossRef]
- Jelenik, T.; Flogel, U.; Alvarez-Hernandez, E.; Scheiber, D.; Zweck, E.; Ding, Z.; Rothe, M.; Mastrototaro, L.; Kohlhaas, V.; Kotzka, J.; et al. Insulin Resistance and Vulnerability to Cardiac Ischemia. Diabetes 2018, 67, 2695–2702. [Google Scholar] [CrossRef] [PubMed]
- Bergman, B.C.; Hunerdosse, D.M.; Kerege, A.; Playdon, M.C.; Perreault, L. Localisation and composition of skeletal muscle diacylglycerol predicts insulin resistance in humans. Diabetologia 2012, 55, 1140–1150. [Google Scholar] [CrossRef] [PubMed]
- Perreault, L.; Newsom, S.A.; Strauss, A.; Kerege, A.; Kahn, D.E.; Harrison, K.A.; Snell-Bergeon, J.K.; Nemkov, T.; D’Alessandro, A.; Jackman, M.R.; et al. Intracellular localization of diacylglycerols and sphingolipids influences insulin sensitivity and mitochondrial function in human skeletal muscle. JCI Insight 2018, 3. [Google Scholar] [CrossRef]
- Jelenik, T.; Kaul, K.; Sequaris, G.; Flogel, U.; Phielix, E.; Kotzka, J.; Knebel, B.; Fahlbusch, P.; Horbelt, T.; Lehr, S.; et al. Mechanisms of Insulin Resistance in Primary and Secondary Nonalcoholic Fatty Liver. Diabetes 2017, 66, 2241–2253. [Google Scholar] [CrossRef] [PubMed]
- Ritter, O.; Jelenik, T.; Roden, M. Lipid-mediated muscle insulin resistance: Different fat, different pathways? J. Mol. Med. (Berl) 2015, 93, 831–843. [Google Scholar] [CrossRef]
- Krahmer, N.; Najafi, B.; Schueder, F.; Quagliarini, F.; Steger, M.; Seitz, S.; Kasper, R.; Salinas, F.; Cox, J.; Uhlenhaut, N.H.; et al. Organellar Proteomics and Phospho-Proteomics Reveal Subcellular Reorganization in Diet-Induced Hepatic Steatosis. Dev. Cell 2018, 47, 205–221. [Google Scholar] [CrossRef]
- Szendroedi, J.; Saxena, A.; Weber, K.S.; Strassburger, K.; Herder, C.; Burkart, V.; Nowotny, B.; Icks, A.; Kuss, O.; Ziegler, D.; et al. Cohort profile: The German Diabetes Study (GDS). Cardiovasc. Diabetol. 2016, 15, 59. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Tilg, H.; Moschen, A.R.; Roden, M. NAFLD and diabetes mellitus. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 32–42. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Kumashiro, N.; Erion, D.M.; Zhang, D.; Kahn, M.; Beddow, S.A.; Chu, X.; Still, C.D.; Gerhard, G.S.; Han, X.; Dziura, J.; et al. Cellular mechanism of insulin resistance in nonalcoholic fatty liver disease. Proc. Natl. Acad. Sci. USA 2011, 108, 16381–16385. [Google Scholar] [CrossRef]
- Shimomura, I.; Hammer, R.E.; Richardson, J.A.; Ikemoto, S.; Bashmakov, Y.; Goldstein, J.L.; Brown, M.S. Insulin resistance and diabetes mellitus in transgenic mice expressing nuclear SREBP-1c in adipose tissue: Model for congenital generalized lipodystrophy. Genes Dev. 1998, 12, 3182–3194. [Google Scholar] [CrossRef]
- Erion, D.M.; Shulman, G.I. Diacylglycerol-mediated insulin resistance. Nat. Med. 2010, 16, 400–402. [Google Scholar] [CrossRef]
- Marignani, P.A.; Epand, R.M.; Sebaldt, R.J. Acyl chain dependence of diacylglycerol activation of protein kinase C activity in vitro. Biochem. Biophys. Res. Commun. 1996, 225, 469–473. [Google Scholar] [CrossRef]
- Gancheva, S.; Jelenik, T.; Alvarez-Hernandez, E.; Roden, M. Interorgan Metabolic Crosstalk in Human Insulin Resistance. Physiol. Rev. 2018, 98, 1371–1415. [Google Scholar] [CrossRef]
- Senkal, C.E.; Salama, M.F.; Snider, A.J.; Allopenna, J.J.; Rana, N.A.; Koller, A.; Hannun, Y.A.; Obeid, L.M. Ceramide Is Metabolized to Acylceramide and Stored in Lipid Droplets. Cell Metab. 2017, 25, 686–697. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Class | Lipid Species | Ion | MRM | CE (V) | RT (min) | LOD (fmol) On Column |
|---|---|---|---|---|---|---|
| DAG | 14:0 14:0 | [M + NH4]+ | 530.2 → 285.0 | 16 | 4.50 | 2.0 |
| DAG | 16:1 16:1 | [M + NH4]+ | 582.2 → 311.1 | 26 | 4.84 | 1.5 |
| DAG | 18:2 18:2 | [M + NH4]+ | 634.3 → 337.2 | 26 | 5.30 | 1.6 |
| DAG | 16:0 18:2 | [M + NH4]+ | 610.2 → 313.2 | 26 | 5.63 | 1.7 |
| DAG | 16:0 16:0 | [M + NH4]+ | 586.2 → 313.0 | 16 | 5.96 | 1.8 |
| DAG | 16:0 18:1 | [M + NH4]+ | 612.1 → 339.1 | 30 | 6.20 | 1.7 |
| DAG | 18:1 18:1 | [M + NH4]+ | 638.2 → 339.1 | 28 | 6.43 | 1.6 |
| DAG | 18:0 20:4 | [M + NH4]+ | 662.2 → 341.3 | 20 | 6.45 | 1.6 |
| DAG | 18:0 18:2 | [M + NH4]+ | 638.1 → 341.0 | 24 | 6.50 | 1.6 |
| DAG | 18:0 16:0 | [M + NH4]+ | 614.2 → 313.1 | 24 | 6.89 | 1.7 |
| DAG | 18:0 18:1 | [M + NH4]+ | 640.2 → 341.2 | 30 | 7.15 | 1.6 |
| DAG | 18:0 18:0 | [M + NH4]+ | 642.1 → 341.0 | 28 | 7.92 | 1.6 |
| CER | d18:1/14:0 | [M + H]+ | 510.3 → 264.0 | 34 | 3.93 | 2.0 |
| CER | d18:1/16:0 | [M + H]+ | 538.2 → 264.0 | 32 | 4.44 | 1.9 |
| CER | d18:1/18:1 | [M + H]+ | 564.2 → 264.0 | 36 | 4.61 | 1.8 |
| CER | d18:1/18:0 | [M + H]+ | 566.1 → 264.1 | 30 | 5.07 | 1.8 |
| CER | d18:1/20:0 | [M + H]+ | 594.2 → 264.0 | 36 | 5.86 | 1.7 |
| CER | d18:1/24:1 | [M + H]+ | 648.3 → 264.1 | 34 | 6.91 | 1.5 |
| CER | d18:1/24:0 | [M + H]+ | 650.5 → 264.0 | 34 | 7.78 | 1.5 |
| Lipid Class | Lipid Species | Calibration Range (ng) | Calibration Range (pmol) | IS Added (ng; pmol) | Correlation Coefficient (mean ± SD) |
|---|---|---|---|---|---|
| DAG | 14:0 14:0 | 1–1000 | 1.95–1950 | 500; 830 | 0.999 ± 0.001 |
| DAG | 16:1 16:1 | 1–1000 | 1.77–1770 | 500; 830 | 0.998 ± 0.001 |
| DAG | 18:2 18:2 | 1–1000 | 1.62–1621 | 500; 830 | 0.998 ± 0.001 |
| DAG | 16:0 18:2 | 1–1000 | 1.69–1687 | 500; 830 | 0.998 ± 0.001 |
| DAG | 16:0 16:0 | 1–1000 | 1.76–1758 | 500; 830 | 0.999 ± 0.000 |
| DAG | 16:0 18:1 | 1–1000 | 1.68–1681 | 500; 830 | 0.998 ± 0.000 |
| DAG | 18:1 18:1 | 1–1000 | 1.61–1610 | 500; 830 | 0.999 ± 0.000 |
| DAG | 18:0 20:4 | 1–1000 | 1.55–1550 | 500; 830 | 0.998 ± 0.000 |
| DAG | 18:0 18:2 | 1–1000 | 1.61–1610 | 500; 830 | 0.998 ± 0.000 |
| DAG | 18:0 16:0 | 1–1000 | 1.68–1675 | 500; 830 | 0.999 ± 0.001 |
| DAG | 18:0 18:1 | 1–1000 | 1.61–1605 | 500; 830 | 0.997 ± 0.001 |
| DAG | 18:0 18:0 | 1–1000 | 1.77–1585 | 500; 830 | 0.999 ± 0.001 |
| CER | d18:1/14:0 | 1–200 | 1.96–392 | 100; 181 | 0.997 ± 0.002 |
| CER | d18:1/16:0 | 1–200 | 1.86–372 | 100; 181 | 0.999 ± 0.001 |
| CER | d18:1/18:1 | 1–200 | 1.77–355 | 100; 181 | 0.999 ± 0.000 |
| CER | d18:1/18:0 | 1–200 | 1.77–353 | 100; 181 | 0.995 ± 0.003 |
| CER | d18:1/20:0 | 1–200 | 1.68–337 | 100; 181 | 0.997 ± 0.002 |
| CER | d18:1/24:1 | 1–200 | 1.54–309 | 100; 181 | 0.997 ± 0.002 |
| CER | d18:1/24:0 | 1–200 | 1.54–308 | 100; 181 | 0.997 ± 0.001 |
| Lipid Class | Lipid Species | Spiked Amount (ng; pmol) | Recovery (%) |
|---|---|---|---|
| DAG | 14:0 14:0 | 100; 195 | 73.6 ± 3.4 |
| DAG | 16:1 16:1 | 100; 177 | 86.7 ± 2.7 |
| DAG | 18:2 18:2 | 100; 162 | 88.1 ± 4.2 |
| DAG | 16:0 18:2 | 100; 169 | 91.7 ± 0.9 |
| DAG | 16:0 16:0 | 100; 176 | 105.2 ± 3.5 |
| DAG | 16:0 18:1 | 100; 168 | 105.3 ± 3.2 |
| DAG | 18:1 18:1 | 100; 161 | 115.1 ± 1.0 |
| DAG | 18:0 20:4 | 100; 155 | 97.0 ± 4.2 |
| DAG | 18:0 18:2 | 100; 161 | 97.1 ± 2.7 |
| DAG | 18:0 16:0 | 100; 168 | 79.7 ± 0.5 |
| DAG | 18:0 18:1 | 100; 161 | 83.0 ± 1.0 |
| DAG | 18:0 18:0 | 100; 160 | 72.4 ± 5.3 |
| CER | d18:1/14:0 | 70; 137 | 97.0 ± 4.3 |
| CER | d18:1/16:0 | 70; 130 | 91.0 ± 2.9 |
| CER | d18:1/18:1 | 70; 124 | 87.2 ± 2.8 |
| CER | d18:1/18:0 | 70; 124 | 81.3 ± 2.5 |
| CER | d18:1/20:0 | 70; 118 | 118.3 ± 3.3 |
| CER | d18:1/24:1 | 70; 108 | 89.2 ± 2.3 |
| CER | d18:1/24:0 | 70; 108 | 118.3 ± 0.6 |
| Lipid Class | Lipid Species | Spiked Amount (ng; pmol) | CV (%) | Accuracy (%) |
|---|---|---|---|---|
| DAG | 14:0 14:0 | 125; 244 | 6.4 | 109 ± 3.6 |
| DAG | 16:1 16:1 | 125; 221 | 9.0 | 108 ± 4.5 |
| DAG | 18:2 18:2 | 125; 203 | 11.0 | 102 ± 5.9 |
| DAG | 16:0 18:2 | 125; 211 | 8.6 | 87.0 ± 8.1 |
| DAG | 16:0 16:0 | 125; 220 | 7.4 | 106 ± 5.4 |
| DAG | 16:0 18:1 | 125; 210 | 6.3 | 111 ± 10.3 |
| DAG | 18:1 18:1 | 125; 201 | 8.3 | 103 ± 14.8 |
| DAG | 18:0 20:4 | 125; 194 | 8.3 | 114 ± 5.7 |
| DAG | 18:0 18:2 | 125; 201 | 9.0 | 111 ± 6.2 |
| DAG | 18:0 16:0 | 125; 209 | 6.3 | 106 ± 4.0 |
| DAG | 18:0 18:1 | 125; 201 | 5.4 | 98.9 ± 6.0 |
| DAG | 18:0 18:0 | 125; 201 | 1.3 | 95.0 ± 0.6 |
| CER | d18:1/14:0 | 100; 196 | 4.9 | 83.6 ± 1.7 |
| CER | d18:1/16:0 | 100; 186 | 4.1 | 107 ± 2.1 |
| CER | d18:1/18:1 | 100; 177 | 5.4 | 96.6 ± 2.2 |
| CER | d18:1/18:0 | 100; 177 | 4.1 | 116 ± 2.4 |
| CER | d18:1/20:0 | 100; 168 | 7.7 | 123 ± 4.3 |
| CER | d18:1/24:1 | 100; 154 | 9.0 | 110 ± 5.2 |
| CER | d18:1/24:0 | 100; 154 | 7.2 | 118 ± 4.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Preuss, C.; Jelenik, T.; Bódis, K.; Müssig, K.; Burkart, V.; Szendroedi, J.; Roden, M.; Markgraf, D.F. A New Targeted Lipidomics Approach Reveals Lipid Droplets in Liver, Muscle and Heart as a Repository for Diacylglycerol and Ceramide Species in Non-Alcoholic Fatty Liver. Cells 2019, 8, 277. https://doi.org/10.3390/cells8030277
Preuss C, Jelenik T, Bódis K, Müssig K, Burkart V, Szendroedi J, Roden M, Markgraf DF. A New Targeted Lipidomics Approach Reveals Lipid Droplets in Liver, Muscle and Heart as a Repository for Diacylglycerol and Ceramide Species in Non-Alcoholic Fatty Liver. Cells. 2019; 8(3):277. https://doi.org/10.3390/cells8030277
Chicago/Turabian StylePreuss, Christina, Tomas Jelenik, Kálmán Bódis, Karsten Müssig, Volker Burkart, Julia Szendroedi, Michael Roden, and Daniel F. Markgraf. 2019. "A New Targeted Lipidomics Approach Reveals Lipid Droplets in Liver, Muscle and Heart as a Repository for Diacylglycerol and Ceramide Species in Non-Alcoholic Fatty Liver" Cells 8, no. 3: 277. https://doi.org/10.3390/cells8030277
APA StylePreuss, C., Jelenik, T., Bódis, K., Müssig, K., Burkart, V., Szendroedi, J., Roden, M., & Markgraf, D. F. (2019). A New Targeted Lipidomics Approach Reveals Lipid Droplets in Liver, Muscle and Heart as a Repository for Diacylglycerol and Ceramide Species in Non-Alcoholic Fatty Liver. Cells, 8(3), 277. https://doi.org/10.3390/cells8030277