Activation of COUP-TFI by a Novel Diindolylmethane Derivative

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines, Chemicals, and Reagents
2.2. Transfection Assays
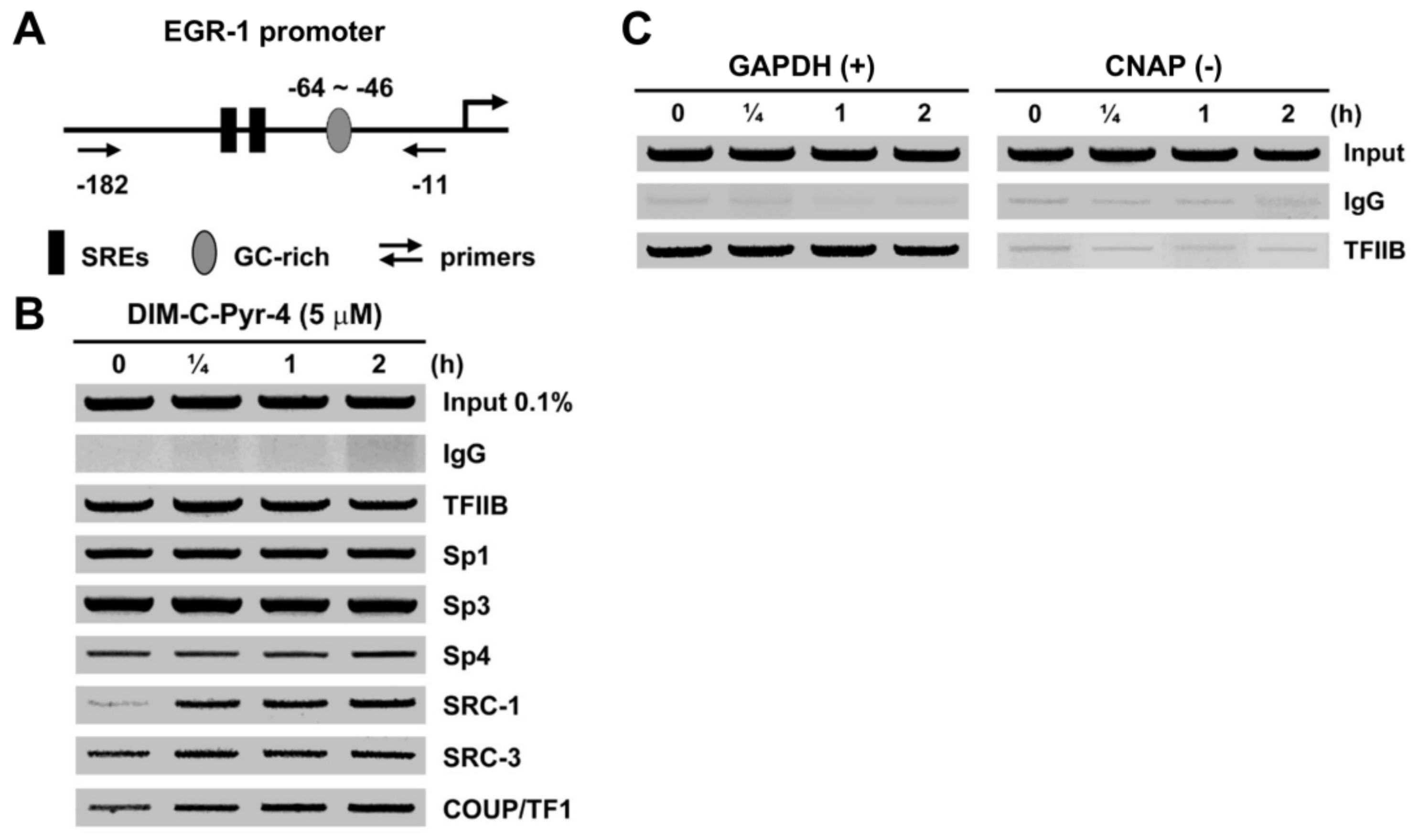
2.3. Chromatin Immunoprecipitation (ChIP) Assay
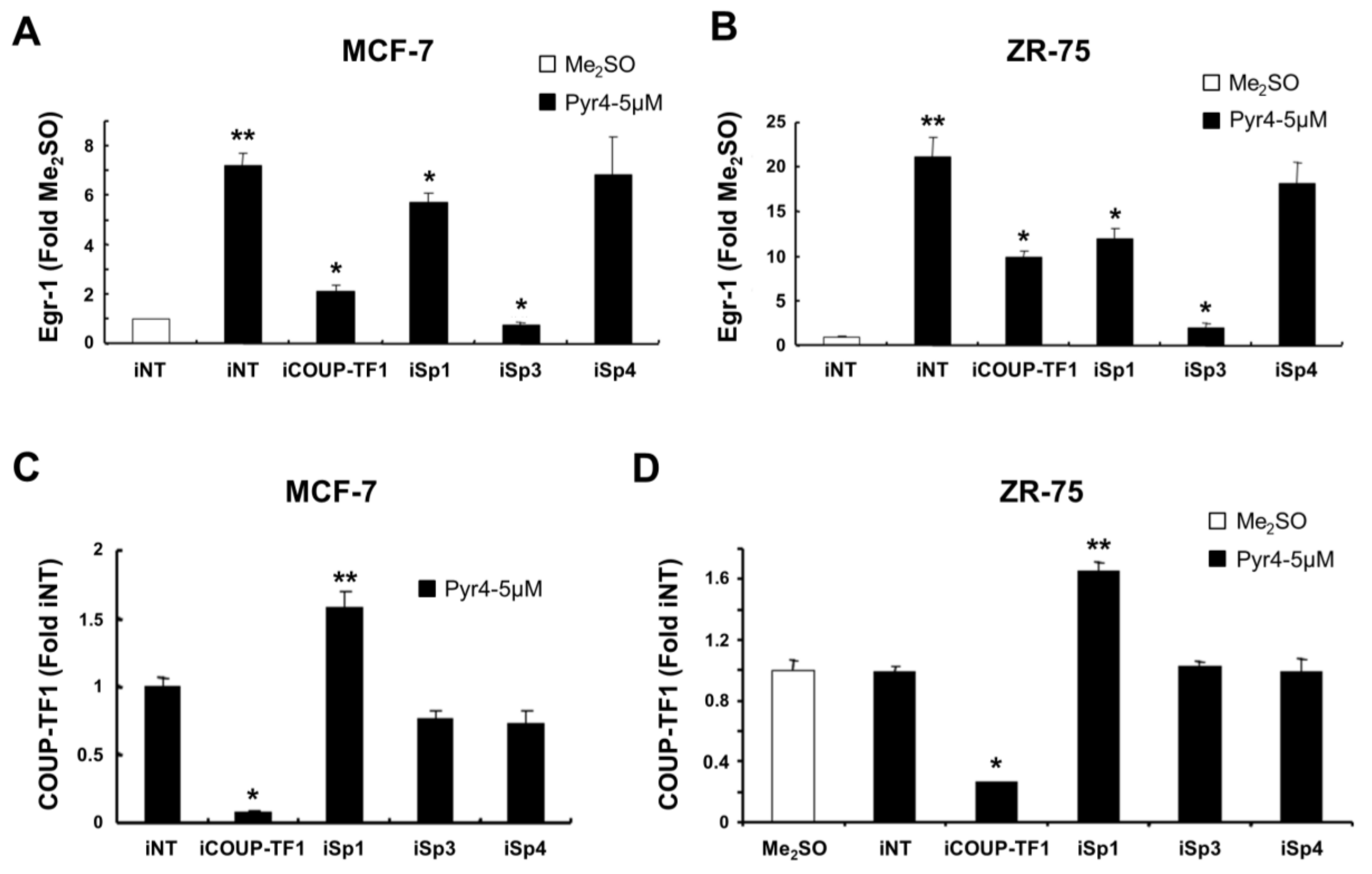
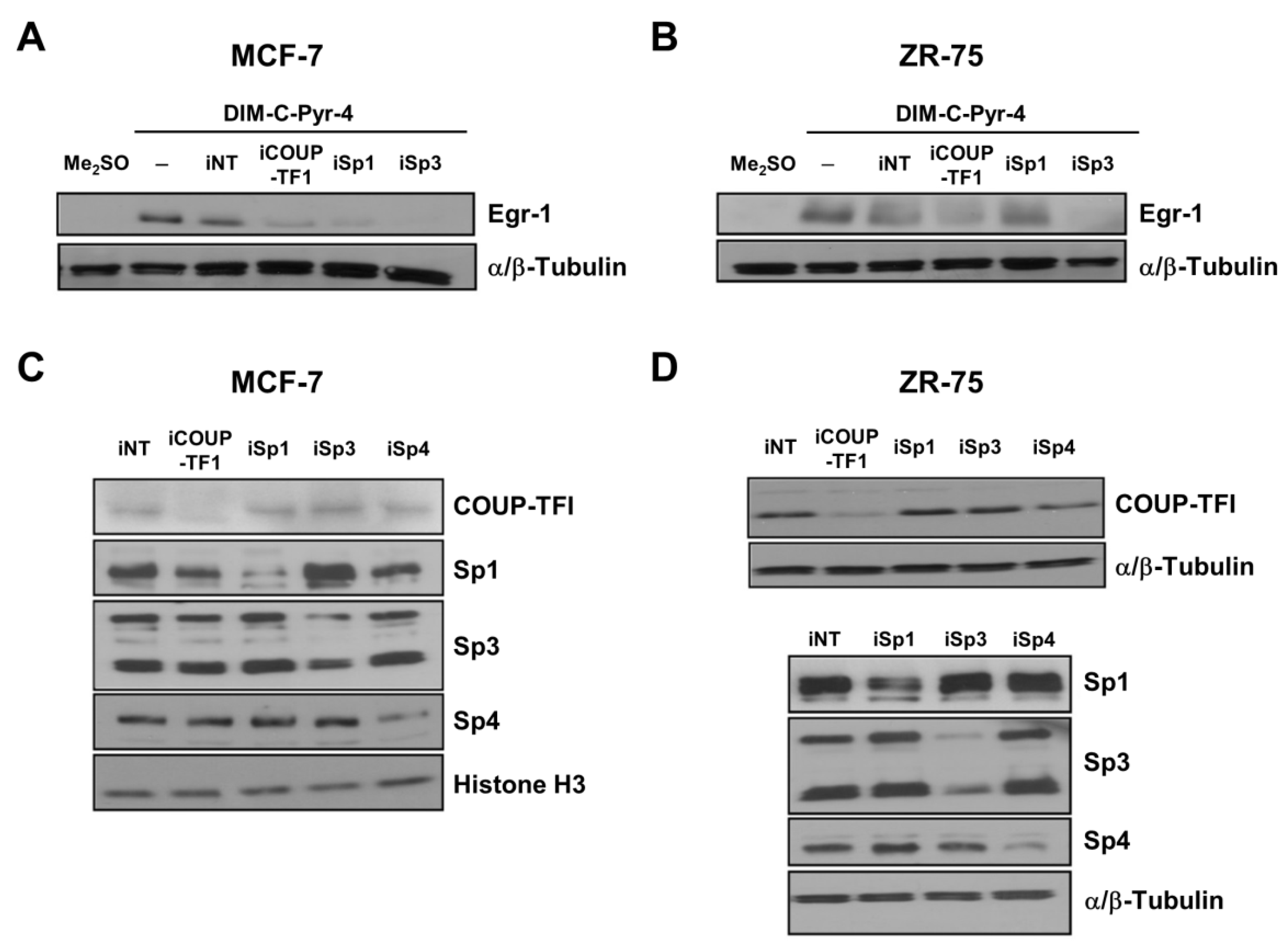
2.4. RNA Interference Studies
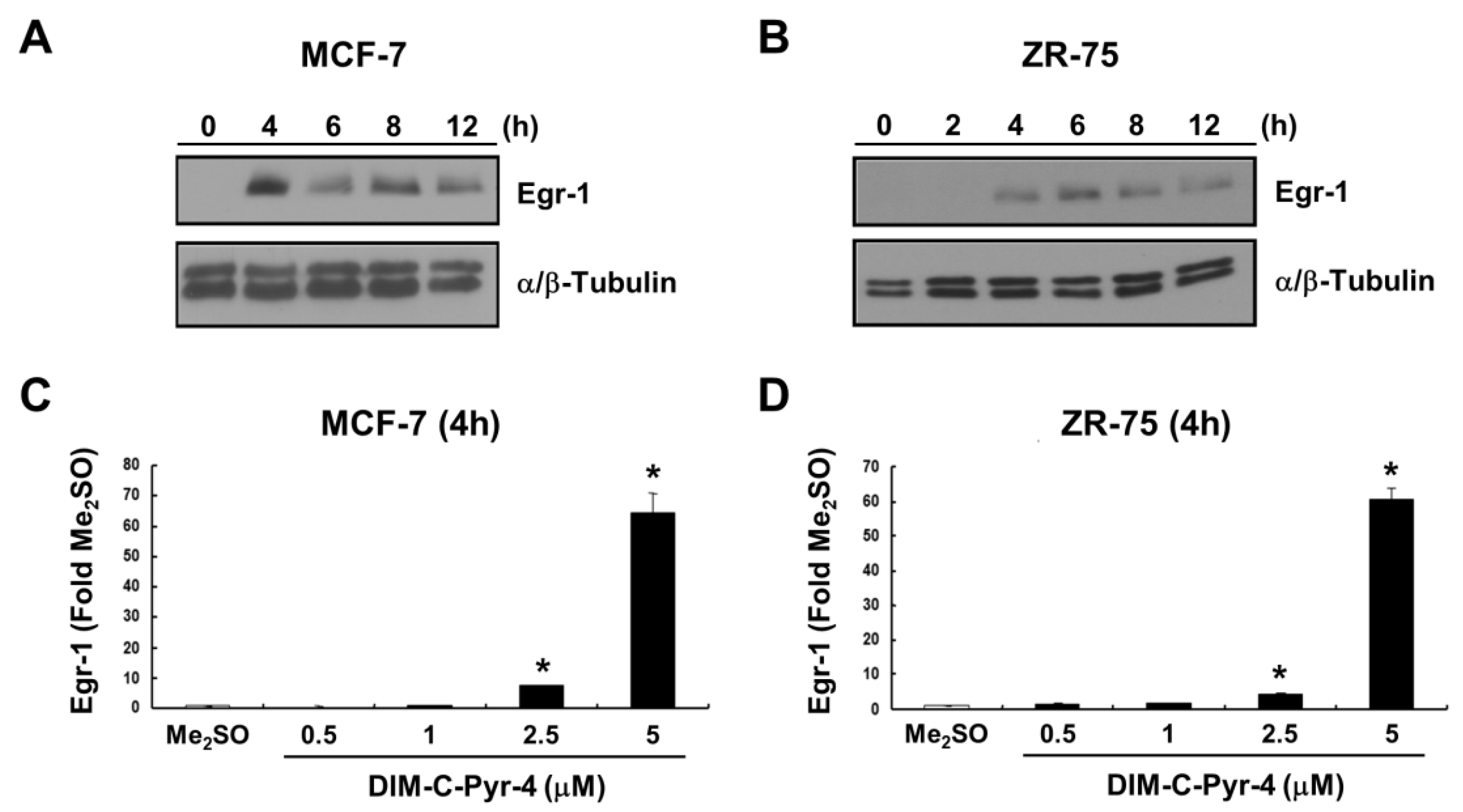
2.5. Induction of Egr-1 Protein/mRNA by DIM-C-Pyr-4
2.6. Clones and Reagents
2.7. Computation Analysis of COUP-TFI and COUP-TFII Ligand Binding
2.8. Statistical Analysis
3. Results
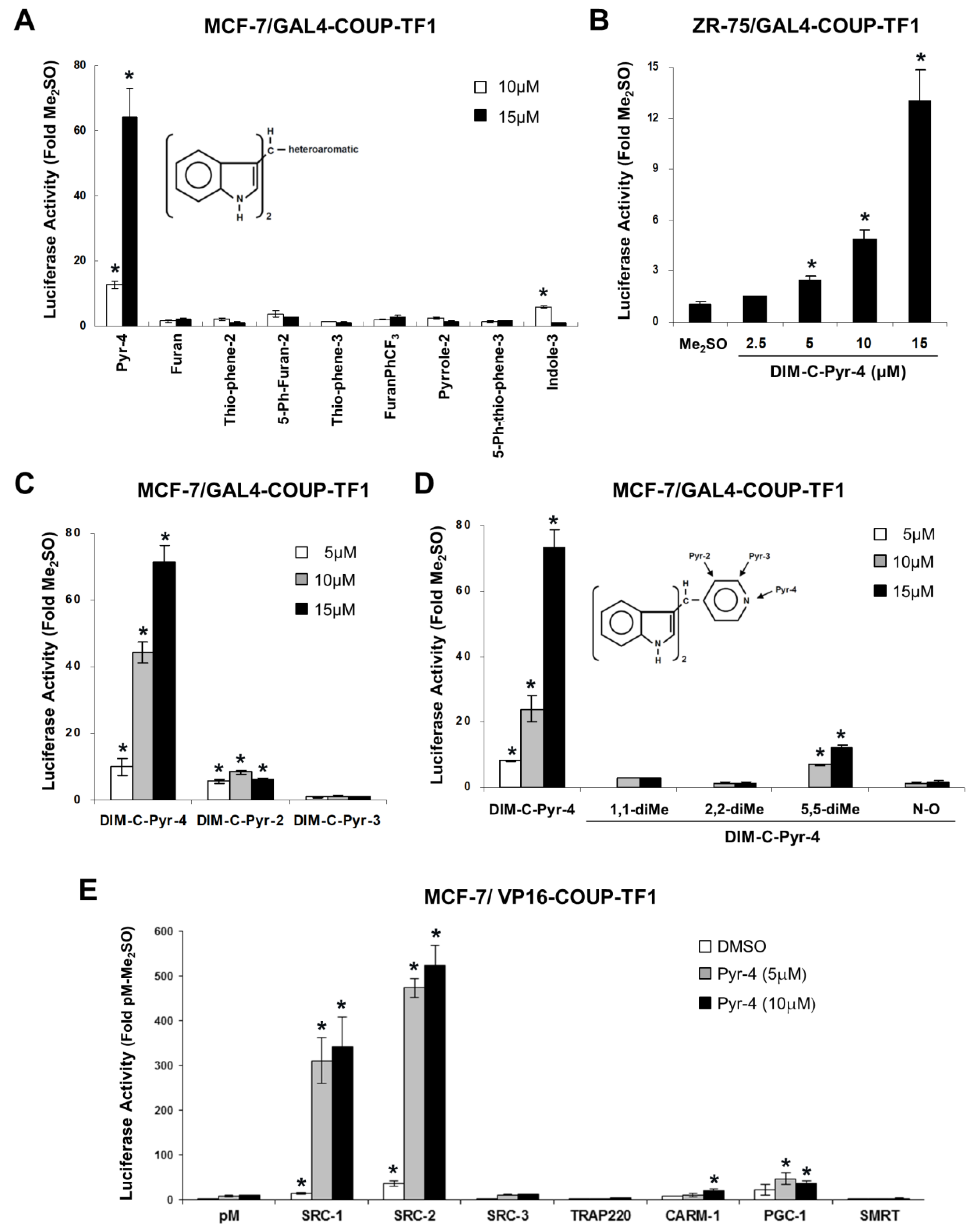
3.1. Structure-Dependent Activation of COUP-TFI and Interaction with Coactivators and Silencing Mediator of Retinoic Acid and Thyroid Hormone Receptor (SMRT)
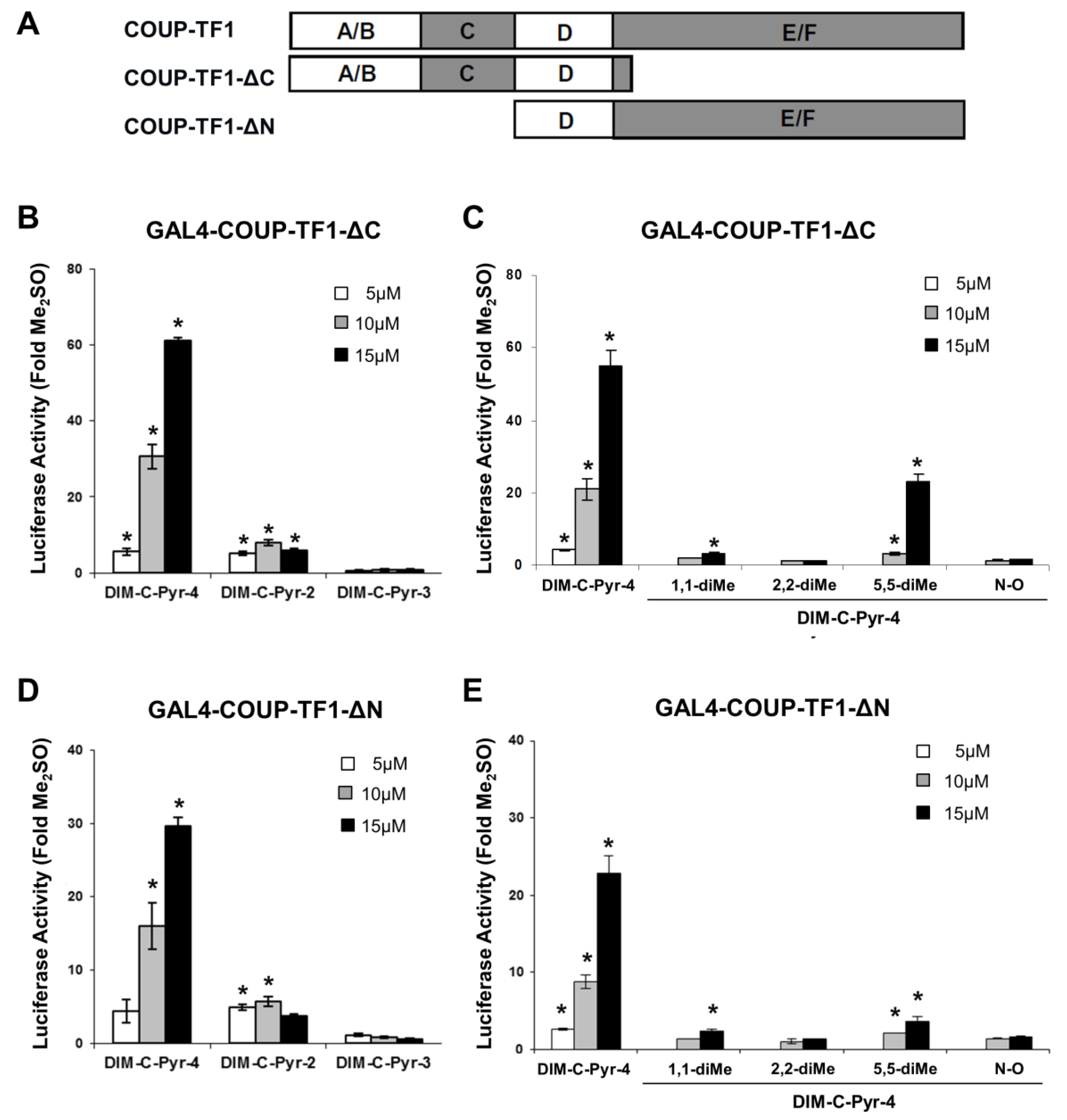
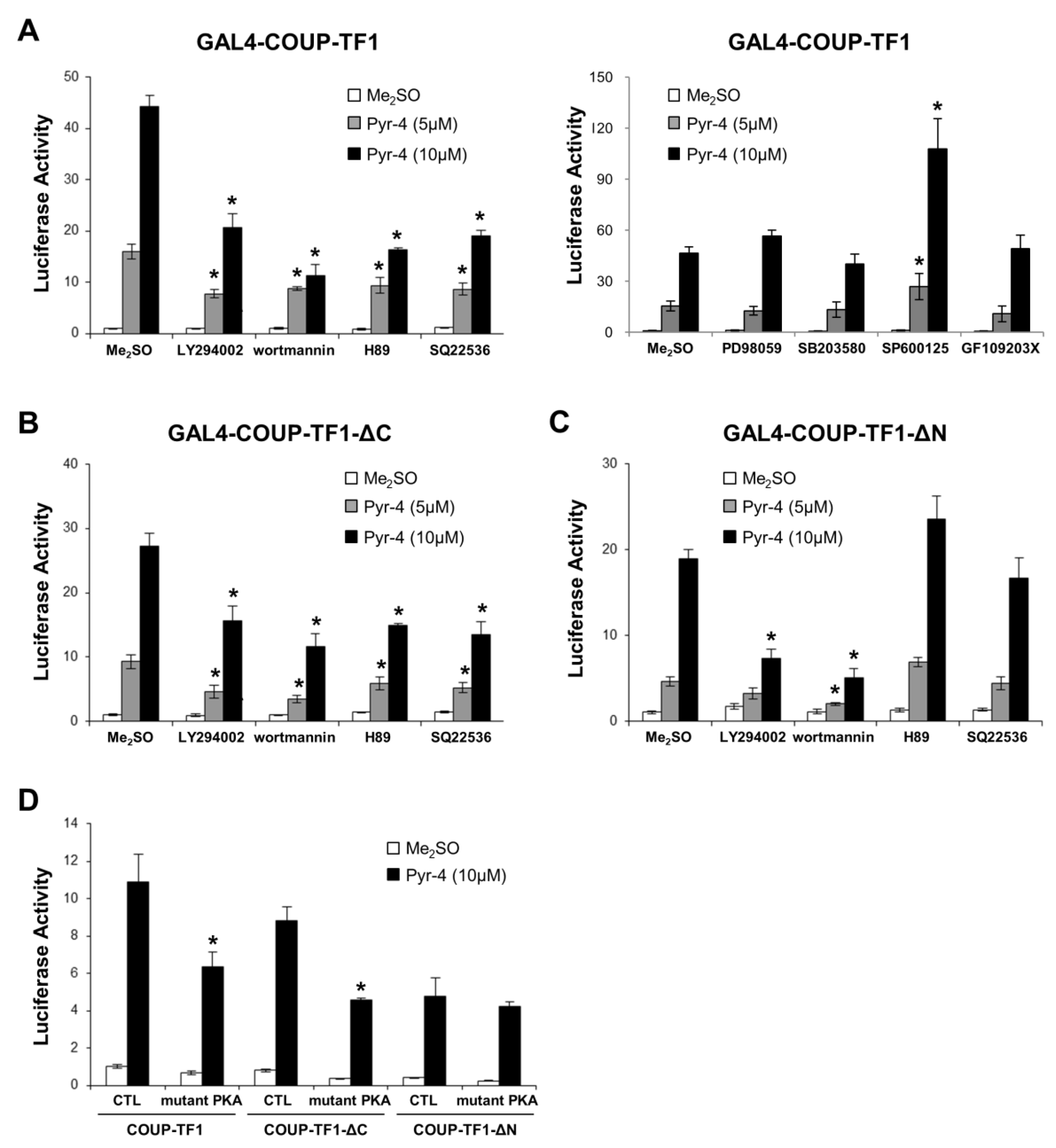
3.2. Activation of Variant GAL4-COUP-TFI Chimeras by DIM-C-Pyr-4 and Related Compounds
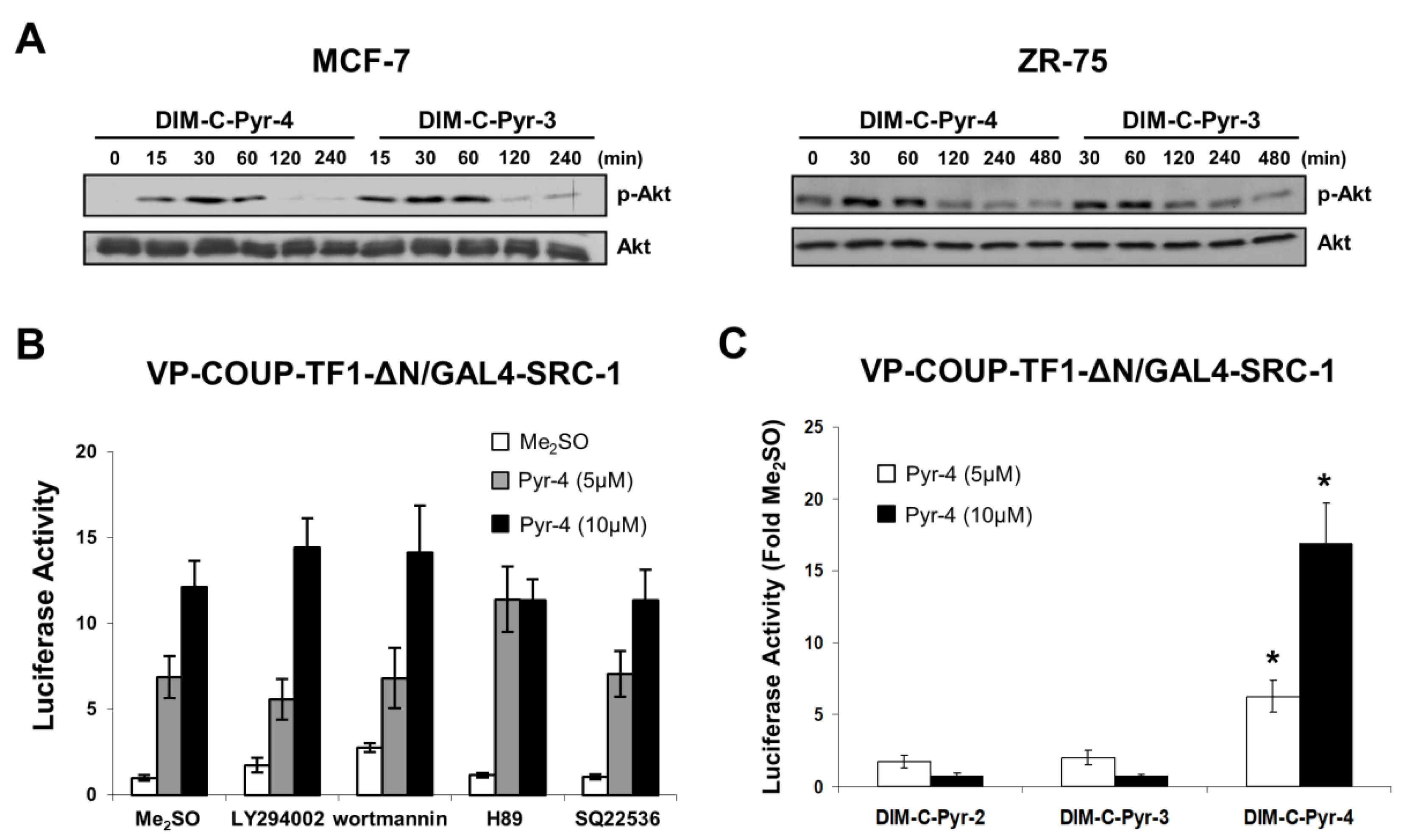
3.3. DIM-C-Pyr-4 Activation of Egr-1 Is Dependent on COUP-TFI and Sp Proteins
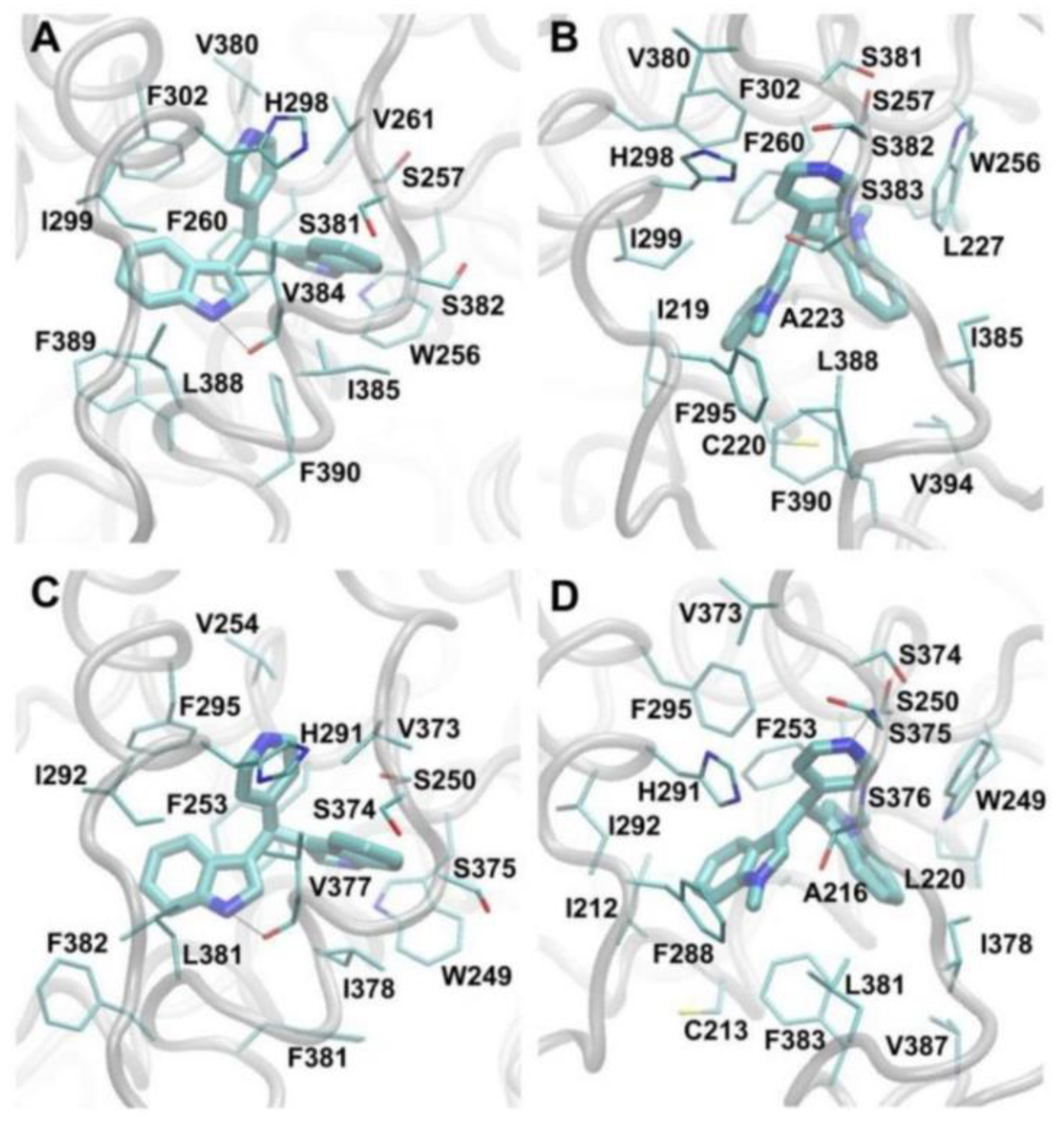
3.4. Molecular Modeling Studies
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ing, N.H.; Beekman, J.M.; Tsai, S.Y.; Tsai, M.J.; O’Malley, B.W. Members of the steroid hormone receptor superfamily interact with TFIIB (S300-II). J. Biol. Chem. 1992, 267, 17617–17623. [Google Scholar] [PubMed]
- Pastorcic, M.; Wang, H.; Elbrecht, A.; Tsai, S.Y.; Tsai, M.J.; O’Malley, B.W. Control of transcription initiation in vitro requires binding of a transcription factor to the distal promoter of the ovalbumin gene. Mol. Cell. Biol. 1986, 6, 2784–2791. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.H.; Tsai, S.Y.; Cook, R.G.; Beattie, W.G.; Tsai, M.J.; O’Malley, B.W. COUP transcription factor is a member of the steroid receptor superfamily. Nature 1989, 340, 163–166. [Google Scholar] [CrossRef] [PubMed]
- Ladias, J.A.; Hadzopoulou-Cladaras, M.; Kardassis, D.; Cardot, P.; Cheng, J.; Zannis, V.; Cladaras, C. Transcriptional regulation of human apolipoprotein genes ApoB, ApoCIII, and ApoAII by members of the steroid hormone receptor superfamily HNF-4, ARP-1, EAR-2, and EAR-3. J. Biol. Chem. 1992, 267, 15849–15860. [Google Scholar] [PubMed]
- Cooney, A.J.; Lee, C.T.; Lin, S.C.; Tsai, S.Y.; Tsai, M.J. Physiological function of the orphans GCNF and COUP-TF. Trends Endocrinol. Metab. 2001, 12, 247–251. [Google Scholar] [CrossRef]
- Tsai, S.Y.; Tsai, M.J. Chick ovalbumin upstream promoter-transcription factors (COUP-TFs): Coming of age. Endocr. Rev. 1997, 18, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dufau, M.L. Gene silencing by nuclear orphan receptors. Vitam. Horm. 2004, 68, 1–48. [Google Scholar] [PubMed]
- Cooney, A.J.; Tsai, S.Y.; O’Malley, B.W.; Tsai, M.J. Chicken ovalbumin upstream promoter transcription factor (COUP-TF) dimers bind to different GGTCA response elements, allowing COUP-TF to repress hormonal induction of the vitamin D3, thyroid hormone, and retinoic acid receptors. Mol. Cell. Biol. 1992, 12, 4153–4163. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, Y.; Toyoshima, K.; Yamamoto, T. Ear3/COUP-TF binds most tightly to a response element with tandem repeat separated by one nucleotide. Biochem. Biophys. Res. Commun. 1992, 183, 492–498. [Google Scholar] [CrossRef]
- Marcus, S.L.; Capone, J.P.; Rachubinski, R.A. Identification of COUP-TFII as a peroxisome proliferator response element binding factor using genetic selection in yeast: COUP-TFII activates transcription in yeast but antagonizes PPAR signaling in mammalian cells. Mol. Cell. Endocrinol. 1996, 120, 31–39. [Google Scholar] [CrossRef]
- Nishiyama, C.; Hi, R.; Osada, S.; Osumi, T. Functional interactions between nuclear receptors recognizing a common sequence element, the direct repeat motif spaced by one nucleotide (DR-1). J. Biochem. 1998, 123, 1174–1179. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, N.; Teng, C.T. COUP-TF acts as a competitive repressor for estrogen receptor-mediated activation of the mouse lactoferrin gene. Mol. Cell. Biol. 1993, 13, 1836–1846. [Google Scholar] [CrossRef] [PubMed]
- Mietus-Snyder, M.; Sladek, F.M.; Ginsburg, G.S.; Kuo, C.F.; Ladias, J.A.; Darnell, J.E., Jr.; Karathanasis, S.K. Antagonism between apolipoprotein AI regulatory protein 1, Ear3/COUP-TF, and hepatocyte nuclear factor 4 modulates apolipoprotein CIII gene expression in liver and intestinal cells. Mol. Cell. Biol. 1992, 12, 1708–1718. [Google Scholar] [CrossRef] [PubMed]
- Yanai, K.; Hirota, K.; Taniguchi-Yanai, K.; Shigematsu, Y.; Shimamoto, Y.; Saito, T.; Chowdhury, S.; Takiguchi, M.; Arakawa, M.; Nibu, Y.; et al. Regulated expression of human angiotensinogen gene by hepatocyte nuclear factor 4 and chicken ovalbumin upstream promoter-transcription factor. J. Biol. Chem. 1999, 274, 34605–34612. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Huang, X.; Sigmund, C.D. Identification of a nuclear orphan receptor (Ear2) as a negative regulator of renin gene transcription. Circ. Res. 2003, 92, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shushan, E.; Sharir, H.; Pikarsky, E.; Bergman, Y. A dynamic balance between ARP-1/COUP-TFII, EAR-3/COUP-TFI, and retinoic acid receptor:retinoid X receptor heterodimers regulates Oct-3/4 expression in embryonal carcinoma cells. Mol. Cell. Biol. 1995, 15, 1034–1048. [Google Scholar] [CrossRef] [PubMed]
- Stephanou, A.; Shah, M.; Richardson, B.; Handwerger, S. The ARP-1 orphan receptor represses steroid-mediated stimulation of human placental lactogen gene expression. J. Mol. Endocrinol. 1996, 16, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Tran, P.; Zhang, X.K.; Salbert, G.; Hermann, T.; Lehmann, J.M.; Pfahl, M. COUP orphan receptors are negative regulators of retinoic acid response pathways. Mol. Cell. Biol. 1992, 12, 4666–4676. [Google Scholar] [CrossRef] [PubMed]
- Wehrenberg, U.; Ivell, R.; Jansen, M.; von Goedecke, S.; Walther, N. Two orphan receptors binding to a common site are involved in the regulation of the oxytocin gene in the bovine ovary. Proc. Natl. Acad. Sci. USA 1994, 91, 1440–1444. [Google Scholar] [CrossRef] [PubMed]
- Xing, W.; Danilovich, N.; Sairam, M.R. Orphan receptor chicken ovalbumin upstream promoter transcription factors inhibit steroid factor-1, upstream stimulatory factor, and activator protein-1 activation of ovine follicle-stimulating hormone receptor expression via composite cis-elements. Biol. Reprod. 2002, 66, 1656–1666. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.K.; Hoffmann, B.; Tran, P.B.; Graupner, G.; Pfahl, M. Retinoid X receptor is an auxiliary protein for thyroid hormone and retinoic acid receptors. Nature 1992, 355, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Kliewer, S.A.; Umesono, K.; Heyman, R.A.; Mangelsdorf, D.J.; Dyck, J.A.; Evans, R.M. Retinoid X receptor-COUP-TF interactions modulate retinoic acid signaling. Proc. Natl. Acad. Sci. USA 1992, 89, 1448–1452. [Google Scholar] [CrossRef] [PubMed]
- Widom, R.L.; Rhee, M.; Karathanasis, S.K. Repression by ARP-1 sensitizes apolipoprotein AI gene responsiveness to RXR alpha and retinoic acid. Mol. Cell. Biol. 1992, 12, 3380–3389. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.J.; Parker, M.G. COUP-TF II homodimers are formed in preference to heterodimers with RXR alpha or TR beta in intact cells. Nucleic Acids Res. 1995, 23, 4143–4150. [Google Scholar] [CrossRef] [PubMed]
- Bailey, P.J.; Dowhan, D.H.; Franke, K.; Burke, L.J.; Downes, M.; Muscat, G.E. Transcriptional repression by COUP-TF II is dependent on the C-terminal domain and involves the N-CoR variant, RIP13delta1. J. Steroid Biochem. Mol. Biol. 1997, 63, 165–174. [Google Scholar] [CrossRef]
- Shibata, H.; Nawaz, Z.; Tsai, S.Y.; O’Malley, B.W.; Tsai, M.J. Gene silencing by chicken ovalbumin upstream promoter-transcription factor I (COUP-TFI) is mediated by transcriptional corepressors, nuclear receptor-corepressor (N-CoR) and silencing mediator for retinoic acid receptor and thyroid hormone receptor (SMRT). Mol. Endocrinol. 1997, 11, 714–724. [Google Scholar] [CrossRef] [PubMed]
- Pipaon, C.; Tsai, S.Y.; Tsai, M.J. COUP-TF upregulates NGFI-A gene expression through an Sp1 binding site. Mol. Cell. Biol. 1999, 19, 2734–2745. [Google Scholar] [CrossRef] [PubMed]
- Shibata, H.; Kobayashi, S.; Kurihara, I.; Suda, N.; Yokota, K.; Murai, A.; Ikeda, Y.; Saito, I.; Rainey, W.E.; Saruta, T. COUP-TF and transcriptional co-regulators in adrenal steroidogenesis. Endocr. Res. 2004, 30, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Wang, J.C.; Scott, D.K.; Granner, D.K. Transcription activation by the orphan nuclear receptor, chicken ovalbumin upstream promoter-transcription factor I (COUP-TFI). Definition of the domain involved in the glucocorticoid response of the phosphoenolpyruvate carboxykinase gene. J. Biol. Chem. 2000, 275, 3446–3454. [Google Scholar] [CrossRef] [PubMed]
- Grocock, C.A.; Charlton, H.M.; Pike, M.C. Role of the fetal pituitary in cryptorchidism induced by exogenous maternal oestrogen during pregnancy in mice. J. Reprod. Fertil. 1988, 83, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, I.; Shibata, H.; Kobayashi, S.; Suda, N.; Ikeda, Y.; Yokota, K.; Murai, A.; Saito, I.; Rainey, W.E.; Saruta, T. Ubc9 and Protein Inhibitor of Activated STAT 1 Activate Chicken Ovalbumin Upstream Promoter-Transcription Factor I-mediated Human CYP11B2 Gene Transcription. J. Biol. Chem. 2005, 280, 6721–6730. [Google Scholar] [CrossRef] [PubMed]
- Compe, E.; de Sousa, G.; Francois, K.; Roche, R.; Rahmani, R.; Torresani, J.; Raymondjean, M.; Planells, R. Spot 14 protein interacts and co-operates with chicken ovalbumin upstream promoter-transcription factor 1 in the transcription of the L-type pyruvate kinase gene through a specificity protein 1 (Sp1) binding site. Biochem. J. 2001, 358, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.J.; Horowitz, J.M.; Eling, T.E. Molecular cloning and characterization of human nonsteroidal anti-inflammatory drug-activated gene promoter. Basal transcription is mediated by Sp1 and Sp3. J. Biol. Chem. 2001, 276, 33384–33392. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dufau, M.L. Repression of the luteinizing hormone receptor gene promoter by cross talk among EAR3/COUP-TFI, Sp1/Sp3, and TFIIB. Mol. Cell. Biol. 2003, 23, 6958–6972. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Morrow, D.; Stewart, J.; Spencer, K.; Porter, W.; Smith, R., 3rd; Phillips, T.; Abdelrahim, M.; Samudio, I.; Safe, S. A new class of peroxisome proliferator-activated receptor gamma (PPARgamma) agonists that inhibit growth of breast cancer cells: 1,1-Bis(3′-indolyl)-1-(p-substituted phenyl)methanes. Mol. Cancer Ther. 2004, 3, 247–260. [Google Scholar] [PubMed]
- Hong, J.; Samudio, I.; Liu, S.; Abdelrahim, M.; Safe, S. Peroxisome proliferator-activated receptor gamma-dependent activation of p21 in Panc-28 pancreatic cancer cells involves Sp1 and Sp4 proteins. Endocrinology 2004, 145, 5774–5785. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.O.; Abdelrahim, M.; Yoon, K.; Chintharlapalli, S.; Papineni, S.; Kim, K.; Wang, H.; Safe, S. Inactivation of the orphan nuclear receptor TR3/Nur77 inhibits pancreatic cancer cell and tumor growth. Cancer Res. 2010, 70, 6824–6836. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.O.; Li, X.; Hedrick, E.; Jin, U.H.; Tjalkens, R.B.; Backos, D.S.; Li, L.; Zhang, Y.; Wu, Q.; Safe, S. Diindolylmethane analogs bind NR4A1 and are NR4A1 antagonists in colon cancer cells. Mol. Endocrinol. 2014, 28, 1729–1739. [Google Scholar] [CrossRef] [PubMed]
- Kruse, S.W.; Suino-Powell, K.; Zhou, X.E.; Kretschman, J.E.; Reynolds, R.; Vonrhein, C.; Xu, Y.; Wang, L.; Tsai, S.Y.; Tsai, M.J.; et al. Identification of COUP-TFII orphan nuclear receptor as a retinoic acid-activated receptor. PLoS Biol. 2008, 6, e227. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.; Sali, A. Comparative Protein Structure Modeling Using MODELLER. Curr. Protoc. Bioinform. 2016, 54. [Google Scholar] [CrossRef]
- Tamamis, P.; Morikis, D.; Floudas, C.A.; Archontis, G. Species specificity of the complement inhibitor compstatin investigated by all-atom molecular dynamics simulations. Proteins 2010, 78, 2655–2667. [Google Scholar] [CrossRef] [PubMed]
- Orr, A.A.; Wördehoff, M.M.; Hoyer, W.; Tamamis, P. Uncovering the Binding and Specificity of β-Wrapins for Amyloid-β and α-Synuclein. J. Phys. Chem. B 2016, 120, 12781–12794. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Jin, U.H.; Davidson, L.A.; Chapkin, R.S.; Jayaraman, A.; Tamamis, P.; Orr, A.; Allred, C.; Denison, M.S.; Soshilov, A.; et al. Editor’s Highlight: Microbial-Derived 1,4-Dihydroxy-2-naphthoic Acid and Related Compounds as Aryl Hydrocarbon Receptor Agonists/Antagonists: Structure-Activity Relationships and Receptor Modeling. Toxicol. Sci. 2017, 155, 458–473. [Google Scholar] [CrossRef] [PubMed]
- Orr, A.A.; Jayaraman, A.; Tamamis, P. Molecular Modeling of Chemoreceptor:Ligand Interactions. Methods Mol. Biol. 2018, 1729, 353–372. [Google Scholar] [PubMed]
- Jin, U.H.; Park, H.; Li, X.; Davidson, L.A.; Allred, C.; Patil, B.; Jayaprakasha, G.; Orr, A.A.; Mao, L.; Chapkin, R.S.; et al. Structure-Dependent Modulation of Aryl Hydrocarbon Receptor-Mediated Activities by Flavonoids. Toxicol. Sci. 2018, 164, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Orr, A.A.; Shaykhalishahi, H.; Mirecka, E.; Jonnalagadda, S.V.R.; Hoyer, W.; Tamamis, P. Elucidating Multi-Targeted Anti-Amyloid Activity and Enhanced Islet Amyloid Polypeptide Binding of β-wrapins. Comput. Chem. Eng. 2018, 116, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Orr, A.A.; Gonzalez-Rivera, J.C.; Wilson, M.; Bhikha, P.R.; Wang, D.; Contreras, L.M.; Tamamis, P. A high-throughput and rapid computational method for screening of RNA post-transcriptional modifications that can be recognized by target proteins. Methods 2018, 143, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Mohan, R.R.; Wilson, M.; Gorham, R.D., Jr.; Harrison, R.E.S.; Morikis, V.A.; Kieslich, C.A.; Orr, A.A.; Coley, A.V.; Tamamis, P.; Morikis, D. Virtual Screening of Chemical Compounds for Discovery of Complement C3 Ligands. ACS Omega 2018, 3, 6427–6438. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.R.; Brooks, C.L., 3rd; Mackerell, A.D., Jr.; Nilsson, L.; Petrella, R.J.; Roux, B.; Won, Y.; Archontis, G.; Bartels, C.; Boresch, S.; et al. CHARMM: The biomolecular simulation program. J. Comput. Chem. 2009, 30, 1545–1614. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; MacKerell, A.D., Jr. CHARMM36 all-atom additive protein force field: Validation based on comparison to NMR data. J. Comput. Chem. 2013, 34, 2135–2145. [Google Scholar] [CrossRef] [PubMed]
- Krivov, G.G.; Shapovalov, M.V.; Dunbrack, R.L., Jr. Improved prediction of protein side-chain conformations with SCWRL4. Proteins 2009, 77, 778–795. [Google Scholar] [CrossRef] [PubMed]
- Lannigan, D.A. Estrogen receptor phosphorylation. Steroids 2003, 68, 1–9. [Google Scholar] [CrossRef]
- Weigel, N.L. Steroid hormone receptors and their regulation by phosphorylation. Biochem. J. 1996, 319 Pt 3, 657–667. [Google Scholar] [CrossRef]
- Gianni, M.; Bauer, A.; Garattini, E.; Chambon, P.; Rochette-Egly, C. Phosphorylation by p38MAPK and recruitment of SUG-1 are required for RA-induced RAR gamma degradation and transactivation. EMBO J. 2002, 21, 3760–3769. [Google Scholar] [CrossRef] [PubMed]
- Gianni, M.; Kopf, E.; Bastien, J.; Oulad-Abdelghani, M.; Garattini, E.; Chambon, P.; Rochette-Egly, C. Down-regulation of the phosphatidylinositol 3-kinase/Akt pathway is involved in retinoic acid-induced phosphorylation, degradation, and transcriptional activity of retinoic acid receptor gamma 2. J. Biol. Chem. 2002, 277, 24859–24862. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.C.; Smith, C.L.; O’Malley, B.W. Transcriptional regulation by steroid receptor coactivator phosphorylation. Endocr. Rev. 2005, 26, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Safe, S.; Kim, K. Nuclear receptor-mediated transactivation through interaction with Sp proteins. Prog. Nucleic Acid Res. Mol. Biol. 2004, 77, 1–36. [Google Scholar] [PubMed]
- Higgins, K.J.; Liu, S.; Abdelrahim, M.; Yoon, K.; Vanderlaag, K.; Porter, W.; Metz, R.P.; Safe, S. Vascular endothelial growth factor receptor-2 expression is induced by 17beta-estradiol in ZR-75 breast cancer cells by estrogen receptor alpha/Sp proteins. Endocrinology 2006, 147, 3285–3295. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Cooney, A.J.; Kuratani, S.; DeMayo, F.J.; Tsai, S.Y.; Tsai, M.J. Spatiotemporal expression patterns of chicken ovalbumin upstream promoter-transcription factors in the developing mouse central nervous system: Evidence for a role in segmental patterning of the diencephalon. Proc. Natl. Acad. Sci. USA 1994, 91, 4451–4455. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Pereira, F.A.; DeMayo, F.J.; Lydon, J.P.; Tsai, S.Y.; Tsai, M.J. Null mutation of mCOUP-TFI results in defects in morphogenesis of the glossopharyngeal ganglion, axonal projection, and arborization. Genes Dev. 1997, 11, 1925–1937. [Google Scholar] [CrossRef] [PubMed]
- Pereira, F.A.; Qiu, Y.; Zhou, G.; Tsai, M.J.; Tsai, S.Y. The orphan nuclear receptor COUP-TFII is required for angiogenesis and heart development. Genes Dev. 1999, 13, 1037–1049. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, H.; Zhou, C.; Lin, S.C.; Durand, B.; Tsai, S.Y.; Tsai, M.J. The nuclear orphan receptor COUP-TFI is important for differentiation of oligodendrocytes. Dev. Biol. 2004, 266, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Borgen, E.; Rypdal, M.C.; Sosa, M.S.; Renolen, A.; Schlichting, E.; Lonning, P.E.; Synnestvedt, M.; Aguirre-Ghiso, J.A.; Naume, B. NR2F1 stratifies dormant disseminated tumor cells in breast cancer patients. Breast Cancer Res. 2018, 20, 120. [Google Scholar] [CrossRef] [PubMed]
- Klinge, C.M.; Silver, B.F.; Driscoll, M.D.; Sathya, G.; Bambara, R.A.; Hilf, R. Chicken ovalbumin upstream promoter-transcription factor interacts with estrogen receptor, binds to estrogen response elements and half-sites, and inhibits estrogen-induced gene expression. J. Biol. Chem. 1997, 272, 31465–31474. [Google Scholar] [CrossRef] [PubMed]
- Le Dily, F.; Metivier, R.; Gueguen, M.M.; Le Peron, C.; Flouriot, G.; Tas, P.; Pakdel, F. COUP-TFI modulates estrogen signaling and influences proliferation, survival and migration of breast cancer cells. Breast Cancer Res. Treat. 2008, 110, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Lemon, B.D.; Freedman, L.P. Nuclear receptor cofactors as chromatin remodelers. Curr. Opin. Genet. Dev. 1999, 9, 499–504. [Google Scholar] [CrossRef]
- Klinge, C.M. Estrogen receptor interaction with co-activators and co-repressors. Steroids 2000, 65, 227–251. [Google Scholar] [CrossRef]
- Smith, C.L.; O’Malley, B.W. Coregulator function: A key to understanding tissue specificity of selective receptor modulators. Endocr. Rev. 2004, 25, 45–71. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.P. Coregulatory proteins in nuclear hormone receptor action. Vitam. Horm. 1999, 55, 165–218. [Google Scholar] [PubMed]
- Naar, A.M.; Lemon, B.D.; Tjian, R. Transcriptional coactivator complexes. Annu. Rev. Biochem. 2001, 70, 475–501. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, M.G.; Glass, C.K. Coregulator codes of transcriptional regulation by nuclear receptors. J. Biol. Chem. 2001, 276, 36865–36868. [Google Scholar] [CrossRef] [PubMed]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, K.; Chen, C.-C.; Orr, A.A.; Barreto, P.N.; Tamamis, P.; Safe, S. Activation of COUP-TFI by a Novel Diindolylmethane Derivative. Cells 2019, 8, 220. https://doi.org/10.3390/cells8030220
Yoon K, Chen C-C, Orr AA, Barreto PN, Tamamis P, Safe S. Activation of COUP-TFI by a Novel Diindolylmethane Derivative. Cells. 2019; 8(3):220. https://doi.org/10.3390/cells8030220
Chicago/Turabian StyleYoon, Kyungsil, Chien-Cheng Chen, Asuka A. Orr, Patricia N. Barreto, Phanourios Tamamis, and Stephen Safe. 2019. "Activation of COUP-TFI by a Novel Diindolylmethane Derivative" Cells 8, no. 3: 220. https://doi.org/10.3390/cells8030220
APA StyleYoon, K., Chen, C.-C., Orr, A. A., Barreto, P. N., Tamamis, P., & Safe, S. (2019). Activation of COUP-TFI by a Novel Diindolylmethane Derivative. Cells, 8(3), 220. https://doi.org/10.3390/cells8030220