Adenosine A1-A2A Receptor-Receptor Interaction: Contribution to Guanosine-Mediated Effects

 ,
,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. OGD Protocol
2.4. Cellular Viability Evaluation
2.5. Measurement of ROS Production
2.6. Plasmid Constructs
2.7. Cell Culture and Transfection
2.8. NanoBRET Experiments
2.9. cAMP Assay
2.10. Intracellular Calcium Determinations
2.11. Label-Free Cellular Impedance Assay
2.12. Statistics
3. Results
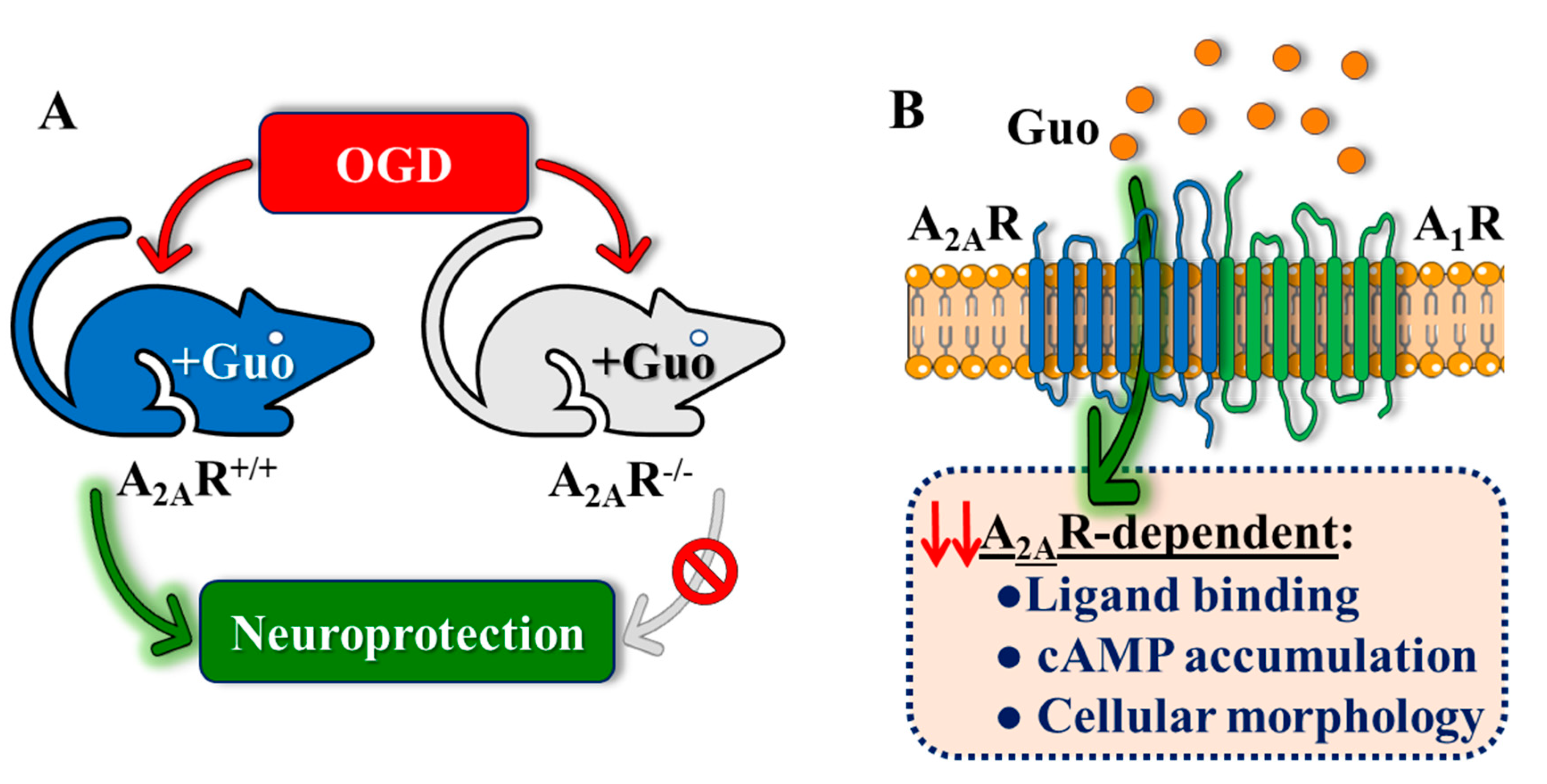
3.1. Guanosine-Mediated Neuroprotection in Hippocampal Slices Depends on A2AR Expression
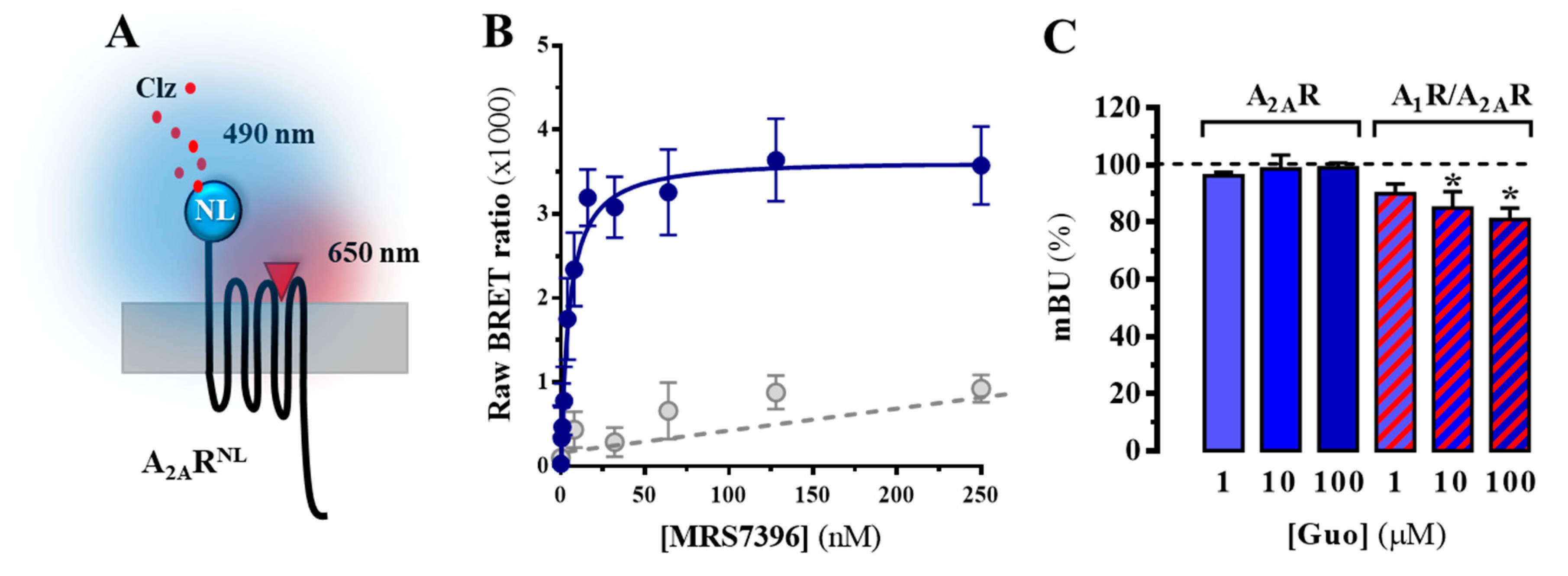
3.2. A2AR Ligand Binding is Affected by Guanosine upon A1R Coexpression
3.3. A2AR Signalling, but Not A1R, is Modulated by Guanosine in an A1R Coexpression-Dependent Manner
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Materials and Methods
Appendix A.1.1. Immunocytochemistry
Appendix A.1.2. BRET
Appendix A.2. Results

References
- Lanznaster, D.; Dal-Cim, T.; Piermartiri, T.C.B.; Tasca, C.I. Guanosine: A Neuromodulator with Therapeutic Potential in Brain Disorders. Aging Dis. 2016, 7, 657–679. [Google Scholar] [CrossRef] [PubMed]
- Di Liberto, V.; Mudò, G.; Garozzo, R.; Frinchi, M.; Fernandez-Dueñas, V.; Di Iorio, P.; Ciccarelli, R.; Caciagli, F.; Condorelli, D.F.; Ciruela, F.; et al. The guanine-based purinergic system: The tale of an orphan neuromodulation. Front. Pharmacol. 2016, 7, 158. [Google Scholar] [CrossRef] [PubMed]
- Durukan, A.; Tatlisumak, T. Acute ischemic stroke: Overview of major experimental rodent models, pathophysiology, and therapy of focal cerebral ischemia. Pharmacol. Biochem. Behav. 2007, 87, 179–197. [Google Scholar] [CrossRef] [PubMed]
- Candelario-Jalil, E. Injury and repair mechanisms in ischemic stroke: Considerations for the development of novel neurotherapeutics. Current Opin. Investig. Drugs (Lond. Engl. 2000) 2009, 10, 644–654. [Google Scholar]
- Dal-Cim, T.; Martins, W.C.; Santos, A.R.S.; Tasca, C.I. Guanosine is neuroprotective against oxygen/glucose deprivation in hippocampal slices via large conductance Ca2+-activated K+ channels, phosphatidilinositol-3 kinase/protein kinase B pathway activation and glutamate uptake. Neuroscience 2011, 183, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Dal-Cim, T.; Molz, S.; Egea, J.; Parada, E.; Romero, A.; Budni, J.; Martín de Saavedra, M.D.; del Barrio, L.; Tasca, C.I.; López, M.G. Guanosine protects human neuroblastoma SH-SY5Y cells against mitochondrial oxidative stress by inducing heme oxigenase-1 via PI3K/Akt/GSK-3β pathway. Neurochem. Int. 2012, 61, 397–404. [Google Scholar] [CrossRef]
- Ganzella, M.; de Oliveira, E.D.A.; Comassetto, D.D.; Cechetti, F.; Cereser, V.H.; Moreira, J.D.; Hansel, G.; Almeida, R.F.; Ramos, D.B.; Figueredo, Y.N.; et al. Effects of chronic guanosine treatment on hippocampal damage and cognitive impairment of rats submitted to chronic cerebral hypoperfusion. Neurol. Sci. 2012, 33, 985–997. [Google Scholar] [CrossRef]
- Tasca, C.; Llorente, J.; Dal-Cim, T.; Fernandez-Duenas, V.; Gomez-Soler, M.; Gandia, J.; Ciruela, F. The neuroprotective agent Guanosine activates big conductance Ca2+-activated Potassium channels (BK) transfected to HEK-293 cells. J. Neurochem. 2013, 125, 273. [Google Scholar]
- Hansel, G.; Tonon, A.C.; Guella, F.L.; Pettenuzzo, L.F.; Duarte, T.; Duarte, M.M.M.F.; Oses, J.P.; Achaval, M.; Souza, D.O. Guanosine Protects Against Cortical Focal Ischemia. Involvement of Inflammatory Response. Mol. Neurobiol. 2015, 52, 1791–1803. [Google Scholar] [CrossRef]
- Dal-Cim, T.; Martins, W.C.; Thomaz, D.T.; Coelho, V.; Poluceno, G.G.; Lanznaster, D.; Vandresen-Filho, S.; Tasca, C.I. Neuroprotection Promoted by Guanosine Depends on Glutamine Synthetase and Glutamate Transporters Activity in Hippocampal Slices Subjected to Oxygen/Glucose Deprivation. Neurotox. Res. 2016, 29, 460–468. [Google Scholar] [CrossRef]
- Dal-Cim, T.; Ludka, F.K.; Martins, W.C.; Reginato, C.; Parada, E.; Egea, J.; López, M.G.; Tasca, C.I. Guanosine controls inflammatory pathways to afford neuroprotection of hippocampal slices under oxygen and glucose deprivation conditions. J. Neurochem. 2013, 126, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Traversa, U.; Bombi, G.; Di Iorio, P.; Ciccarelli, R.; Werstiuk, E.S.; Rathbone, M.P. Specific [(3)H]-guanosine binding sites in rat brain membranes. Br. J. Pharmacol. 2002, 135, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Traversa, U.; Bombi, G.; Camaioni, E.; Macchiarulo, A.; Costantino, G.; Palmieri, C.; Caciagli, F.; Pellicciari, R. Rat brain guanosine binding site. Biological studies and pseudo-receptor construction. Bioorganic Med. Chem. 2003, 11, 5417–5425. [Google Scholar] [CrossRef] [PubMed]
- Volpini, R.; Marucci, G.; Buccioni, M.; Dal Ben, D.; Lambertucci, C.; Lammi, C.; Mishra, R.C.; Thomas, A.; Cristalli, G. Evidence for the existence of a specific g protein-coupled receptor activated by guanosine. ChemMedChem 2011, 6, 1074–1080. [Google Scholar] [CrossRef] [PubMed]
- Dal-Cim, T.; Poluceno, G.G.; Lanznaster, D.; de Oliveira, K.A.; Nedel, C.B.; Tasca, C.I. Guanosine prevents oxidative damage and glutamate uptake impairment induced by oxygen/glucose deprivation in cortical astrocyte cultures: Involvement of A1 and A2A adenosine receptors and PI3K, MEK, and PKC pathways. Purinergic Signal. 2019, in press. [Google Scholar] [CrossRef]
- Ciruela, F. Guanosine behind the scene. J. Neurochem. 2013, 126, 425–427. [Google Scholar] [CrossRef]
- Ciruela, F.; Casadó, V.; Rodrigues, R.J.; Luján, R.; Burgueño, J.; Canals, M.; Borycz, J.; Rebola, N.; Goldberg, S.R.; Mallol, J.; et al. Presynaptic control of striatal glutamatergic neurotransmission by adenosine A1-A2Areceptor heteromers. J. Neurosci. 2006, 26, 2080–2087. [Google Scholar] [CrossRef]
- Ciruela, F.; Ferre, S.; Casado, V.; Cortes, A.; Cunha, R.A.; Lluis, C.; Franco, R. Heterodimeric adenosine receptors: A device to regulate neurotransmitter release. Cell. Mol. Life Sci. 2006, 63, 2427–2431. [Google Scholar] [CrossRef]
- Duroux, R.; Ciancetta, A.; Mannes, P.; Yu, J.; Boyapati, S.; Gizewski, E.; Yous, S.; Ciruela, F.; Auchampach, J.A.; Gao, Z.-G.; et al. Bitopic fluorescent antagonists of the A2A adenosine receptor based on pyrazolo[4,3-: E] [1,2,4]triazolo[1,5- c] pyrimidin-5-amine functionalized congeners. MedChemComm 2017, 8, 1659–1667. [Google Scholar] [CrossRef]
- Ledent, C.; Vaugeois, J.M.; Schiffmann, S.N.; Pedrazzini, T.; El Yacoubi, M.; Vanderhaeghen, J.J.; Costentin, J.; Heath, J.K.; Vassart, G.; Parmentier, M. Aggressiveness, hypoalgesia and high blood pressure in mice lacking the adenosine A2a receptor. Nature 1997, 388, 674–678. [Google Scholar] [CrossRef]
- Clark, J.D.; Gebhart, G.F.; Gonder, J.C.; Keeling, M.E.; Kohn, D.F. Special report: The 1996 guide for the care and use of laboratory animals. Ilar J. /Natl. Res. Counc. Inst. Lab. Anim. Resour. 1997, 38, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Peterson, D.A.; Kimura, H.; Schubert, D. Mechanism of cellular 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) reduction. J. Neurochem. 1997, 69, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.G.K.; da Cunha, A.A.; Machado, F.R.; Pederzolli, C.D.; Dalazen, G.R.; de Assis, A.M.; Lamers, M.L.; dos Santos, M.F.; Dutra-Filho, C.S.; Wyse, A.T.S. Experimental hyperprolinemia induces mild oxidative stress, metabolic changes, and tissue adaptation in rat liver. J. Cell. Biochem. 2012, 113, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Duenas, V.; Gomez-Soler, M.; Jacobson, K.A.; Kumar, S.T.; Fuxe, K.; Borroto-Escuela, D.O.; Ciruela, F. Molecular determinants of A(2A) R-D(2) R allosterism: Role of the intracellular loop 3 of the D(2) R. J. Neurochem. 2012, 123, 373–384. [Google Scholar] [CrossRef]
- Stoddart, L.A.; Johnstone, E.K.M.; Wheal, A.J.; Goulding, J.; Robers, M.B.; Machleidt, T.; Wood, K.V.; Hill, S.J.; Pfleger, K.D.G. Application of BRET to monitor ligand binding to GPCRs. Nat. Methods 2015, 12, 661–663. [Google Scholar] [CrossRef]
- Taura, J.; Fernández-Dueñas, V.; Ciruela, F. Determination of GPCR-mediated cAMP accumulation in rat striatal synaptosomes. Neuromethods 2016, 110, 455–464. [Google Scholar]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Tarakanov, A.O.; Ciruela, F.; Agnati, L.F.; Fuxe, K. On the existence of a possible A2A-D2-beta-Arrestin2 complex: A2A agonist modulation of D2 agonist-induced beta-arrestin2 recruitment. J. Mol. Biol. 2011, 406, 687–699. [Google Scholar] [CrossRef]
- Xu, Y.; Xie, X.; Duan, Y.; Wang, L.; Cheng, Z.; Cheng, J. A review of impedance measurements of whole cells. Biosens. Bioelectron. 2016, 77, 824–836. [Google Scholar] [CrossRef]
- Hillger, J.M.; Schoop, J.; Boomsma, D.I.; Slagboom, P.E.; IJzerman, A.P.; Heitman, L.H. Whole-cell biosensor for label-free detection of GPCR-mediated drug responses in personal cell lines. Biosens. Bioelectron. 2015, 74, 233–242. [Google Scholar] [CrossRef]
- Stallaert, W.; Dorn, J.F.; van der Westhuizen, E.; Audet, M.; Bouvier, M. Impedance responses reveal β₂-adrenergic receptor signaling pluridimensionality and allow classification of ligands with distinct signaling profiles. PLoS ONE 2012, 7, e29420. [Google Scholar] [CrossRef]
- Fernández-Dueñas, V.; Taura, J.J.; Cottet, M.; Gómez-Soler, M.; López-Cano, M.; Ledent, C.; Watanabe, M.; Trinquet, E.; Pin, J.-P.; Luján, R.; et al. Untangling dopamine-adenosine receptor-receptor assembly in experimental parkinsonism in rats. Dis. Models Mech. 2015, 8, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Thomaz, D.T.; Dal-Cim, T.A.; Martins, W.C.; Cunha, M.P.; Lanznaster, D.; de Bem, A.F.; Tasca, C.I. Guanosine prevents nitroxidative stress and recovers mitochondrial membrane potential disruption in hippocampal slices subjected to oxygen/glucose deprivation. Purinergic Signal. 2016, 12, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Tasca, C.I.; Lanznaster, D.; Oliveira, K.A.; Fernández-Dueñas, V.; Ciruela, F. Neuromodulatory effects of guanine-based purines in health and disease. Front. Cell. Neurosci. 2018, 12, 376. [Google Scholar] [CrossRef] [PubMed]
- Núñez, F.; Taura, J.; Camacho, J.; López-Cano, M.; Fernández-Dueñas, V.; Castro, N.; Castro, J.; Ciruela, F. PBF509, an Adenosine A2A Receptor Antagonist with Efficacy in Rodent Models of Movement Disorders. Front. Pharmacol. 2018, 9, 1200. [Google Scholar] [CrossRef]
- Almeida, R.F.; Comasseto, D.D.; Ramos, D.B.; Hansel, G.; Zimmer, E.R.; Loureiro, S.O.; Ganzella, M.; Souza, D.O. Guanosine Anxiolytic-Like Effect Involves Adenosinergic and Glutamatergic Neurotransmitter Systems. Mol. Neurobiol. 2017, 54, 423–436. [Google Scholar] [CrossRef]
- Dobrachinski, F.; Gerbatin, R.R.; Sartori, G.; Golombieski, R.M.; Antoniazzi, A.; Nogueira, C.W.; Royes, L.F.; Fighera, M.R.; Porciúncula, L.O.; Cunha, R.A.; et al. Guanosine Attenuates Behavioral Deficits After Traumatic Brain Injury by Modulation of Adenosinergic Receptors. Mol. Neurobiol. 2019, 56, 3145–3158. [Google Scholar] [CrossRef]
- Ciruela, F.; Gómez-Soler, M.; Guidolin, D.; Borroto-Escuela, D.O.; Agnati, L.F.; Fuxe, K.; Fernández-Dueñas, V. Adenosine receptor containing oligomers: Their role in the control of dopamine and glutamate neurotransmission in the brain. Biochim. Biophys. Acta Biomembr. 2011, 1808, 1245–1255. [Google Scholar] [CrossRef]
- Ferré, S.; Casadó, V.; Devi, L.A.; Filizola, M.; Jockers, R.; Lohse, M.J.; Milligan, G.; Pin, J.-P.; Guitart, X. G Protein–Coupled Receptor Oligomerization Revisited: Functional and Pharmacological Perspectives. Pharmacol. Rev. 2014, 66, 413–434. [Google Scholar] [CrossRef]
- Orru, M.; Bakešová, J.; Brugarolas, M.; Quiroz, C.; Beaumont, V.; Goldberg, S.R.; Lluís, C.; Cortés, A.; Franco, R.; Casadó, V.; et al. Striatal pre- and postsynaptic profile of adenosine A2A receptor antagonists. PLoS ONE 2011, 6, e16088. [Google Scholar] [CrossRef]
- Navarro, G.; Cordomí, A.; Brugarolas, M.; Moreno, E.; Aguinaga, D.; Pérez-Benito, L.; Ferre, S.; Cortés, A.; Casadó, V.; Mallol, J.; et al. Cross-communication between G i and G s in a G-protein-coupled receptor heterotetramer guided by a receptor C-terminal domain. BMC Biol. 2018, 16, 24. [Google Scholar] [CrossRef]
- Ciruela, F.; Fernández-Dueñas, V. GPCR oligomerization analysis by means of BRET and dFRAP. In G Protein-Coupled Receptor Screening Assays; Humana Press: New York, NY, USA, 2015. [Google Scholar]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lanznaster, D.; Massari, C.M.; Marková, V.; Šimková, T.; Duroux, R.; Jacobson, K.A.; Fernández-Dueñas, V.; Tasca, C.I.; Ciruela, F. Adenosine A1-A2A Receptor-Receptor Interaction: Contribution to Guanosine-Mediated Effects. Cells 2019, 8, 1630. https://doi.org/10.3390/cells8121630
Lanznaster D, Massari CM, Marková V, Šimková T, Duroux R, Jacobson KA, Fernández-Dueñas V, Tasca CI, Ciruela F. Adenosine A1-A2A Receptor-Receptor Interaction: Contribution to Guanosine-Mediated Effects. Cells. 2019; 8(12):1630. https://doi.org/10.3390/cells8121630
Chicago/Turabian StyleLanznaster, Débora, Caio M. Massari, Vendula Marková, Tereza Šimková, Romain Duroux, Kenneth A. Jacobson, Víctor Fernández-Dueñas, Carla I. Tasca, and Francisco Ciruela. 2019. "Adenosine A1-A2A Receptor-Receptor Interaction: Contribution to Guanosine-Mediated Effects" Cells 8, no. 12: 1630. https://doi.org/10.3390/cells8121630
APA StyleLanznaster, D., Massari, C. M., Marková, V., Šimková, T., Duroux, R., Jacobson, K. A., Fernández-Dueñas, V., Tasca, C. I., & Ciruela, F. (2019). Adenosine A1-A2A Receptor-Receptor Interaction: Contribution to Guanosine-Mediated Effects. Cells, 8(12), 1630. https://doi.org/10.3390/cells8121630