The Expression of miRNAs in Human Ovaries, Oocytes, Extracellular Vesicles, and Early Embryos: A Systematic Review

 ,
, 

Abstract
1. Introduction
2. Material and Methods
2.1. Search Strategy and Systematic Review Registration
2.2. Study Selection and Eligibility
2.3. Data Extraction and Quality Assessment
3. Results
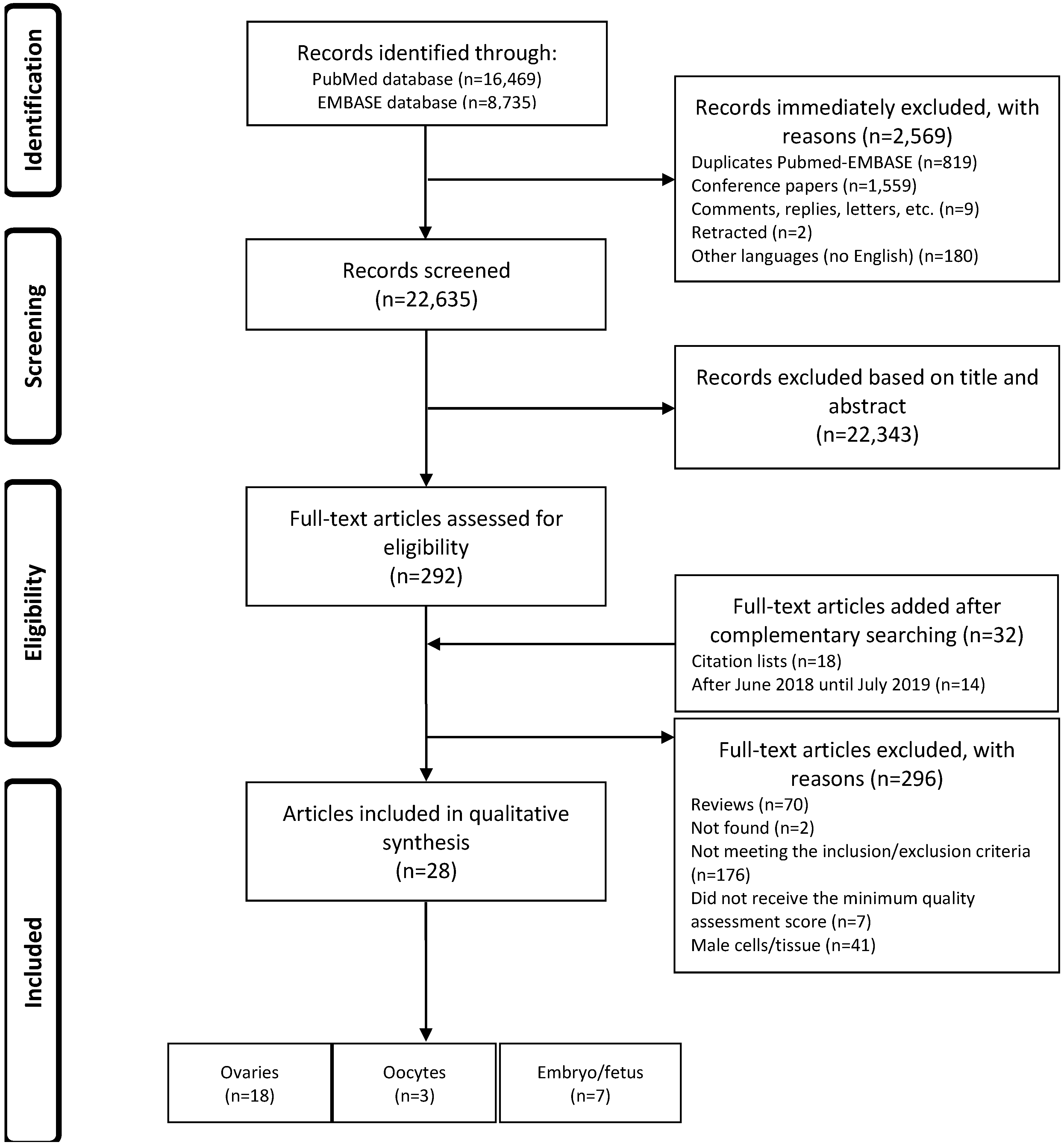
3.1. Identification and Selection of the Articles
3.2. Summary of Selected Studies
3.2.1. Ovaries and/or Extracellular Vesicles (Exosomes and Microvesicles)
3.2.2. Oocytes
3.2.3. Embryos
4. Discussion
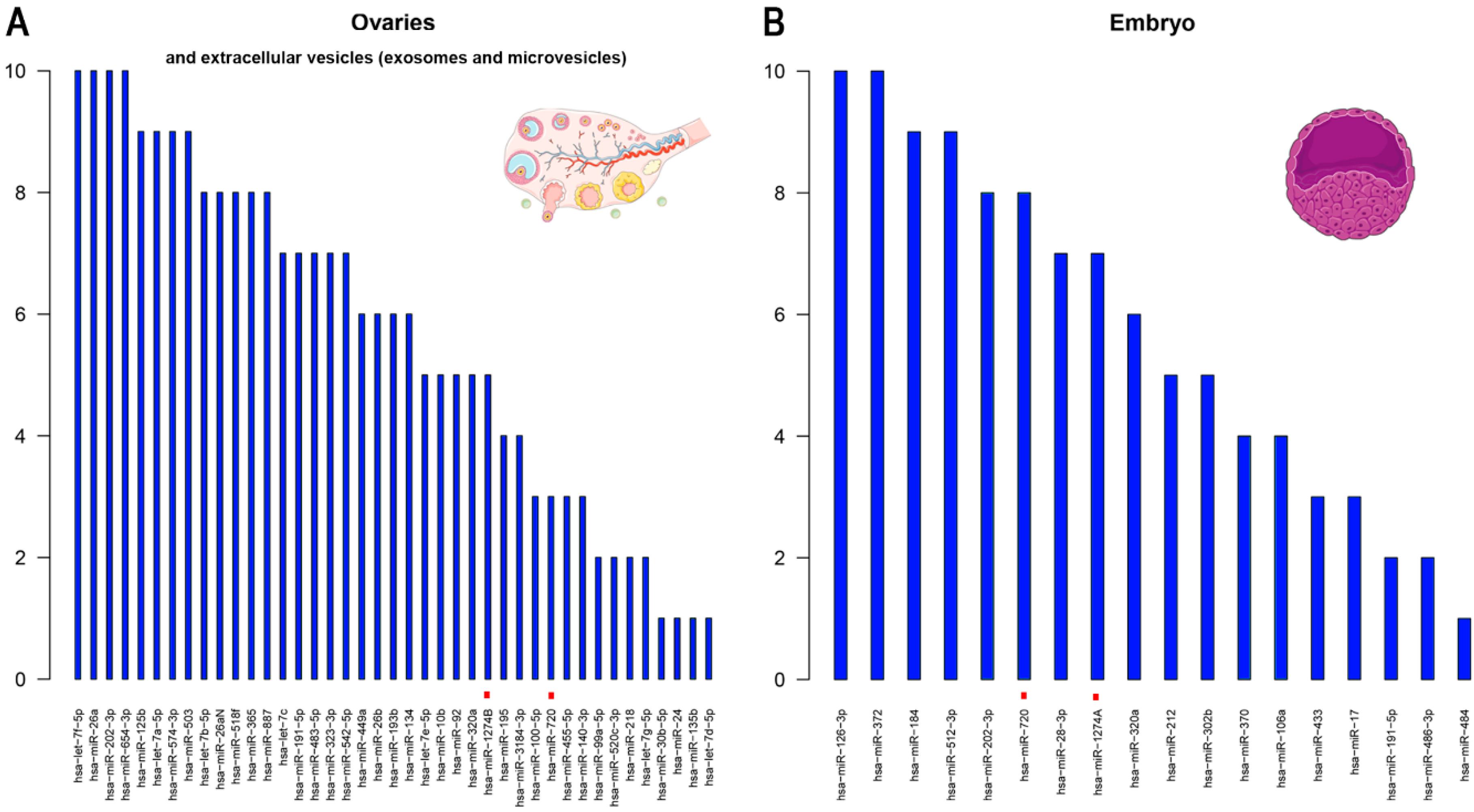
4.1. Highly Expressed miRNAs in Female Reproductive Cells/Tissues and Embryos
4.2. miRNA Deregulations as a Risk of Infertility or Embryo Development Errors
4.3. Looking Beyond the Present Systematic Review
4.4. Concerns, Limitations and Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Prospero Registration Number
References
- Ambros, V. microRNAs: Tiny regulators with great potential. Cell 2001, 107, 823–826. [Google Scholar] [CrossRef]
- Pasquinelli, A.E. MicroRNAs and their targets: Recognition, regulation and an emerging reciprocal relationship. Nat. Rev. Genet. 2012, 13, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Boyd, S.D. Everything you wanted to know about small RNA but were afraid to ask. Lab. Investig. 2008, 88, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Inui, M.; Martello, G.; Piccolo, S. MicroRNA control of signal transduction. Nat. Rev. Mol. Cell Biol. 2010, 11, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Tétreault, N.; De Guire, V. miRNAs: Their discovery, biogenesis and mechanism of action. Clin. Biochem. 2013, 46, 842–845. [Google Scholar] [CrossRef] [PubMed]
- Alles, J.; Fehlmann, T.; Fischer, U.; Backes, C.; Galata, V.; Minet, M.; Hart, M.; Abu-halima, M.; Lenhof, H.; Keller, A.; et al. An estimate of the total number of true human miRNAs. Nucleic Acids Res. 2019, 47, 3353–3364. [Google Scholar] [CrossRef]
- Ostermeier, G.C.; Dix, D.J.; Miller, D.; Khatri, P.; Krawetz, S.A. Spermatozoal RNA profiles of normal fertile men. Lancet 2002, 360, 772–777. [Google Scholar] [CrossRef]
- Ostermeier, G.C.; Goodrich, R.J.; Moldenhauer, J.S.; Diamond, M.P.; Krawetz, S. A suite of novel human spermatozoal RNAs. J. Androl. 2005, 26, 70–74. [Google Scholar]
- Liang, Y.; Ridzon, D.; Wong, L.; Chen, C. Characterization of microRNA expression profiles in normal human tissues. BMC Genom. 2007, 8, 166. [Google Scholar] [CrossRef]
- Barad, O.; Meiri, E.; Avniel, A.; Aharonov, R.; Barzilai, A.; Bentwich, I.; Einav, U.; Gilad, S.; Hurban, P.; Karov, Y.; et al. MicroRNA expression detected by oligonucleotide microarrays: System establishment and expression profiling in human tissues. Genome Res. 2004, 14, 2486–2494. [Google Scholar] [CrossRef]
- Yadav, R.P.; Kotaja, N. Small RNAs in spermatogenesis. Mol. Cell. Endocrinol. 2013, 382, 498–508. [Google Scholar] [CrossRef] [PubMed]
- Toloubeydokhti, T.; Bukulmez, O.; Chegini, N. Potential Regulatory Functions of MicroRNA in the Ovary. Endocrinology 2009, 26, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Galliano, D.; Pellicer, A. MicroRNA and implantation. Fertil. Steril. 2014, 101, 1531–1544. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Garcia, I.; Miska, E. MicroRNA functions in animal development and human disease. Development 2005, 132, 4653–4662. [Google Scholar] [CrossRef] [PubMed]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Goetzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. J. Clin. Epidemiol. 2009, 62, e1000100. [Google Scholar] [CrossRef]
- Cai, G.; Ma, X.; Chen, B.; Huang, Y.; Liu, S.; Yang, H.; Zou, W. MicroRNA-145 Negatively Regulates Cell Proliferation Through Targeting IRS1 in Isolated Ovarian Granulosa Cells from Patients with Polycystic Ovary Syndrome. Reprod. Sci. 2017, 24, 902–910. [Google Scholar] [CrossRef]
- Roth, L.W.; McCallie, B.; Alvero, R.; Schoolcraft, W.B.; Minjarez, D.; Katz-Jaffe, M.G. Altered microRNA and gene expression in the follicular fluid of women with polycystic ovary syndrome. J. Assist. Reprod. Genet. 2014, 31, 355–362. [Google Scholar] [CrossRef]
- Scalici, E.; Traver, S.; Mullet, T.; Molinari, N.; Ferrières, A.; Brunet, C.; Belloc, S.; Hamamah, S. Circulating microRNAs in follicular fluid, powerful tools to explore in vitro fertilization process. Sci. Rep. 2016, 6, 24976. [Google Scholar] [CrossRef]
- Shi, L.; Liu, S.; Zhao, W.; Shi, J. miR-483-5p and miR-486-5p are down-regulated in cumulus cells of metaphase II oocytes from women with polycystic ovary syndrome. Reprod. Biomed. Online 2015, 31, 565–572. [Google Scholar] [CrossRef]
- Wang, M.; Sun, J.; Xu, B.; Chrusciel, M.; Gao, J.; Bazert, M.; Stelmaszewska, J.; Xu, Y.; Zhang, H.; Pawelczyk, L.; et al. Functional characterization of MicroRNA-27a-3p expression in human polycystic ovary syndrome. Endocrinology 2018, 159, 297–309. [Google Scholar] [CrossRef]
- Xu, B.; Zhang, Y.W.; Tong, X.H.; Liu, Y.S. Characterization of microRNA profile in human cumulus granulosa cells: Identification of microRNAs that regulate Notch signaling and are associated with PCOS. Mol. Cell. Endocrinol. 2015, 404, 26–36. [Google Scholar] [CrossRef]
- Sang, Q.; Yao, Z.; Wang, H.; Feng, R.; Wang, H.; Zhao, X.; Xing, Q.; Jin, L.; He, L.; Wu, L.; et al. Identification of MicroRNAs in Human Follicular Fluid: Characterization of MicroRNAs That Govern Steroidogenesis in Vitro and are Associated with Polycystic Ovary Syndrome in Vivo. J. Clin. Endocrinol. Metab. 2013, 98, 3068–3079. [Google Scholar] [CrossRef]
- Santonocito, M.; Vento, M.; Guglielmino, M.R.; Battaglia, R.; Wahlgren, J.; Ragusa, M.; Barbagallo, D.; Borzì, P.; Rizzari, S.; Maugeri, M.; et al. Molecular characterization of exosomes and their microRNA cargo in human follicular fluid: Bioinformatic analysis reveals that exosomal microRNAs control pathways involved in follicular maturation. Fertil. Steril. 2014, 102, 1751–1761. [Google Scholar] [CrossRef]
- Tong, X.H.; Xu, B.; Zhang, Y.W.; Liu, Y.S.; Ma, C.H. Research resources: Comparative microRNA profiles in human corona radiata cells and cumulus oophorus cells detected by next-generation small RNA sequencing. PLoS ONE 2014, 9, e106706. [Google Scholar] [CrossRef]
- Chen, D.; Zhang, Z.; Chen, B.; Ji, D.; Hao, Y.; Zhou, P.; Wei, Z.; Cao, Y. Altered microRNA and Piwi-interacting RNA profiles in cumulus cells from patients with diminished ovarian reserve. Biol. Reprod. 2017, 97, 91–103. [Google Scholar] [CrossRef]
- Diez-Fraile, A.; Lammens, T.; Tilleman, K.; Witkowski, W.; Verhasselt, B.; De Sutter, P.; Benoit, Y.; Espeel, M.; D’Herde, K. Age-associated differential microRNA levels in human follicular fluid reveal pathways potentially determining fertility and success of in vitro fertilization. Hum. Fertil. 2014, 17, 90–98. [Google Scholar] [CrossRef]
- Eisenberg, I.; Nahmias, N.; Novoselsky Persky, M.; Greenfield, C.; Goldman-Wohl, D.; Hurwitz, A.; Haimov-Kochman, R.; Yagel, S.; Imbar, T. Elevated circulating micro-ribonucleic acid (miRNA)-200b and miRNA-429 levels in anovulatory women. Fertil. Steril. 2017, 107, 269–275. [Google Scholar] [CrossRef]
- Fu, J.; Qu, R.; Zhang, Y.; Gu, R.; Li, X.; Sun, Y.; Wang, L.; Sang, Q.; Sun, X. Screening of miRNAs in human follicular fluid reveals an inverse relationship between microRNA-663b expression and blastocyst formation. Reprod. Biomed. Online 2018, 37, 25–32. [Google Scholar] [CrossRef]
- Karakaya, C.; Guzeloglu-Kayisli, O.; Uyar, A.; Kallen, A.N.; Babayev, E.; Bozkurt, N.; Unsal, E.; Karabacak, O.; Seli, E. Poor ovarian response in women undergoing in vitro fertilization is associated with altered microRNA expression in cumulus cells. Fertil. Steril. 2015, 103, 1469–1476. [Google Scholar] [CrossRef]
- Luo, H.; Han, Y.; Liu, J.; Zhang, Y. Identification of microRNAs in granulosa cells from patients with different levels of ovarian reserve function and the potential regulatory function of miR-23a in granulosa cell apoptosis. Gene 2019, 686, 250–260. [Google Scholar] [CrossRef]
- Martinez, R.M.; Liang, L.; Racowsky, C.; Dioni, L.; Mansur, A.; Adir, M.; Bollati, V.; Baccarelli, A.A.; Hauser, R.; Machtinger, R. Extracellular microRNAs profile in human follicular fluid and IVF outcomes. Sci. Rep. 2018, 8, 17036. [Google Scholar] [CrossRef]
- Moreno, J.M.; Núñez, M.J.; Quiñonero, A.; Martínez, S.; De La Orden, M.; Simón, C.; Pellicer, A.; Díaz-García, C.; Domínguez, F. Follicular fluid and mural granulosa cells microRNA profiles vary in in vitro fertilization patients depending on their age and oocyte maturation stage. Fertil. Steril. 2015, 104, 1037–1046. [Google Scholar] [CrossRef]
- Barragán, M.; Pons, J.; Ferrer-Vaquer, A.; Cornet-Bartolomé, D.; Schweitzer, A.; Hubbard, J.; Auer, H.; Rodolosse, A.; Vassena, R. The transcriptome of human oocytes is related to age and ovarian reserve. Mol. Hum. Reprod. 2017, 23, 535–548. [Google Scholar] [CrossRef]
- Xu, Y.W.; Wang, B.; Ding, C.H.; Li, T.; Gu, F.; Zhou, C. Differentially expressed micoRNAs in human oocytes. J. Assist. Reprod. Genet. 2011, 28, 559–566. [Google Scholar] [CrossRef]
- Battaglia, R.; Vento, M.E.; Ragusa, M.; Barbagallo, D.; La Ferlita, A.; Di Emidio, G.; Borzi, P.; Artini, P.G.; Scollo, P.; Tatone, C.; et al. MicroRNAs Are Stored in Human MII Oocyte and Their Expression Profile Changes in Reproductive Aging. Biol. Reprod. 2016, 95, 131. [Google Scholar] [CrossRef]
- Dominguez, F.; Moreno-Moya, J.M.; Lozoya, T.; Romero, A.; Martínez, S.; Monterde, M.; Gurrea, M.; Ferri, B.; Núñez, M.J.; Simón, C.; et al. Embryonic miRNA profiles of normal and ectopic pregnancies. PLoS ONE 2014, 9, e102185. [Google Scholar] [CrossRef]
- McCallie, B.; Schoolcraft, W.B.; Katz-Jaffe, M.G. Aberration of blastocyst microRNA expression is associated with human infertility. Fertil. Steril. 2010, 93, 2374–2382. [Google Scholar] [CrossRef]
- Rosenbluth, E.M.; Shelton, D.N.; Sparks, A.E.T.; Devor, E.; Christenson, L.; Van Voorhis, B.J. MicroRNA expression in the human blastocyst. Fertil. Steril. 2013, 99, 855–861. [Google Scholar] [CrossRef]
- McCallie, B.R.; Parks, J.C.; Strieby, A.L.; Schoolcraft, W.B.; Katz-Jaffe, M.G. Human blastocysts exhibit unique microrna profiles in relation to maternal age and chromosome constitution. J. Assist. Reprod. Genet. 2014, 31, 913–919. [Google Scholar] [CrossRef]
- Lozoya, T.; Domínguez, F.; Romero-Ruiz, A.; Steffani, L.; Martínez, S.; Monterde, M.; Ferri, B.; Núñez, M.J.; Romero-Espinós, A.; Zamora, O.; et al. The Lin28/Let-7 system in early human embryonic tissue and ectopic pregnancy. PLoS ONE 2014, 9, e87698. [Google Scholar] [CrossRef]
- Capalbo, A.; Ubaldi, F.M.; Cimadomo, D.; Noli, L.; Khalaf, Y.; Farcomeni, A.; Ilic, D.; Rienzi, L. MicroRNAs in spent blastocyst culture medium are derived from trophectoderm cells and can be explored for human embryo reproductive competence assessment. Fertil. Steril. 2016, 105, 225–235. [Google Scholar] [CrossRef]
- Rosenbluth, E.M.; Shelton, D.N.; Wells, L.M.; Sparks, A.E.T.; Van Voorhis, B.J. Human embryos secrete microRNAs into culture media—A potential biomarker for implantation. Fertil. Steril. 2014, 101, 1493–1500. [Google Scholar] [CrossRef]
- Lena, A.M.; Mancini, M.; Rivetti di Val Cervo, P.; Saintigny, G.; Mahé, C.; Melino, G.; Candi, E. MicroRNA-191 triggers keratinocytes senescence by SATB1 and CDK6 downregulation. Biochem. Biophys. Res. Commun. 2012, 423, 509–514. [Google Scholar] [CrossRef]
- Cook, J.; Bennett, P.R.; Kim, S.H.; Teoh, T.G.; Sykes, L.; Kindinger, L.M.; Garrett, A.; Binkhamis, R.; MacIntyre, D.A.; Terzidou, V. First Trimester Circulating MicroRNA Biomarkers Predictive of Subsequent Preterm Delivery and Cervical Shortening. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Rodosthenous, R.S.; Burris, H.H.; Sanders, A.P.; Just, A.C.; Dereix, A.E.; Svensson, K.; Solano, M.; Téllez-Rojo, M.M.; Wright, R.O.; Baccarelli, A.A. Second trimester extracellular microRNAs in maternal blood and fetal growth: An exploratory study. Epigenetics 2017, 12, 804–810. [Google Scholar] [CrossRef]
- Singh, K.; Williams, J.; Brown, J.; Wang, E.T.; Lee, B.; Gonzalez, T.L.; Cui, J.; Goodarzi, M.O.; Pisarska, M.D. Up-regulation of microRNA-202-3p in first trimester placenta of pregnancies destined to develop severe preeclampsia, a pilot study. Pregnancy Hypertens. 2017, 10, 7–9. [Google Scholar] [CrossRef]
- Salas-Huetos, A.; James, E.R.; Aston, K.I.; Carrell, D.T.; Jenkins, T.G.; Yeste, M. The role of miRNAs in male human reproduction: A systematic review. Andrology 2019, in press. [Google Scholar] [CrossRef]
- Feng, R.; Sang, Q.; Zhu, Y.; Fu, W.; Liu, M.; Xu, Y.; Shi, H.; Xu, Y.; Qu, R.; Chai, R.; et al. MiRNA-320 in the human follicular fluid is associated with embryo quality in vivo and affects mouse embryonic development in vitro. Sci. Rep. 2015, 5, 8689. [Google Scholar] [CrossRef]
- Sun, Y.X.; Zhang, Y.X.; Zhang, D.; Xu, C.M.; Chen, S.C.; Zhang, J.Y.; Ruan, Y.C.; Chen, F.; Zhang, R.J.; Qian, Y.Q.; et al. XCI-escaping gene KDM5C contributes to ovarian development via downregulating miR-320a. Hum. Genet. 2017, 136, 227–239. [Google Scholar] [CrossRef]
- Zeng, Y.; Rosborough, R.C.; Li, Y.; Gupta, A.R.; Bennett, J. Temporal and spatial regulation of gene expression mediated by the promoter for the human tissue inhibitor of metalloproteinases-3 (TIMP-3)- encoding gene. Dev. Dyn. 1998, 211, 228–237. [Google Scholar] [CrossRef]
- Ketley, A.; Warren, A.; Holmes, E.; Gering, M.; Aboobaker, A.A.; Brook, J.D. The miR-30 MicroRNA Family Targets smoothened to Regulate Hedgehog Signalling in Zebrafish Early Muscle Development. PLoS ONE 2013, 8, e65170. [Google Scholar] [CrossRef]
- Zhao, K.; Chen, Y.; Yang, R.; Bai, Y.; Li, C.; Li, H.; Xiong, C. miR-424/322 is downregulated in the semen of patients with severe DNA damage and may regulate sperm DNA damage. Reprod. Fertil. Dev. 2015, 28, 1598–1607. [Google Scholar] [CrossRef] [PubMed]
- La Ferlita, A.; Battaglia, R.; Andronico, F.; Caruso, S.; Cianci, A.; Purrello, M.; Di Pietro, C. Non-coding RNAs in endometrial physiopathology. Int. J. Mol. Sci. 2018, 19, 2120. [Google Scholar] [CrossRef] [PubMed]
- Chiofalo, B.; Laganà, A.S.; Vaiarelli, A.; La Rosa, V.L.; Rossetti, D.; Palmara, V.; Valenti, G.; Rapisarda, A.M.C.; Granese, R.; Sapia, F.; et al. Do miRNAs play a role in fetal growth restriction? A fresh look to a busy corner. Biomed Res. Int. 2017, 2017, 6073167. [Google Scholar] [CrossRef] [PubMed]
- Lycoudi, A.; Mavreli, D.; Mavrou, A.; Papantoniou, N.; Kolialexi, A. MIRNAs in pregnancy-related complications. Expert Rev. Mol. Diagn. 2015, 15, 999–1010. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Reference | Location | Age (years) | Population Studied | Study Design | Outcome and Aim | Analysis and Normalization Methods | Principal Conclusion | Quality Score |
|---|---|---|---|---|---|---|---|---|
| (A) Ovary and extracellular vesicles (exosomes and microvesicles) | ||||||||
| [9] (Liang et al., 2007) | USA | ND | 1 ovary | Descriptive | To identify miRNA expression patterns in human tissues (including ovary). | Analysis: qRT-PCR. Normalization: Average of miR-30e, miR-92, miR-92N, and miR-423. | A total of 216 miRNAs are expressed in human ovaries. | 4/9 |
| [22] (Sang et al., 2013) | China | Desciptive: ND. Cases: 29.09 ± 0.70 Control: 30.83 ± 0.90 | Follicular fluid and microvesicles. Descriptive: 20 women undergoing ICSI. Cases: 22 PCOS patients Control: 22. | Descriptive | To identify cell-free miRNAs in human follicular fluid and to investigate the function of these miRNAs in vitro and any roles they play in PCOS. | Analysis: qRT-PCR. Normalization: U6 RNA. | Identification of hsa-miRNA-19b, 24, and 222 in human follicular fluid and another 538 known miRNAs. The expression levels of hsa-miRNA-132 and 320 were significantly lower in controls than in PCOS patients. | 4/9 |
| [23] (Santonocito et al., 2014) | Italy | <35 | FF exosomes from 15 women undergoing ICSI | Descriptive | To characterize well-represented miRNAs in human FF and exosomes. | Analysis: qRT-PCR. Normalization: Average of miR-25, miR-28-3p, and miR-145. | 37 miRNAs upregulated in human FF compared with in plasma (15 miRNAs were found to be upregulated in total FF compared with in plasma; 10 miRNAs were carried by exosomes, while five were not). 22 miRNAs were present exclusively in exosomes. | 6/9 |
| [24] (Tong et al., 2014) | China | 29.16 ± 2.7 | 5 women (CRC and COC) | Descriptive | To determine the miRNA expression profiles, via NGS technology, of CRCs and COCs. | Analysis: RNA seq and qRT-PCR validation. Normalization: U6. | A total of 785 and 799 annotated miRNAs were identified in CRCs and COCs respectively. Different expression patterns in CRCs and COCs were detected in 72 annotated miRNAs. | 6/9 |
| [16] (Cai et al., 2017) | China | 29 ± 3.5 | GC. Cases: 25 women with PCOS. Controls 20 women wihtout PCOS. | Case-control | To investigate the effect of miR-145 on cell proliferation and the underlying mechanism of miR-145 in isolated human GCs from the aspirated follicular fluid in women with PCOS. | Analysis: qRT-PCR. Normalization: GAPDH or U6. | miR-145 is downregulated in human GCs from PCOS when compared with control subjects. Insulin receptor substrate 1 (IRS1) gene is a direct target of miR-145. | 6/12 |
| [25] (Chen et al., 2017) | China | NOR: 30.57 ± 2.69. DOR: 33.74 ± 3.25 | COC. Cases: 10 women with DOR. Controls: 10 women with NOR | Case-control | To comprehensively characterize miRNA expression profiles in cumulus cells of DOR patients. | Analysis: RNA seq and qRT-PCR validation. Normalization: U6. | 79 annotated miRNAs, and 5 novel miRNAs were identified differentially expressed between DOR and NOR. mTOR pathway and meiosis-associated biological processes were enriched in the DE-miRNAs | 5/12 |
| [27] (Eisenberg et al., 2017) | Israel | <35. PCOS: 26.9 ± 4.3 MIF: 26.8 ± 4.7 | GC. Cases: 15 normally ovulating with pure male infertility factor, and 18 with PCOS. Controls: 7 normally ovulating. | Case-control | To study the role of micro-RNA (miRNA)-200b and miRNA-429 in human ovulation and to measure their expression levels in ovulatory and anovulatory patients. | Analysis: qRT-PCR. Normalization: U6. | Very low expression levels were detected for the two miRNAs in granulosa cells, with hsa-miRNA-200b expression relatively higher than miRNA-429. No significant expression difference of any of these miRNAs was identified in the GLCs of the two groups analyzed. | 5/12 |
| [28] (Fu et al., 2018) | China | <40. Control: 29.89 ± 3.16. Cases: 30.94 ± 3.82 | 91 individual FF. Cases: 53 no blastocysts. Controls: 38 viable blastocysts | Case-control | To screen miRNAs in human follicular fluid and to explore the relationship between miRNA expression and blastocyst formation | Analysis: qRT-PCR screening and qRT-PCR validation. Normalization: U6. | 13 miRNAs (hsa-miR-185, 202, 22, 224, 29c, 30a-3p, 378, 382, 424, 432, 497, 520c-3p and 663B) were up-regulated, and three miRNAs (hsa-miR-1274a, 139-5p and 150) were down-regulated in cases compared with controls. In the validation, the expression level of miR-663B was found to be significantly higher in cases than in controls. | 5/12 |
| [29] (Karakaya et al., 2015) | Turkey | 4 groups: <35, 35–37, 38–40, >40. | COC in 189 women undergoing IVF–ICSI. Cases: 24 poor-responders. Controls: 32 non-poor responders. | Case-control | To analyze the association of miRNA expression with the number of oocytes retrieved, in women undergoing in vitro fertilization (IVF). | Analysis: Microarray screening and qRT-PCR validation. Normalization: RNU43. | MicroRNA microarray analysis showed up-regulation of 16 miRNAs and down-regulation of 88 miRNAs in poor responders. qRT-PCR confirmed that miR-21-5p expression was significantly up-regulated in poor responders, whereas miR-21-3p expression was significantly lower. | 5/12 |
| [30] (Luo et al., 2019) | China | Poor ovarian response (37 ± 3.16), PCOS (27 ± 3.26), normal patients (29 ± 3.22). | GC. Cases: poor ovarian response (n = 7), PCOS (n = 20). Controls: normal patients (n = 18). | Case-control | To determine the microRNA (miRNA) profiles in GCs from the FF of patients with varying levels of ovarian reserve function. | Analysis: RNA seq. | Identified 20 conserved and 3 novel miRNAs that were upregulated in the poor ovarian response group and 30 conserved miRNAs and 1 novel miRNA that were upregulated in the PCOS group. | 7/12 |
| [17] (Roth et al., 2014) | USA | PCOS: 33.1 ± 4.4. Oocyte donors: 27.1 ± 3.6 | FF. Cases: PCOS (n = 10). Controls: Oocyte donors (n = 10). | Case-control | To determine if miRNAs are differentially expressed in the FF of women with PCOS compared to fertile oocyte donors | Analysis: qRT-PCR screening and qRT-PCR validation. Normalization: U6snRNA. | 29 miRNAs are differentially expressed between PCOS and OD samples. In the validation step only five of these upregulated miRNAs (hsa-miR-9, 18b, 32, 34c, and 135a) displayed a significant increase in expression in the PCOS group compared to OD controls. | 7/12 |
| [18] (Scalici et al., 2016) | France | 19–43 | FF. Cases: 30 women with PCOS. Controls: 91 women with normal ovarian reserve. | Case-control | To investigate the expression profiles of five circulating miRNAs (let-7b, miR-29a, miR-30a, miR-140 and miR-320a) in human FF | Analysis: qRT-PCR. Normalization: miR-16. | Hsa-miR-30a was significantly up-regulated, while miR-140 and let-7b were significantly down-regulated in FF pools from patients with PCOS (n = 30) compared to women with normal ovarian reserve. | 6/12 |
| [19] (Shi et al., 2015) | China | Non-PCOS (28.5 ±3.6). PCOS (28.3 ± 3.3). | COC. Cases: PCOS (n = 24). Controls: Non-PCOS (n = 24). | Case-control | To compare the expression of miRNAs in COC from PCOS and non-PCOS women. | Analysis: qRT-PCR screening and qRT-PCR validation. Normalization: ND. | Hsa-miR-483-5p and 486-5p are significantly decreased in COC of PCOS patients compared with non-PCOS. Four predicted genes, SOCS3, SRF, PTEN and FOXO1, were significantly increased in PCOS COC, and IGF2 was significantly decreased in PCOS COC. | 7/12 |
| [20] (Wang et al., 2018) | China | Non-PCOS (30.00 ± 0.74). PCOS (28.67 ± 0.675) | GC. Cases: 25 PCOS. Controls: 17 non-PCOS healthy women. | Case-control | To characterize the function of microRNA-27a-3p (miR-27a-3p) in PCOS | Analysis: qRT-PCR. Normalization: U6 RNA. | 21 miRNAs were upregulated and 38 were downregulated in PCOS GCs. Hsa-miR-27a-3p was significantly increased in both excised GCs and the ovaries of patients with PCOS compared with the controls. | 6/12 |
| [21] (Xu et al., 2015) | China | Non-PCOS (29.43 ± 3.92). PCOS (28.76 ± 3.51). | GC. Cases: 21 women with PCOS. Controls: 20 women without PCOS. | Case-control | To describe the altered miRNA expression profiles and miRNA targeted signaling pathways in PCOS. | Analysis: RNAseq screening and qRT-PCR validation. Normalization: U6. | A total of 59 known miRNA were identified that are differentially expressed in PCOS cumulus granulosa cells, including 21 miRNAs increased and 38 miRNAs decreased. Notch signaling, regulation of hormone, and energy metabolism of the DE-miRNA target genes. | 6/12 |
| [26] (Diez-Fraile et al., 2014) | Belgium | Younger group (29.3–30.3), older group (38.7–42.4) | 16 women. Young age: < 31 years (n = 8); advanced age: >38 years (n = 8) | Prospective | To report the presence of miRNAs in human FF and identify a set of miRNAs that are differentially expressed in older women compared to younger women. | Analysis: qRT-PCR screening and qRT-PCR validation. Normalization: hsa-miR-483-5p. | Hsa-miR-21-5p was present at significantly higher levels in FF from young women, whereas hsa-miR-99b-3p, 134 and 190b were present at significantly higher levels in the FF from older women. | 8/14 |
| [32] (Moreno et al., 2015) | Spain | Younger group (32.93 ± 2.19), older group (38.20 ± 0.86) | Stage 1: Younger group: <35 (n = 15), older group: >37 (n = 15). Stage 2: MII vs. GV (n = 12) and MI vs. MII (n = 9), each patient acted as her own control. | Prospective | To determine whether there is any difference in the FF miRNA profiles from IVF patients according to their age and oocyte maturation stage. | Analysis: qRT-PCR screening and qRT-PCR validation. Normalization: RNU6B. | hsa-miR-424, which is present in higher proportions in FF from patients with advanced age. When we compared the FF from MII versus GV oocytes, they found 13 differentially expressed miRNAs. When we compared FF from MII versus MI, they found seven differentially expressed miRNAs in MII. | 7/14 |
| [31] (Martinez et al., 2018) | USA | 28.9–33.7 | 126 women. EV-miRNAs in FF from oocytes with normal fertilization (n = 93) and from oocytes that failed to fertilize (n = 33). | Cross-sectional | To assess whether EV miRNAs from FF can serve as biomarkers for fertilization status and day 3 embryo quality. | Analysis: qRT-PCR. Normalization: global mean. | 12 EV-miRNAs were differentially expressed between the normal and failed to fertilize groups. Hsa-miR-92a and miR-130b, were over-expressed in FF samples from oocytes that failed to fertilize compared to those that were normally fertilized. Hsa-miR-888 was over-expressed and miR-214 and miR-454 were underexpressed in samples that resulted in impaired day-3 embryo quality compared to top-quality day-3 embryos. | 8/14 |
| (B) Oocytes | ||||||||
| [33] (Barragán et al., 2017) | Spain | 1. Young women with high AFC (age 21 ± 1 years and 24 ± 3 follicles) and low AFC (age 24 ± 2 years and 8 ± 2 follicles); 2. Old women with high AFC (age 32 ± 2 years and 29 ± 7 follicles) and low AFC (age 34 ± 1 years and 7 ± 1 follicles). | 36 in vivo matured MII oocytes from 30 healthy women recruited for oocyte donation | Retrospective | To identify the coding and noncoding transcriptional profiles of in vivo matured MII human oocytes and evaluate their changes in relation to age and, independently, ovarian reserve. | Analysis: Microarray and qRT-PCR validation. Normalization: Actin B, ubiquitin C and DNA methyltransferase-1. | A set of five miRNAs (ENSG00000221162, hsa-miR-220b, ENSG00000239174, miR-4262 and 1260a) were increased in old group. | 7/14 |
| [35] (Battaglia et al., 2016) | Italy | Young women (28–35 years) and old women (38–40 years) | Six MII oocytes from young women and six from older women | Retrospective | To identify human oocyte miRNAs and demonstrate thatconditions altering oocyte quality, such as reproductive aging | Analysis: qRT-PCR screening and qRT-PCR validation. Normalization: RNU6B. | Twelve miRNAs are differentially expressed in women of advanced reproductive age | 7/14 |
| [34] (Xu et al., 2011) | China | 1. 30.65 ± 3.87 GV, and 2. 32.15 ± 4.63 MII oocytes | 392 oocytes at GV stage and 43 oocytes at MI stage during 251 ICSI cycles | Retrospective | To identify differentially expressed miRNAs and expression patterns of specific miRNAs during meiosis in human oocytes. | Analysis: Microarray and qRT-PCR validation. Normalization: U6 RNA | Compared with GV oocytes, MII oocytes exhibited up-regulation of 4 miRNAs (hsa-miR-193a-5p, 297, 625 and 602), and down-regulation of 11 miRNAs (hsa-miR-888*, 212, 662, 299-5p, 339-5p, 20a, 486-5p, 141*, 768-5p, 376a and 15a). | 7/14 |
| Reference | Location | Age (years) | Population Studied | Study Design | Outcome and Aim | Analysis and Normalization Methods | Principal Conclusion | Quality Score |
|---|---|---|---|---|---|---|---|---|
| [41] (Capalbo et al., 2016) | Italy | ND | ICM-free TE samples and their relative SBM from 5 good-quality human blastocysts | Prospective | To identify miRNAs secreted by human embryos in culture media, which can be used as biomarkers of embryo quality during IVF cycles. | Analysis: qRT-PCR screening and qRT-PCR validation. Normalization: RNU44 and RNU48. | The comparative analysis of TE and SBM samples revealed that 96.6% (57/59) of the miRNAs detected in the SBM were derived from TE cells. MiRNAs analysis of SBM from euploid implanted and unimplanted blastocysts highlighted two miRNAs (hsa-miR-20a and 30c) that showed increased concentrations in the former. | 7/14 |
| [42] (Rosenbluth et al., 2014) | USA | ND | 28 tested blastocysts SBM, from 13 couples | Prospective | To determine whether human blastocysts secrete miRNAs into culture media and whether these reflect embryonic ploidy status and can predict IVF outcomes. | Analysis: qRT-PCR screening and qRT-PCR validation. Normalization: U6. | Ten miRNAs (hsa-miR-106b, 191, 30c, 372, 376a, 548a-3p, 548c-3p, 548d-3p, 576d-3p, and 603) were consistently detected in the spent IVF culture media, but only two miRNAs (hsa-miR-372 and 191) were confirmed by later single assay qRT-PCR. Hsa-miR-191 was more highly concentrated in media from aneuploid embryos, and hsa-miR-191, 372, and 645 were more highly concentrated in media from failed IVF/non-intracytoplasmic sperm injection cycles. | 7/14 |
| [36] (Dominguez et al., 2014) | Spain | Screening: EP: 30.75 ± 1.78, VTOP: 26 ± 3.17. Validation: EP: 30.81 ± 1.55, VTOP: 24.62 ± 1.86. | Cases: 23 patients suffering from tubal EP (8 in screening and 15 in validation). Controls: 29 patients with VTOP (8 in screening and 21 in validation). | Case-control | To investigate the miRNA profile of embryonic tissues in EP and controlled abortions VTOP | Analysis: Microarray screening and qRT-PCR validation. Normalization: SNORD96A. | Four miRNAs (hsa-miR-196b, 30a, 873, and 337-3p) were found to be downregulated in EP versus healthy pregnancy tissues, and three miRNAs (hsa-miR-1288, 451, and 223) were upregulated in EP compared to control pregnancy tissue samples. Validation confirmed the differentially expression of the miRNAs hsa-miR-196 and 223. | 7/12 |
| [40] (Lozoya et al., 2014) | Spain | EP: 30.9, VTOP: 21.1 | Cases: 17 patients suffering from tubal EP. Controls: 23 patients with VTOP. | Case-control | To determine the expression of the elements of the Lin28/Let-7 system, and related miRNAs in early stages of human placentation and ectopic pregnancy | Analysis: qRT-PCR. Normalization: RNU6. | LIN28B mRNA was barely detectable in embryonic tissue from early stages of gestation and sharply increased thereafter to plateau between gestational weeks 7–9. In contrast, expression levels of Let-7, mir-132 and mir-145 were high in embryonic tissue from early gestations (≤6-weeks) and abruptly declined thereafter, especially for Let-7. Opposite trends were detected for mir-323-3p. Embryonic expression of LIN28B mRNA was higher in early stages (≤6-weeks) of ectopic pregnancy than in normal gestation. In contrast, Let-7a expression was significantly lower in early ectopic pregnancies, while miR-132 and miR-145 levels were not altered. Expression of mir-323-3p was also suppressed in ectopic embryonic tissue. | 6/12 |
| [37] (McCallie et al., 2010) | USA | ND | Cryopreserved blastocysts (n = 22). Cases: blastocysts from MF infertility alone (n = 6), and blastocysts from PCOS women (n = 6). Controls: Oocyte donor cycles with no known MF infertility (n = 10) | Case-control | To examine human blastocyst miRNA expression (11 probes) in correlation with human infertility. | Analysis: qRT-PCR. Normalization: RNU48. | Morphologically similar blastocysts derived from patients with PCOS or MF infertility exhibited a significant decrease in the expression of six miRNAs (hsa-let-7a, miR-19a, 19b, 24, 93 and 94) in comparison with donor fertile control blastocysts. Annotation of predicted gene targets for these DE-miRNAs included gene ontology biological processes involved in cell growth and maintenance and transcription. | 6/12 |
| [39] (McCallie et al., 2014) | USA | Chromosomally normal blastocystsfrom young, OD (26.4 years), Chromosomallynormal blastocysts from women in their forties (40–44 years) | Cases: blastocysts produced from women in their forties (n = 5). Controls: young oocyte donor derived blastocysts (n = 5) | Case-control | To determine miRNA expression in human blastocysts relative to advanced maternal age and chromosome constitution. | Analysis: qRT-PCR. Normalization: MammU6. | 42 DE-miRNAs. miR-93 was exclusively expressed in blastocysts from women in their forties and further up-regulated with an abnormal chromosome complement. Up-regulated miR-93 resulted in an inverse down-regulation of targets like SIRT1, resulting in reduced oxidative defense. | 5/12 |
| [38] (Rosenbluth et al., 2013) | USA | ND | Screening (Cases: 5 aneuploids. Controls: 4 male, 5 females euploid.). Validation (Cases: 9 aneuploids. Controls: 7 male, 11 females euploid.). | Case-control and Descriptive | To determine the most highly expressed miRNAs in human blastocysts and to compare miRNAs in euploid versus aneuploid embryos and in male versus female embryos. | Analysis: qRT-PCR screening and qRT-PCR validation. Normalization: snRNA U6. | The most highly expressed miRNA in euploid embryos was miR-372. Screening identified 39 miRNAs that were differentially expressed between euploid (n = 9) and aneuploid (n = 5) embryos, and 21 miRNAs that were differentially expressed between male (n = 4) and female (n = 5). | Case-control: 5/12. Descriptive: 6/9. |
| miRNA Name | miRBase Accession Number | Tissue/Cell Expressed | DE-miRNA Condition(s) | Main Publication(s) |
|---|---|---|---|---|
| hsa-miR-9 | MIMAT0000441 | 1. Granulosa cells 2. Follicular fluid | 1. PCOS vs. oocyte donors 2. PCOS vs. oocyte donors | [17,30] |
| hsa-miR-21 | MIMAT0000076 | 1. Granulosa cells 2. Cumulus cells 3. Follicular fluid | 1. PCOS vs. oocyte donors 2. Poor-responders vs. non-poor responders 3. Older vs. younger women | [26,29,30] |
| hsa-miR-27b | MIMAT0000419 | 1. Follicular fluid 2. Cumulus cells 3. Blastocysts | 1. MII vs. GV 2. Poor-responders vs. non-poor responders 3. Aneuploid vs. euploid | [29,32,38] |
| hsa-miR-29b | MIMAT0000100 | Follicular fluid | MII vs. GV and MII vs. MI | [32] |
| hsa-miR-30a | MIMAT0000087 | 1. Follicular fluid 2. Embryonic tissue | 1. PCOS vs. oocyte donors 2. EP vs. VTOP | [18,36] |
| hsa-miR-146a | MIMAT0000449 | Granulosa cells | PCOS vs. oocyte donors and Poor ovarian response vs. oocyte donors | [30] |
| hsa-miR-150 | MIMAT0000451 | 1. Follicular fluid 2. Cumulus cells | 1. Non-viable vs. viable blastocysts 2. Poor-responders vs. non-poor responders | [28,29] |
| hsa-miR-339-3p | MIMAT0004702 | 1. Follicular fluid 2. Blastocysts | 1. MII vs. GV 2. Aneuploid vs. euploid | [32,38] |
| hsa-miR-424 | MIMAT0001341 | 1. Follicular fluid 2. Cumulus cells 3. Follicular fluid | 1. Non-viable vs. viable blastocysts 2. Poor-responders vs. non-poor responders 3. Older vs. younger women | [28,29,32] |
| hsa-miR-451 | MIMAT0001631 | 1. Follicular fluid 2. Embryonic tissue | 1. MII vs. GV and MII vs. MI 2. EP vs. VTOP | [32,36] |
| hsa-miR-663b | MIMAT0005867 | 1. Follicular fluid 2. Cumulus cells | 1. Non-viable vs. viable blastocysts 2. Poor-responders vs. non-poor responders | [28,29] |
| hsa-miR-1275 | MIMAT0005929 | 1. Cumulus cells 2. Granulosa cells | 1. DOR vs. NOR 2. PCOS vs. oocyte donors | [21,25] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salas-Huetos, A.; James, E.R.; Aston, K.I.; Jenkins, T.G.; Carrell, D.T.; Yeste, M. The Expression of miRNAs in Human Ovaries, Oocytes, Extracellular Vesicles, and Early Embryos: A Systematic Review. Cells 2019, 8, 1564. https://doi.org/10.3390/cells8121564
Salas-Huetos A, James ER, Aston KI, Jenkins TG, Carrell DT, Yeste M. The Expression of miRNAs in Human Ovaries, Oocytes, Extracellular Vesicles, and Early Embryos: A Systematic Review. Cells. 2019; 8(12):1564. https://doi.org/10.3390/cells8121564
Chicago/Turabian StyleSalas-Huetos, Albert, Emma R. James, Kenneth I. Aston, Timothy G. Jenkins, Douglas T. Carrell, and Marc Yeste. 2019. "The Expression of miRNAs in Human Ovaries, Oocytes, Extracellular Vesicles, and Early Embryos: A Systematic Review" Cells 8, no. 12: 1564. https://doi.org/10.3390/cells8121564
APA StyleSalas-Huetos, A., James, E. R., Aston, K. I., Jenkins, T. G., Carrell, D. T., & Yeste, M. (2019). The Expression of miRNAs in Human Ovaries, Oocytes, Extracellular Vesicles, and Early Embryos: A Systematic Review. Cells, 8(12), 1564. https://doi.org/10.3390/cells8121564