MicroRNAs Involved in Carcinogenesis, Prognosis, Therapeutic Resistance, and Applications in Human Triple-Negative Breast Cancer

Abstract
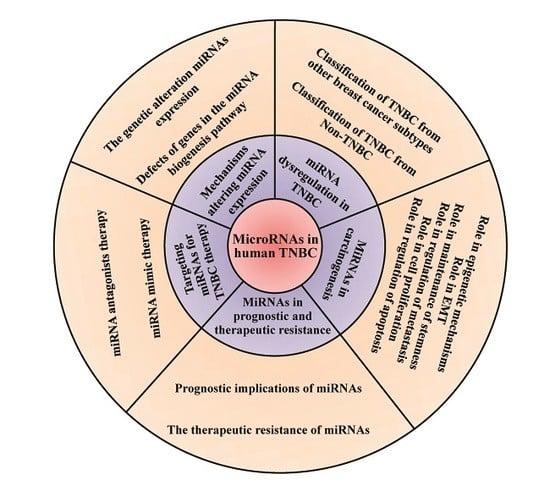
1. Introduction
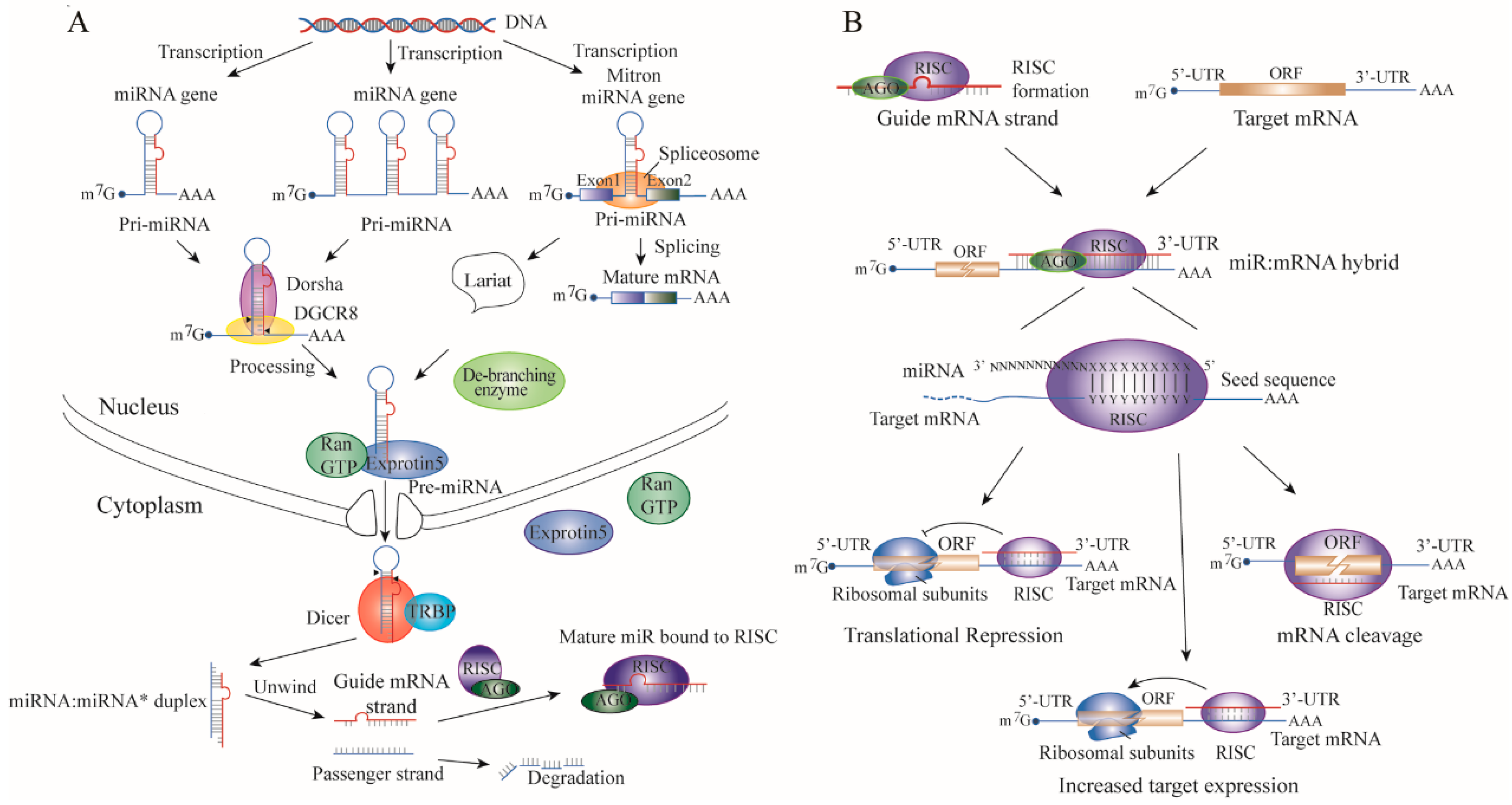
2. The Mechanisms Altering miRNA Expression in TNBC
2.1. The Genetic Alteration of miRNA Expression in TNBC
2.2. Defects of Genes in the miRNA Biogenesis Pathway
3. miRNA Dysregulation in TNBC
3.1. Classification of TNBC from Other Breast Cancer Subtypes with miRNA Signatures
3.2. Classification of TNBC from Non-TNBC with miRNAs Signatures
4. MiRNAs in the Carcinogenesis of TNBC
4.1. MiRNAs in Epigenetic Mechanisms in TNBC
4.2. MiRNAs in EMT in TNBC
4.3. MiRNAs in Maintenance of Stemness in TNBC
4.4. MiRNAs in Regulation of Metastasis in TNBC
4.5. MiRNAs in Cell Proliferation in TNBC
4.6. MiRNAs in Regulation of Apoptosis in TNBC
5. MiRNAs in Prognostic and Therapeutic Resistance in TNBC
5.1. Prognostic Implications of miRNAs in TNBC
5.2. The Therapeutic Resistance of miRNAs in TNBC Therapy
6. Targeting miRNAs for TNBC Therapy
6.1. MiRNA Antagonist Therapy in TNBC
6.2. MiRNA Mimic (miRNA Replacement) Therapy in TNBC
6.3. Challenges and Perspectivse on miRNA Therapy in TNBC
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Perou, C.M.; Sorlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Foulkes, W.D.; Smith, I.E.; Reis-Filho, J.S. Triple-negative breast cancer. N. Engl. J. Med. 2010, 363, 1938–1948. [Google Scholar] [CrossRef] [PubMed]
- Bianchini, G.; Balko, J.M.; Mayer, I.A.; Sanders, M.E.; Gianni, L. Triple-negative breast cancer: Challenges and opportunities of a heterogeneous disease. Nat. Rev. Clin. Oncol. 2016, 13, 674–690. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Lan, Z.; Xiong, X.; Ao, H.; Feng, Y.; Gu, H.; Yu, M.; Cui, Q. The Dual Role of MicroRNAs in Colorectal Cancer Progression. Int. J. Mol. Sci. 2018, 19, 2791. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Valencia-Sanchez, M.A.; Liu, J.; Hannon, G.J.; Parker, R. Control of translation and mRNA degradation by miRNAs and siRNAs. Genes Dev. 2006, 20, 515–524. [Google Scholar] [CrossRef]
- Parker, R.; Song, H. The enzymes and control of eukaryotic mRNA turnover. Nat. Struct. Mol. Biol. 2004, 11, 121–127. [Google Scholar] [CrossRef]
- Mathe, A.; Scott, R.J.; Avery-Kiejda, K.A. MiRNAs and Other Epigenetic Changes as Biomarkers in Triple Negative Breast Cancer. Int. J. Mol. Sci. 2015, 16, 28347–28376. [Google Scholar] [CrossRef]
- Petersen, C.P.; Bordeleau, M.E.; Pelletier, J.; Sharp, P.A. Short RNAs repress translation after initiation in mammalian cells. Mol. Cell 2006, 21, 533–542. [Google Scholar] [CrossRef]
- Sharma, S.; Lu, H.C. MicroRNAs in Neurodegeneration: Current Findings and Potential Impacts. J. Alzheimers Dis. Parkinsonism 2018, 8, 420. [Google Scholar] [CrossRef] [PubMed]
- D’Ippolito, E.; Iorio, M.V. MicroRNAs and triple negative breast cancer. Int. J. Mol. Sci. 2013, 14, 22202–22220. [Google Scholar] [CrossRef] [PubMed]
- Moutinho, C.; Esteller, M. MicroRNAs and Epigenetics. Adv. Cancer Res. 2017, 135, 189–220. [Google Scholar] [PubMed]
- Griffiths-Jones, S.; Saini, H.K.; van Dongen, S.; Enright, A.J. MiRBase: Tools for microRNA genomics. Nucleic Acids Res. 2008, 36, D154–D158. [Google Scholar] [CrossRef]
- Altuvia, Y.; Landgraf, P.; Lithwick, G.; Elefant, N.; Pfeffer, S.; Aravin, A.; Brownstein, M.J.; Tuschl, T.; Margalit, H. Clustering and conservation patterns of human microRNAs. Nucleic Acids Res. 2005, 33, 2697–2706. [Google Scholar] [CrossRef]
- Shaffi, S.K.; Galas, D.; Etheridge, A.; Argyropoulos, C. Role of MicroRNAs in Renal Parenchymal Diseases—A New Dimension. Int. J. Mol. Sci. 2018, 19, 1797. [Google Scholar] [CrossRef]
- Chen, E.; Xu, X.; Liu, R.; Liu, T. Small but Heavy Role: MicroRNAs in Hepatocellular Carcinoma Progression. BioMed Res. Int. 2018, 2018, 6784607. [Google Scholar] [CrossRef]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or Tumor Suppressor? The Duplicity of MicroRNAs in Cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef]
- Paszek, S.; Gablo, N.; Barnas, E.; Szybka, M.; Morawiec, J.; Kolacinska, A.; Zawlik, I. Dysregulation of microRNAs in triple-negative breast cancer. Ginekol. Pol. 2017, 88, 530–536. [Google Scholar] [CrossRef]
- Ventura, A.; Jacks, T. MicroRNAs and cancer: Short RNAs go a long way. Cell 2009, 136, 586–591. [Google Scholar] [CrossRef]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef] [PubMed]
- Melo, S.A.; Esteller, M. Dysregulation of microRNAs in cancer: Playing with fire. FEBS Lett. 2011, 585, 2087–2099. [Google Scholar] [CrossRef] [PubMed]
- Cascione, L.; Gasparini, P.; Lovat, F.; Carasi, S.; Pulvirenti, A.; Ferro, A.; Alder, H.; He, G.; Vecchione, A.; Croce, C.M.; et al. Integrated microRNA and mRNA signatures associated with survival in triple negative breast cancer. PLoS ONE 2013, 8, e55910. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.Y.; Kuo, W.H.; Hung, J.H.; Lee, C.Y.; Lee, Y.H.; Chang, Y.C.; Lin, W.C.; Shen, C.Y.; Huang, C.S.; Hsieh, F.J.; et al. Deregulated microRNAs in triple-negative breast cancer revealed by deep sequencing. Mol. Cancer 2015, 14, 36. [Google Scholar] [CrossRef]
- Cantini, L.; Bertoli, G.; Cava, C.; Dubois, T.; Zinovyev, A.; Caselle, M.; Castiglioni, I.; Barillot, E.; Martignetti, L. Identification of microRNA clusters cooperatively acting on epithelial to mesenchymal transition in triple negative breast cancer. Nucleic Acids Res. 2019, 47, 2205–2215. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Huang, H.Y.; Wang, T.; Wan, Y.; Cui, S.D.; Liu, Z.Z.; Fan, Q.X. Dysregulated expression of dicer and drosha in breast cancer. Pathol. Oncol. Res. 2012, 18, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Dedes, K.J.; Natrajan, R.; Lambros, M.B.; Geyer, F.C.; Lopez-Garcia, M.A.; Savage, K.; Jones, R.L.; Reis-Filho, J.S. Down-regulation of the miRNA master regulators Drosha and Dicer is associated with specific subgroups of breast cancer. Eur. J. Cancer 2011, 47, 138–150. [Google Scholar] [CrossRef]
- Passon, N.; Gerometta, A.; Puppin, C.; Lavarone, E.; Puglisi, F.; Tell, G.; Di Loreto, C.; Damante, G. Expression of Dicer and Drosha in triple-negative breast cancer. J. Clin. Pathol. 2012, 65, 320–326. [Google Scholar] [CrossRef]
- Pickering, B.F.; Yu, D.; Van Dyke, M.W. Nucleolin protein interacts with microprocessor complex to affect biogenesis of microRNAs 15a and 16. J. Biol. Chem. 2011, 286, 44095–44103. [Google Scholar] [CrossRef]
- Pichiorri, F.; Palmieri, D.; De Luca, L.; Consiglio, J.; You, J.; Rocci, A.; Talabere, T.; Piovan, C.; Lagana, A.; Cascione, L.; et al. In vivo NCL targeting affects breast cancer aggressiveness through miRNA regulation. J. Exp. Med. 2013, 210, 951–968. [Google Scholar] [CrossRef]
- Leaderer, D.; Hoffman, A.E.; Zheng, T.; Fu, A.; Weidhaas, J.; Paranjape, T.; Zhu, Y. Genetic and epigenetic association studies suggest a role of microRNA biogenesis gene exportin-5 (XPO5) in breast tumorigenesis. Int. J. Mol. Epidemiol. Genet. 2011, 2, 9–18. [Google Scholar] [PubMed]
- Lin, X.; Wu, M.; Liu, P.; Wei, F.; Li, L.; Tang, H.; Xie, X.; Liu, X.; Yang, L.; Xie, X. Up-regulation and worse prognostic marker of cytoplasmic TARBP2 expression in obstinate breast cancer. Med. Oncol. 2014, 31, 868. [Google Scholar] [CrossRef] [PubMed]
- Tsang, J.; Zhu, J.; van Oudenaarden, A. MicroRNA-mediated feedback and feedforward loops are recurrent network motifs in mammals. Mol. Cell 2007, 26, 753–767. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Lin, J.; Zack, D.J.; Mendell, J.T.; Qian, J. Analysis of regulatory network topology reveals functionally distinct classes of microRNAs. Nucleic Acids Res. 2008, 36, 6494–6503. [Google Scholar] [CrossRef]
- Bertoli, G.; Cava, C.; Castiglioni, I. MicroRNAs: New Biomarkers for Diagnosis, Prognosis, Therapy Prediction and Therapeutic Tools for Breast Cancer. Theranostics 2015, 5, 1122–1143. [Google Scholar] [CrossRef]
- Kawai, S.; Amano, A. BRCA1 regulates microRNA biogenesis via the DROSHA microprocessor complex. J. Cell Biol. 2012, 197, 201–208. [Google Scholar] [CrossRef]
- Shen, J.; Xia, W.; Khotskaya, Y.B.; Huo, L.; Nakanishi, K.; Lim, S.O.; Du, Y.; Wang, Y.; Chang, W.C.; Chen, C.H.; et al. EGFR modulates microRNA maturation in response to hypoxia through phosphorylation of AGO2. Nature 2013, 497, 383–387. [Google Scholar] [CrossRef]
- Van Schooneveld, E.; Wildiers, H.; Vergote, I.; Vermeulen, P.B.; Dirix, L.Y.; Van Laere, S.J. Dysregulation of microRNAs in breast cancer and their potential role as prognostic and predictive biomarkers in patient management. Breast Cancer Res. 2015, 17, 21. [Google Scholar] [CrossRef]
- Boyle, P. Triple-negative breast cancer: Epidemiological considerations and recommendations. Ann. Oncol. 2012, 23, 7–12. [Google Scholar] [CrossRef]
- Farazi, T.A.; Horlings, H.M.; Ten Hoeve, J.J.; Mihailovic, A.; Halfwerk, H.; Morozov, P.; Brown, M.; Hafner, M.; Reyal, F.; van Kouwenhove, M.; et al. MicroRNA sequence and expression analysis in breast tumors by deep sequencing. Cancer Res. 2011, 71, 4443–4453. [Google Scholar] [CrossRef]
- Savad, S.; Mehdipour, P.; Miryounesi, M.; Shirkoohi, R.; Fereidooni, F.; Mansouri, F.; Modarressi, M.H. Expression analysis of MiR-21, MiR-205, and MiR-342 in breast cancer in Iran. Asian Pac. J. Cancer Prev. 2012, 13, 873–877. [Google Scholar] [CrossRef] [PubMed]
- Gasparini, P.; Cascione, L.; Fassan, M.; Lovat, F.; Guler, G.; Balci, S.; Irkkan, C.; Morrison, C.; Croce, C.M.; Shapiro, C.L.; et al. MicroRNA expression profiling identifies a four microRNA signature as a novel diagnostic and prognostic biomarker in triple negative breast cancers. Oncotarget 2014, 5, 1174–1184. [Google Scholar] [CrossRef] [PubMed]
- Calvano Filho, C.M.; Calvano-Mendes, D.C.; Carvalho, K.C.; Maciel, G.A.; Ricci, M.D.; Torres, A.P.; Filassi, J.R.; Baracat, E.C. Triple-negative and luminal A breast tumors: Differential expression of miR-18a-5p, miR-17-5p, and miR-20a-5p. Tumour Biol. 2014, 35, 7733–7741. [Google Scholar] [CrossRef] [PubMed]
- Thakur, S.; Grover, R.K.; Gupta, S.; Yadav, A.K.; Das, B.C. Identification of Specific miRNA Signature in Paired Sera and Tissue Samples of Indian Women with Triple Negative Breast Cancer. PLoS ONE 2016, 11, e0158946. [Google Scholar] [CrossRef] [PubMed]
- Medimegh, I.; Omrane, I.; Privat, M.; Uhrhummer, N.; Ayari, H.; Belaiba, F.; Benayed, F.; Benromdhan, K.; Mader, S.; Bignon, I.J.; et al. MicroRNAs expression in triple negative vs non triple negative breast cancer in Tunisia: Interaction with clinical outcome. PLoS ONE 2014, 9, e111877. [Google Scholar] [CrossRef]
- Lyng, M.B.; Laenkholm, A.V.; Sokilde, R.; Gravgaard, K.H.; Litman, T.; Ditzel, H.J. Global microRNA expression profiling of high-risk ER+ breast cancers from patients receiving adjuvant tamoxifen mono-therapy: A DBCG study. PLoS ONE 2012, 7, e36170. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, W.; Shen, Y.; Guan, X. Identification of dysregulated microRNAs in triple-negative breast cancer (review). Int. J. Oncol. 2015, 46, 927–932. [Google Scholar] [CrossRef]
- Chen, J.; Chen, Z.; Huang, J.; Chen, F.; Ye, W.; Ding, G.; Wang, X. Bioinformatics identification of dysregulated microRNAs in triple negative breast cancer based on microRNA expression profiling. Oncol. Lett. 2018, 15, 3017–3023. [Google Scholar] [CrossRef]
- Yang, R.; Xing, L.; Zheng, X.; Sun, Y.; Wang, X.; Chen, J. The circRNA circAGFG1 acts as a sponge of miR-195-5p to promote triple-negative breast cancer progression through regulating CCNE1 expression. Mol. Cancer 2019, 18, 4. [Google Scholar] [CrossRef]
- Naorem, L.D.; Muthaiyan, M.; Venkatesan, A. Identification of dysregulated miRNAs in triple negative breast cancer: A meta-analysis approach. J. Cell Physiol. 2019, 234, 11768–11779. [Google Scholar] [CrossRef]
- Hu, J.; Xu, J.; Wu, Y.; Chen, Q.; Zheng, W.; Lu, X.; Zhou, C.; Jiao, D. Identification of microRNA-93 as a functional dysregulated miRNA in triple-negative breast cancer. Tumour Biol. 2015, 36, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Piasecka, D.; Braun, M.; Kordek, R.; Sadej, R.; Romanska, H. MicroRNAs in regulation of triple-negative breast cancer progression. J. Cancer Res. Clin. Oncol. 2018, 144, 1401–1411. [Google Scholar] [CrossRef] [PubMed]
- Augoff, K.; McCue, B.; Plow, E.F.; Sossey-Alaoui, K. MiR-31 and its host gene lncRNA LOC554202 are regulated by promoter hypermethylation in triple-negative breast cancer. Mol. Cancer 2012, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Liu, J.; Li, X.; Xiao, G.; Wang, H.; Yang, G.; Li, Y.; Tang, S.C.; Qin, S.; Du, N.; et al. MYC and DNMT3A-mediated DNA methylation represses microRNA-200b in triple negative breast cancer. J. Cell. Mol. Med. 2018, 22, 6262–6274. [Google Scholar] [CrossRef]
- Wee, E.J.; Peters, K.; Nair, S.S.; Hulf, T.; Stein, S.; Wagner, S.; Bailey, P.; Lee, S.Y.; Qu, W.J.; Brewster, B.; et al. Mapping the regulatory sequences controlling 93 breast cancer-associated miRNA genes leads to the identification of two functional promoters of the Hsa-mir-200b cluster, methylation of which is associated with metastasis or hormone receptor status in advanced breast cancer. Oncogene 2012, 31, 4182–4195. [Google Scholar]
- Damiano, V.; Brisotto, G.; Borgna, S.; di Gennaro, A.; Armellin, M.; Perin, T.; Guardascione, M.; Maestro, R.; Santarosa, M. Epigenetic silencing of miR-200c in breast cancer is associated with aggressiveness and is modulated by ZEB1. Genes Chromosomes Cancer 2017, 56, 147–158. [Google Scholar] [CrossRef]
- Taube, J.H.; Malouf, G.G.; Lu, E.; Sphyris, N.; Vijay, V.; Ramachandran, P.P.; Ueno, K.R.; Gaur, S.; Nicoloso, M.S.; Rossi, S.; et al. Epigenetic silencing of microRNA-203 is required for EMT and cancer stem cell properties. Sci. Rep. 2013, 3, 2687. [Google Scholar] [CrossRef]
- Lodygin, D.; Tarasov, V.; Epanchintsev, A.; Berking, C.; Knyazeva, T.; Korner, H.; Knyazev, P.; Diebold, J.; Hermeking, H. Inactivation of miR-34a by aberrant CpG methylation in multiple types of cancer. Cell Cycle 2008, 7, 2591–2600. [Google Scholar] [CrossRef]
- Lehmann, U.; Hasemeier, B.; Christgen, M.; Muller, M.; Romermann, D.; Langer, F.; Kreipe, H. Epigenetic inactivation of microRNA gene hsa-mir-9-1 in human breast cancer. J. Pathol. 2008, 214, 17–24. [Google Scholar] [CrossRef]
- Hoffman, A.E.; Zheng, T.; Yi, C.; Leaderer, D.; Weidhaas, J.; Slack, F.; Zhang, Y.; Paranjape, T.; Zhu, Y. MicroRNA miR-196a-2 and breast cancer: A genetic and epigenetic association study and functional analysis. Cancer Res. 2009, 69, 5970–5977. [Google Scholar] [CrossRef]
- Fabbri, M.; Garzon, R.; Cimmino, A.; Liu, Z.; Zanesi, N.; Callegari, E.; Liu, S.; Alder, H.; Costinean, S.; Fernandez-Cymering, C.; et al. MicroRNA-29 family reverts aberrant methylation in lung cancer by targeting DNA methyltransferases 3A and 3B. Proc. Natl. Acad. Sci. USA 2007, 104, 15805–15810. [Google Scholar] [CrossRef] [PubMed]
- Duursma, A.M.; Kedde, M.; Schrier, M.; le Sage, C.; Agami, R. MiR-148 targets human DNMT3b protein coding region. RNA 2008, 14, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, M.; Calin, G.A. Epigenetics and miRNAs in human cancer. Adv. Genet. 2010, 70, 87–99. [Google Scholar] [PubMed]
- Ng, E.K.; Li, R.; Shin, V.Y.; Siu, J.M.; Ma, E.S.; Kwong, A. MicroRNA-143 is downregulated in breast cancer and regulates DNA methyltransferases 3A in breast cancer cells. Tumour Biol. 2014, 35, 2591–2598. [Google Scholar] [CrossRef] [PubMed]
- Mekala, J.R.; Naushad, S.M.; Ponnusamy, L.; Arivazhagan, G.; Sakthiprasad, V.; Pal-Bhadra, M. Epigenetic regulation of miR-200 as the potential strategy for the therapy against triple-negative breast cancer. Gene 2018, 641, 248–258. [Google Scholar] [CrossRef]
- Tryndyak, V.P.; Beland, F.A.; Pogribny, I.P. E-cadherin transcriptional up by epigenetic and microRNA-200 family alterations is related to mesenchymal and drug-resistant phenotypes in human breast cancer cells. Int. J. Cancer 2010, 126, 2575–2583. [Google Scholar]
- Felipe Lima, J.; Nofech-Mozes, S.; Bayani, J.; Bartlett, J.M. EMT in Breast Carcinoma-A Review. J. Clin. Med. 2016, 5, 65. [Google Scholar] [CrossRef]
- Gwak, J.M.; Kim, H.J.; Kim, E.J.; Chung, Y.R.; Yun, S.; Seo, A.N.; Lee, H.J.; Park, S.Y. MicroRNA-9 is associated with epithelial-mesenchymal transition, breast cancer stem cell phenotype, and tumor progression in breast cancer. Breast Cancer Res. Treat. 2014, 147, 39–49. [Google Scholar] [CrossRef]
- Ma, L.; Young, J.; Prabhala, H.; Pan, E.; Mestdagh, P.; Muth, D.; Teruya-Feldstein, J.; Reinhardt, F.; Onder, T.T.; Valastyan, S.; et al. MiR-9, a MYC/MYCN-activated microRNA, regulates E-cadherin and cancer metastasis. Nat. Cell Biol. 2010, 12, 247–256. [Google Scholar] [CrossRef]
- Stinson, S.; Lackner, M.R.; Adai, A.T.; Yu, N.; Kim, H.J.; O’Brien, C.; Spoerke, J.; Jhunjhunwala, S.; Boyd, Z.; Januario, T.; et al. MiR-221/222 targeting of trichorhinophalangeal 1 (TRPS1) promotes epithelial-to-mesenchymal transition in breast cancer. Sci. Signal 2011, 4, pt5. [Google Scholar] [CrossRef]
- Hwang, M.S.; Yu, N.; Stinson, S.Y.; Yue, P.; Newman, R.J.; Allan, B.B.; Dornan, D. MiR-221/222 targets adiponectin receptor 1 to promote the epithelial-to-mesenchymal transition in breast cancer. PLoS ONE 2013, 8, e66502. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.A.; Sossey-Alaoui, K.; Thompson, C.L.; Danielpour, D.; Schiemann, W.P. TGF-beta upregulates miR-181a expression to promote breast cancer metastasis. J. Clin. Investig. 2013, 123, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Johansson, J.; Berg, T.; Kurzejamska, E.; Pang, M.F.; Tabor, V.; Jansson, M.; Roswall, P.; Pietras, K.; Sund, M.; Religa, P.; et al. MiR-155-mediated loss of C/EBPbeta shifts the TGF-beta response from growth inhibition to epithelial-mesenchymal transition, invasion and metastasis in breast cancer. Oncogene 2013, 32, 5614–5624. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Yan, S.; Weijie, Z.; Feng, W.; Liuxing, W.; Mengquan, L.; Qingxia, F. Critical role of miR-10b in transforming growth factor-beta1-induced epithelial-mesenchymal transition in breast cancer. Cancer Gene Ther. 2014, 21, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Martello, G.; Rosato, A.; Ferrari, F.; Manfrin, A.; Cordenonsi, M.; Dupont, S.; Enzo, E.; Guzzardo, V.; Rondina, M.; Spruce, T.; et al. A MicroRNA targeting dicer for metastasis control. Cell 2010, 141, 1195–1207. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.F.; Xu, L.Y.; Li, E.M. A family of pleiotropically acting microRNAs in cancer progression, miR-200: Potential cancer therapeutic targets. Curr. Pharm. Des. 2014, 20, 1896–1903. [Google Scholar] [CrossRef] [PubMed]
- Humphries, B.; Wang, Z.; Oom, A.L.; Fisher, T.; Tan, D.; Cui, Y.; Jiang, Y.; Yang, C. MicroRNA-200b targets protein kinase Calpha and suppresses triple-negative breast cancer metastasis. Carcinogenesis 2014, 35, 2254–2263. [Google Scholar] [CrossRef]
- Korpal, M.; Lee, E.S.; Hu, G.; Kang, Y. The miR-200 family inhibits epithelial-mesenchymal transition and cancer cell migration by direct targeting of E-cadherin transcriptional repressors ZEB1 and ZEB2. J. Biol. Chem. 2008, 283, 14910–14914. [Google Scholar] [CrossRef]
- Gregory, P.A.; Bert, A.G.; Paterson, E.L.; Barry, S.C.; Tsykin, A.; Farshid, G.; Vadas, M.A.; Khew-Goodall, Y.; Goodall, G.J. The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat. Cell Biol. 2008, 10, 593–601. [Google Scholar] [CrossRef]
- Howe, E.N.; Cochrane, D.R.; Richer, J.K. Targets of miR-200c mediate suppression of cell motility and anoikis resistance. Breast Cancer Res. 2011, 13, R45. [Google Scholar] [CrossRef]
- Rhodes, L.V.; Martin, E.C.; Segar, H.C.; Miller, D.F.; Buechlein, A.; Rusch, D.B.; Nephew, K.P.; Burow, M.E.; Collins-Burow, B.M. Dual regulation by microRNA-200b-3p and microRNA-200b-5p in the inhibition of epithelial-to-mesenchymal transition in triple-negative breast cancer. Oncotarget 2015, 6, 16638–16652. [Google Scholar] [CrossRef] [PubMed]
- Soung, Y.H.; Chung, H.; Yan, C.; Ju, J.; Chung, J. Arrestin Domain Containing 3 Reverses Epithelial to Mesenchymal Transition and Chemo-Resistance of TNBC Cells by Up-Regulating Expression of miR-200b. Cells 2019, 8, 692. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Shin, V.Y.; Siu, M.T.; Ho, J.C.; Cheuk, I.; Kwong, A. MiR-199a-5p confers tumor-suppressive role in triple-negative breast cancer. BMC Cancer 2016, 16, 887. [Google Scholar] [CrossRef] [PubMed]
- Kong, P.; Chen, L.; Yu, M.; Tao, J.; Liu, J.; Wang, Y.; Pan, H.; Zhou, W.; Wang, S. MiR-3178 inhibits cell proliferation and metastasis by targeting Notch1 in triple-negative breast cancer. Cell Death Dis. 2018, 9, 1059. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.D.; Yang, D.X.; Liu, X.P.; Jin, L.Y.; Wang, X.G.; Yang, Z.C.; Liu, D.; Zhao, J.J.; Kong, B.; Li, F.N.; et al. MiR-212-5p Suppresses the Epithelial-Mesenchymal Transition in Triple-Negative Breast Cancer by Targeting Prrx2. Cell Physiol. Biochem. 2017, 44, 1785–1795. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.D.; Kong, B.; Liu, X.P.; Jin, L.Y.; Dong, Q.; Li, F.N.; Wang, H.B. MiR-655 suppresses epithelial-to-mesenchymal transition by targeting Prrx1 in triple-negative breast cancer. J. Cell. Mol. Med. 2016, 20, 864–873. [Google Scholar] [CrossRef]
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef]
- Idowu, M.O.; Kmieciak, M.; Dumur, C.; Burton, R.S.; Grimes, M.M.; Powers, C.N.; Manjili, M.H. CD44(+)/CD24(−/low) cancer stem/progenitor cells are more abundant in triple-negative invasive breast carcinoma phenotype and are associated with poor outcome. Hum. Pathol. 2012, 43, 364–373. [Google Scholar] [CrossRef]
- Jang, G.B.; Kim, J.Y.; Cho, S.D.; Park, K.S.; Jung, J.Y.; Lee, H.Y.; Hong, I.S.; Nam, J.S. Blockade of Wnt/beta-catenin signaling suppresses breast cancer metastasis by inhibiting CSC-like phenotype. Sci. Rep. 2015, 5, 12465. [Google Scholar] [CrossRef]
- Dong, G.; Liang, X.; Wang, D.; Gao, H.; Wang, L.; Wang, L.; Liu, J.; Du, Z. High expression of miR-21 in triple-negative breast cancers was correlated with a poor prognosis and promoted tumor cell in vitro proliferation. Med. Oncol. 2014, 31, 57. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, Y.; Tsuyada, A.; Ren, X.; Wu, X.; Stubblefield, K.; Rankin-Gee, E.K.; Wang, S.E. Transforming growth factor-beta regulates the sphere-initiating stem cell-like feature in breast cancer through miRNA-181 and ATM. Oncogene 2011, 30, 1470–1480. [Google Scholar] [CrossRef] [PubMed]
- Malla, R.R.; Kumari, S.; Gavara, M.M.; Badana, A.K.; Gugalavath, S.; Kumar, D.K.G.; Rokkam, P. A perspective on the diagnostics, prognostics, and therapeutics of microRNAs of triple-negative breast cancer. Biophys. Rev. 2019, 11, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Hwang-Verslues, W.W.; Chang, P.H.; Wei, P.C.; Yang, C.Y.; Huang, C.K.; Kuo, W.H.; Shew, J.Y.; Chang, K.J.; Lee, E.Y.; Lee, W.H. MiR-495 is upregulated by E12/E47 in breast cancer stem cells, and promotes oncogenesis and hypoxia resistance via downregulation of E-cadherin and REDD1. Oncogene 2011, 30, 2463–2474. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Lu, Y.; Yu, L.; Han, X.; Wang, H.; Mao, J.; Shen, J.; Wang, B.; Tang, J.; Li, C.; et al. MiR-221/222 promote cancer stem-like cell properties and tumor growth of breast cancer via targeting PTEN and sustained Akt/NF-kappaB/COX-2 activation. Chem. Biol. Interact. 2017, 277, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Cuiffo, B.G.; Campagne, A.; Bell, G.W.; Lembo, A.; Orso, F.; Lien, E.C.; Bhasin, M.K.; Raimo, M.; Hanson, S.E.; Marusyk, A.; et al. MSC-regulated microRNAs converge on the transcription factor FOXP2 and promote breast cancer metastasis. Cell Stem Cell 2014, 15, 762–774. [Google Scholar] [CrossRef]
- Wang, B.; Wang, Q.; Wang, Z.; Jiang, J.; Yu, S.C.; Ping, Y.F.; Yang, J.; Xu, S.L.; Ye, X.Z.; Xu, C.; et al. Metastatic consequences of immune escape from NK cell cytotoxicity by human breast cancer stem cells. Cancer Res. 2014, 74, 5746–5757. [Google Scholar] [CrossRef]
- Shimono, Y.; Zabala, M.; Cho, R.W.; Lobo, N.; Dalerba, P.; Qian, D.; Diehn, M.; Liu, H.; Panula, S.P.; Chiao, E.; et al. Downregulation of miRNA-200c links breast cancer stem cells with normal stem cells. Cell 2009, 138, 592–603. [Google Scholar] [CrossRef]
- Iliopoulos, D.; Lindahl-Allen, M.; Polytarchou, C.; Hirsch, H.A.; Tsichlis, P.N.; Struhl, K. Loss of miR-200 inhibition of Suz12 leads to polycomb-mediated repression required for the formation and maintenance of cancer stem cells. Mol. Cell 2010, 39, 761–772. [Google Scholar] [CrossRef]
- Rokavec, M.; Wu, W.; Luo, J.L. IL6-mediated suppression of miR-200c directs constitutive activation of inflammatory signaling circuit driving transformation and tumorigenesis. Mol. Cell 2012, 45, 777–789. [Google Scholar] [CrossRef]
- Iliopoulos, D.; Hirsch, H.A.; Struhl, K. An epigenetic switch involving NF-kappaB, Lin28, Let-7 MicroRNA, and IL6 links inflammation to cell transformation. Cell 2009, 139, 693–706. [Google Scholar] [CrossRef]
- Yu, F.; Yao, H.; Zhu, P.; Zhang, X.; Pan, Q.; Gong, C.; Huang, Y.; Hu, X.; Su, F.; Lieberman, J.; et al. Let-7 regulates self renewal and tumorigenicity of breast cancer cells. Cell 2007, 131, 1109–1123. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Deng, H.; Yao, H.; Liu, Q.; Su, F.; Song, E. Mir-30 reduction maintains self-renewal and inhibits apoptosis in breast tumor-initiating cells. Oncogene 2010, 29, 4194–4204. [Google Scholar] [PubMed]
- Guo, L.; Chen, C.; Shi, M.; Wang, F.; Chen, X.; Diao, D.; Hu, M.; Yu, M.; Qian, L.; Guo, N. Stat3-coordinated Lin-28-let-7-HMGA2 and miR-200-ZEB1 circuits initiate and maintain oncostatin M-driven epithelial-mesenchymal transition. Oncogene 2013, 32, 5272–5282. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Ding, C.; Zhang, H.; Gao, J. Let7 miRNAs sensitize breast cancer stem cells to radiationinduced repression through inhibition of the cyclin D1/Akt1/Wnt1 signaling pathway. Mol. Med. Rep. 2016, 14, 3285–3292. [Google Scholar] [CrossRef]
- Polytarchou, C.; Iliopoulos, D.; Struhl, K. An integrated transcriptional regulatory circuit that reinforces the breast cancer stem cell state. Proc. Natl. Acad. Sci. USA 2012, 109, 14470–14475. [Google Scholar] [CrossRef]
- DeCastro, A.J.; Dunphy, K.A.; Hutchinson, J.; Balboni, A.L.; Cherukuri, P.; Jerry, D.J.; DiRenzo, J. MiR203 mediates subversion of stem cell properties during mammary epithelial differentiation via repression of DeltaNP63alpha and promotes mesenchymal-to-epithelial transition. Cell Death Dis. 2013, 4, e514. [Google Scholar] [CrossRef]
- Chao, C.H.; Chang, C.C.; Wu, M.J.; Ko, H.W.; Wang, D.; Hung, M.C.; Yang, J.Y.; Chang, C.J. MicroRNA-205 signaling regulates mammary stem cell fate and tumorigenesis. J. Clin. Investig. 2014, 124, 3093–3106. [Google Scholar] [CrossRef]
- Cheng, S.; Huang, Y.; Lou, C.; He, Y.; Zhang, Y.; Zhang, Q. FSTL1 enhances chemoresistance and maintains stemness in breast cancer cells via integrin beta3/Wnt signaling under miR-137 regulation. Cancer Biol. Ther. 2019, 20, 328–337. [Google Scholar]
- Chen, F.; Luo, N.; Hu, Y.; Li, X.; Zhang, K. MiR-137 Suppresses Triple-Negative Breast Cancer Stemness and Tumorigenesis by Perturbing BCL11A-DNMT1 Interaction. Cell Physiol. Biochem. 2018, 47, 2147–2158. [Google Scholar] [CrossRef]
- Sun, X.; Li, Y.; Zheng, M.; Zuo, W.; Zheng, W. MicroRNA-223 Increases the Sensitivity of Triple-Negative Breast Cancer Stem Cells to TRAIL-Induced Apoptosis by Targeting HAX-1. PLoS ONE 2016, 11, e0162754. [Google Scholar] [CrossRef]
- Chu, J.; Li, Y.; Fan, X.; Ma, J.; Li, J.; Lu, G.; Zhang, Y.; Huang, Y.; Li, W.; Huang, X.; et al. MiR-4319 Suppress the Malignancy of Triple-Negative Breast Cancer by Regulating Self-Renewal and Tumorigenesis of Stem Cells. Cell Physiol. Biochem. 2018, 48, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.D.; Zheng, S.R.; Cai, Y.J.; Chen, D.L.; Shen, Y.Y.; Lin, C.Q.; Hu, X.Q.; Wang, X.H.; Shi, H.; Guo, G.L. IMP3 promotes TNBC stem cell property through miRNA-34a regulation. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 2688–2696. [Google Scholar] [PubMed]
- Zhang, H.; Cai, K.; Wang, J.; Wang, X.; Cheng, K.; Shi, F.; Jiang, L.; Zhang, Y.; Dou, J. MiR-7, inhibited indirectly by lincRNA HOTAIR, directly inhibits SETDB1 and reverses the EMT of breast cancer stem cells by downregulating the STAT3 pathway. Stem Cells 2014, 32, 2858–2868. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Liu, A.Y.; Fan, C.; Zheng, H.; Li, Y.; Zhang, C.; Wu, S.; Yu, D.; Huang, Z.; Liu, F.; et al. MicroRNA-33b Inhibits Breast Cancer Metastasis by Targeting HMGA2, SALL4 and Twist1. Sci. Rep. 2015, 5, 9995. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Y.; Li, X.; Zhang, Y.J.; Li, J.; Zheng, Y.Q.; Liu, M.; Song, X.; Li, X.R. Expression and regulatory function of miRNA-182 in triple-negative breast cancer cells through its targeting of profilin 1. Tumour Biol. 2013, 34, 1713–1722. [Google Scholar] [CrossRef]
- Zhang, X.; Ma, G.; Liu, J.; Zhang, Y. MicroRNA-182 promotes proliferation and metastasis by targeting FOXF2 in triple-negative breast cancer. Oncol. Lett. 2017, 14, 4805–4811. [Google Scholar] [CrossRef]
- Chen, W.; Cai, F.; Zhang, B.; Barekati, Z.; Zhong, X.Y. The level of circulating miRNA-10b and miRNA-373 in detecting lymph node metastasis of breast cancer: Potential biomarkers. Tumour Biol. 2013, 34, 455–462. [Google Scholar] [CrossRef]
- Chen, D.; Dang, B.L.; Huang, J.Z.; Chen, M.; Wu, D.; Xu, M.L.; Li, R.; Yan, G.R. MiR-373 drives the epithelial-to-mesenchymal transition and metastasis via the miR-373-TXNIP-HIF1alpha-TWIST signaling axis in breast cancer. Oncotarget 2015, 6, 32701–32712. [Google Scholar]
- Ma, L.; Teruya-Feldstein, J.; Weinberg, R.A. Tumour invasion and metastasis initiated by microRNA-10b in breast cancer. Nature 2007, 449, 682–688. [Google Scholar] [CrossRef]
- Yan, L.X.; Huang, X.F.; Shao, Q.; Huang, M.Y.; Deng, L.; Wu, Q.L.; Zeng, Y.X.; Shao, J.Y. MicroRNA miR-21 overexpression in human breast cancer is associated with advanced clinical stage, lymph node metastasis and patient poor prognosis. RNA 2008, 14, 2348–2360. [Google Scholar] [CrossRef]
- Avery-Kiejda, K.A.; Braye, S.G.; Mathe, A.; Forbes, J.F.; Scott, R.J. Decreased expression of key tumour suppressor microRNAs is associated with lymph node metastases in triple negative breast cancer. BMC Cancer 2014, 14, 51. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Lim, M.; Zhao, S.; Sano, Y.; Simone, B.A.; Savage, J.E.; Wickstrom, E.; Camphausen, K.; Pestell, R.G.; Simone, N.L. The metastatic potential of triple-negative breast cancer is decreased via caloric restriction-mediated reduction of the miR-17~92 cluster. Breast Cancer Res. Treat. 2014, 146, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Song, C.; Tang, H.; Zhang, C.; Tang, J.; Li, X.; Chen, B.; Xie, X. MiR-629-3p may serve as a novel biomarker and potential therapeutic target for lung metastases of triple-negative breast cancer. Breast Cancer Res. 2017, 19, 72. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Meng, Q.; Pan, A.; Wu, X.; Cui, J.; Wang, Y.; Li, L. MicroRNA-455-3p promotes invasion and migration in triple negative breast cancer by targeting tumor suppressor EI24. Oncotarget 2017, 8, 19455–19466. [Google Scholar] [CrossRef]
- Nie, J.; Jiang, H.C.; Zhou, Y.C.; Jiang, B.; He, W.J.; Wang, Y.F.; Dong, J. MiR-125b regulates the proliferation and metastasis of triple negative breast cancer cells via the Wnt/beta-catenin pathway and EMT. Biosci. Biotechnol. Biochem. 2019, 83, 1062–1071. [Google Scholar] [CrossRef]
- Ren, Y.; Han, X.; Yu, K.; Sun, S.; Zhen, L.; Li, Z.; Wang, S. MicroRNA-200c downregulates XIAP expression to suppress proliferation and promote apoptosis of triple-negative breast cancer cells. Mol. Med. Rep. 2014, 10, 315–321. [Google Scholar] [CrossRef]
- Lee, S.T.; Feng, M.; Wei, Y.; Li, Z.; Qiao, Y.; Guan, P.; Jiang, X.; Wong, C.H.; Huynh, K.; Wang, J.; et al. Protein tyrosine phosphatase UBASH3B is overexpressed in triple-negative breast cancer and promotes invasion and metastasis. Proc. Natl. Acad. Sci. USA 2013, 110, 11121–11126. [Google Scholar] [CrossRef]
- Berber, U.; Yilmaz, I.; Narli, G.; Haholu, A.; Kucukodaci, Z.; Demirel, D. MiR-205 and miR-200c: Predictive Micro RNAs for Lymph Node Metastasis in Triple Negative Breast Cancer. J. Breast Cancer 2014, 17, 143–148. [Google Scholar] [CrossRef]
- Aceto, N.; Sausgruber, N.; Brinkhaus, H.; Gaidatzis, D.; Martiny-Baron, G.; Mazzarol, G.; Confalonieri, S.; Quarto, M.; Hu, G.; Balwierz, P.J.; et al. Tyrosine phosphatase SHP2 promotes breast cancer progression and maintains tumor-initiating cells via activation of key transcription factors and a positive feedback signaling loop. Nat. Med. 2012, 18, 529–537. [Google Scholar]
- Ma, J.; Yang, Y.; Huo, D.; Wang, Z.; Zhai, X.; Chen, J.; Sun, H.; An, W.; Jie, J.; Yang, P. LincRNA-RoR/miR-145 promote invasion and metastasis in triple-negative breast cancer via targeting MUC1. Biochem. Biophys. Res. Commun. 2018, 500, 614–620. [Google Scholar] [CrossRef]
- Eades, G.; Wolfson, B.; Zhang, Y.; Li, Q.; Yao, Y.; Zhou, Q. LincRNA-RoR and miR-145 regulate invasion in triple-negative breast cancer via targeting ARF6. Mol. Cancer Res. 2015, 13, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Gotte, M.; Mohr, C.; Koo, C.Y.; Stock, C.; Vaske, A.K.; Viola, M.; Ibrahim, S.A.; Peddibhotla, S.; Teng, Y.H.; Low, J.Y.; et al. MiR-145-dependent targeting of junctional adhesion molecule A and modulation of fascin expression are associated with reduced breast cancer cell motility and invasiveness. Oncogene 2010, 29, 6569–6580. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tsouko, E.; Jonsson, P.; Bergh, J.; Hartman, J.; Aydogdu, E.; Williams, C. MiR-206 inhibits cell migration through direct targeting of the actin-binding protein coronin 1C in triple-negative breast cancer. Mol. Oncol. 2014, 8, 1690–1702. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qiu, H.; Tang, R.; Song, H.; Pan, H.; Feng, Z.; Chen, L. MiR30a inhibits epithelialmesenchymal transition and metastasis in triplenegative breast cancer by targeting ROR1. Oncol. Rep. 2018, 39, 2635–2643. [Google Scholar]
- Liu, W.; Xu, Y.; Guan, H.; Meng, H. Clinical potential of miR-940 as a diagnostic and prognostic biomarker in breast cancer patients. Cancer Biomark. 2018, 22, 487–493. [Google Scholar] [CrossRef]
- Si, C.; Yu, Q.; Yao, Y. Effect of miR-146a-5p on proliferation and metastasis of triple-negative breast cancer via regulation of SOX5. Exp. Ther. Med. 2018, 15, 4515–4521. [Google Scholar] [CrossRef]
- Tang, W.; Xu, P.; Wang, H.; Niu, Z.; Zhu, D.; Lin, Q.; Tang, L.; Ren, L. MicroRNA-150 suppresses triple-negative breast cancer metastasis through targeting HMGA2. Onco Targets Ther. 2018, 11, 2319–2332. [Google Scholar] [CrossRef]
- Ji, H.; Sang, M.; Liu, F.; Ai, N.; Geng, C. MiR-124 regulates EMT based on ZEB2 target to inhibit invasion and metastasis in triple-negative breast cancer. Pathol. Res. Pract. 2019, 215, 697–704. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, Y.; Jasper, J.; Lykken, E.; Alexander, P.B.; Markowitz, G.J.; McDonnell, D.P.; Li, Q.J.; Wang, X.F. MiR-148a functions to suppress metastasis and serves as a prognostic indicator in triple-negative breast cancer. Oncotarget 2016, 7, 20381–20394. [Google Scholar] [CrossRef]
- Hong, Z.; Hong, C.; Ma, B.; Wang, Q.; Zhang, X.; Li, L.; Wang, C.; Chen, D. MicroRNA1263p inhibits the proliferation, migration, invasion, and angiogenesis of triplenegative breast cancer cells by targeting RGS3. Oncol. Rep. 2019, 42, 1569–1579. [Google Scholar]
- Guo, S.J.; Zeng, H.X.; Huang, P.; Wang, S.; Xie, C.H.; Li, S.J. MiR-508-3p inhibits cell invasion and epithelial-mesenchymal transition by targeting ZEB1 in triple-negative breast cancer. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6379–6385. [Google Scholar] [PubMed]
- Xiong, H.; Yan, T.; Zhang, W.; Shi, F.; Jiang, X.; Wang, X.; Li, S.; Chen, Y.; Chen, C.; Zhu, Y. MiR-613 inhibits cell migration and invasion by downregulating Daam1 in triple-negative breast cancer. Cell. Signal. 2018, 44, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Song, H.; Wu, T.; Xie, D.; Hu, J.; Zhao, J.; Shen, Q.; Fang, L. MiR-519d-3p suppresses breast cancer cell growth and motility via targeting LIM domain kinase 1. Mol. Cell. Biochem. 2018, 444, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Tang, H.; Chen, B.; He, Z.; Deng, M.; Wu, M.; Liu, X.; Yang, L.; Ye, F.; Xie, X. MiR-26a suppresses tumour proliferation and metastasis by targeting metadherin in triple negative breast cancer. Cancer Lett. 2015, 357, 384–392. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, M.; Huang, J.; Li, Y.; Wang, S.; Harrington, C.A.; Qian, D.Z.; Sun, X.X.; Dai, M.S. MicroRNA-130a suppresses breast cancer cell migration and invasion by targeting FOSL1 and upregulating ZO-1. J. Cell. Biochem. 2018, 119, 4945–4956. [Google Scholar] [CrossRef]
- Garcia, A.I.; Buisson, M.; Bertrand, P.; Rimokh, R.; Rouleau, E.; Lopez, B.S.; Lidereau, R.; Mikaelian, I.; Mazoyer, S. Down-regulation of BRCA1 expression by miR-146a and miR-146b-5p in triple negative sporadic breast cancers. EMBO Mol. Med. 2011, 3, 279–290. [Google Scholar] [CrossRef]
- Matamala, N.; Vargas, M.T.; Gonzalez-Campora, R.; Arias, J.I.; Menendez, P.; Andres-Leon, E.; Yanowsky, K.; Llaneza-Folgueras, A.; Minambres, R.; Martinez-Delgado, B.; et al. MicroRNA deregulation in triple negative breast cancer reveals a role of miR-498 in regulating BRCA1 expression. Oncotarget 2016, 7, 20068–20079. [Google Scholar] [CrossRef]
- Chen, H.; Pan, H.; Qian, Y.; Zhou, W.; Liu, X. MiR-25-3p promotes the proliferation of triple negative breast cancer by targeting BTG2. Mol. Cancer 2018, 17, 4. [Google Scholar] [CrossRef]
- Bai, X.; Han, G.; Liu, Y.; Jiang, H.; He, Q. MiRNA-20a-5p promotes the growth of triple-negative breast cancer cells through targeting RUNX3. Biomed. Pharmacother 2018, 103, 1482–1489. [Google Scholar] [CrossRef]
- Lv, Z.D.; Xin, H.N.; Yang, Z.C.; Wang, W.J.; Dong, J.J.; Jin, L.Y.; Li, F.N. miR-135b promotes proliferation and metastasis by targeting APC in triple-negative breast cancer. J. Cell Physiol. 2019, 234, 10819–10826. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, X.; Shen, C.; Shi, Y. MicroRNA-203 suppresses cell proliferation and migration by targeting BIRC5 and LASP1 in human triple-negative breast cancer cells. J. Exp. Clin. Cancer Res. 2012, 31, 58. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zhu, S.; Mo, Y.Y. Suppression of cell growth and invasion by miR-205 in breast cancer. Cell Res. 2009, 19, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Piovan, C.; Palmieri, D.; Di Leva, G.; Braccioli, L.; Casalini, P.; Nuovo, G.; Tortoreto, M.; Sasso, M.; Plantamura, I.; Triulzi, T.; et al. Oncosuppressive role of p53-induced miR-205 in triple negative breast cancer. Mol. Oncol. 2012, 6, 458–472. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Cao, M.; Zhang, J.; Hu, K.; Yin, Z.; Zhou, Z.; Xiao, X.; Yang, Y.; Sheng, W.; Wu, Y.; et al. Hyaluronic acid-chitosan nanoparticles for co-delivery of MiR-34a and doxorubicin in therapy against triple negative breast cancer. Biomaterials 2014, 35, 4333–4344. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Chao, Z.; Kung, H.F.; Lin, M.C.; Dress, A.; Wardle, F.; Jiang, B.H.; Lai, L. MiR-34a modulates ErbB2 in breast cancer. Cell Biol. Int. 2017, 41, 93–101. [Google Scholar] [CrossRef]
- Adams, B.D.; Wali, V.B.; Cheng, C.J.; Inukai, S.; Booth, C.J.; Agarwal, S.; Rimm, D.L.; Gyorffy, B.; Santarpia, L.; Pusztai, L.; et al. MiR-34a Silences c-SRC to Attenuate Tumor Growth in Triple-Negative Breast Cancer. Cancer Res. 2016, 76, 927–939. [Google Scholar] [CrossRef]
- Hou, L.; Chen, M.; Yang, H.; Xing, T.; Li, J.; Li, G.; Zhang, L.; Deng, S.; Hu, J.; Zhao, X.; et al. MiR-940 Inhibited Cell Growth and Migration in Triple-Negative Breast Cancer. Med. Sci. Monit. 2016, 22, 3666–3672. [Google Scholar] [CrossRef]
- Chen, L.L.; Zhang, Z.J.; Yi, Z.B.; Li, J.J. MicroRNA-211-5p suppresses tumour cell proliferation, invasion, migration and metastasis in triple-negative breast cancer by directly targeting SETBP1. Br. J. Cancer 2017, 117, 78–88. [Google Scholar] [CrossRef]
- Wu, Q.; Chen, Z.; Zhang, G.; Zhou, W.; Peng, Y.; Liu, R.; Chen, C.; Feng, J. EZH2 induces the expression of miR-1301 as a negative feedback control mechanism in triple negative breast cancer. Acta. Biochim. Biophys. Sin. (Shanghai) 2018, 50, 693–700. [Google Scholar] [CrossRef]
- Jia, Z.; Liu, Y.; Gao, Q.; Han, Y.; Zhang, G.; Xu, S.; Cheng, K.; Zou, W. MiR-490-3p inhibits the growth and invasiveness in triple-negative breast cancer by repressing the expression of TNKS2. Gene 2016, 593, 41–47. [Google Scholar] [CrossRef]
- Li, D.; Hu, J.; Song, H.; Xu, H.; Wu, C.; Zhao, B.; Xie, D.; Wu, T.; Zhao, J.; Fang, L. MiR-143-3p targeting LIM domain kinase 1 suppresses the progression of triple-negative breast cancer cells. Am. J. Transl. Res. 2017, 9, 2276–2285. [Google Scholar] [PubMed]
- Li, J.; Lai, Y.; Ma, J.; Liu, Y.; Bi, J.; Zhang, L.; Chen, L.; Yao, C.; Lv, W.; Chang, G.; et al. MiR-17-5p suppresses cell proliferation and invasion by targeting ETV1 in triple-negative breast cancer. BMC Cancer 2017, 17, 745. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.X.; Zhang, B.; Wei, J.; Jiang, G.Q.; Wu, Y.L.; Leng, B.J.; Xing, C.G. MiR-539 inhibits proliferation and migration of triple-negative breast cancer cells by down-regulating LAMA4 expression. Cancer Cell Int. 2018, 18, 16. [Google Scholar] [CrossRef] [PubMed]
- Hong, L.; Pan, F.; Jiang, H.; Zhang, L.; Liu, Y.; Cai, C.; Hua, C.; Luo, X.; Sun, J.; Chen, Z. MiR-125b inhibited epithelial-mesenchymal transition of triple-negative breast cancer by targeting MAP2K7. Onco Targets Ther. 2016, 9, 2639–2648. [Google Scholar]
- Zhou, W.; Song, F.; Wu, Q.; Liu, R.; Wang, L.; Liu, C.; Peng, Y.; Mao, S.; Feng, J.; Chen, C. MiR-217 inhibits triple-negative breast cancer cell growth, migration, and invasion through targeting KLF5. PLoS ONE 2017, 12, e0176395. [Google Scholar] [CrossRef]
- Chu, J. MicroRNA-589 serves as a tumor suppressor microRNA through directly targeting metastasis-associated protein 2 in breast cancer. Oncol. Lett. 2019, 18, 2232–2239. [Google Scholar] [CrossRef]
- Ding, L.; Gu, H.; Lan, Z.; Lei, Q.; Wang, W.; Ruan, J.; Yu, M.; Lin, J.; Cui, Q. Downregulation of cyclooxygenase1 stimulates mitochondrial apoptosis through the NFkappaB signaling pathway in colorectal cancer cells. Oncol. Rep. 2019, 41, 559–569. [Google Scholar]
- Wo, L.; Lu, D.; Gu, X. Knockdown of miR-182 promotes apoptosis via regulating RIP1 deubiquitination in TNF-alpha-treated triple-negative breast cancer cells. Tumour Biol. 2016, 37, 13733–13742. [Google Scholar] [CrossRef]
- Song, H.; Li, D.; Wu, T.; Xie, D.; Hua, K.; Hu, J.; Deng, X.; Ji, C.; Deng, Y.; Fang, L. MicroRNA-301b promotes cell proliferation and apoptosis resistance in triple-negative breast cancer by targeting CYLD. BMB Rep. 2018, 51, 602–607. [Google Scholar] [CrossRef]
- Wang, Q.; Li, C.; Zhu, Z.; Teng, Y.; Che, X.; Wang, Y.; Ma, Y.; Wang, Y.; Zheng, H.; Liu, Y.; et al. MiR-155-5p antagonizes the apoptotic effect of bufalin in triple-negative breast cancer cells. Anticancer Drugs 2016, 27, 9–16. [Google Scholar] [CrossRef]
- Li, X.; Wu, B.; Chen, L.; Ju, Y.; Li, C.; Meng, S. Urokinase-type plasminogen activator receptor inhibits apoptosis in triple-negative breast cancer through miR-17/20a suppression of death receptors 4 and 5. Oncotarget 2017, 8, 88645–88657. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ju, H.; Shen, C.; Tong, Z. MiR-429 mediates delta-tocotrienol-induced apoptosis in triple-negative breast cancer cells by targeting XIAP. Int. J. Clin. Exp. Med. 2015, 8, 15648–15656. [Google Scholar]
- Korner, C.; Keklikoglou, I.; Bender, C.; Worner, A.; Munstermann, E.; Wiemann, S. MicroRNA-31 sensitizes human breast cells to apoptosis by direct targeting of protein kinase C epsilon (PKCepsilon). J. Biol. Chem. 2013, 288, 8750–8761. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, J.; Zhang, G. MiR-4458 regulates cell proliferation and apoptosis through targeting SOCS1 in triple-negative breast cancer. J. Cell Biochem. 2019, 120, 12943–12948. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xu, C.; Niu, R.; Hu, G.; Gu, Z.; Zhuang, Z. MiR-890 inhibits proliferation and invasion and induces apoptosis in triple-negative breast cancer cells by targeting CD147. BMC Cancer 2019, 19, 577. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Liu, Z.; Han, Y.; Wang, X.; Yang, Z. Overexpression of miR-509 Increases Apoptosis and Inhibits Invasion via Suppression of Tumor Necrosis Factor-alpha in Triple-Negative Breast Cancer Hs578T Cells. Oncol. Res. 2016, 24, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Wu, Z.; Wu, A.; Huang, Z.; He, N.; Xie, X. MiR-145 promotes TNF-alpha-induced apoptosis by facilitating the formation of RIP1-FADDcaspase-8 complex in triple-negative breast cancer. Tumour Biol. 2016, 37, 8599–8607. [Google Scholar] [CrossRef]
- Phan, B.; Majid, S.; Ursu, S.; de Semir, D.; Nosrati, M.; Bezrookove, V.; Kashani-Sabet, M.; Dar, A.A. Tumor suppressor role of microRNA-1296 in triple-negative breast cancer. Oncotarget 2016, 7, 19519–19530. [Google Scholar] [CrossRef]
- Ke, K.; Lou, T. MicroRNA-10a suppresses breast cancer progression via PI3K/Akt/mTOR pathway. Oncol. Lett. 2017, 14, 5994–6000. [Google Scholar] [CrossRef]
- Jang, M.H.; Kim, H.J.; Gwak, J.M.; Chung, Y.R.; Park, S.Y. Prognostic value of microRNA-9 and microRNA-155 expression in triple-negative breast cancer. Hum. Pathol. 2017, 68, 69–78. [Google Scholar] [CrossRef]
- Song, X.; Liu, Z.; Yu, Z. LncRNA NEF is downregulated in triple negative breast cancer and correlated with poor prognosis. Acta. Biochim. Biophys. Sin. (Shanghai) 2019, 51, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; He, L.; Richards, E.J.; Challa, S.; Xu, C.X.; Permuth-Wey, J.; Lancaster, J.M.; Coppola, D.; Sellers, T.A.; Djeu, J.Y.; et al. Upregulation of miRNA-155 promotes tumour angiogenesis by targeting VHL and is associated with poor prognosis and triple-negative breast cancer. Oncogene 2014, 33, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Toyama, T.; Kondo, N.; Endo, Y.; Sugiura, H.; Yoshimoto, N.; Iwasa, M.; Takahashi, S.; Fujii, Y.; Yamashita, H. High expression of microRNA-210 is an independent factor indicating a poor prognosis in Japanese triple-negative breast cancer patients. Jpn. J. Clin. Oncol. 2012, 42, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Liu, Y.; Cao, Z.; Li, J.; Huang, Y.; Hu, X.; Shao, Z. MicroRNA-493 is a prognostic factor in triple-negative breast cancer. Cancer Sci. 2018, 109, 2294–2301. [Google Scholar] [CrossRef]
- Son, D.; Kim, Y.; Lim, S.; Kang, H.G.; Kim, D.H.; Park, J.W.; Cheong, W.; Kong, H.K.; Han, W.; Park, W.Y.; et al. MiR-374a-5p promotes tumor progression by targeting ARRB1 in triple negative breast cancer. Cancer Lett. 2019, 454, 224–233. [Google Scholar] [CrossRef]
- Tormo, E.; Ballester, S.; Adam-Artigues, A.; Burgues, O.; Alonso, E.; Bermejo, B.; Menendez, S.; Zazo, S.; Madoz-Gurpide, J.; Rovira, A.; et al. The miRNA-449 family mediates doxorubicin resistance in triple-negative breast cancer by regulating cell cycle factors. Sci. Rep. 2019, 9, 5316. [Google Scholar] [CrossRef]
- Tsiakou, A.; Zagouri, F.; Zografos, E.; Samelis, G.; Gazouli, M.; Kalapanida, D.; Giannos, A.; Marinopoulos, S.; Dimitrakakis, K.; Lazaris, C.A.; et al. Prognostic significance of miR-34 rs4938723 T > C polymorphism in triple negative breast cancer patients. Clin. Biochem. 2019, 68, 9–14. [Google Scholar] [CrossRef]
- Liu, M.; Gong, C.; Xu, R.; Chen, Y.; Wang, X. MicroRNA-5195-3p enhances the chemosensitivity of triple-negative breast cancer to paclitaxel by downregulating EIF4A2. Cell. Mol. Biol. Lett. 2019, 24, 47. [Google Scholar] [CrossRef]
- Sha, L.Y.; Zhang, Y.; Wang, W.; Sui, X.; Liu, S.K.; Wang, T.; Zhang, H. MiR-18a upregulation decreases Dicer expression and confers paclitaxel resistance in triple negative breast cancer. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 2201–2208. [Google Scholar]
- Hou, X.; Niu, Z.; Liu, L.; Guo, Q.; Li, H.; Yang, X.; Zhang, X. MiR-1207-5p regulates the sensitivity of triple-negative breast cancer cells to Taxol treatment via the suppression of LZTS1 expression. Oncol. Lett. 2019, 17, 990–998. [Google Scholar] [CrossRef]
- Wu, C.; Zhao, A.; Tan, T.; Wang, Y.; Shen, Z. Overexpression of microRNA-620 facilitates the resistance of triple negative breast cancer cells to gemcitabine treatment by targeting DCTD. Exp. Ther. Med. 2019, 18, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liang, Y.; Sang, Y.; Song, X.; Zhang, H.; Liu, Y.; Jiang, L.; Yang, Q. MiR-770 suppresses the chemo-resistance and metastasis of triple negative breast cancer via direct targeting of STMN1. Cell Death Dis. 2018, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, S.; Cangemi, A.; Galvano, A.; Fanale, D.; Buscemi, S.; Ciaccio, M.; Russo, A.; Castorina, S.; Bazan, V. Analysis of miRNA expression profile induced by short term starvation in breast cancer cells treated with doxorubicin. Oncotarget 2017, 8, 71924–71932. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Cheng, Y.; She, Q.; Jiang, Y.; Chen, Y.; Yang, W.; Li, Y. Long non-coding RNA TUG1 sponges miR-197 to enhance cisplatin sensitivity in triple negative breast cancer. Biomed. Pharmacother. 2018, 107, 338–346. [Google Scholar] [CrossRef]
- Wang, Q.; Cheng, Y.; Wang, Y.; Fan, Y.; Li, C.; Zhang, Y.; Wang, Y.; Dong, Q.; Ma, Y.; Teng, Y.E.; et al. Tamoxifen reverses epithelial-mesenchymal transition by demethylating miR-200c in triple-negative breast cancer cells. BMC Cancer 2017, 17, 492. [Google Scholar] [CrossRef]
- Li, H.Y.; Liang, J.L.; Kuo, Y.L.; Lee, H.H.; Calkins, M.J.; Chang, H.T.; Lin, F.C.; Chen, Y.C.; Hsu, T.I.; Hsiao, M.; et al. MiR-105/93-3p promotes chemoresistance and circulating miR-105/93-3p acts as a diagnostic biomarker for triple negative breast cancer. Breast Cancer Res. 2017, 19, 133. [Google Scholar] [CrossRef]
- De Mattos-Arruda, L.; Bottai, G.; Nuciforo, P.G.; Di Tommaso, L.; Giovannetti, E.; Peg, V.; Losurdo, A.; Perez-Garcia, J.; Masci, G.; Corsi, F.; et al. MicroRNA-21 links epithelial-to-mesenchymal transition and inflammatory signals to confer resistance to neoadjuvant trastuzumab and chemotherapy in HER2-positive breast cancer patients. Oncotarget 2015, 6, 37269–37280. [Google Scholar] [CrossRef]
- Huang, X.; Taeb, S.; Jahangiri, S.; Emmenegger, U.; Tran, E.; Bruce, J.; Mesci, A.; Korpela, E.; Vesprini, D.; Wong, C.S.; et al. MiRNA-95 mediates radioresistance in tumors by targeting the sphingolipid phosphatase SGPP1. Cancer Res. 2013, 73, 6972–6986. [Google Scholar] [CrossRef]
- Liang, Z.; Ahn, J.; Guo, D.; Votaw, J.R.; Shim, H. MicroRNA-302 replacement therapy sensitizes breast cancer cells to ionizing radiation. Pharm. Res. 2013, 30, 1008–1016. [Google Scholar] [CrossRef]
- Ren, Y.Q.; Fu, F.; Han, J. MiR-27a modulates radiosensitivity of triple-negative breast cancer (TNBC) cells by targeting CDC27. Med. Sci. Monit. 2015, 21, 1297–1303. [Google Scholar]
- Gasparini, P.; Lovat, F.; Fassan, M.; Casadei, L.; Cascione, L.; Jacob, N.K.; Carasi, S.; Palmieri, D.; Costinean, S.; Shapiro, C.L.; et al. Protective role of miR-155 in breast cancer through RAD51 targeting impairs homologous recombination after irradiation. Proc. Natl. Acad. Sci. USA 2014, 111, 4536–4541. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Peng, J.; Fu, Y.; An, S.; Rezaei, K.; Tabbara, S.; Teal, C.B.; Man, Y.G.; Brem, R.F.; Fu, S.W. MiR-638 mediated regulation of BRCA1 affects DNA repair and sensitivity to UV and cisplatin in triple-negative breast cancer. Breast Cancer Res. 2014, 16, 435. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.W.; Hao, W.J.; Li, Y.W.; Li, Y.X.; Zhao, B.C.; Lu, D. Hsa-miRNA-143-3p Reverses Multidrug Resistance of Triple-Negative Breast Cancer by Inhibiting the Expression of Its Target Protein Cytokine-Induced Apoptosis Inhibitor 1 In Vivo. J. Breast Cancer 2018, 21, 251–258. [Google Scholar] [CrossRef]
- Jafri, M.A.; Zaidi, S.K.; Ansari, S.A.; Al-Qahtani, M.H.; Shay, J.W. MicroRNAs as potential drug targets for therapeutic intervention in colorectal cancer. Expert Opin. Ther. Targets 2015, 19, 1705–1723. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Reinhardt, F.; Pan, E.; Soutschek, J.; Bhat, B.; Marcusson, E.G.; Teruya-Feldstein, J.; Bell, G.W.; Weinberg, R.A. Therapeutic silencing of miR-10b inhibits metastasis in a mouse mammary tumor model. Nat. Biotechnol. 2010, 28, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Shu, D.; Li, H.; Shu, Y.; Xiong, G.; Carson, W.E., 3rd; Haque, F.; Xu, R.; Guo, P. Systemic Delivery of Anti-miRNA for Suppression of Triple Negative Breast Cancer Utilizing RNA Nanotechnology. ACS Nano 2015, 9, 9731–9740. [Google Scholar] [CrossRef]
- Yin, H.; Xiong, G.; Guo, S.; Xu, C.; Xu, R.; Guo, P.; Shu, D. Delivery of Anti-miRNA for Triple-Negative Breast Cancer Therapy Using RNA Nanoparticles Targeting Stem Cell Marker CD133. Mol. Ther. 2019, 27, 1252–1261. [Google Scholar] [CrossRef]
- Devulapally, R.; Sekar, N.M.; Sekar, T.V.; Foygel, K.; Massoud, T.F.; Willmann, J.K.; Paulmurugan, R. Polymer nanoparticles mediated codelivery of antimiR-10b and antimiR-21 for achieving triple negative breast cancer therapy. ACS Nano 2015, 9, 2290–2302. [Google Scholar] [CrossRef]
- Zhou, Z.; Kennell, C.; Lee, J.Y.; Leung, Y.K.; Tarapore, P. Calcium phosphate-polymer hybrid nanoparticles for enhanced triple negative breast cancer treatment via co-delivery of paclitaxel and miR-221/222 inhibitors. Nanomedicine 2017, 13, 403–410. [Google Scholar] [CrossRef]
- Ahir, M.; Bhattacharya, S.; Karmakar, S.; Mukhopadhyay, A.; Mukherjee, S.; Ghosh, S.; Chattopadhyay, S.; Patra, P.; Adhikary, A. Tailored-CuO-nanowire decorated with folic acid mediated coupling of the mitochondrial-ROS generation and miR425-PTEN axis in furnishing potent anti-cancer activity in human triple negative breast carcinoma cells. Biomaterials 2016, 76, 115–132. [Google Scholar] [CrossRef]
- Ebert, M.S.; Neilson, J.R.; Sharp, P.A. MicroRNA sponges: Competitive inhibitors of small RNAs in mammalian cells. Nat. Methods 2007, 4, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Chen, Y.; Wu, W.; Ouyang, N.; Chen, J.; Li, H.; Liu, X.; Su, F.; Lin, L.; Yao, Y. MiR-150 promotes human breast cancer growth and malignant behavior by targeting the pro-apoptotic purinergic P2X7 receptor. PLoS ONE 2013, 8, e80707. [Google Scholar] [CrossRef] [PubMed]
- Beg, M.S.; Brenner, A.J.; Sachdev, J.; Borad, M.; Kang, Y.K.; Stoudemire, J.; Smith, S.; Bader, A.G.; Kim, S.; Hong, D.S. Phase I study of MRX34, a liposomal miR-34a mimic, administered twice weekly in patients with advanced solid tumors. Investig. New Drugs 2017, 35, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Van Roosbroeck, K.; Calin, G.A. Cancer Hallmarks and MicroRNAs: The Therapeutic Connection. Adv. Cancer Res. 2017, 135, 119–149. [Google Scholar] [PubMed]
- Di Martino, M.T.; Leone, E.; Amodio, N.; Foresta, U.; Lionetti, M.; Pitari, M.R.; Cantafio, M.E.; Gulla, A.; Conforti, F.; Morelli, E.; et al. Synthetic miR-34a mimics as a novel therapeutic agent for multiple myeloma: In vitro and in vivo evidence. Clin. Cancer Res. 2012, 18, 6260–6270. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, J.; Wang, Y.; Chen, M. Hyaluronic acid-coated PEI-PLGA nanoparticles mediated co-delivery of doxorubicin and miR-542-3p for triple negative breast cancer therapy. Nanomedicine 2016, 12, 411–420. [Google Scholar] [CrossRef]
- Petrs-Silva, H.; Linden, R. Advances in recombinant adeno-associated viral vectors for gene delivery. Curr. Gene Ther. 2013, 13, 335–345. [Google Scholar] [CrossRef]
- Kota, J.; Chivukula, R.R.; O’Donnell, K.A.; Wentzel, E.A.; Montgomery, C.L.; Hwang, H.W.; Chang, T.C.; Vivekanandan, P.; Torbenson, M.; Clark, K.R.; et al. Therapeutic microRNA delivery suppresses tumorigenesis in a murine liver cancer model. Cell 2009, 137, 1005–1017. [Google Scholar] [CrossRef]
- Trepel, M.; Korbelin, J.; Spies, E.; Heckmann, M.B.; Hunger, A.; Fehse, B.; Katus, H.A.; Kleinschmidt, J.A.; Muller, O.J.; Michelfelder, S. Treatment of multifocal breast cancer by systemic delivery of dual-targeted adeno-associated viral vectors. Gene Ther. 2015, 22, 848. [Google Scholar] [CrossRef]
- Bader, A.G.; Brown, D.; Stoudemire, J.; Lammers, P. Developing therapeutic microRNAs for cancer. Gene Ther. 2011, 18, 1121–1126. [Google Scholar] [CrossRef]
- Gupta, I.; Sareyeldin, R.M.; Al-Hashimi, I.; Al-Thawadi, H.A.; Al Farsi, H.; Vranic, S.; Al Moustafa, A.E. Triple Negative Breast Cancer Profile, from Gene to microRNA, in Relation to Ethnicity. Cancers 2019, 11, 363. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wu, D.; Zhu, J.; Jiang, O.; Shi, Q.; Tian, J.; Weng, Y. Upregulation of miR-195 increases the sensitivity of breast cancer cells to Adriamycin treatment through inhibition of Raf-1. Oncol. Rep. 2013, 30, 877–889. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Yao, Y.; Wang, Y.; Liu, B.; Wu, W.; Chen, J.; Su, F.; Yao, H.; Song, E. Up-regulation of miR-21 mediates resistance to trastuzumab therapy for breast cancer. J. Biol. Chem. 2011, 286, 19127–19137. [Google Scholar] [CrossRef] [PubMed]

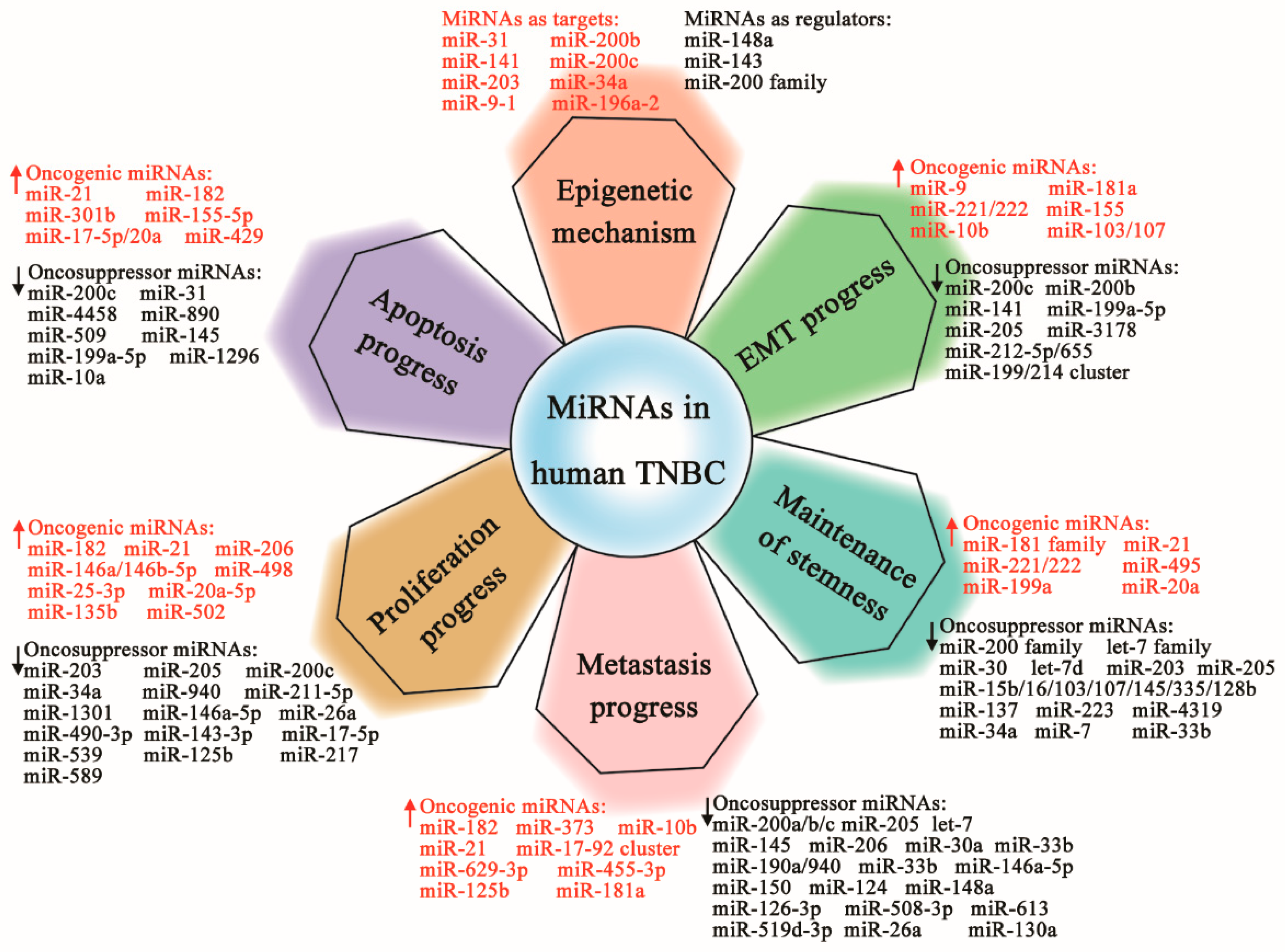
 ) or oncosuppressive genes (Downregulated,
) or oncosuppressive genes (Downregulated,  ) associated with epithelial–mesenchymal transition (EMT), stemness maintenance, invasion and metastases, cell proliferation and survival, and apoptotic regulation via special targets, resulting in the relapse, development, or inhibition of TNBC.
) or oncosuppressive genes (Downregulated, ) associated with epithelial–mesenchymal transition (EMT), stemness maintenance, invasion and metastases, cell proliferation and survival, and apoptotic regulation via special targets, resulting in the relapse, development, or inhibition of TNBC.
) associated with epithelial–mesenchymal transition (EMT), stemness maintenance, invasion and metastases, cell proliferation and survival, and apoptotic regulation via special targets, resulting in the relapse, development, or inhibition of TNBC.
) or oncosuppressive genes (Downregulated, ) associated with epithelial–mesenchymal transition (EMT), stemness maintenance, invasion and metastases, cell proliferation and survival, and apoptotic regulation via special targets, resulting in the relapse, development, or inhibition of TNBC.

{kind=link}
{kind=link}
{kind=link}
| miRNA | Type of Deregulation | Samples Type | Reference |
|---|---|---|---|
| Differentially Expressed miRNAs Between TNBC and other breast cancer (BC) Subtypes | |||
| miR-17-92 cluster | Upregulated | Tumor tissues | [40] |
| miR-205, miR-342 | Downregulated | Tumor samples | [38,41] |
| miR-155, miR-493, miR-30e, miR-27a | Deregulated | Tumor samples | [42] |
| miR-17-5p, miR-18a-5p, miR-20a-5 | Upregulated | Tumor samples | [43] |
| miR-21, miR-221, miR-210, let-7a | Upregulated | Tumor samples | [44] |
| miR-195, miR-145 | Downregulated | Tumor samples | [44] |
| Differentially Expressed miRNAs Between TNBC and Non-TNBC | |||
| miR-21, miR-146a, miR-182, miR-10b | Upregulated | Tumor samples | [45] |
| miR-190a, miR-136-5p, miR-126-5p, miR-135b-5p, miR-182-5p | Downregulated | Serum | [46] |
| miR-146a, miR-100, miR-125b, miR-29a, miR-222, miR-221, miR-138-5p, miR-4324, miR-4800-3p, miR-6836-3p | Upregulated | TNBC cell lines | [47] |
| miR-200a, miR-200c, miR-141, miR-375, miR-203, miR-363-5p, miR-182-5p, miR-141-3p, miR-339-5p, miR-4655-3p, miR-4784, miR-664b-5p, miR-6787-5p | Downregulated | TNBC cell lines | [47] |
| miR-190a, miR-136-5p, miR-126-5p | Downregulated | Tumor samples | [19] |
| miR-135b-5p, miR-182-5p | Upregulated | Tumor samples | [19] |
| miR-195-5p | Downregulated | Tumor samples | [49] |
| miR-135b-5p, miR-18a-5p, miR-9-5p, miR-522-3p | Upregulated | Tumor samples | [50] |
| miR-190b, miR-449a | Downregulated | Tumor samples | [50] |
| miR-93 | Upregulated | Tumor samples | [51] |
| MiRNA | Direct/Indirect Targets | Functions | Reference |
|---|---|---|---|
| MiRNAs as the Targets of Aberrant Epigenetic Modulation | |||
| miR-31 | WAVE3, RhoA, Radexin, integrin | Metastasis suppressor | [53] |
| miR-200b | ZEB1, SOX2, CD133 | Inhibit the capacities of migration, invasion, and mammosphere formation | [54,55] |
| miR-200c/141 | ZEB1 | Lymph node metastasis, cellular plasticity | [56] |
| miR-203 | DKK1 | EMT and cancer stem cell properties | [57] |
| miR-34a | CDK6 | Tumor suppressor gene | [58] |
| miR-9-1 | Unknown | Associated with the early and frequent event in breast cancer development | [59] |
| miR-196a-2 | HOXB2, HOXB3, HOXC13, HOXB5 | A delay in the G2/M phase of the cell cycle | [60] |
| MiRNAs as the Regulators of Aberrant Epigenetic Modulation | |||
| miR-148a | DNMT3B | Associated with the primary human breast cancer development | [59,63] |
| miR-143 | DNMT3A | Inhibits proliferation and soft agar colony formation, downregulates the expression of DNMT3A | [64] |
| miR-200 family | HDAC4 | Influences the mesenchymal phenotype and drug-resistant phenotypes | [65,66] |
| MiRNA | Direct/Indirect Targets | Functions | Reference |
|---|---|---|---|
| Oncogenic miRNAs | |||
| miR-9 | CDH1 | Increases cell motility and invasiveness, tumor angiogenesis | [68,69] |
| miR-221/222 | ZEB2, TRPS1, ADIPOR1 | Induce EMT and increase cell invasion, activate the NF-kB | [70,71] |
| miR-181a | Bim | Enhances TGF-β-mediated EMT, migratory, and invasive phenotypes | [72] |
| miR-155 | C/EBPβ | Promotes EMT, invasion, and metastasis | [73] |
| miR-10b | TGF-β1 | Promotes EMT, invasion, and proliferation | [74] |
| miR-103/107 | Dicer | Promotes migration, invasion, and EMT | [75,76] |
| Oncosuppressor miRNAs | |||
| miR-200 family | ZEB1/2 | Reverse EMT | [77,78,79] |
| miR-200c | ZEB1/2, FN1, MSN, NTRK2, LEPR, ARHGAP19 | Maintain the epithelial phenotype, suppress cell migration | [80] |
| miR-200b | ZEB1/2, PKCα | Reverse EMT phenotypes | [74,81,82] |
| miR-141 | Unknown | Epithelial phenotype maintenance | [47] |
| miR-205 | ZEB1/2 | Reduce TGF-β-induced EMT | [79] |
| miR-199a-5p | PIK3CD | Alters EMT-related genes expression, reduces cell motility and invasiveness, represses tumor cell growth | [83] |
| miR-3178 | Notch-1 | Inhibits EMT, suppress proliferation, invasion, and migration | [84] |
| miR-212-5p/655 | Prrx1/2 | Inhibits EMT phenotype | [85,86] |
| miR-199/214 cluster | Col1 | Decreases TNBC phenotype via its control of proliferation and EMT | [25] |
| MiRNA | Direct/Indirect Targets | Functions | Reference |
|---|---|---|---|
| Oncogenic miRNAs | |||
| miR-21 | PTEN | Increases EMT process, induces breast cancer stem cell (BCSC)-like phenotype, promotes migration and invasion | [51,74,90] |
| miR-181 family | ATM | Induces a stem cell phenotype | [72,91] |
| miR-495 | E-cadherin, REDD1 | Maintain stem cell-like features, promote cell invasion and proliferation in hypoxia | [92,93] |
| miR-221/222 | PTEN | Promotes BCSC properties and tumor growth | [94] |
| miR-199a | FOXP2 | Enhances BCSC properties, promotes metastasis | [95] |
| miR-20a | NKG2D, MICA/B | Promote the lung metastasis by enhancement of BCSC resistance to NK cell cytotoxicity | [96] |
| Oncosuppressor miRNAs | |||
| miR-200 family | Bmi-1, Suz12, | Regulate BCSC formation and growth | [97,98,99] |
| let-7 family | H-RAS, HMGA2 | Regulate the mammosphere formation, BCSC self-renewal and metastasis | [100,101] |
| miR-30 | Ubc9, ITGB3 | Inhibit the self-renewal of breast tumor-initiating cells (BT-ICs), trigger apoptosis | [102] |
| let-7d | Cyclin D1 | Induces stem cells radiation sensitization | [104] |
| miR-15b/16/103/107/145/335/128b | Bmi-1, Suz12, ZEB1/2, Klf4 | Inhibit cancer stem cell (CSC) growth | [105] |
| miR-203 | ΔNp63α | Forfeiture of self-renewing capacity associated with epithelial stem cells, suppresses proliferation and colony formation | [106] |
| miR-205 | Notch-2 | Inhibits EMT and stem cell properties | [107] |
| miR-137 | FSTL1 | Suppresses TNBC stemness | [108,109] |
| miR-223 | HAX-1 | Re-sensitizes TNBC stem cells to tumor necrosis factor-related apoptosis | [110] |
| miR-4319 | E2F2 | Suppresses the self-renewal and malignancy in stem cells | [111] |
| miR-34a | IMP3 | Regulates TNBC stem cell property | [112] |
| miR-7 | SETDB1 | Inhibits cell invasion and metastasis, decreases the BCSC population, and partially reverses EMT | [113] |
| miR-33b | HMGA2, SALL4, Twist1 | Regulate cell stemness and metastasis | [114] |
| MiRNA | Direct/Indirect Targets | Functions | Reference |
|---|---|---|---|
| Oncogenic miRNAs | |||
| miR-182 | PFN1, FOXF2 | Promote cell proliferation, invasion, and migration | [115,116] |
| miR-373 | TXNIP | Induces cancer cell EMT and metastasis | [117,118] |
| miR-10b | HOXD10 | Promotes invasion and metastasis | [119] |
| miR-21 | PTEN | Lymph node metastasis | [120] |
| miR-17/92 cluster | COL4A3, LAMA3, TIMP2/3 | Lymph node metastases, promote metastasis | [121,122] |
| miR-629-3p | LIFR | Lung metastases | [123] |
| miR-455-3p | EI24 | Promotes proliferation, invasion, and migration | [124] |
| miR-125b | APC | Promotes proliferation, metastasis, and EMT | [125] |
| miR-181a | Bim | Promotes EMT, migratory, and invasive | [72] |
| Oncosuppressor miRNAs | |||
| miR-200a/b/c | PKCα, UBASH3B, XIAP | Suppress proliferation, migration, invasion, and metastasis, promote apoptosis | [77,126,127] |
| miR-205 | Unknown | Lymph node metastasis | [128] |
| let-7 | RAS, c-Myc | Block growth and reduce metastasis | [129] |
| miR-145 | MUC1, Arf6, JAM-A, Fascin | Reduce cell motility, invasiveness, and metastasis | [130,131,132] |
| miR-206 | CORO1C | Regulates metastasis | [133] |
| miR-30a | ROR1 | Associates with high histological grade and more lymph node metastasis | [134] |
| miR-190a/940 | Unknown | Prevents metastasis and cell invasion | [19,135] |
| miR-33b | HMGA2, SALL4, Twist1 | Inhibit metastasis | [114] |
| miR-146a-5p | SOX5 | Inhibits proliferation and metastasis | [136] |
| miR-150 | HMGA2 | Inhibits metastasis | [137] |
| miR-124 | ZEB2 | Inhibits invasion and metastasis | [138] |
| miR-148a | WNT1, NRP1 | Suppress metastasis | [139] |
| miR-126-3p | RGS3 | Inhibits proliferation, migration, invasion, and angiogenesis | [140] |
| miR-508-3p | ZEB1 | Inhibits cell invasion and EMT | [141] |
| miR-613 | Daam1 | Inhibits cell migration and invasion | [142] |
| miR-519d-3p | LIMK1 | Suppresses growth and motility | [143] |
| miR-26a | MTDH | Suppresses proliferation and metastasis | [144] |
| miR-130a | FOSL1, ZO-1 | Suppress migration and invasion | [145] |
| miRNA | Direct/Indirect Targets | Functions | Reference |
|---|---|---|---|
| Oncogenic miRNAs | |||
| miR-182 | PFN1 | Promotes TNBC cell proliferation | [115] |
| miR-21 | PTEN | Promotes TNBC cell proliferation | [90] |
| miR-206 | RASA1, SPRED1 | Promote TNBC cell proliferation | [47] |
| miR-146a/146b-5p | BRCA1 | Increases BRCA1-mediated proliferation | [146] |
| miR-498 | BRCA1 | Promotes TNBC cell proliferation | [147] |
| miR-25-3p | BTG2 | Promotes TNBC cell proliferation | [148] |
| miR-20a-5p | RUNX3, Bim, p21 | Promote TNBC cell proliferation | [149] |
| miR-135b | APC | Promotes proliferation and metastasis | [150] |
| miR-502 | STE8 | Promotes cell proliferation and cell cycle | [25] |
| Oncosuppressor miRNAs | |||
| miR-203 | BIRC5, LASP1 | Suppress cell proliferation and migration | [151] |
| miR-205 | ErbB3, VEGF-A, E2F1, LAMC1 | Inhibit cell proliferation, cell invasion, cell cycle arrest, clonogenic potential | [152,153] |
| miR-200c | XIAP | Inhibits proliferation, induces apoptosis | [126] |
| miR-34a | Notch-1, ErbB2, c-SRC | Inhibit cell growth and invasion, activate senescence, sensitize to dasatinib | [154,155,156] |
| miR-940 | ZNF24 | Inhibits cell proliferation and migration | [157] |
| miR-211-5p | SETBP1 | Inhibits cell proliferation and migration | [158] |
| miR-1301 | EZH2 | Suppresses proliferation, migration, and colony formation | [159] |
| miR-146a-5p | SOX5 | Inhibits the proliferation and metastasis | [136] |
| miR-26a | MTDH | Suppresses tumor proliferation and metastasis | [144] |
| miR-490-3p | TNKS2 | Inhibits the growth and invasiveness | [160] |
| miR-143-3p | LIMK1 | Suppresses the growth | [161] |
| miR-17-5p | ETV1 | Suppresses cell proliferation and invasion | [162] |
| miR-539 | LAMA4 | Inhibits proliferation and migration | [163] |
| miR-125b | MAP2K7 | Inhibits proliferation | [164] |
| miR-217 | KLF5 | Inhibits cell growth, migration | [165] |
| miR-589 | MTA2 | Decreases cell proliferation, migration, and invasion | [166] |
| MiRNA | Direct/Indirect Targets | Functions | Reference |
|---|---|---|---|
| Oncogenic miRNAs | |||
| miR-21 | PTEN | Promotes the tumor proliferation and inhibits cell apoptosis | [90] |
| miR-182 | PFN1, RIP1 | Promote the tumor proliferation, inhibit cell apoptosis and migration | [115,168] |
| miR-301b | CYLD | Inhibits cell apoptosis induced by 5-FU | [169] |
| miR-155-5p | FOXO3A | Promotes proliferation and reduces bufalin-induced apoptosis | [170] |
| miR-17-5p/20a | DR4/DR5 | Inhibit apoptosis | [171] |
| miR-429 | XIAP | Regulates apoptosis | [172] |
| Oncosuppressor miRNAs | |||
| miR-200c | XIAP | Induces apoptosis | [126] |
| miR-31 | Bcl-2, PKCε | Induce apoptosis, increase sensitivity of chemo- and radiosensitivity | [173] |
| miR-4458 | SOCS1 | Inhibits proliferation and promotes apoptosis | [174] |
| miR-890 | CD147 | Inhibits the cell proliferation and invasion, induces apoptosis | [175] |
| miR-509 | TNF-α | Increases apoptosis and inhibits invasion | [176] |
| miR-145 | cIAP1 | Promotes TNF-α-induced apoptosis | [177] |
| miR-199a-5p | TGF-β2, PIK3CD | Proliferate inhibition, cell cycle arrest, and increase apoptosis | [83] |
| miR-1296 | CCND1 | Suppresses proliferation, cell cycle arrest accompanied by induction of apoptosis | [178] |
| miR-10a | PIK3CA | Inhibits the proliferation and migration, promotes apoptosis | [179] |
| MiRNA | Targets | Functions | Prognostic | Reference |
|---|---|---|---|---|
| miR-9 | CDH1 | Invasion and metastasis | Highly expressed with poor disease-free and distant metastasis-free survival | [180] |
| miR-155 | VHL | Migration and invasion, tumor angiogenesis | Highly expressed with poor prognosis | [181,182] |
| miR-210 | ISCU1/2, SDHD | Mitochondrial dysfunction | Highly expressed with worse disease-free and overall survival | [183] |
| miR-493 | Unknown | Subclassify core basal and five negative phenotype subtypes | Highly expressed with better disease-free survival | [42,184] |
| miR-374a-5p | ARRB1 | Promotes cell survival, proliferation, and migration | Highly expressed with poor prognosis | [185] |
| miR-449 | CDK2, CCNE2 | Doxorubicin resistance | Elevated expression with better survival in chemotherapy-treated | [186] |
| miR-34 | GATA-X transcription factor | Tumor suppression | TC and CC alleles are associated with unfavorable prognosis | [187] |
| MiRNA | Targets | Functions | Drugs Involved | Reference |
|---|---|---|---|---|
| miR-5195-3p | EIF4A2 | Enhances the chemosensitivity | Paclitaxel | [188] |
| miR-18a | Dicer | Increases PTX IC50 and reduces PTX-induced cell apoptosis | Paclitaxel | [189] |
| miR-1207-5p | LZTS1 | Enhances cell growth arrest and cell apoptosis, a predictor of sensitivity towards Taxol | Taxol | [190] |
| miR-620 | DCTD | Induces cell apoptosis and cell growth arrest, facilitates the resistance of gemcitabine | Gemcitabine | [191] |
| miR-770 | STMN1 | Regulates apoptosis and tumor microenvironment, suppresses the doxorubicin resistance and metastasis | Doxorubicin | [192] |
| miR-15b/23a/26a/29a/ 106b/128/149/181a/192 /193b/195/324-3p/494 | Unknown | Mediates the anticancer effects of short-term starvation in doxorubicin-treated breast cancer cells | Doxorubicin | [193] |
| miR-197 | NLK | Enhances cisplatin sensitivity, cell proliferation inhibition | Cisplatin | [194] |
| miR-200c | Vimentin, E-cadherin | Inhibit cell migration and enhances chemosensitivity of mesenchymal by reversing their EMT-like property | Tamoxifen | [195] |
| miR-105/93-3p | SFRP1 | Promotes stemness, chemoresistance, and metastasis | Cisplatin | [196] |
| miR-21 | PTEN, PDCD4 | Sustain EMT and shape the tumor immune microenvironment, confer resistance to trastuzumab and chemotherapy | Trastuzumab | [197] |
| miR-95 | SGPP1 | Increass tumor growth and resistance to radiation treatment | Ionizing radiation | [198] |
| miR-302 | AKT1, RAD52 | Sensitize to radiotherapy | Ionizing radiation | [199] |
| miR-27a | CDC27 | Modulates proliferation and radiosensitivity | Irradiation treatment | [200] |
| miR-155 | RAD51 | Decreases the efficiency of homologous recombination DNA repair and enhances sensitivity to ionizing radiation | Ionizing radiation | [201] |
| miR-638 | BRCA1 | Reduces proliferation, invasive ability, and DNA repair capabilities | UV/cisplatin | [202] |
| miR-143-3p | CIAPIN1 | Effectively reverses multidrug resistance | Multidrug resistance | [203] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, L.; Gu, H.; Xiong, X.; Ao, H.; Cao, J.; Lin, W.; Yu, M.; Lin, J.; Cui, Q. MicroRNAs Involved in Carcinogenesis, Prognosis, Therapeutic Resistance, and Applications in Human Triple-Negative Breast Cancer. Cells 2019, 8, 1492. https://doi.org/10.3390/cells8121492
Ding L, Gu H, Xiong X, Ao H, Cao J, Lin W, Yu M, Lin J, Cui Q. MicroRNAs Involved in Carcinogenesis, Prognosis, Therapeutic Resistance, and Applications in Human Triple-Negative Breast Cancer. Cells. 2019; 8(12):1492. https://doi.org/10.3390/cells8121492
Chicago/Turabian StyleDing, Lei, Huan Gu, Xianhui Xiong, Hongshun Ao, Jiaqi Cao, Wen Lin, Min Yu, Jie Lin, and Qinghua Cui. 2019. "MicroRNAs Involved in Carcinogenesis, Prognosis, Therapeutic Resistance, and Applications in Human Triple-Negative Breast Cancer" Cells 8, no. 12: 1492. https://doi.org/10.3390/cells8121492
APA StyleDing, L., Gu, H., Xiong, X., Ao, H., Cao, J., Lin, W., Yu, M., Lin, J., & Cui, Q. (2019). MicroRNAs Involved in Carcinogenesis, Prognosis, Therapeutic Resistance, and Applications in Human Triple-Negative Breast Cancer. Cells, 8(12), 1492. https://doi.org/10.3390/cells8121492