Target-Specific Fluorescence-Mediated Tomography for Non-Invasive and Dynamic Assessment of Early Neutrophil Infiltration in Murine Experimental Colitis

 ,
,  and
and 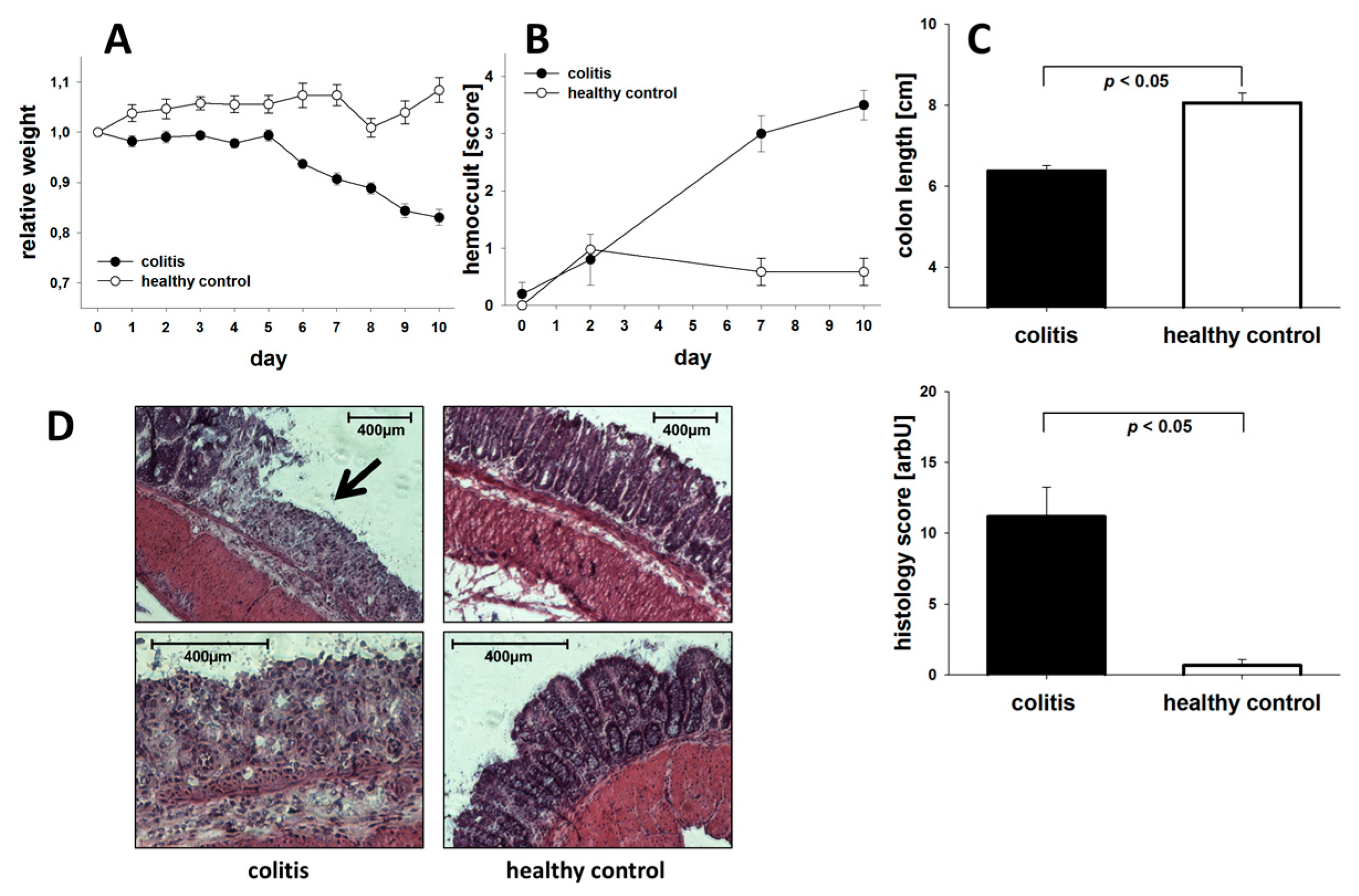{kind=link}
{kind=link}
{kind=link}
{kind=link}
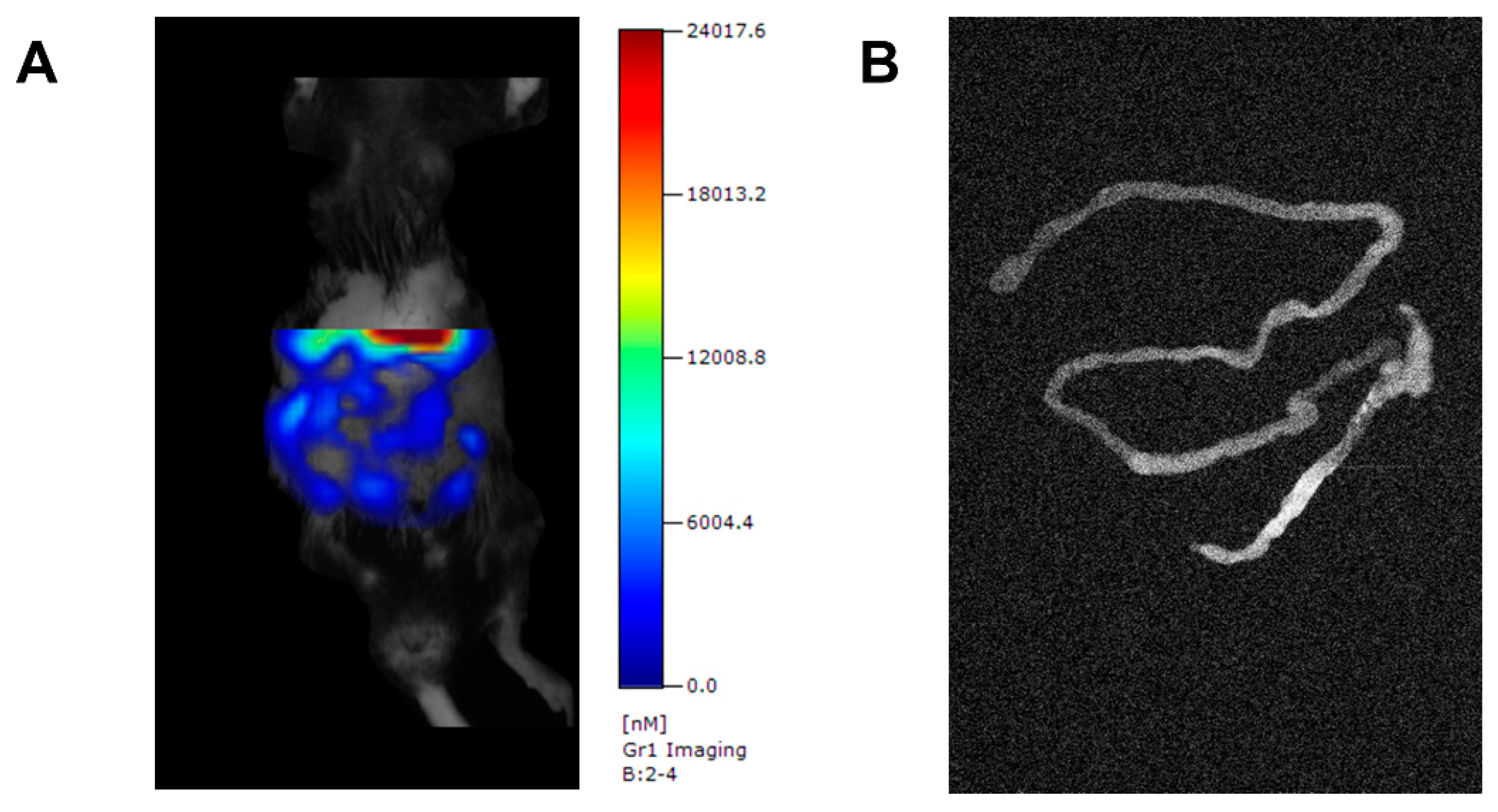{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Colitis Induction
2.2. Assessment of Colitis
2.3. Tissue Myeloperoxidase (MPO) Assay
2.4. Real Time (RT)-PCR Analysis
2.5. Fluorescence-Mediated Tomography
2.6. Endoscopy
2.7. Preparation of Leukocytes and Flow Cytometry
2.8. Experimental Design and Statistical Analysis
3. Results
3.1. General Assessment of Colitis
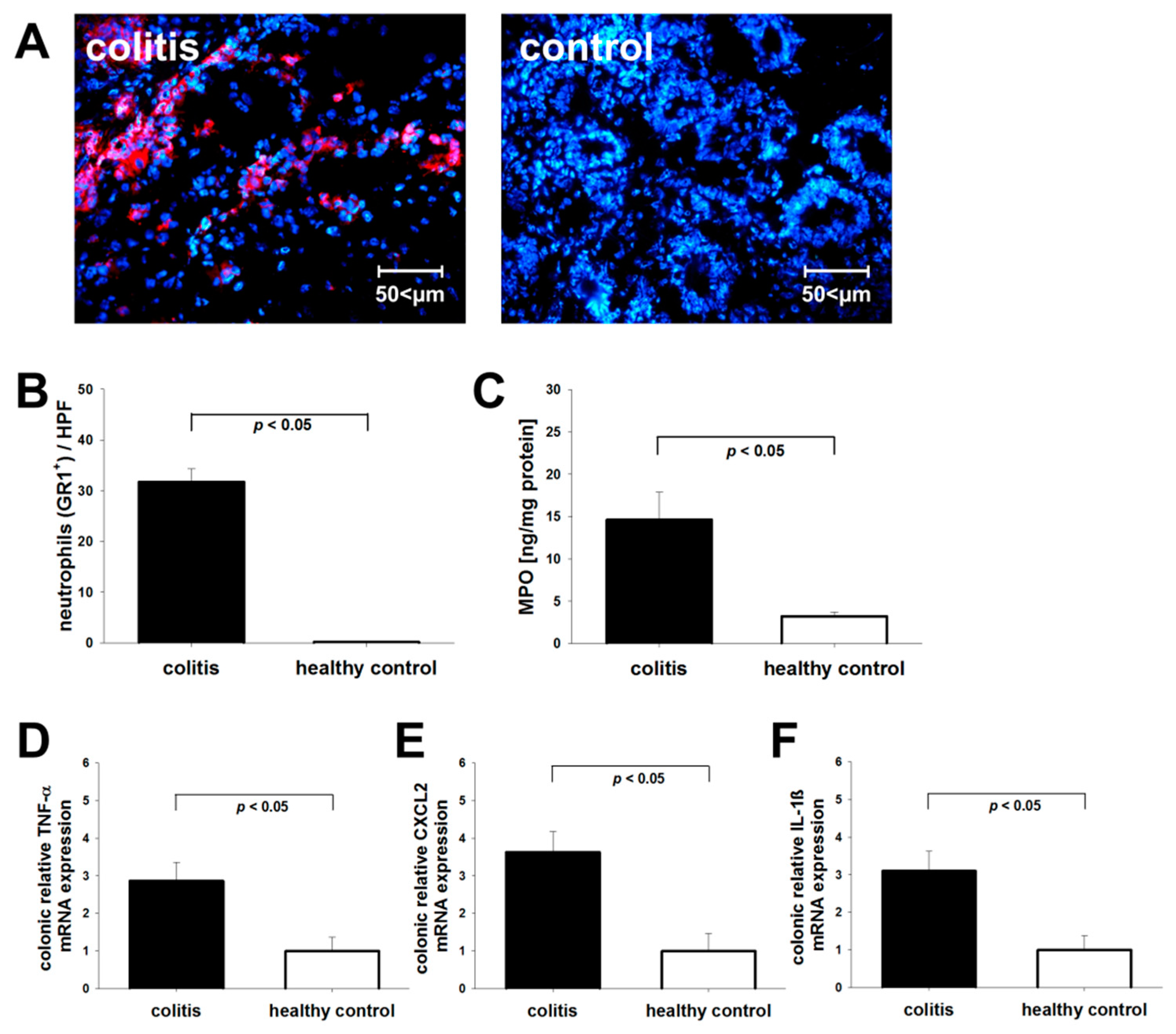
3.2. Post-Mortem Assessment of Neutrophil Recruitment
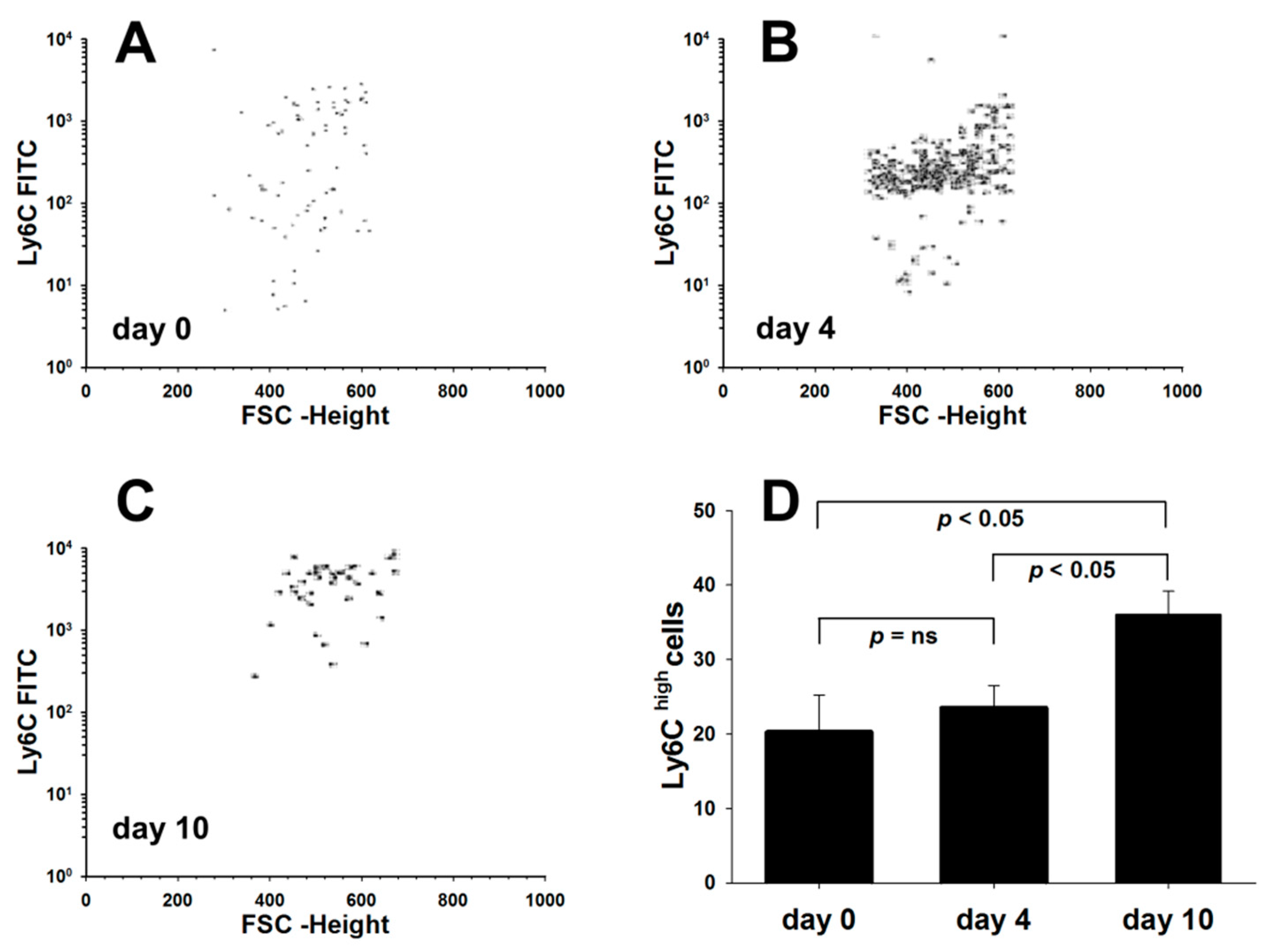
3.3. Dss-Colitis Induces Systemic Changes in Macrophage Subpopulations
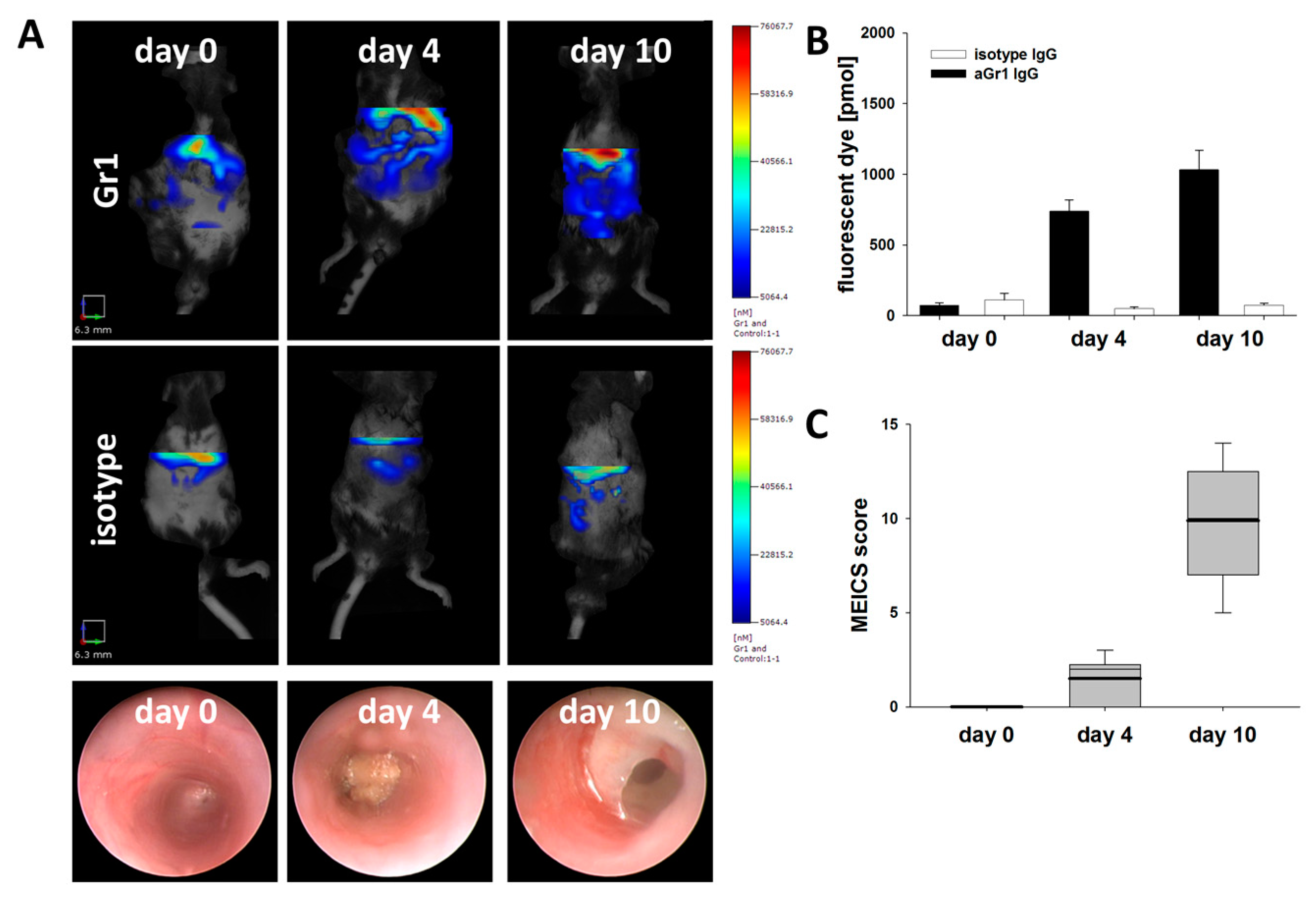
3.4. Fluorescence Mediated Tomography Scan Detects Early Neutrophil Infiltration in Murine Colitis
3.5. Specificity of FMT Detected Gr-1 Tracer Signal
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Kucharzik, T.; Maaser, C.; Lügering, A.; Kagnoff, M.; Mayer, L.; Targan, S.; Domschke, W. Recent understanding of IBD pathogenesis: Implications for future therapies. Inflamm. Bowel Dis. 2006, 12, 1068–1083. [Google Scholar] [CrossRef] [PubMed]
- Burisch, J.; Jess, T.; Martinato, M.; Lakatos, P.L. The burden of inflammatory bowel disease in Europe. J. Crohns Colitis 2013, 7, 322–337. [Google Scholar] [CrossRef] [PubMed]
- Habtezion, A.; Nguyen, L.P.; Hadeiba, H.; Butcher, E.C. Leukocyte Trafficking to the Small Intestine and Colon. Gastroenterology 2016, 150, 340–354. [Google Scholar] [CrossRef] [PubMed]
- Rutgeerts, P.; Sandborn, W.J.; Axler, J.; Kim, H.-J.; Danese, S.; Sankoh, S.; Wyant, T.; Feagan, B.G.; Sands, B.E.; Hanauer, S.; et al. Vedolizumab as Induction and Maintenance Therapy for Ulcerative Colitis. N. Engl. J. Med. 2013, 369, 699–710. [Google Scholar]
- Vermeire, S.; O’Byrne, S.; Keir, M.; Williams, M.; Lu, T.T.; Mansfield, J.C.; Lamb, C.A.; Feagan, B.G.; Panés, J.; Salas, A.; et al. Etrolizumab as induction therapy for ulcerative colitis: A randomised, controlled, phase 2 trial. Lancet 2014, 384, 309–318. [Google Scholar] [CrossRef]
- Nowacki, T.M.; Remaley, A.T.; Bettenworth, D.; Eisenblätter, M.; Vowinkel, T.; Becker, F.; Vogl, T.; Roth, J.; Tietge, U.J.; Lügering, A.; et al. The 5A apolipoprotein A-I (apoA-I) mimetic peptide ameliorates experimental colitis by regulating monocyte infiltration. Br. J. Pharmacol. 2016, 173, 2780–2792. [Google Scholar] [CrossRef]
- Rivera-Nieves, J.; Burcin, T.L.; Olson, T.S.; Morris, M.A.; McDuffie, M.; Cominelli, F.; Ley, K. Critical role of endothelial P-selectin glycoprotein ligand 1 in chronic murine ileitis. J. Exp. Med. 2006, 203, 907–917. [Google Scholar] [CrossRef]
- Sandborn, W.J.; Cyrille, M.; Hansen, M.B.; Feagan, B.G.; Loftus, E.V.; Rogler, G.; Vermeire, S.; Cruz, M.L.; Yang, J.; Boedigheimer, M.J.; et al. Efficacy and Safety of Abrilumab in a Randomized, Placebo-Controlled Trial for Moderate-to-Severe Ulcerative Colitis. Gastroenterology 2019, 156, 946–957.e18. [Google Scholar] [CrossRef]
- Targan, S.R.; Feagan, B.G.; Fedorak, R.N.; Lashner, B.A.; Panaccione, R.; Present, D.H.; Spehlmann, M.E.; Rutgeerts, P.J.; Tulassay, Z.; Volfova, M.; et al. Natalizumab for the Treatment of Active Crohn’s Disease: Results of the ENCORE Trial. Gastroenterology 2007, 132, 1672–1683. [Google Scholar] [CrossRef] [PubMed]
- Hindryckx, P.; Vande Casteele, N.; Novak, G.; Khanna, R.; D’Haens, G.; Sandborn, W.J.; Danese, S.; Jairath, V.; Feagan, B.G. The Expanding Therapeutic Armamentarium for Inflammatory Bowel Disease: How to Choose the Right Drug[s] for Our Patients? J. Crohns Colitis 2018, 12, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Fournier, B.M.; Parkos, C.A. The role of neutrophils during intestinal inflammation. Mucosal Immunol. 2012, 5, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Wera, O.; Lancellotti, P.; Oury, C. The Dual Role of Neutrophils in Inflammatory Bowel Diseases. J. Clin. Med. 2016, 5, 118. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, S.; Popp, V.; Kindermann, M.; Gerlach, K.; Weigmann, B.; Fichtner-Feigl, S.; Neurath, M.F. Chemically induced mouse models of acute and chronic intestinal inflammation. Nat. Protoc. 2017, 12, 1295–1309. [Google Scholar] [CrossRef] [PubMed]
- Brückner, M.; Lenz, P.; Nowacki, T.M.; Pott, F.; Foell, D.; Bettenworth, D. Murine Endoscopy for In Vivo Multimodal Imaging of Carcinogenesis and Assessment of Intestinal Wound Healing and Inflammation. J. Vis. Exp. 2014, 90. [Google Scholar] [CrossRef] [PubMed]
- Nowacki, T.M.; Bettenworth, D.; Brückner, M.; Cordes, F.; Lenze, F.; Becker, A.; Wildgruber, M.; Eisenblätter, M. Fluorescence-mediated Tomography for the Detection and Quantification of Macrophage-related Murine Intestinal Inflammation. J. Vis. Exp. 2017, 130. [Google Scholar] [CrossRef]
- Ntziachristos, V.; Bremer, C.; Weissleder, R. Fluorescence imaging with near-infrared light: New technological advances that enable in vivo molecular imaging. Eur. Radiol. 2003, 13, 195–208. [Google Scholar]
- Ntziachristos, V.; Tung, C.-H.; Bremer, C.; Weissleder, R. Fluorescence molecular tomography resolves protease activity in vivo. Nat. Med. 2002, 8, 757–761. [Google Scholar] [CrossRef]
- Ntziachristos, V.; Bremer, C.; Graves, E.E.; Ripoll, J.; Weissleder, R. In Vivo Tomographic Imaging of Near-Infrared Fluorescent Probes. Mol. Imaging 2002, 1, 82–88. [Google Scholar] [CrossRef]
- Fleming, T.J.; Fleming, M.L.; Malek, T.R. Selective expression of Ly-6G on myeloid lineage cells in mouse bone marrow. RB6-8C5 mAb to granulocyte-differentiation antigen (Gr-1) detects members of the Ly-6 family. J. Immunol. 1993, 151, 2399–2408. [Google Scholar] [PubMed]
- Dieleman, L.A.; Palmen, M.J.H.J.; Akol, H.; Bloemena, E.; Peña, A.S.; Meuwissen, S.G.M.; Van Rees, E.P. Chronic experimental colitis induced by dextran sulphate sodium (DSS) is characterized by Th1 and Th2 cytokines. Clin. Exp. Immunol. 1998, 114, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Kucharzik, T.; Hudson, J.T., 3rd; Lugering, A.; Abbas, J.A.; Bettini, M.; Lake, J.G.; Evans, M.E.; Ziegler, T.R.; Merlin, D.; Madara, J.L.; et al. Acute induction of human IL-8 production by intestinal epithelium triggers neutrophil infiltration without mucosal injury. Gut 2005, 54, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.; Fantini, M.C.; Neurath, M.F. High resolution colonoscopy in live mice. Nat. Protoc. 2006, 1, 2900–2904. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.; Fantini, M.C.; Wirtz, S.; Nikolaev, A.; Kiesslich, R.; Lehr, H.A.; Galle, P.R.; Neurath, M.F. In vivo imaging of colitis and colon cancer development in mice using high resolution chromoendoscopy. Gut 2005, 54, 950–954. [Google Scholar] [CrossRef]
- Sunderkötter, C.; Nikolic, T.; Dillon, M.J.; Van Rooijen, N.; Stehling, M.; Drevets, D.A.; Leenen, P.J.M. Subpopulations of mouse blood monocytes differ in maturation stage and inflammatory response. J. Immunol. 2004, 172, 4410–4417. [Google Scholar] [CrossRef]
- Segal, A.W. The role of neutrophils in the pathogenesis of Crohn’s disease. Eur. J. Clin. Investig. 2018, 48, e12983. [Google Scholar] [CrossRef]
- Mencarelli, A.; Cipriani, S.; Francisci, D.; Santucci, L.; Baldelli, F.; Distrutti, E.; Fiorucci, S. Highly specific blockade of CCR5 inhibits leukocyte trafficking and reduces mucosal inflammation in murine colitis. Sci. Rep. 2016, 6, 30802. [Google Scholar] [CrossRef]
- Perez-Jeldres, T.; Tyler, C.J.; Boyer, J.D.; Karuppuchamy, T.; Bamias, G.; Dulai, P.S.; Boland, B.S.; Sandborn, W.J.; Patel, D.R.; Rivera-Nieves, J. Cell Trafficking Interference in Inflammatory Bowel Disease: Therapeutic Interventions Based on Basic Pathogenesis Concepts. Inflamm. Bowel Dis. 2019, 25, 270–282. [Google Scholar] [CrossRef]
- Zundler, S.; Becker, E.; Weidinger, C.; Siegmund, B. Anti-Adhesion Therapies in Inflammatory Bowel Disease—Molecular and Clinical Aspects. Front. Immunol. 2017, 8, 891. [Google Scholar] [CrossRef]
- Sandborn, W.J.; Feagan, B.G.; Wolf, D.C.; D’Haens, G.; Vermeire, S.; Hanauer, S.B.; Ghosh, S.; Smith, H.; Cravets, M.; Frohna, P.A.; et al. Ozanimod Induction and Maintenance Treatment for Ulcerative Colitis. N. Engl. J. Med. 2016, 374, 1754–1762. [Google Scholar] [CrossRef]
- Nemoto, Y.; Kanai, T.; Tohda, S.; Totsuka, T.; Okamoto, R.; Tsuchiya, K.; Nakamura, T.; Sakamoto, N.; Fukuda, T.; Miura, O.; et al. Negative feedback regulation of colitogenic CD4+ T cells by increased granulopoiesis. Inflamm. Bowel Dis. 2008, 14, 1491–1503. [Google Scholar] [CrossRef] [PubMed]
- Raab, Y.; Gerdin, B.; Ahlstedt, S.; Hällgren, R. Neutrophil mucosal involvement is accompanied by enhanced local production of interleukin-8 in ulcerative colitis. Gut 1993, 34, 1203–1206. [Google Scholar] [CrossRef] [PubMed]
- Harbord, M.W.N.; Marks, D.J.B.; Forbes, A.; Bloom, S.L.; Day, R.M.; Segal, A.W. Impaired neutrophil chemotaxis in Crohn’s disease relates to reduced production of chemokines and can be augmented by granulocyte-colony stimulating factor. Aliment. Pharmacol. Ther. 2006, 24, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Vazeille, E.; Buisson, A.; Bringer, M.A.; Goutte, M.; Ouchchane, L.; Hugot, J.P.; de Vallee, A.; Barnich, N.; Bommelaer, G.; Darfeuille-Michaud, A. Monocyte-derived macrophages from Crohn’s disease patients are impaired in the ability to control intracellular adherent-invasive Escherichia coli and exhibit disordered cytokine secretion profile. J. Crohns Colitis 2015, 9, 410–420. [Google Scholar] [CrossRef]
- Couper, R.; Kapelushnik, J.; Griffiths, A.M. Neutrophil dysfunction in glycogen storage disease Ib: Association with Crohn’s-like colitis. Gastroenterology 1991, 100, 549–554. [Google Scholar] [CrossRef]
- Werlin, S.L.; Chusid, M.J.; Caya, J.; Oechler, H.W. Colitis in chronic granulomatous disease. Gastroenterology 1982, 82, 328–331. [Google Scholar] [CrossRef]
- Roe, T.F.; Thomas, D.W.; Gilsanz, V.; Isaacs, H.; Atkinson, J.B. Inflammatory bowel disease in glycogen storage disease type Ib. J. Pediatr. 1986, 109, 55–59. [Google Scholar] [CrossRef]
- Natsui, M.; Kawasaki, K.; Takizawa, H.; Hayashi, S.-I.; Matsuda, Y.; Sugimura, K.; Seki, K.; Narisawa, R.; Sendo, F.; Asakura, H. Selective depletion of neutrophils by a monoclonal antibody, RP-3, suppresses dextran sulphate sodium-induced colitis in rats. J. Gastroenterol. Hepatol. 1997, 12, 801–808. [Google Scholar] [CrossRef]
- Kühl, A.A.; Kakirman, H.; Janotta, M.; Dreher, S.; Cremer, P.; Pawlowski, N.N.; Loddenkemper, C.; Heimesaat, M.M.; Grollich, K.; Zeitz, M.; et al. Aggravation of Different Types of Experimental Colitis by Depletion or Adhesion Blockade of Neutrophils. Gastroenterology 2007, 133, 1882–1892. [Google Scholar] [CrossRef]
- Baggiolini, M.; Horisberger, U.; Gennaro, R.; Dewald, B. Identification of three types of granules in neutrophils of ruminants. Ultrastructure of circulating and maturing cells. Lab. Investig. 1985, 52, 151–158. [Google Scholar]
- Kiesler, P.; Fuss, I.J.; Strober, W. Experimental Models of Inflammatory Bowel Diseases. Cell. Mol. Gastroenterol. Hepatol. 2015, 1, 154–170. [Google Scholar] [CrossRef] [PubMed]
- Wagnerova, A.; Babickova, J.; Liptak, R.; Vlkova, B.; Celec, P.; Gardlik, R. Sex Differences in the Effect of Resveratrol on DSS-Induced Colitis in Mice. Gastroenterol. Res. Pr. 2017, 2017, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mahler, M.; Bristol, I.J.; Leiter, E.H.; Workman, A.E.; Birkenmeier, E.H.; Elson, C.O.; Sundberg, J.P. Differential susceptibility of inbred mouse strains to dextran sulfate sodium-induced colitis. Am. J. Physiol. Liver Physiol. 1998, 274, G544–G551. [Google Scholar] [CrossRef] [PubMed]
- Krieglstein, C.F.; Cerwinka, W.H.; Sprague, A.G.; Koteliansky, V.E.; Senninger, N.; Laroux, F.S.; Grisham, M.B.; Granger, D.N.; De Fougerolles, A.R. Collagen-binding integrin α1β1 regulates intestinal inflammation in experimental colitis. J. Clin. Investig. 2002, 110, 1773–1782. [Google Scholar] [CrossRef]
- Weissleder, R.; Mahmood, U. Molecular Imaging. Radiology 2001, 219, 316–333. [Google Scholar] [CrossRef]
- Ntziachristos, V.; Ripoll, J.; Wang, L.V.; Weissleder, R. Looking and listening to light: The evolution of whole-body photonic imaging. Nat. Biotechnol. 2005, 23, 313–320. [Google Scholar] [CrossRef]
- Stuker, F.; Ripoll, J.; Rudin, M. Fluorescence Molecular Tomography: Principles and Potential for Pharmaceutical Research. Pharmaceutics 2011, 3, 229–274. [Google Scholar] [CrossRef]
- Bézière, N.; Ntziachristos, V. Optoacoustic Imaging: An Emerging Modality for the Gastrointestinal Tract. Gastroenterology 2011, 141, 1979–1985. [Google Scholar] [CrossRef]
- Ribechini, E.; Leenen, P.J.M.; Lutz, M.B. Gr-1 antibody induces STAT signaling, macrophage marker expression and abrogation of myeloid-derived suppressor cell activity in BM cells. Eur. J. Immunol. 2009, 39, 3538–3551. [Google Scholar] [CrossRef]
- Chames, P.; Van Regenmortel, M.; Weiss, E.; Baty, D. Therapeutic antibodies: Successes, limitations and hopes for the future. Br. J. Pharmacol. 2009, 157, 220–233. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowacki, T.M.; Lenz, P.; Bettenworth, D.; Brückner, M.; Bokemeyer, A.; Tepasse, P.R.; Helfen, A.; Wildgruber, M.; Eisenblätter, M. Target-Specific Fluorescence-Mediated Tomography for Non-Invasive and Dynamic Assessment of Early Neutrophil Infiltration in Murine Experimental Colitis. Cells 2019, 8, 1328. https://doi.org/10.3390/cells8111328
Nowacki TM, Lenz P, Bettenworth D, Brückner M, Bokemeyer A, Tepasse PR, Helfen A, Wildgruber M, Eisenblätter M. Target-Specific Fluorescence-Mediated Tomography for Non-Invasive and Dynamic Assessment of Early Neutrophil Infiltration in Murine Experimental Colitis. Cells. 2019; 8(11):1328. https://doi.org/10.3390/cells8111328
Chicago/Turabian StyleNowacki, Tobias M., Philipp Lenz, Dominik Bettenworth, Markus Brückner, Arne Bokemeyer, Phil R. Tepasse, Anne Helfen, Moritz Wildgruber, and Michel Eisenblätter. 2019. "Target-Specific Fluorescence-Mediated Tomography for Non-Invasive and Dynamic Assessment of Early Neutrophil Infiltration in Murine Experimental Colitis" Cells 8, no. 11: 1328. https://doi.org/10.3390/cells8111328
APA StyleNowacki, T. M., Lenz, P., Bettenworth, D., Brückner, M., Bokemeyer, A., Tepasse, P. R., Helfen, A., Wildgruber, M., & Eisenblätter, M. (2019). Target-Specific Fluorescence-Mediated Tomography for Non-Invasive and Dynamic Assessment of Early Neutrophil Infiltration in Murine Experimental Colitis. Cells, 8(11), 1328. https://doi.org/10.3390/cells8111328