Silencing Heat Shock Protein 27 Inhibits the Progression and Metastasis of Colorectal Cancer (CRC) by Maintaining the Stability of Stromal Interaction Molecule 1 (STIM1) Proteins



Abstract

1. Introduction
2. Materials and Methods
2.1. Chemicals, Reagents, and Cell Culture
2.2. Tissue Samples and Immunohistochemistry
2.3. HSP27 Expression Manipulation in Colon Cancer Cells
2.4. Protein Extraction and Western Blot Analysis
2.5. Cell Cycle Analysis by Flow Cytometry
2.6. Evaluation of Cell Proliferation and Cell Migration Using the Xcelligence Biosensor System
2.7. SRB Colorimetric Assay for Cytotoxicity Screening Stain
2.8. In Vivo Tumor Xenograft Experiments
2.9. Transwell Migration Assay and Invasion Assay
2.10. Determination of Calcium Concentrations
2.11. Immunocytochemistry
2.12. Immunoprecipitation Assay
2.13. RT-PCR and Quantitative RT-PCR Analysis
2.14. Statistical Analysis
3. Results
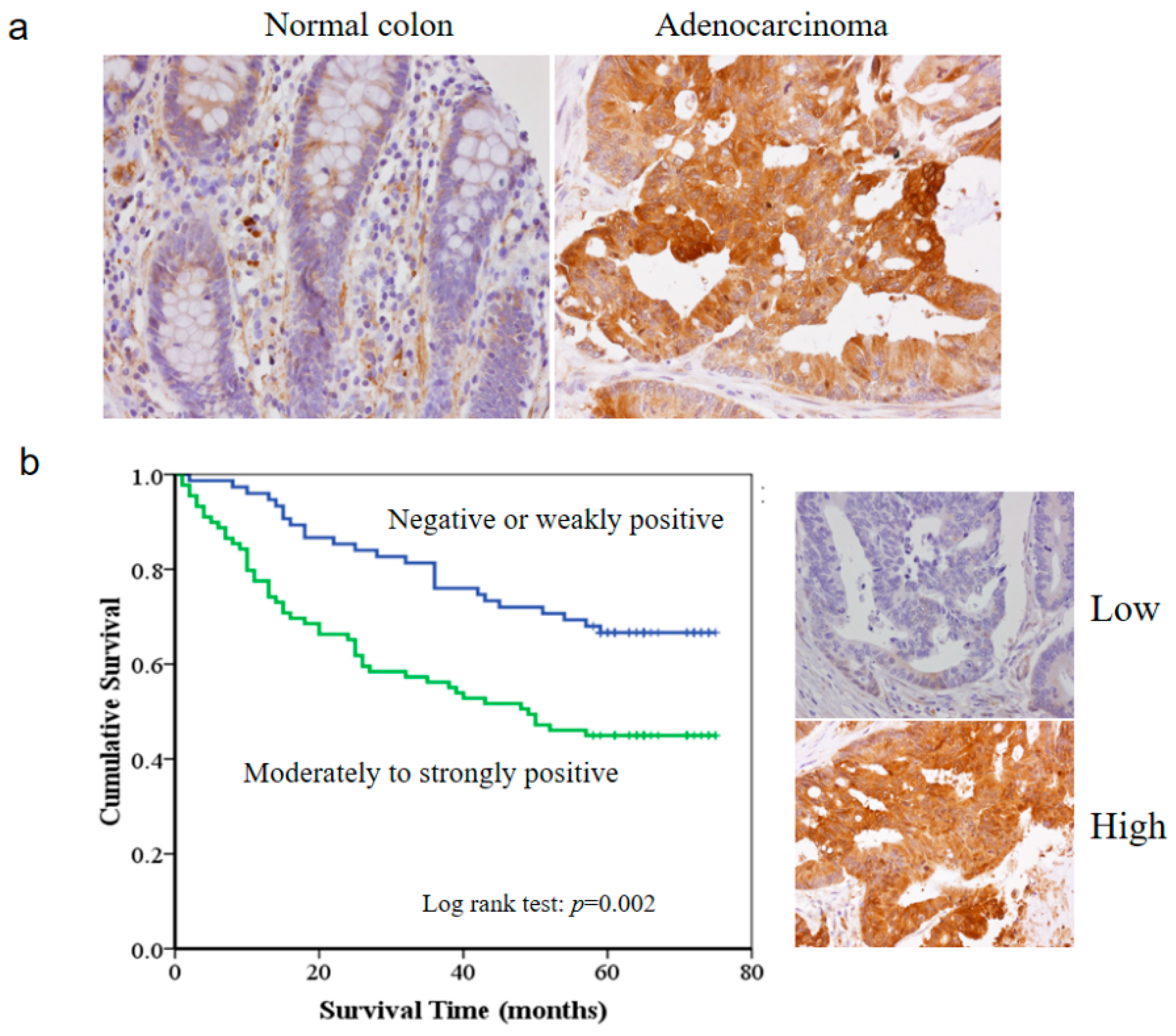
3.1. The HSP27 Expression and Clinicopathological Significance in Colon Adenocarcinoma
3.2. HSP27 Expression and Roles in Cell Growth of CRC Cell Lines
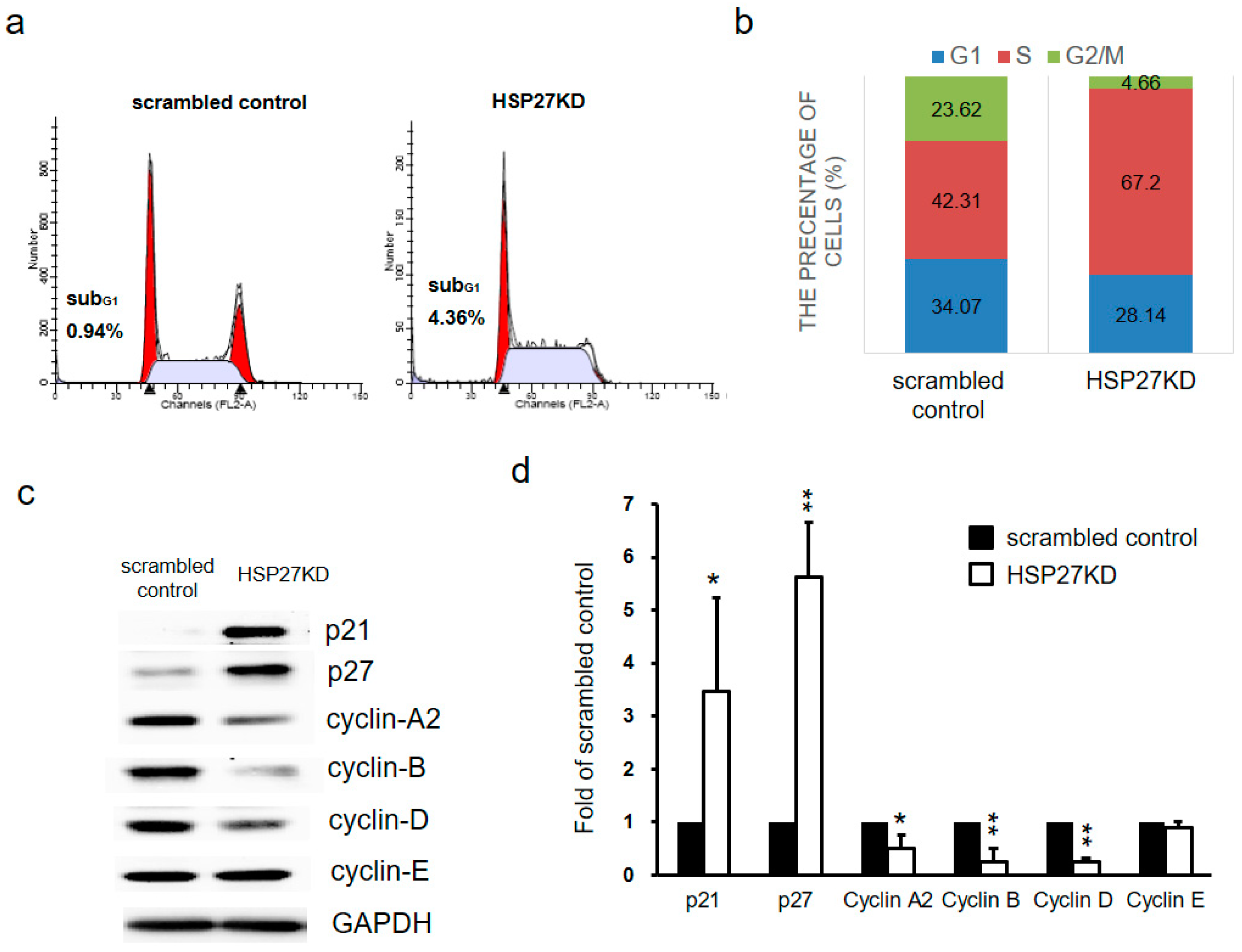
3.3. Silencing HSP27 Decreased Cell Proliferation via Cell Cycle Arrest
3.4. Silencing HSP27 Caused Cell Cycle Arrest at S phase
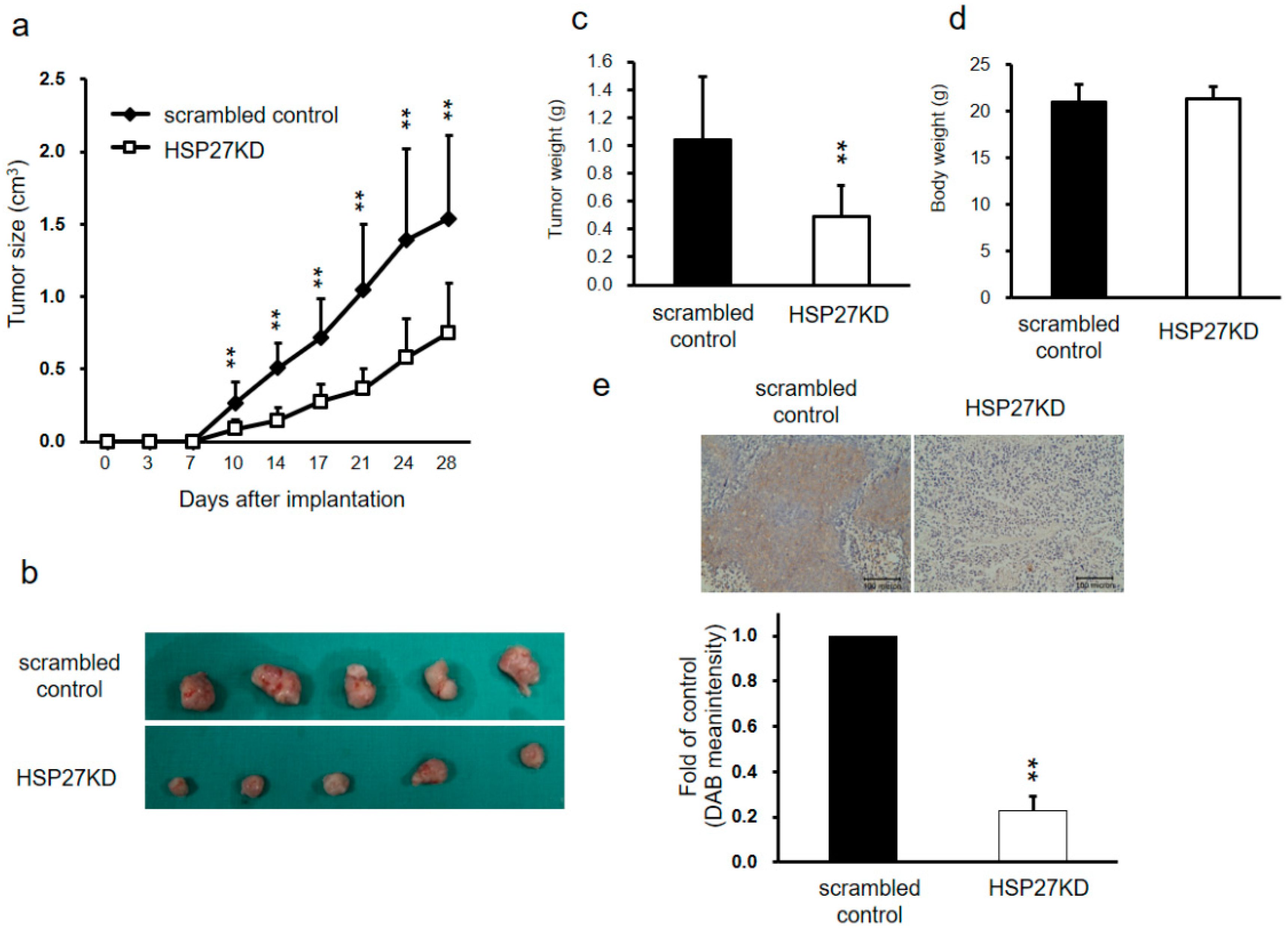
3.5. HSP27 Regulated Cancer Progression in Xenograft Mouse Models
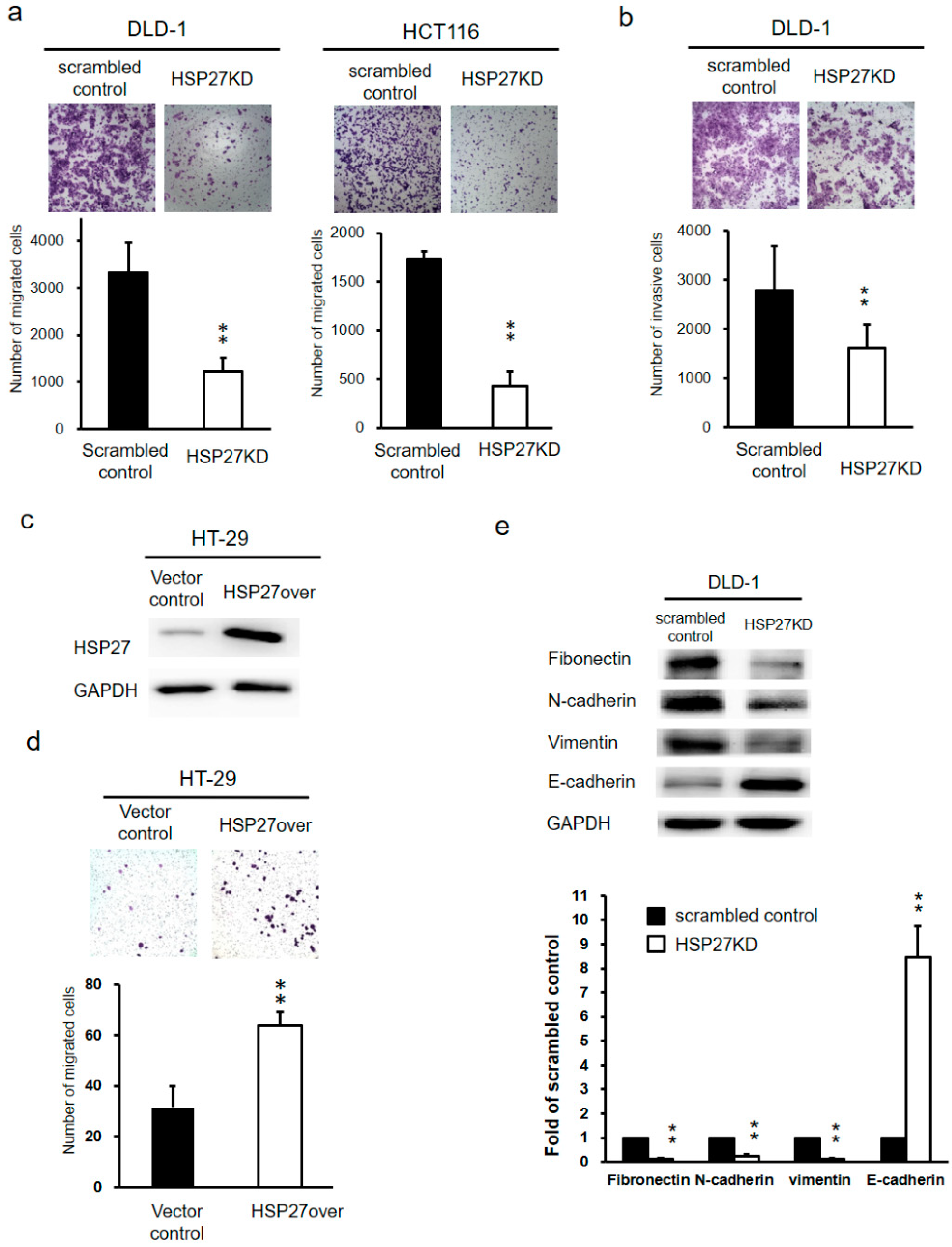
3.6. Silencing HSP27 Expression Inhibited Cell Migration and Invasion of CRC cells
3.7. Silencing HSP27 Influenced the Epithelial-Mesenchymal Transition (EMT)
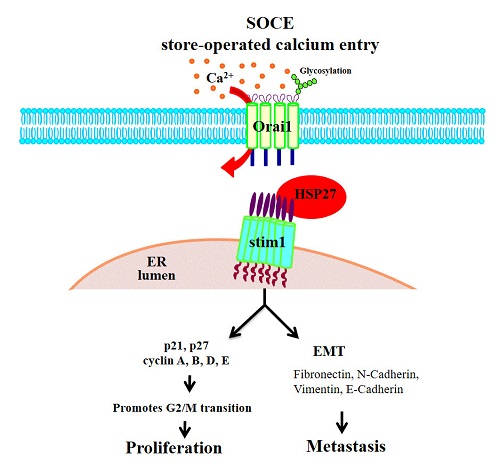
3.8. Silencing HSP27 Expression Abolished Calcium Channel Signals
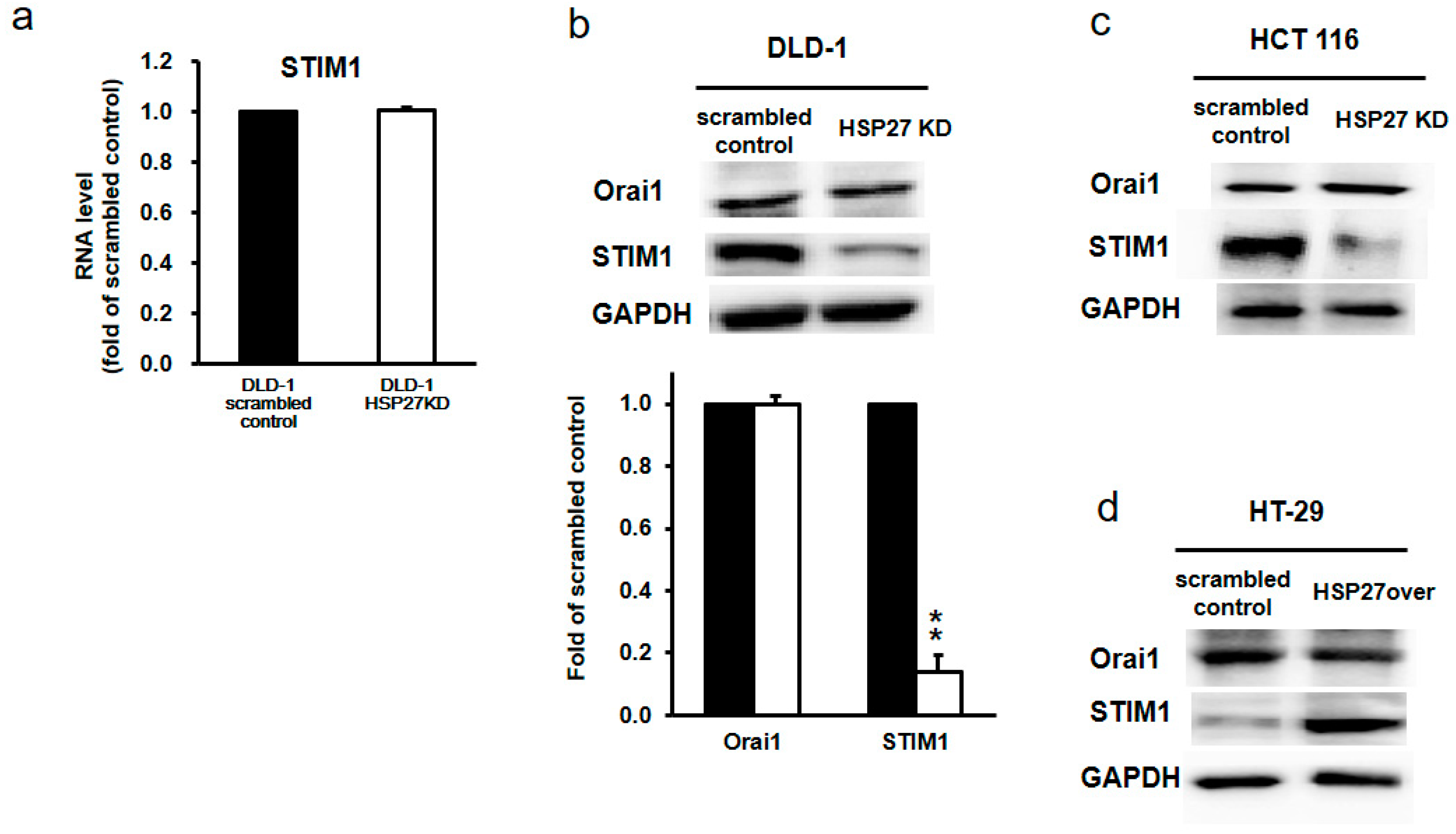
3.9. Silencing HSP27 Reduced STIM1 Levels
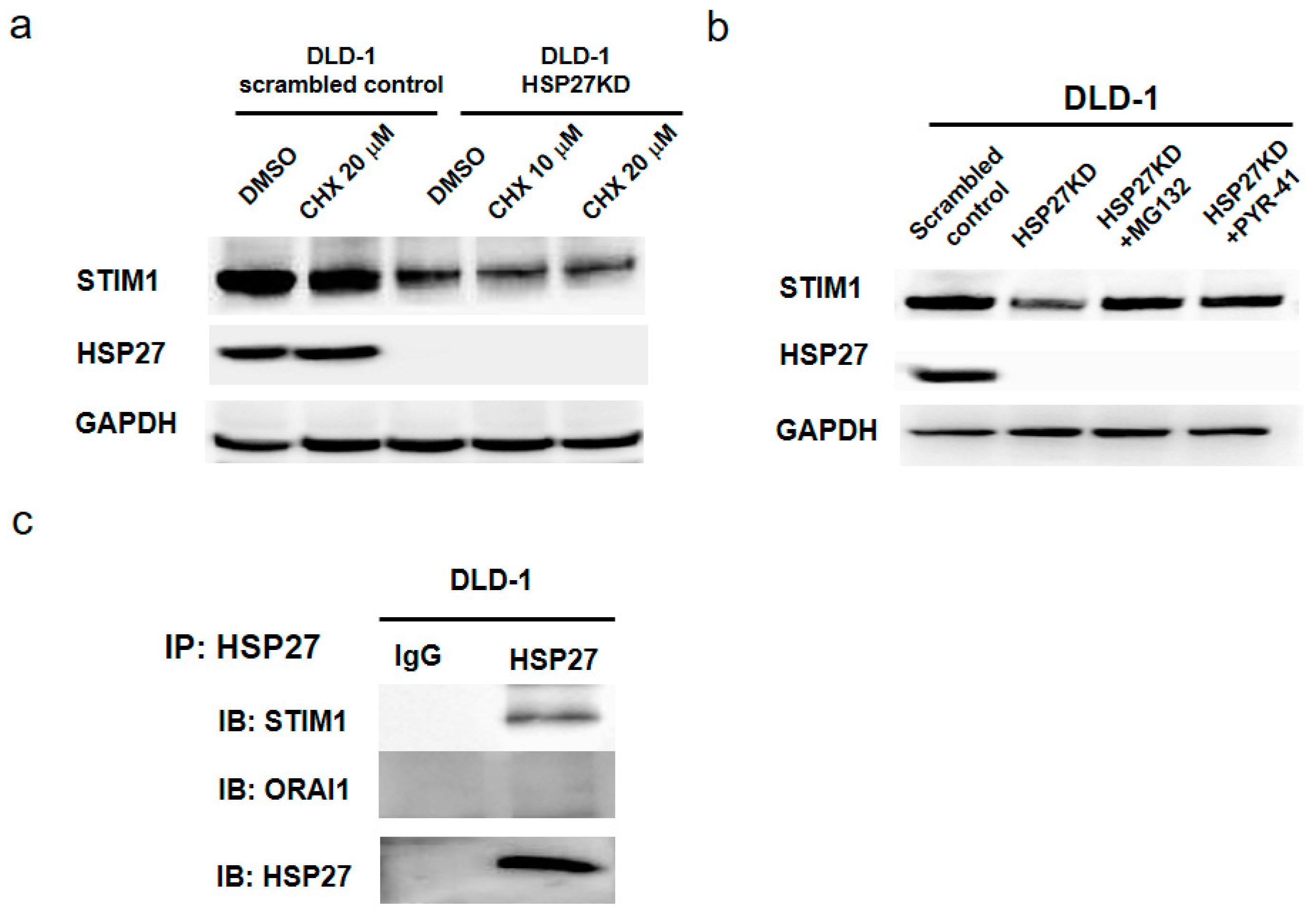
3.10. HSP27 Expression Maintained the Stability of STIM1 Proteins
3.11. STIM1 Was the Client Protein of HSP27
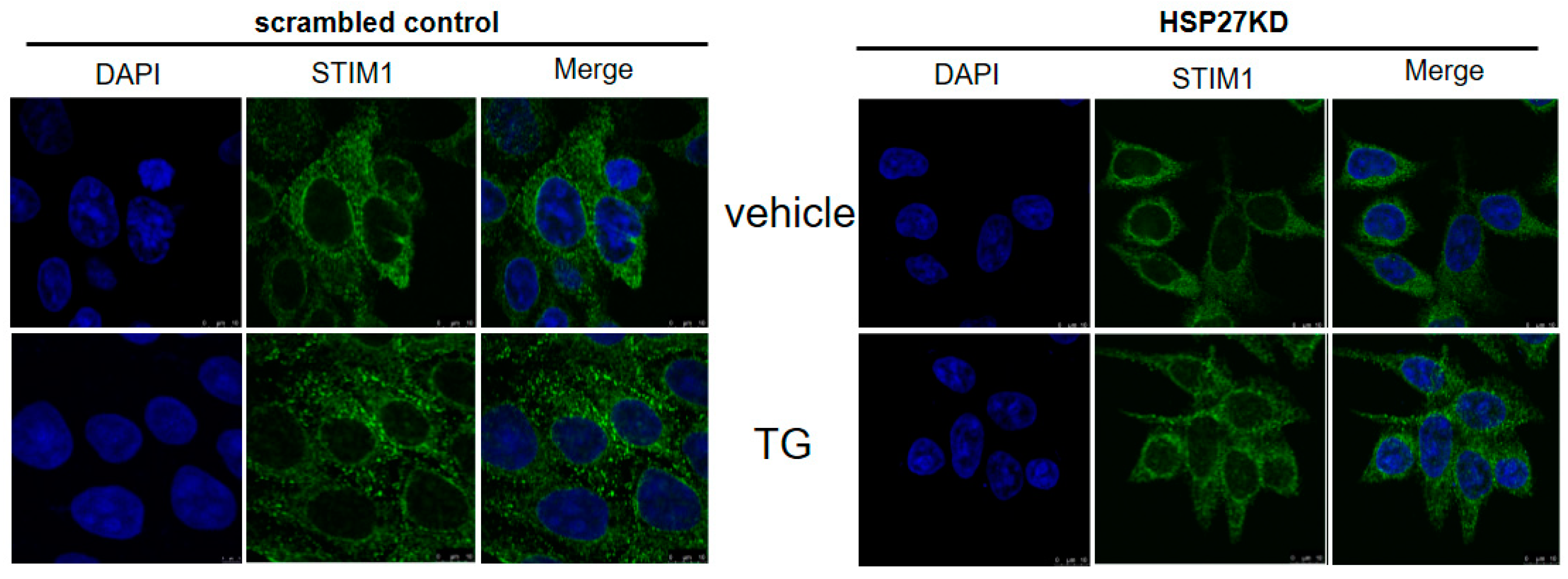
3.12. Silencing HSP27 Caused a Decrease in Puncta Formation of STIM1 under TG Induction
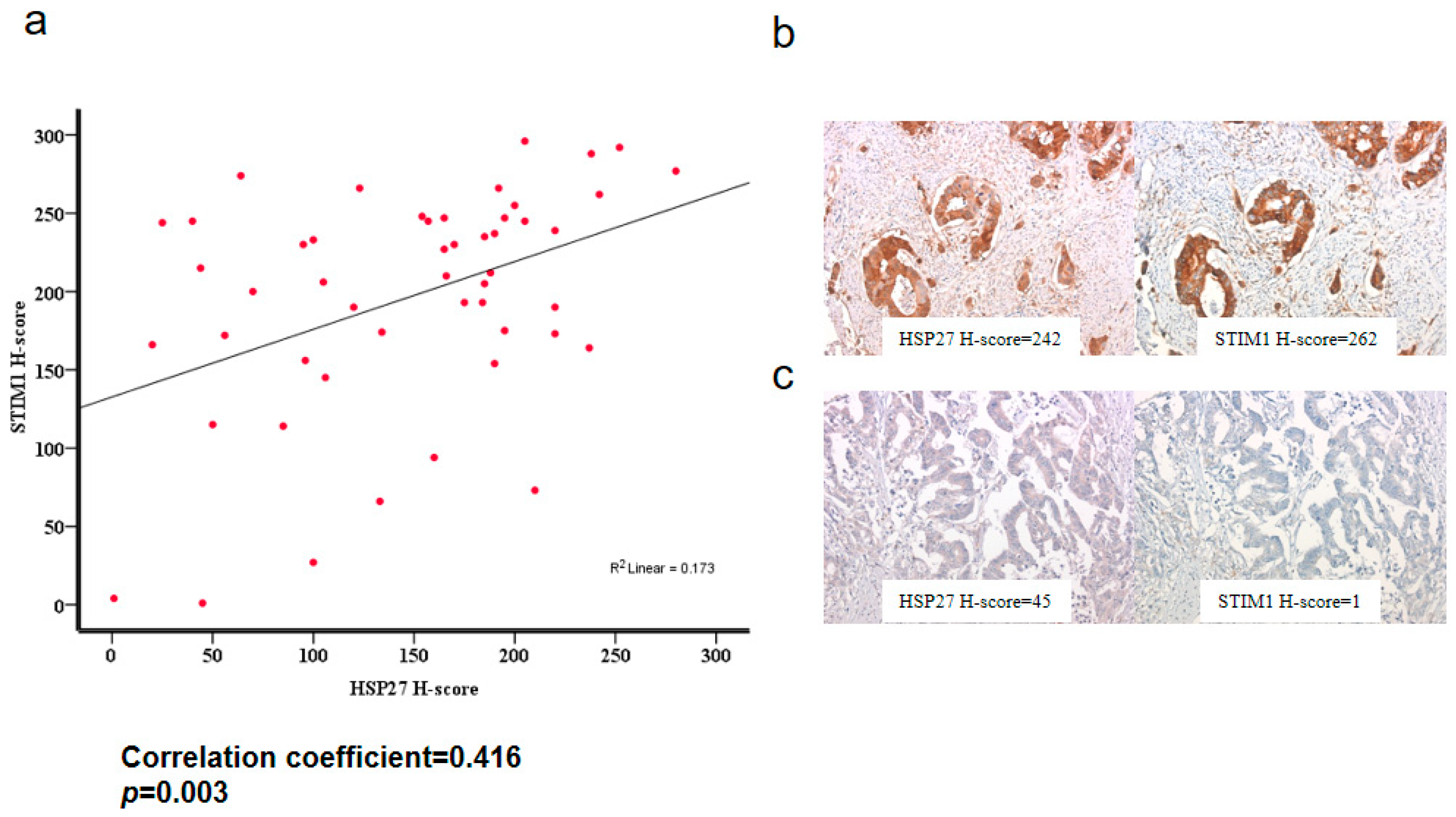
3.13. HSP27 Was Associated with STIM1 Expression in CRC Specimens
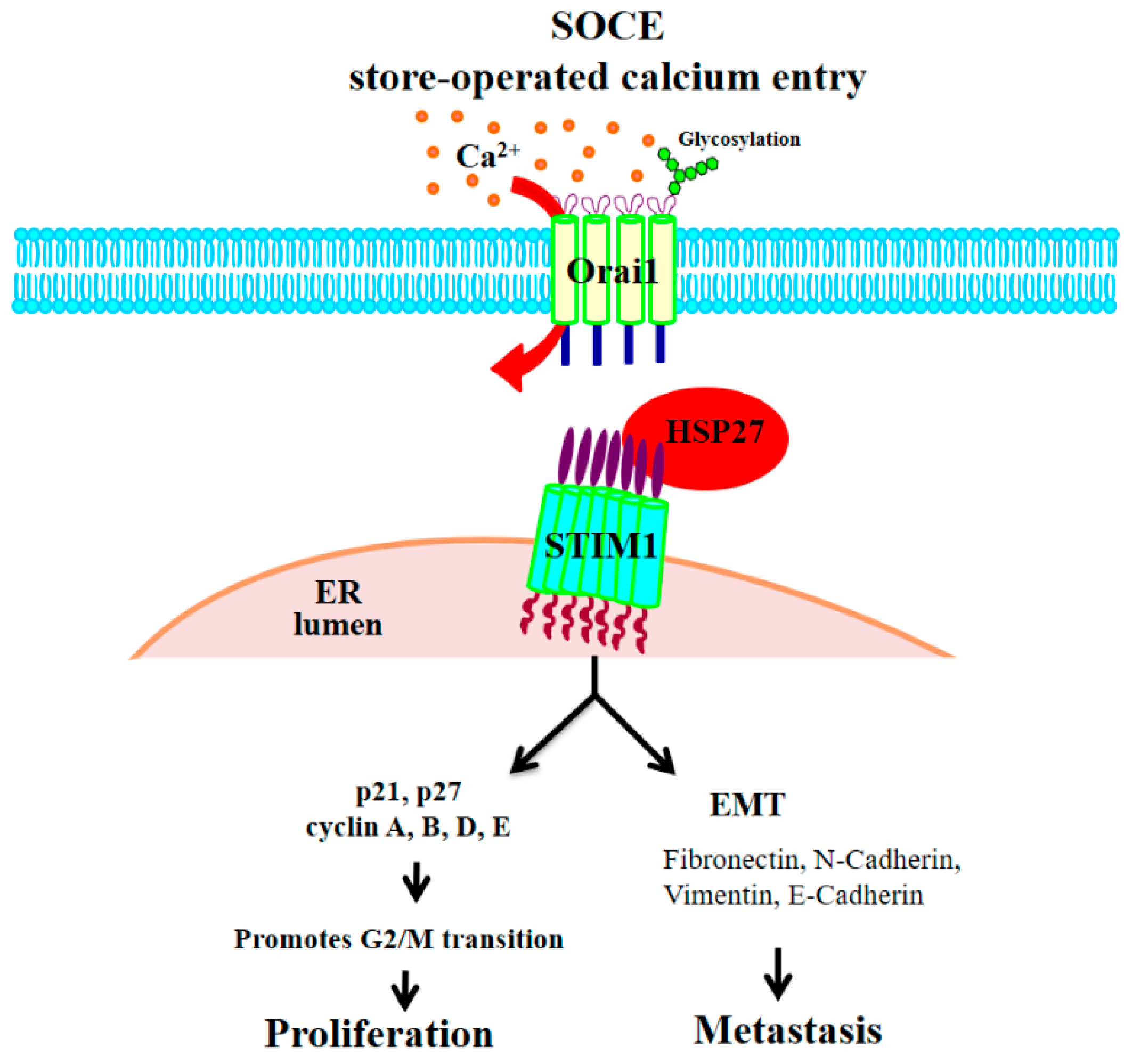
4. Discussion
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, D.; Atkin, W.; Lenz, H.J.; Lynch, H.T.; Minsky, B.; Nordlinger, B.; Starling, N. Colorectal cancer. Lancet 2010, 375, 1030–1047. [Google Scholar] [CrossRef]
- Fahy, B.N.; D’Angelica, M.; DeMatteo, R.P.; Blumgart, L.H.; Weiser, M.R.; Ostrovnaya, I.; Gonen, M.; Jarnagin, W.R. Synchronous hepatic metastases from colon cancer: Changing treatment strategies and results of surgical intervention. Ann. Surg. Oncol. 2009, 16, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Schuller, J.; Cassidy, J.; Dumont, E.; Roos, B.; Banken, L.; Mori, K.; Reigner, B.; Utoh, M.; Weidekamm, E.; Durston, S. Preferential activation of capecitabine in tumor following oral administration to colorectal cancer patients. Cancer Chemother. Pharmacol. 2000, 45, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.C.; Huang, C.Y.; Pan, T.L.; Chen, W.Y.; Ho, C.T.; Liu, T.Z.; Chang, Y.J. Proteomic Characterization of Annexin l (ANX1) and Heat Shock Protein 27 (HSP27) as Biomarkers for Invasive Hepatocellular Carcinoma Cells. PLoS ONE 2015, 10, e0139232. [Google Scholar] [CrossRef]
- Thomaidis, T.; Maderer, A.; Formentini, A.; Bauer, S.; Trautmann, M.; Schwarz, M.; Neumann, W.; Kittner, J.M.; Schad, A.; Link, K.-H.; et al. Proteins of the vegfr and egfr pathway as predictive markers for adjuvant treatment in patients with stage ii/iii colorectal cancer: Results of the fogt-4 trial. J. Exp. Clin. Cancer Res. 2014, 33. [Google Scholar] [CrossRef]
- Schweiger, T.; Nikolowsky, C.; Starlinger, P.; Traxler, D.; Zimmermann, M.; Birner, P.; Hegedüs, B.; Dome, B.; Bergmann, M.; Mildner, M.; et al. Stromal expression of heat-shock protein 27 is associated with worse clinical outcome in patients with colorectal cancer lung metastases. PLoS ONE 2015, 10, e0120724. [Google Scholar] [CrossRef]
- Kampinga, H.H.; Hageman, J.; Vos, M.J.; Kubota, H.; Tanguay, R.M.; Bruford, E.A.; Cheetham, M.E.; Chen, B.; Hightower, L.E. Guidelines for the nomenclature of the human heat shock proteins. Cell Stress Chaperones 2009, 14, 105–111. [Google Scholar] [CrossRef]
- Bakthisaran, R.; Tangirala, R.; Rao Ch, M. Small heat shock proteins: Role in cellular functions and pathology. Biochim. Biophys. Acta 2015, 1854, 291–319. [Google Scholar] [CrossRef]
- Zoubeidi, A.; Gleave, M. Small heat shock proteins in cancer therapy and prognosis. Int. J. Biochem. Cell Biol. 2012, 44, 1646–1656. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, Y.; Guo, K.; Wang, N.; Jin, H.; Liu, Y.; Qin, W. Heat shock proteins in hepatocellular carcinoma: Molecular mechanism and therapeutic potential. Int. J. Cancer 2016, 138, 1824–1834. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Ye, J.; Huang, Q.; Chen, W.; Wang, L.; Lin, W.; Lin, J.; Lin, X. Heat shock protein 27 is over-expressed in tumor tissues and increased in sera of patients with gastric adenocarcinoma. Clin. Chem. Lab. Med. 2010, 48, 263–269. [Google Scholar] [CrossRef]
- Arts, H.J.; Hollema, H.; Lemstra, W.; Willemse, P.H.; De Vries, E.G.; Kampinga, H.H.; Van der Zee, A.G. Heat-shock-protein-27 (hsp27) expression in ovarian carcinoma: Relation in response to chemotherapy and prognosis. Int. J. Cancer 1999, 84, 234–238. [Google Scholar] [CrossRef]
- Conroy, S.; Sasieni, P.; Amin, V.; Wang, D.; Smith, P.; Fentiman, I.; Latchman, D. Antibodies to heat-shock protein 27 are associated with improved survival in patients with breast cancer. Columbia J. Cancer 1998, 77, 1875–1879. [Google Scholar] [CrossRef]
- Bubendorf, L.; Kolmer, M.; Kononen, J.; Koivisto, P.; Mousses, S.; Chen, Y.; Mahlamaki, E.; Schraml, P.; Moch, H.; Willi, N.; et al. Hormone therapy failure in human prostate cancer: Analysis by complementary dna and tissue microarrays. JNCI J. Natl. Cancer Inst. 1999, 91, 1758–1764. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, P. Increased hsp27 after androgen ablation facilitates androgen-independent progression in prostate cancer via signal transducers and activators of transcription 3-mediated suppression of apoptosis. Cancer Res. 2005, 65, 11083–11093. [Google Scholar] [CrossRef]
- Ciocca, D.R.; Calderwood, S.K. Heat shock proteins in cancer: Diagnostic, prognostic, predictive, and treatment implications. Cell Stress Chaperones 2005, 10, 86–103. [Google Scholar] [CrossRef]
- Doshi, B.M.; Hightower, L.E.; Lee, J. The role of hsp27 and actin in the regulation of movement in human cancer cells responding to heat shock. Cell Stress Chaperones 2009, 14, 445–457. [Google Scholar] [CrossRef]
- Stope, M.; Weiss, M.; Preuss, M.; Streitbörger, A.; Ritter, C.; Zimmermann, U.; Walther, R.; Burchardt, M. Immediate and transient phosphorylation of the heat shock protein 27 initiates chemoresistance in prostate cancer cells. Oncol. Rep. 2014, 32, 2380–2386. [Google Scholar] [CrossRef]
- Chi, K.N.; Yu, E.Y.; Jacobs, C.; Bazov, J.; Kollmannsberger, C.; Higano, C.S.; Mukherjee, S.D.; Gleave, M.E.; Stewart, P.S.; Hotte, S.J. A phase i dose-escalation study of apatorsen (ogx-427), an antisense inhibitor targeting heat shock protein 27 (hsp27), in patients with castration-resistant prostate cancer and other advanced cancers. Ann. Oncol. 2016, 27, 1116–1122. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, J.C.; Donakonda, S.; Haupt, V.J.; Lennig, P.; Zhang, Y.; Schroeder, M. New HSP27 inhibitors efficiently suppress drug resistance development in cancer cells. Oncotarget 2016, 7, 68156–68169. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium signalling: Dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-F.; Chiu, W.-T.; Chen, Y.-T.; Lin, P.-Y.; Huang, H.-J.; Chou, C.-Y.; Chang, H.-C.; Tang, M.-J.; Shen, M.-R. Calcium store sensor stromal-interaction molecule 1-dependent signaling plays an important role in cervical cancer growth, migration, and angiogenesis. Proc. Natl. Acad. Sci. 2011, 108, 15225–15230. [Google Scholar] [CrossRef] [PubMed]
- Prevarskaya, N.; Skryma, R.; Shuba, Y. Calcium in tumour metastasis: New roles for known actors. Nat. Rev. Cancer 2011, 11, 609–618. [Google Scholar] [CrossRef]
- Prevarskaya, N.; Skryma, R.; Shuba, Y. Ion channels and the hallmarks of cancer. Trends Mol. Med. 2010, 16, 107–121. [Google Scholar] [CrossRef]
- Wong, H.S.; Chang, W.C. Correlation of clinical features and genetic profiles of stromal interaction molecule 1 (STIM1) in colorectal cancers. Oncotarget 2015, 6, 42169–42182. [Google Scholar] [CrossRef]
- Chen, Y.F.; Chen, Y.-T.; Chiu, W.-T.; Shen, M.-R. Remodeling of calcium signaling in tumor progression. J. Biomed. Sci. 2013, 20, 23. [Google Scholar] [CrossRef]
- Jardin, I.; Rosado, J.A. STIM and calcium channel complexes in cancer. Biochim. Biophys. Acta 2016, 1863, 1418–1426. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, X.; Feng, B.; Liu, N.; Wu, Q.; Han, Y.; Nie, Y.; Wu, K.; Shi, Y.; Fan, D. Stim1, a direct target of microrna-185, promotes tumor metastasis and is associated with poor prognosis in colorectal cancer. Oncogene 2014, 34, 4808–4820. [Google Scholar] [CrossRef]
- Umemura, M.; Baljinnyam, E.; Feske, S.; De Lorenzo, M.S.; Xie, L.-H.; Feng, X.; Oda, K.; Makino, A.; Fujita, T.; Yokoyama, U.; et al. Store-operated Ca2+ entry (soce) regulates melanoma proliferation and cell migration. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zhang, J.J.; Huang, X.-Y. Orai1 and stim1 are critical for breast tumor cell migration and metastasis. Cancer Cell 2009, 15, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Tang, Y.; Wang, F.; Zhang, H.; Xu, D.; Shen, Y.; Sun, S.; Yang, G. Blockade of store-operated ca2+ entry inhibits hepatocarcinoma cell migration and invasion by regulating focal adhesion turnover. Cancer Lett. 2013, 330, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Vashisht, A.; Trebak, M.; Motiani, R.K. Stim and orai proteins as novel targets for cancer therapy. A Review in the Theme: Cell and Molecular Processes in Cancer Metastasis. Am. J. Physiol. Cell Physiol. 2015, 309, C457–C469. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.S.; Huang, C.-Y.; Lee, C.-H.; Chen, W.-Y.; Huang, M.-T.; Wei, P.-L.; Chang, Y.-J. IGFBP2 plays an important role in heat shock protein 27-mediated cancer progression and metastasis. Oncotarget 2017, 8, 54978–54992. [Google Scholar] [CrossRef] [PubMed]
- Ozbey, O.; Sahin, Z.; Acar, N.; Ustunel, I. Distribution of CD105 and CD166 positive cells in the proximal epiphysis of developing rat humerus. Histol. Histopathol. 2010, 25, 1437–1445. [Google Scholar] [PubMed]
- Huang, C.-Y.; Batzorig, U.; Cheng, W.-L.; Huang, M.-T.; Chen, W.-Y.; Wei, P.-L.; Chang, Y.-J. Glucose-regulated protein 94 mediates cancer progression via akt and enos in hepatocellular carcinoma. Tumor Biol. 2015, 37, 4295–4304. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.-L.; Huang, C.-Y.; Tai, C.-J.; Batzorig, U.; Cheng, W.-L.; Hunag, M.-T.; Chang, Y.-J. Glucose-regulated protein 94 mediates metastasis by cct8 and the jnk pathway in hepatocellular carcinoma. Tumor Biol. 2016, 37, 8219–8227. [Google Scholar] [CrossRef]
- Chang, Y.-J.; Li, L.-T.; Chen, H.-A.; Hung, C.-S.; Wei, P.-L. Silencing survivin activates autophagy as an alternative survival pathway in hcc cells. Tumor Biol. 2014, 35, 9957–9966. [Google Scholar] [CrossRef]
- Chang, Y.J.; Cheng, Y.W.; Lin, R.K.; Huang, C.C.; Chen, W.K.; Ke, T.W.; Wei, P.L. Thrombomodulin influences the survival of patients with non-metastatic colorectal cancer through epithelial-to-mesenchymal transition (EMT). PLoS ONE 2016, 11, e0160550. [Google Scholar] [CrossRef]
- Kuo, L.-J.; Hung, C.-S.; Chen, W.-Y.; Chang, Y.-J.; Wei, P.-L. Glucose-regulated protein 78 silencing down-regulates vascular endothelial growth factor/vascular endothelial growth factor receptor 2 pathway to suppress human colon cancer tumor growth. J. Surg. Res. 2013, 185, 264–272. [Google Scholar] [CrossRef]
- Rosen, L.B.; Ginty, D.D.; Greenberg, M.E. Calcium regulation of gene expression. Adv. Second Messenger Phosphoprot. Res. 1995, 30, 225–253. [Google Scholar]
- Thastrup, O.; Cullen, P.J.; Drobak, B.K.; Hanley, M.R.; Dawson, A.P. Thapsigargin, a tumor promoter, discharges intracellular Ca2+ stores by specific inhibition of the endoplasmic reticulum Ca2(+)-atpase. Proc. Natl. Acad. Sci. 1990, 87, 2466–2470. [Google Scholar] [CrossRef]
- Cheng, J.; Lv, Z.; Weng, X.; Ye, S.; Shen, K.; Li, M.; Qin, Y.; Hu, C.; Zhang, C.; Wu, J.; et al. Hsp27 acts as a master molecular chaperone and plays an essential role in hepatocellular carcinoma progression. Digestion 2015, 92, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tao, X.; Jin, G.; Jin, H.; Wang, N.; Hu, F.; Luo, Q.; Shu, H.; Zhao, F.; Yao, M.; et al. A targetable molecular chaperone hsp27 confers aggressiveness in hepatocellular carcinoma. Theranostics 2016, 6, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Andrieu, C.; Taieb, D.; Baylot, V.; Ettinger, S.; Soubeyran, P.; De-Thonel, A.; Nelson, C.; Garrido, C.; So, A.; Fazli, L.; et al. Heat shock protein 27 confers resistance to androgen ablation and chemotherapy in prostate cancer cells through eif4e. Oncogene 2010, 29, 1883–1896. [Google Scholar] [CrossRef] [PubMed]
- Srivats, S.; Balasuriya, D.; Pasche, M.; Vistal, G.; Edwardson, J.M.; Taylor, C.W.; Murrell-Lagnado, R.D. Sigma1 receptors inhibit store-operated ca2+entry by attenuating coupling of stim1 to orai1. J. Cell Biol. 2016, 213, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Park, C.Y.; Hoover, P.J.; Mullins, F.M.; Bachhawat, P.; Covington, E.D.; Raunser, S.; Walz, T.; Garcia, K.C.; Dolmetsch, R.E.; Lewis, R.S. Stim1 clusters and activates crac channels via direct binding of a cytosolic domain to orai1. Cell 2009, 136, 876–890. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Meraner, P.; Kwon, H.T.; Machnes, D.; Oh-hora, M.; Zimmer, J.; Huang, Y.; Stura, A.; Rao, A.; Hogan, P.G. Stim1 gates the store-operated calcium channel orai1 in vitro. Nat. Struct. Mol. Biol. 2010, 17, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, M.; Xu, L.; Lin, D.; Cai, S.; Zou, F. The apoptosis of non-small cell lung cancer induced by cisplatin through modulation of stim1. Exp. Toxicol. Pathol. 2013, 65, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.D.; Xia, X.; Lv, X.F.; Yu, B.X.; Yuan, J.N.; Mai, X.Y.; Shang, J.Y.; Zhou, J.G.; Liang, S.J.; Pang, R.P. Inhibition of Orai1-mediated Ca2+ entry enhances chemosensitivity of HepG2 hepatocarcinoma cells to 5-fluorouracil. J. Cell Mol. Med. 2017, 21, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wei, Q.; Cheng, J.; Bian, Y.; Tian, C.; Hu, Y.; Li, H. Enhanced Stim1 expression is associated with acquired chemo-resistance of cisplatin in osteosarcoma cells. Hum. Cell 2017, 30, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Bauer, K.; Nitsche, U.; Slotta-Huspenina, J.; Drecoll, E.; von Weyhern, C.H.; Rosenberg, R.; Höfler, H.; Langer, R. High hsp27 and hsp70 expression levels are independent adverse prognostic factors in primary resected colon cancer. Cell Oncol. 2012, 35, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Sherr, C.J.; Roberts, J.M. CDK inhibitors: Positive and negative regulators of G1-phase progression. Genes Dev. 1999, 13, 1501–1512. [Google Scholar] [CrossRef]
- Zhou, B.B.; Elledge, S.J. The DNA damage response: Putting checkpoints in perspective. Nature 2000, 408, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, G.K.; Shah, M.A. Targeting the cell cycle: A new approach to cancer therapy. J. Clin. Oncol. 2005, 23, 9408–9421. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Tow Way Direction | Sequence | TM °C |
|---|---|---|---|
| STIM1 | forward reversed | 5′-CTGGGATCTCAGAGGGATTTGA-3′ 5′-GCTGGCGGTCACTCATGTG-3′ | 60 |
| Orai1 | forward reversed | 5′-GCTCATGATCAGCACCTGCAT-3′ 5′-GGGACTCCTTGACCGAGTTG-3′ | 60 |
| GAPDH | forward reversed | 5′-ATACTCCTGCTTGCTGATCC-3′ 5′-CCTGTACGCCAACACAGTGC-3′ | 60 |
| Characteristics | HSP27 Expression * | p Value | |
|---|---|---|---|
| Low (n = 83) | High (n = 94) | ||
| Gender | 0.891 | ||
| Male | 45 (54%) | 50 (53%) | |
| Female | 38 (46%) | 44 (47%) | |
| Mean age (years) ± SD | 66.6 ± 11.4 | 66.7 ± 10.1 | 0.974 |
| Age | 0.831 | ||
| <65 years | 34 (62%) | 40 (51%) | |
| ≧65 years | 49 (38%) | 54 (49%) | |
| Grading of carcinoma | 0.458 | ||
| Well differentiated | 7 (8%) | 7 (8%) | |
| Moderately differentiated | 53 (64%) | 68 (72%) | |
| Poorly differentiated | 23 (28%) | 19 (20%) | |
| Invasive depth of tumor ** | 0.009 | ||
| T1 + T2 | 8 (10%) | 1 (1%) | |
| T3 + T4 | 74 (90%) | 93 (99%) | |
| Lymph node metastasis | 0.848 | ||
| Negative | 55 (66%) | 61 (65%) | |
| Positive | 28 (34%) | 33 (35%) | |
| Distant metastasis | 0.877 | ||
| Negative | 80 (96%) | 91 (97%) | |
| Positive | 3 (4%) | 3 (3%) | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.-Y.; Wei, P.-L.; Chen, W.-Y.; Chang, W.-C.; Chang, Y.-J. Silencing Heat Shock Protein 27 Inhibits the Progression and Metastasis of Colorectal Cancer (CRC) by Maintaining the Stability of Stromal Interaction Molecule 1 (STIM1) Proteins. Cells 2018, 7, 262. https://doi.org/10.3390/cells7120262
Huang C-Y, Wei P-L, Chen W-Y, Chang W-C, Chang Y-J. Silencing Heat Shock Protein 27 Inhibits the Progression and Metastasis of Colorectal Cancer (CRC) by Maintaining the Stability of Stromal Interaction Molecule 1 (STIM1) Proteins. Cells. 2018; 7(12):262. https://doi.org/10.3390/cells7120262
Chicago/Turabian StyleHuang, Chien-Yu, Po-Li Wei, Wei-Yu Chen, Wei-Chiao Chang, and Yu-Jia Chang. 2018. "Silencing Heat Shock Protein 27 Inhibits the Progression and Metastasis of Colorectal Cancer (CRC) by Maintaining the Stability of Stromal Interaction Molecule 1 (STIM1) Proteins" Cells 7, no. 12: 262. https://doi.org/10.3390/cells7120262
APA StyleHuang, C.-Y., Wei, P.-L., Chen, W.-Y., Chang, W.-C., & Chang, Y.-J. (2018). Silencing Heat Shock Protein 27 Inhibits the Progression and Metastasis of Colorectal Cancer (CRC) by Maintaining the Stability of Stromal Interaction Molecule 1 (STIM1) Proteins. Cells, 7(12), 262. https://doi.org/10.3390/cells7120262