Effect of shRNA Mediated Silencing of YB-1 Protein on the Expression of Matrix Collagenases in Malignant Melanoma Cell In Vitro



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
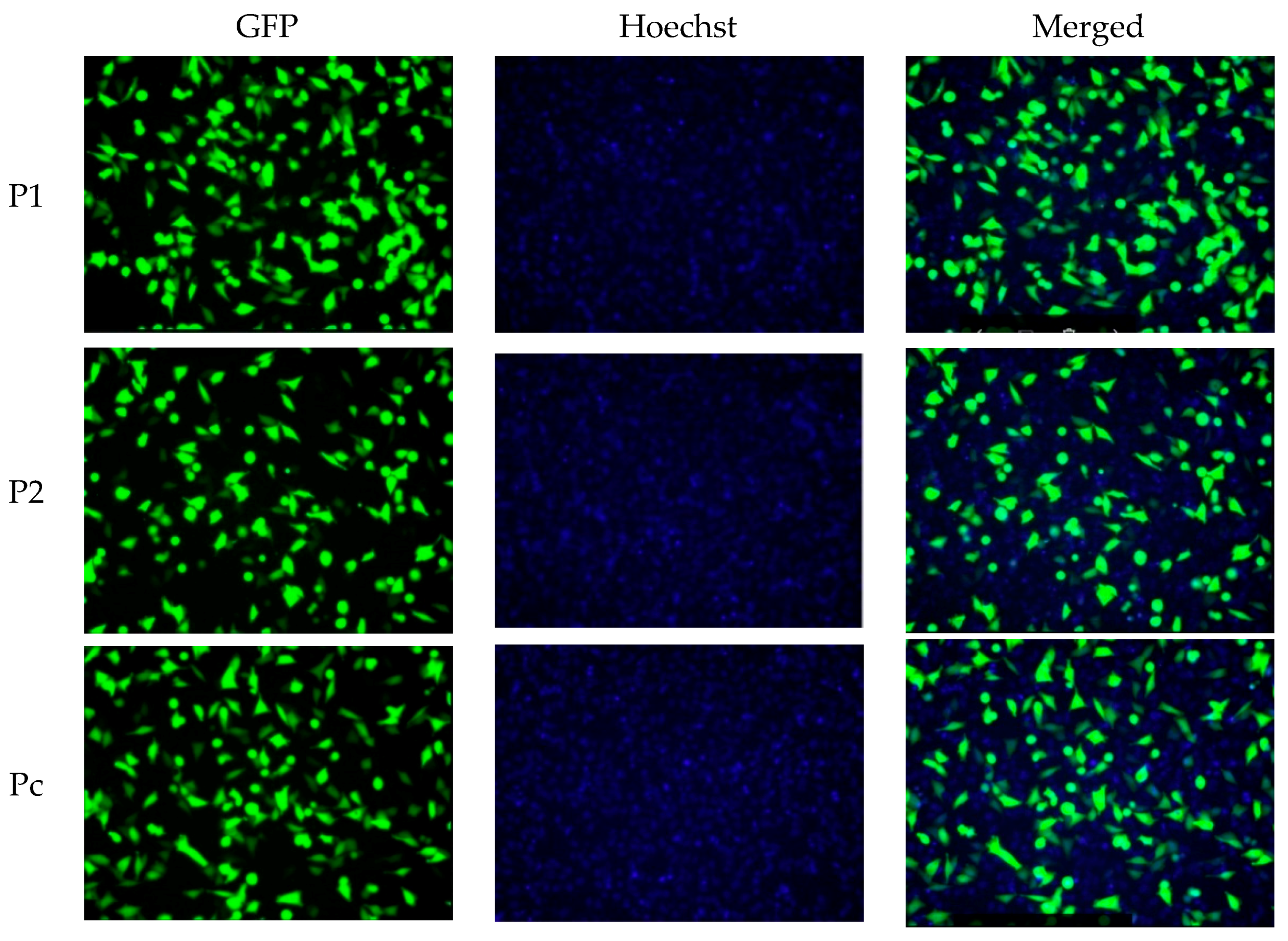
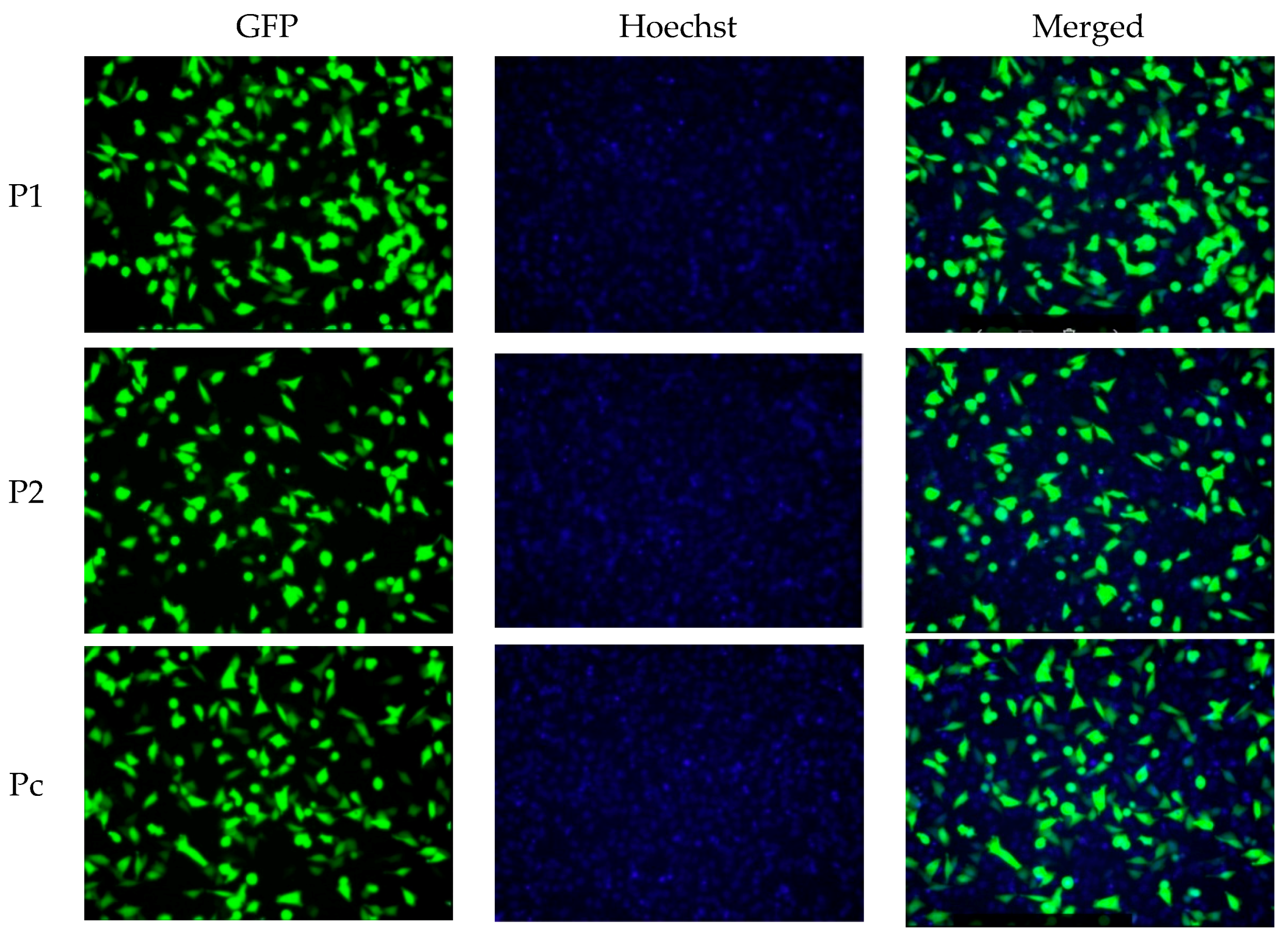
2.1. YB-1 Silencing via shRNA Mediated RNA Interference in A375 Malignant Melanoma Cell Line
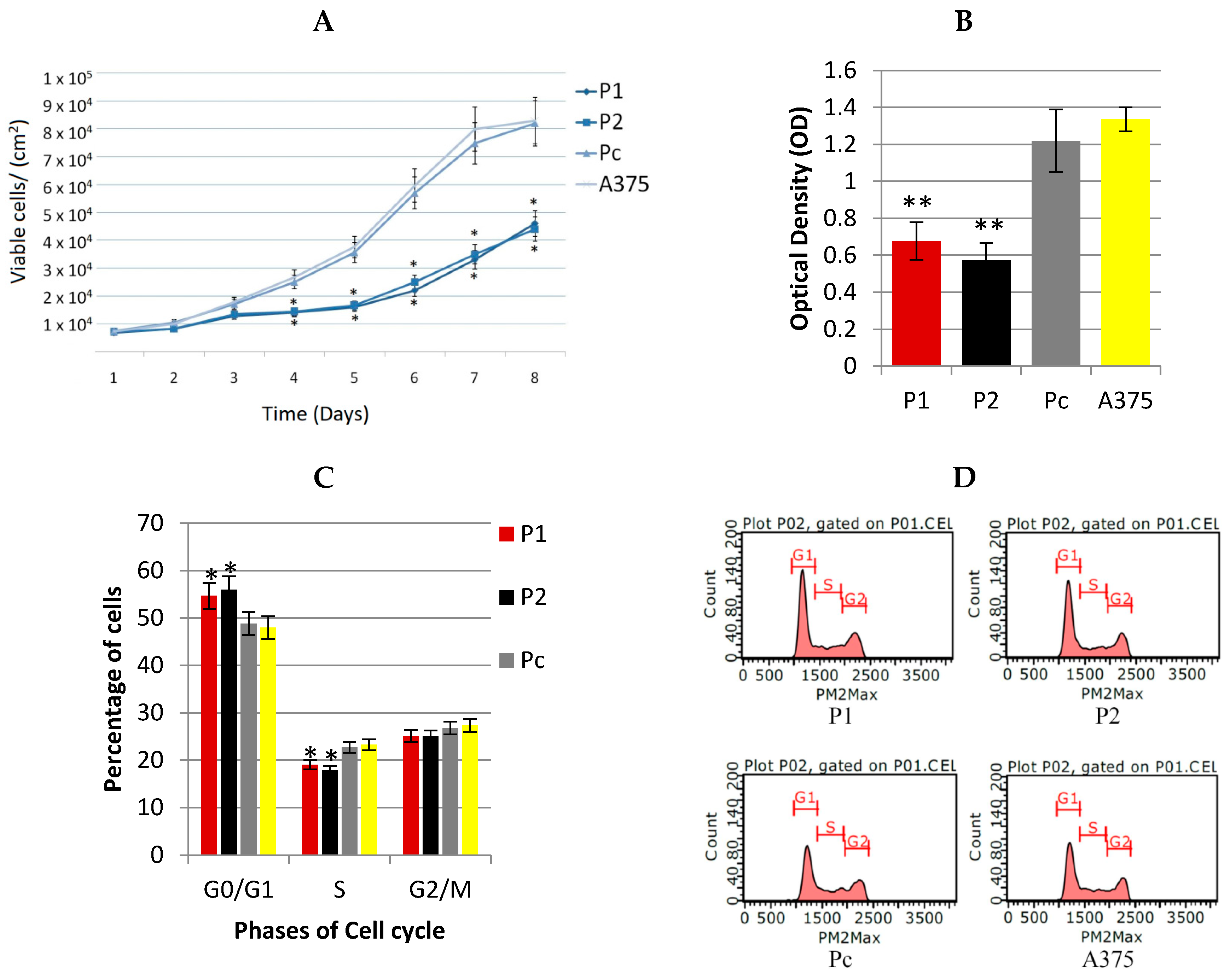
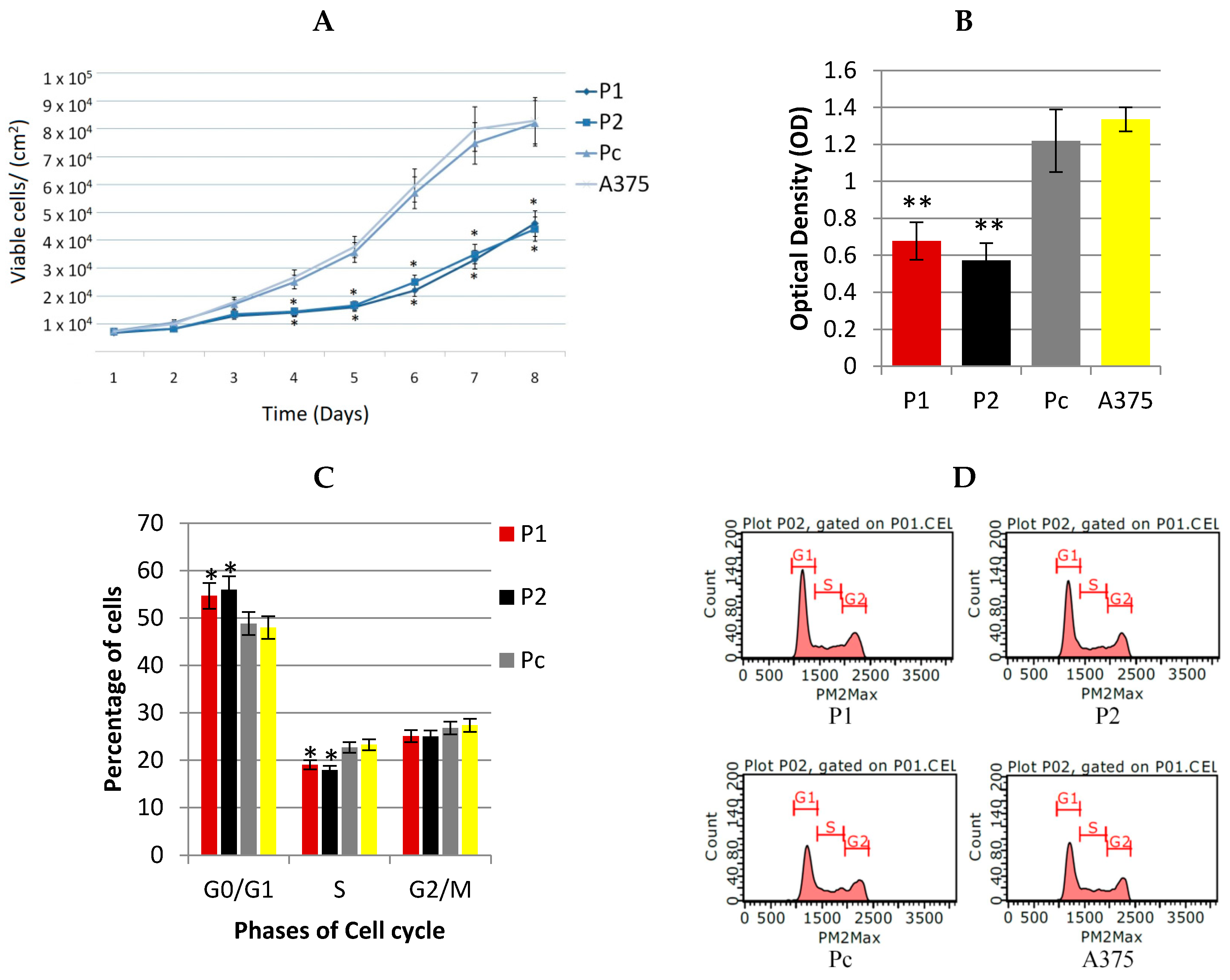
2.2. Antiproliferative Effect of YB-1 Silencing in A375 Cell Line
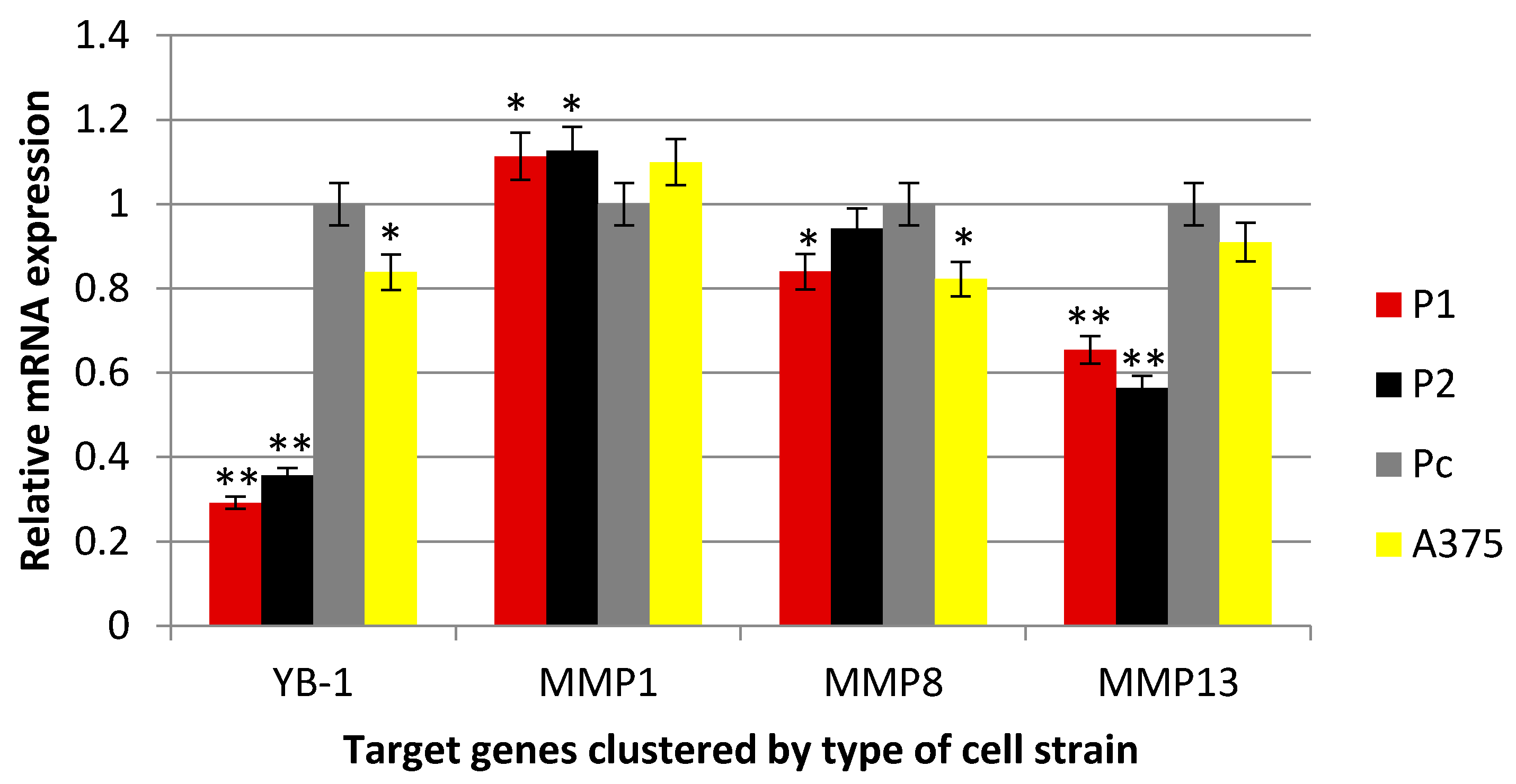
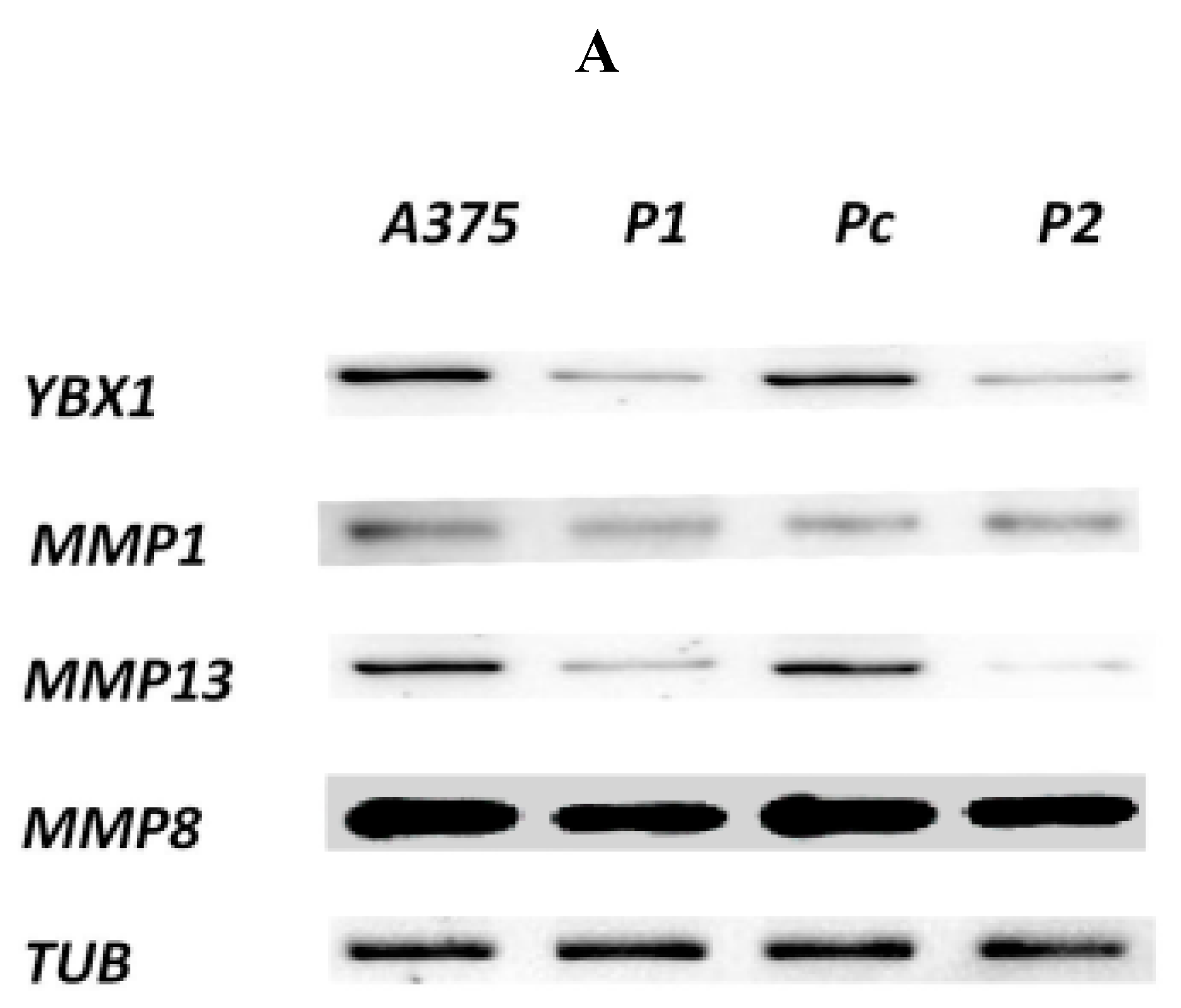
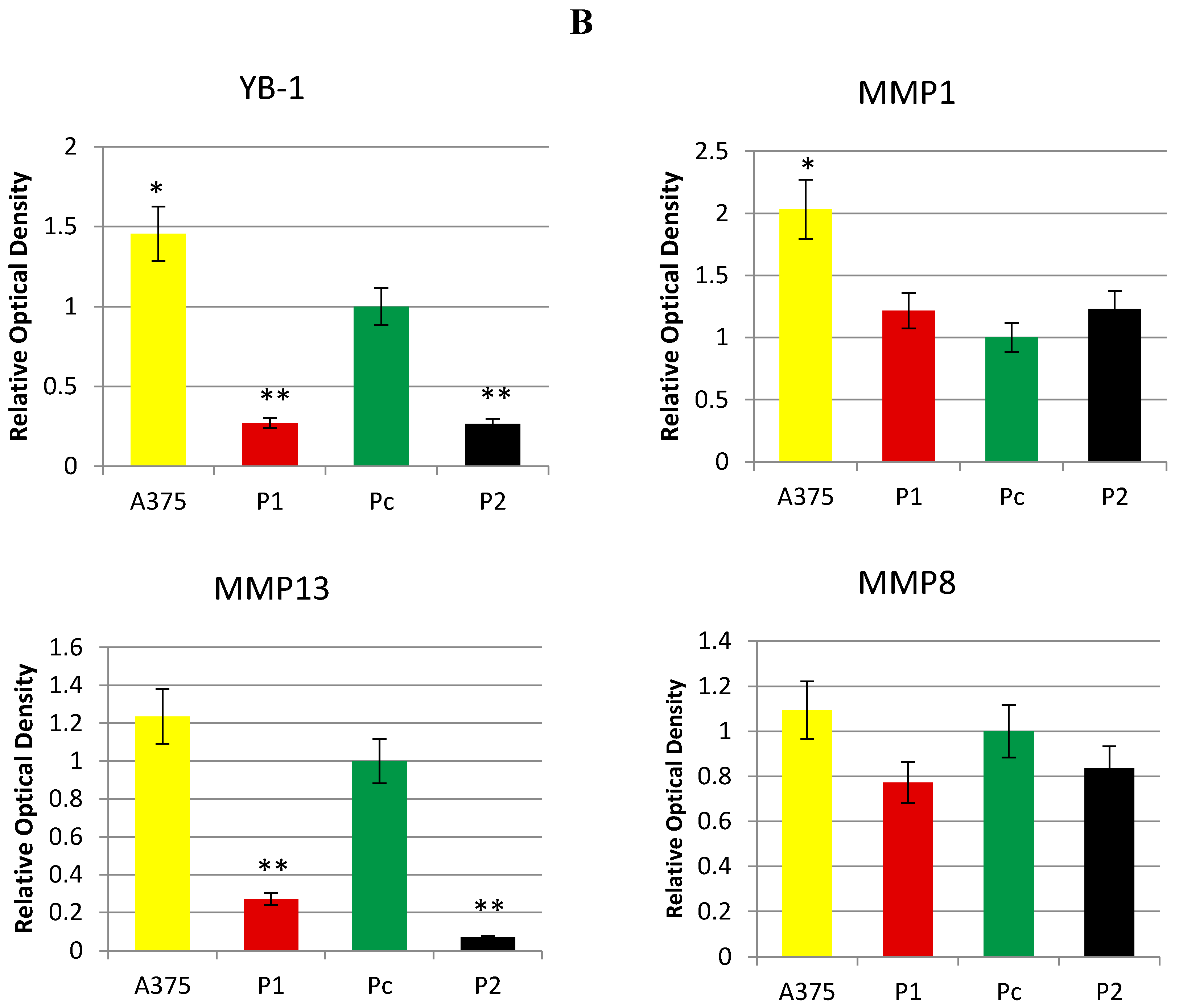
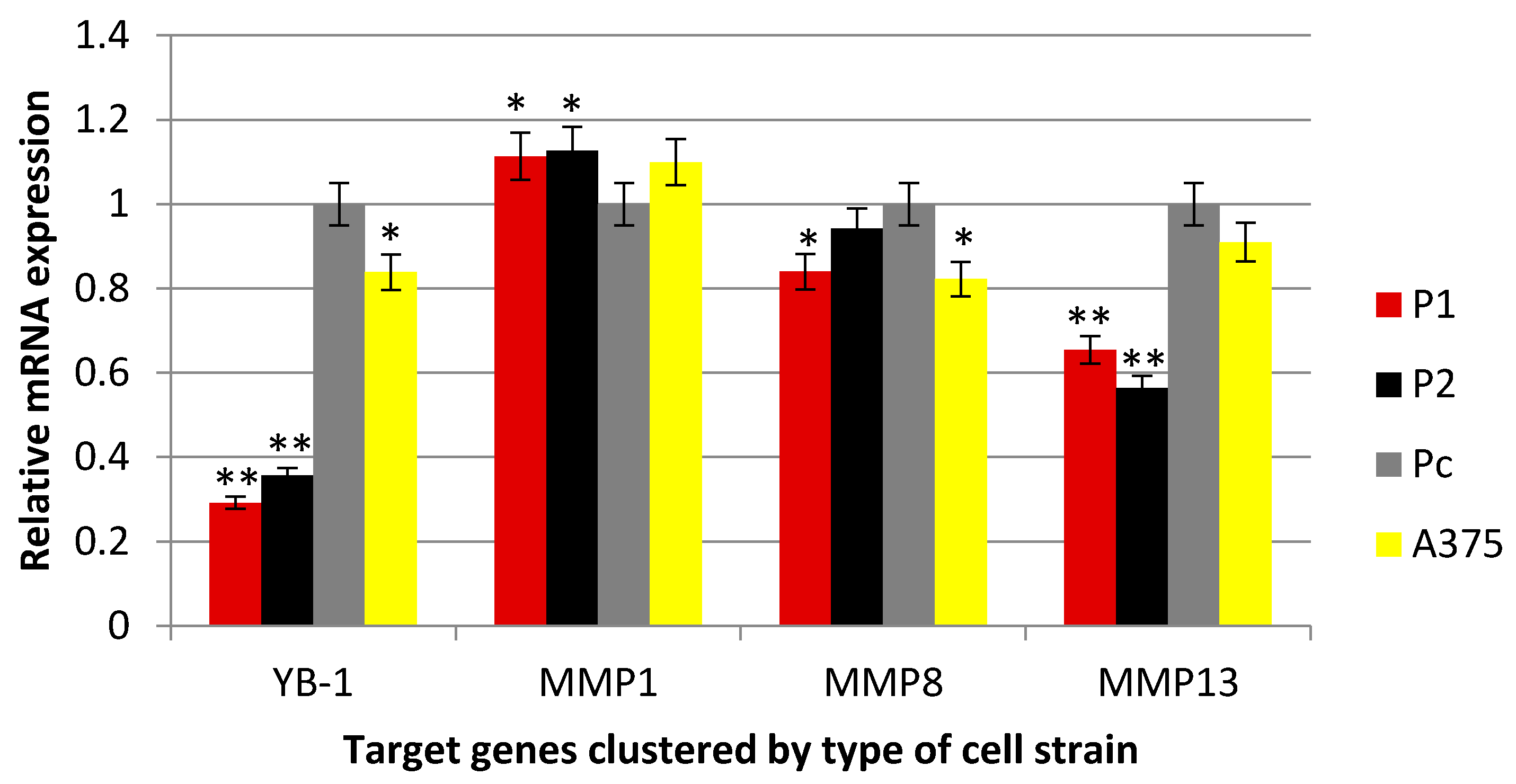
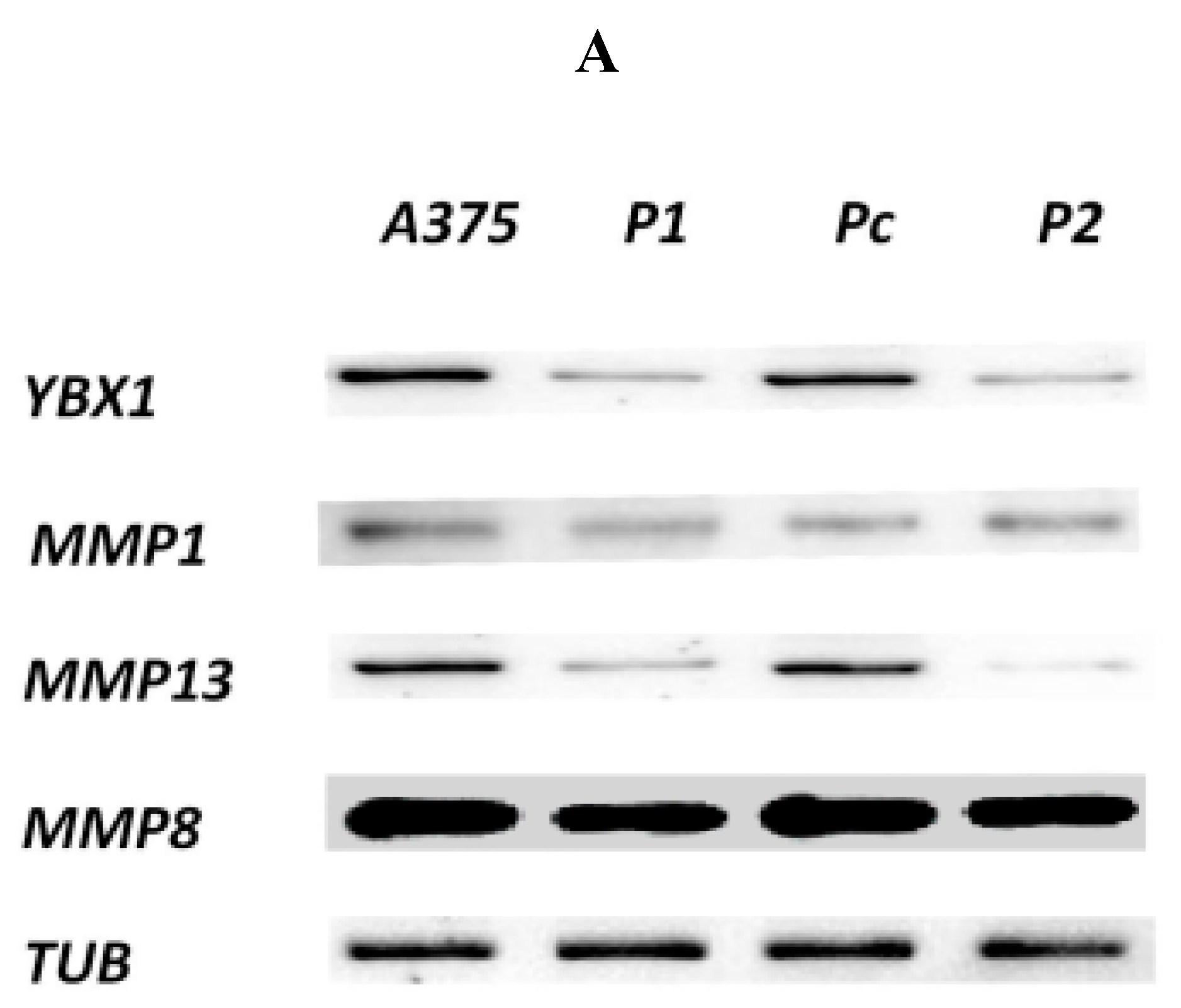
2.3. Effect of YB-1 Silencing on Expression of Matrix Collagenases in A375 Cell Line
2.4. Effect of YB-1 Silencing on A375 Cell Line Migration Assay In Vitro
3. Discussion
3.1. YB-1 Protein and A375 Cancer Cell Proliferation
3.2. YB-1 Protein and the Expression of Collagenases MMPs in A375 Cancer Cells
3.3. YB-1 Protein and A375 Cancer Cell Migration
4. Materials and Methods
4.1. Establishing the Stable YB-1 Silenced Malignant Melanoma Cell Line
4.2. Real-Time PCR
4.3. Protein Extraction and Western Blot Analysis
4.4. MTT Assay and Cell Counting
4.5. Immuno-Fluorescence Staining
4.6. Hoechst Nuclear Staining
4.7. Flow Cytometry for Cell Cycle Interference
4.8. Wound Healing Assay
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kumar, V.; Abbas, A.K.; Aster, J.C. Robbins Basic Pathology; Elsevier Health Sciences: Philadelphia, PA, USA, 2012. [Google Scholar]
- Lens, M.; Dawes, M. Global perspectives of contemporary epidemiological trends of cutaneous malignant melanoma. Br. J. Dermatol. 2004, 150, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Gialeli, C.; Theocharis, A.D.; Karamanos, N.K. Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting. FEBS J. 2011, 278, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Rohani, M.G.; Parks, W.C. Matrix remodeling by MMPs during wound repair. Matrix Biol. 2015, 44, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Srivastava, S.K.; Chaudhuri, T.K.; Upadhyay, G. Multifaceted role of matrix metalloproteinases (MMPs). Front. Mol. Biosci. 2015, 2, 19. [Google Scholar] [CrossRef] [PubMed]
- Fanjul-Fernández, M.; Folgueras, A.R.; Cabrera, S.; López-Otín, C. Matrix metalloproteinases: Evolution, gene regulation and functional analysis in mouse models. Biochim. Biophys. Acta 2010, 1803, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Gaffney, J.; Solomonov, I.; Zehorai, E.; Sagi, I. Multilevel regulation of matrix metalloproteinases in tissue homeostasis indicates their molecular specificity in vivo. Matrix Biol. 2015, 44, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Curry, T.E., Jr.; Osteen, K.G. The matrix metalloproteinase system: Changes, regulation, and impact throughout the ovarian and uterine reproductive cycle. Endocr. Rev. 2003, 24, 428–465. [Google Scholar] [CrossRef] [PubMed]
- Noël, A.; Gutiérrez-Fernández, A.; Sounni, N.E.; Behrendt, N.; Maquoi, E.; Lund, I.K.; Cal, S.; Houer-Hansen, G.; López-Otín, C. New and paradoxical roles of matrix metalloproteinases in the tumor microenvironment. Front. Pharmacol. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Moro, N.; Mauch, C.; Zigrino, P. Metalloproteinases in melanoma. Eur. J. Cell Biol. 2014, 93, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Martinet, Y.; Hirsch, F.R.; Mulshine, J.; Vignaud, J.-M. Clinical and Biological Basis of Lung Cancer Prevention; Birkhäuser: Basel, Switzerland, 2012. [Google Scholar]
- Kim, K.S.; Choi, H.M.; Lee, Y.-A.; Choi, I.A.; Lee, S.-H.; Hong, S.-J.; Yang, H.-I.; Yoo, M.C. Expression levels and association of gelatinases MMP-2 and MMP-9 and collagenases MMP-1 and MMP-13 with VEGF in synovial fluid of patients with arthritis. Rheumatol. Int. 2011, 31, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Schröpfer, A.; Kammerer, U.; Kapp, M.; Dietl, J.; Feix, S.; Anacker, J. Expression pattern of matrix metalloproteinases in human gynecological cancer cell lines. BMC Cancer 2010, 10, 553. [Google Scholar] [CrossRef] [PubMed]
- Thirkettle, S.; Decock, J.; Arnold, H.; Pennington, C.J.; Jaworski, D.M.; Edwards, D.R. Matrix metalloproteinase 8 (collagenase 2) induces the expression of interleukins 6 and 8 in breast cancer cells. J. Biol. Chem. 2013, 288, 16282–16294. [Google Scholar] [CrossRef] [PubMed]
- Lasham, A.; Woolley, A.G.; Dunn, S.E.; Braithwaite, A.W. YB-1: Oncoprotein, prognostic marker and therapeutic target? Biochem. J. 2013, 449, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Eliseeva, I.; Kim, E.; Guryanov, S.; Ovchinnikov, L.; Lyabin, D. Y-box-binding protein 1 (YB-1) and its functions. Biochemistry 2011, 76, 1402–1433. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, R.; Lee, J.; Mahimkar, R.; Lovett, D. P53, YB-1 and the AP-1 family of transcription factors mediate MMP-2 transcription induced by skeletal muscle ischemia. J. Am. Coll. Surg. 2004, 199, 103. [Google Scholar] [CrossRef]
- Schittek, B.; Psenner, K.; Sauer, B.; Meier, F.; Iftner, T.; Garbe, C. The increased expression of Y box-binding protein 1 in melanoma stimulates proliferation and tumor invasion, antagonizes apoptosis and enhances chemoresistance. Int. J. Cancer 2007, 120, 2110–2118. [Google Scholar] [CrossRef] [PubMed]
- Adley, B.P.; Gleason, K.J.; Yang, X.J.; Stack, M.S. Expression of membrane type 1 matrix metalloproteinase (MMP-14) in epithelial ovarian cancer: High level expression in clear cell carcinoma. Gynecol. Oncol. 2009, 112, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Samuel, S.; Beifuss, K.K.; Bernstein, L.R. YB-1 binds to the MMP-13 promoter sequence and represses MMP-13 transactivation via the AP-1 site. Biochim. Biophys. Acta 2007, 1769, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Wisam, N.I.; Ridhwan, A.W.; Syaiful, A.R. Expression of Collagenases Matrix Metalloproteinases and YB-1 Oncogenic Factor in Malignant Melanoma Cancer Cells and its Regulation by Stromal Fibroblasts. Int. J. Cancer Res. 2017, 13, 17–25. [Google Scholar] [CrossRef]
- Wang, H.; Sun, R.; Gu, M.; Li, S.; Zhang, B.; Chi, Z.; Hao, L. shRNA-Mediated Silencing of Y-Box Binding Protein-1 (YB-1) Suppresses Growth of Neuroblastoma Cell SH-SY5Y In Vitro and In Vivo. PLoS ONE 2015, 10, e0127224. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Zhou, L.; Chen, Q.; Chen, C.; Fang, L.; Fang, X.; Shen, H.L. Effect of YB-1 gene knockdown on human leukemia cell line K562/A02. Chin. J. Med. Genet. 2009, 26, 400–405. [Google Scholar]
- Gao, Y.; Fotovati, A.; Lee, C.; Wang, M.; Cote, G.; Guns, E.; Toyota, B.; Faury, D.; Jabado, N.; Dunn, S.E. Inhibition of Y-box binding protein-1 slows the growth of glioblastoma multiforme and sensitizes to temozolomide independent O6-methylguanine-DNA methyltransferase. Mol. Cancer Ther. 2009, 8, 3276–3284. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wei, J.; Zhang, S.; Zhang, X. Shrimp miR-S8 suppresses the stemness of human melanoma stem-like cells by targeting the transcription factor YB-1. Cancer Res. 2017, 77, 5543–5553. [Google Scholar] [CrossRef] [PubMed]
- Shamsuddin, S. Immunoprecipitation of 11 ZN fingers Domains, CTCF and BORIS to Multifunctional Y-box DNA/RNA-binding factor, YB-1 in Glioma-RGBM. J. Biomed. Pharm. Res. 2016, 5, 89–95. [Google Scholar]
- Meierjohann, S.; Hufnagel, A.; Wende, E.; Kleinschmidt, M.A.; Wolf, K.; Friedl, P.; Gaubatz, S.; Schartl, M. MMP13 mediates cell cycle progression in melanocytes and melanoma cells: In vitro studies of migration and proliferation. Mol. Cancer 2010, 9, 201. [Google Scholar] [CrossRef] [PubMed]
- Evdokimova, V.; Tognon, C.; Ng, T.; Ruzanov, P.; Melnyk, N.; Fink, D.; Sorokin, A.; Ovchinnikov, L.P.; Davicioni, E.; Triche, T.J.; et al. Translational activation of snail1 and other developmentally regulated transcription factors by YB-1 promotes an epithelial-mesenchymal transition. Cancer Cell 2009, 15, 402–415. [Google Scholar] [CrossRef] [PubMed]
- Mouneimne, G.; Brugge, J.S. YB-1 translational control of epithelial-mesenchyme transition. Cancer Cell 2009, 15, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Justus, C.R.; Leffler, N.; Ruiz-Echevarria, M.; Yang, L.V. In vitro cell migration and invasion assays. J. Vis. Exp. JoVE 2014. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, W.N.; Doolaanea, A.A.; Bin Abdull Rasad, M.S.B. Effect of shRNA Mediated Silencing of YB-1 Protein on the Expression of Matrix Collagenases in Malignant Melanoma Cell In Vitro. Cells 2018, 7, 7. https://doi.org/10.3390/cells7010007
Ibrahim WN, Doolaanea AA, Bin Abdull Rasad MSB. Effect of shRNA Mediated Silencing of YB-1 Protein on the Expression of Matrix Collagenases in Malignant Melanoma Cell In Vitro. Cells. 2018; 7(1):7. https://doi.org/10.3390/cells7010007
Chicago/Turabian StyleIbrahim, Wisam Nabeel, Abd Almonem Doolaanea, and Mohammad Syaiful Bahari Bin Abdull Rasad. 2018. "Effect of shRNA Mediated Silencing of YB-1 Protein on the Expression of Matrix Collagenases in Malignant Melanoma Cell In Vitro" Cells 7, no. 1: 7. https://doi.org/10.3390/cells7010007
APA StyleIbrahim, W. N., Doolaanea, A. A., & Bin Abdull Rasad, M. S. B. (2018). Effect of shRNA Mediated Silencing of YB-1 Protein on the Expression of Matrix Collagenases in Malignant Melanoma Cell In Vitro. Cells, 7(1), 7. https://doi.org/10.3390/cells7010007