
EFR3A, an Intriguing Gene, and Protein with a Scaffolding Function


 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Physiological Roles of EFR3A
2.1. Drosophila Melanogaster Phototransduction
2.2. EFR3 Proteins Affect GPCR Responsiveness by Regulating Receptor Phosphorylation
2.3. EFR3A and Insulin-Mediated Dispersal of GLUT4
2.4. Brain-Specific EfrA Knockout Promotes Hippocampal Neurogenesis in Mice
3. Association of EFR3A Protein with Disease States
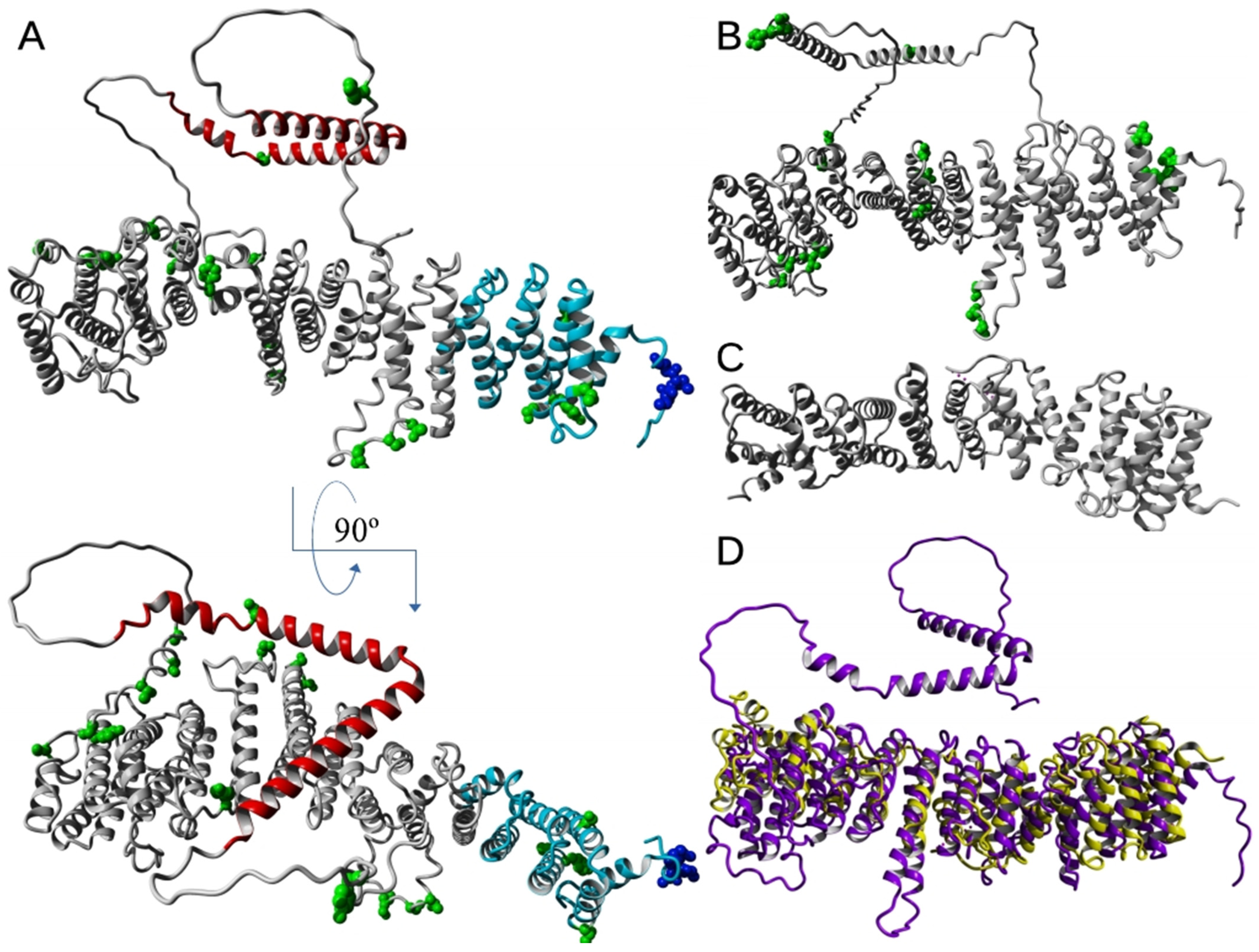
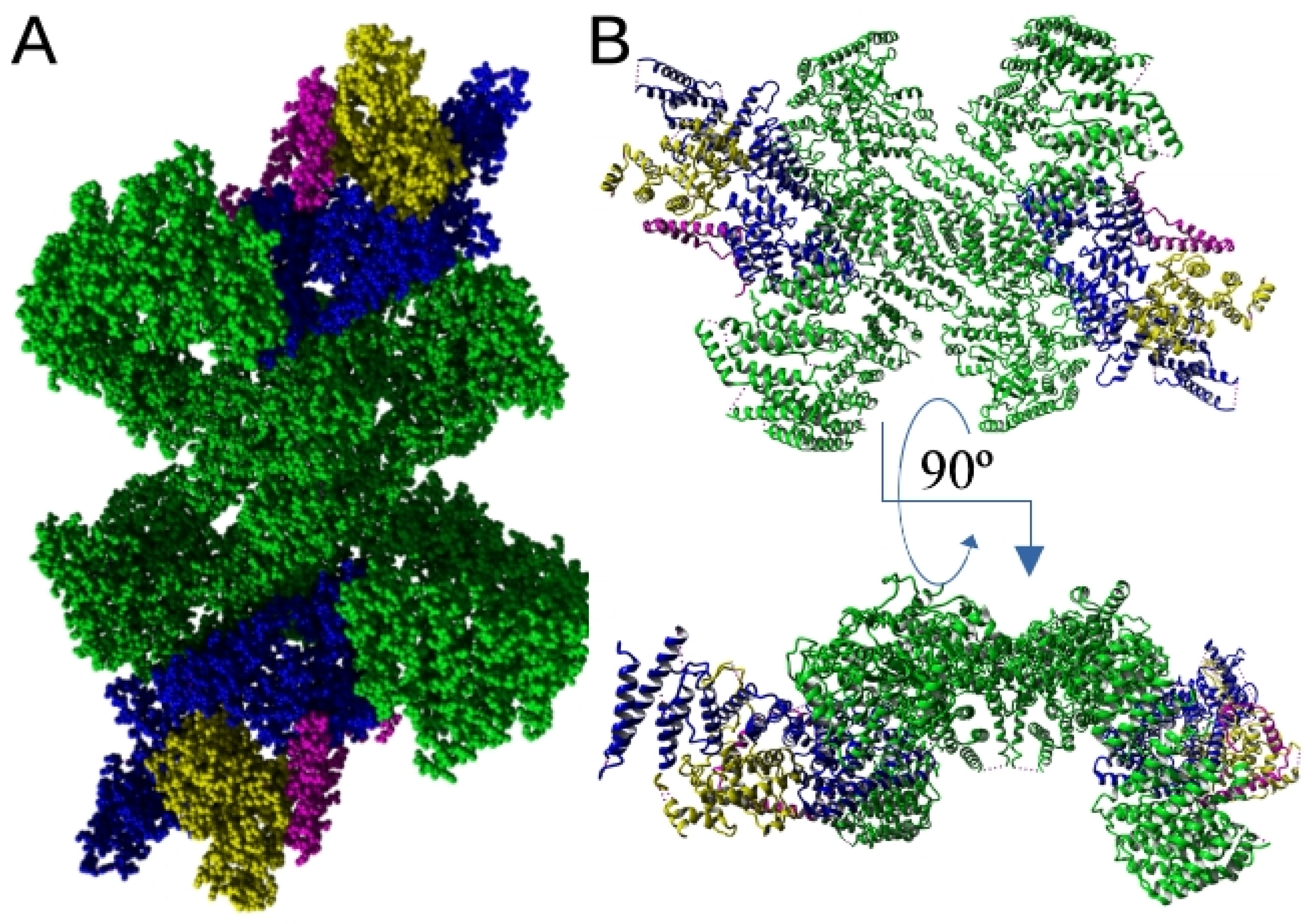
4. EFR3 Structural Features: Atomic Structure and Domain Organization
5. Post-Translational Modifications
5.1. S-Palmitoylation
5.2. Phosphorylation
5.3. Other Post-Translational Modifications
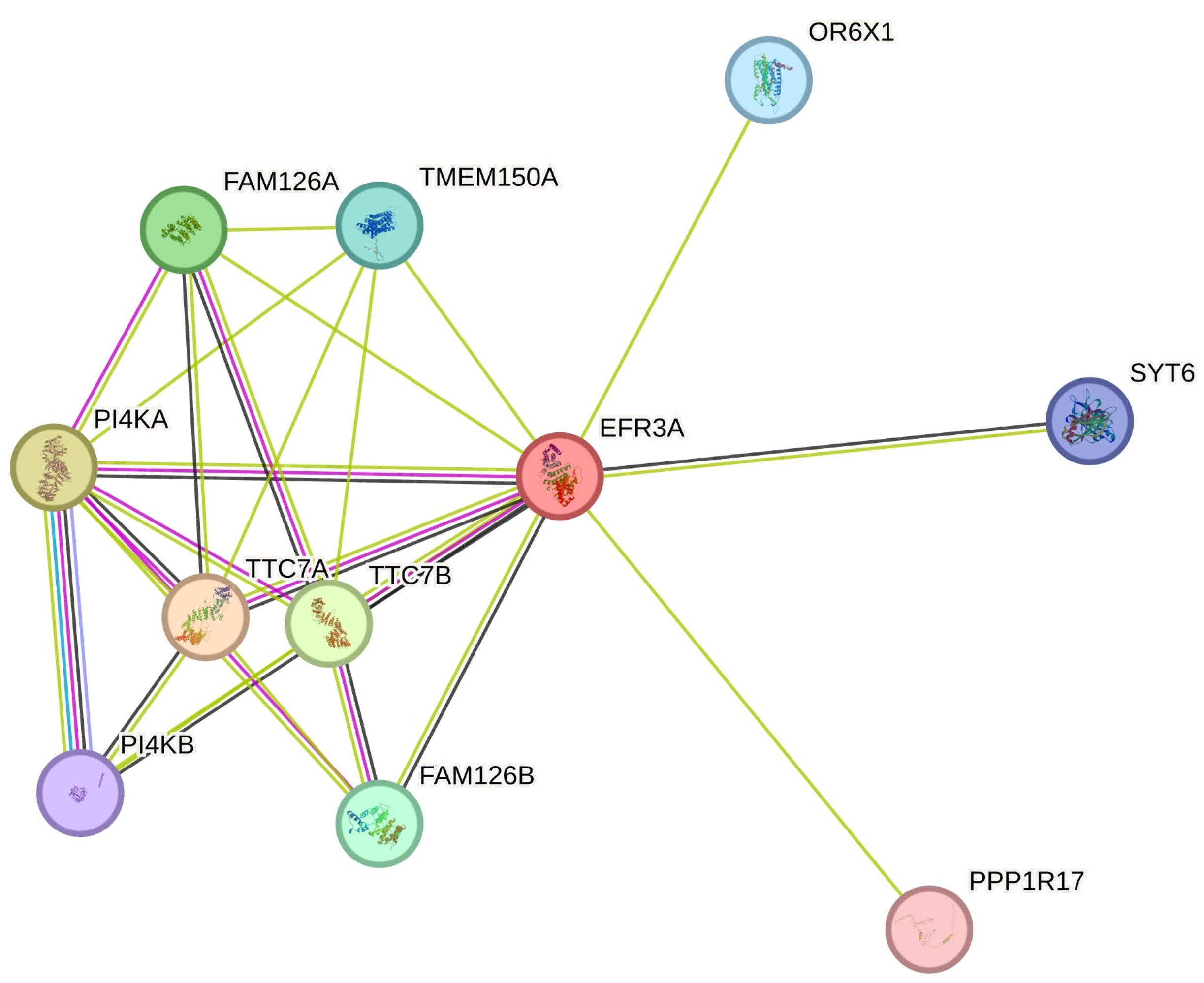
6. Functional Complexes
6.1. PI4K Anchoring in the Plasma Membrane
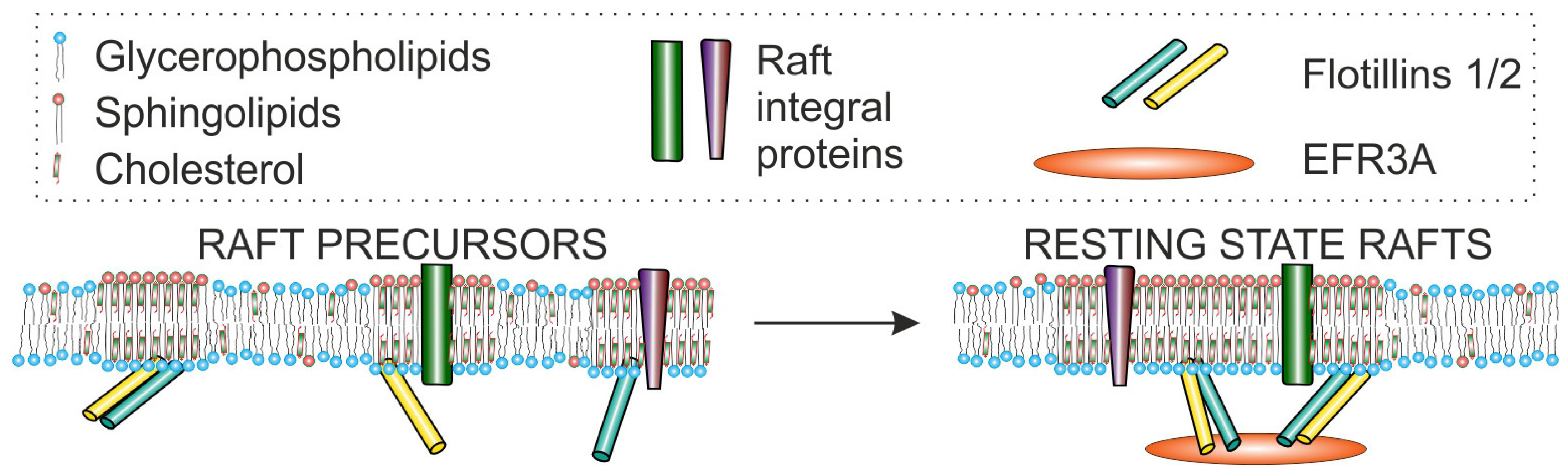
6.2. EFRA Possibly Plays a Role as Membrane Raft Organizer—Interaction with Flotillin-2
7. Concluding Remarks and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACS | acute coronary syndromes |
| ARM | armadillo |
| ASD | autism spectrum disorders |
| AT1R | angiotensin II receptor type 1 |
| BDNF | brain-derived neurotrophic factor |
| CAD | coronary artery disease |
| CID-MIA | combined immunodeficiency with multiple intestinal atresias |
| CRC | colorectal cancer |
| DAG | diacylglycerol |
| DRM | detergent-resistant membrane |
| EFR3A | eighty-five requiring 3A |
| GPMVs | giant PM vesicles |
| IRV | insulin-responding vesicles |
| NPC | nasopharyngeal carcinoma |
| PDAC | pancreatic ductal adenocarcinoma |
| PI(3,4,5)P3 | phosphatidyl inositol (3,4,5)-trisphosphate |
| PI(4,5P)2 | phosphatidyl inositol (4,5)-bisphosphate |
| PI4KA | phosphatidylinositol 4-kinase alpha |
| PLCγ1 | phospholipase Cγ1 |
| PM | plasma membrane |
| SAD | single-wavelength anomalous dispersion |
| SNP | single nucleotide polymorphism |
| TRP | transient receptor potential |
| TTC7 | tetratricopeptide repeat domain 7 |
References
- Lenburg, M.E.; O’Shea, E.K. Genetic evidence for a morphogenetic function of the Saccharomyces cerevisiae Pho85 cyclin-dependent kinase. Genetics 2001, 157, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, D.L.; Dockendorff, T.C.; Jongens, T.A. Clonal analysis of cmp44E, which encodes a conserved putative transmembrane protein, indicates a requirement for cell viability in Drosophila. Dev. Genet. 1998, 23, 264–274. [Google Scholar] [CrossRef]
- Huang, F.D.; Woodruff, E.; Mohrmann, R.; Broadie, K. Rolling blackout is required for synaptic vesicle exocytosis. J. Neurosci. 2006, 26, 2369–2379. [Google Scholar] [CrossRef]
- Available online: https://www.ncbi.nlm.nih.gov/protein/ (accessed on 15 January 2025).
- Available online: https://www.ebi.ac.uk/interpro/protein/UniProt/Q14156/entry/,pfam/#table (accessed on 20 December 2024).
- Hoellerbauer, P.; Biery, M.C.; Arora, S.; Rao, Y.; Girard, E.J.; Mitchell, K.; Dighe, P.; Kufeld, M.; Kuppers, D.A.; Herman, J.A.; et al. Functional genomic analysis of adult and pediatric brain tumor isolates. bioRxiv 2023. [Google Scholar] [CrossRef]
- The EMBL-EBI Job Dispatcher Sequence Analysis Tools Framework in 2024. Available online: https://www.ebi.ac.uk/jdispatcher/msa/clustalo (accessed on 4 November 2024).
- Noack, L.C.; Bayle, V.; Armengot, L.; Rozier, F.; Mamode-Cassim, A.; Stevens, F.D.; Caillaud, M.C.; Munnik, T.; Mongrand, S.; Pleskot, R.; et al. A nanodomain-anchored scaffolding complex is required for the function and localization of phosphatidylinositol 4-kinase alpha in plants. Plant Cell 2022, 34, 302–332. [Google Scholar] [CrossRef] [PubMed]
- Thierry-Mieg, D.; Thierry-Mieg, J. AceView: A comprehensive cDNA-supported gene and transcripts annotation. Genome Biol. 2006, 7 (Suppl. S1), S12. [Google Scholar] [CrossRef]
- Available online: https://www.ncbi.nlm.nih.gov/gene/22979 (accessed on 15 January 2025).
- Liu, C.H.; Bollepalli, M.K.; Long, S.V.; Asteriti, S.; Tan, J.; Brill, J.A.; Hardie, R.C. Genetic dissection of the phosphoinositide cycle in Drosophila photoreceptors. J. Cell Sci. 2018, 131, jcs214478. [Google Scholar] [CrossRef]
- Hardie, R.C.; Juusola, M. Phototransduction in Drosophila. Curr. Opin. Neurobiol. 2015, 34, 37–45. [Google Scholar] [CrossRef]
- Montell, C. Drosophila visual transduction. Trends Neurosci. 2012, 35, 356–363. [Google Scholar] [CrossRef]
- Hardie, R.C.; Liu, C.H.; Randall, A.S.; Sengupta, S. In vivo tracking of phosphoinositides in Drosophila photoreceptors. J. Cell Sci. 2015, 128, 4328–4340. [Google Scholar] [CrossRef]
- Balakrishnan, S.S.; Basu, U.; Shinde, D.; Thakur, R.; Jaiswal, M.; Raghu, P. Regulation of PI4P levels by PI4KIIIalpha during G-protein-coupled PLC signaling in Drosophila photoreceptors. J. Cell Sci. 2018, 131, jcs217257. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.ncbi.nlm.nih.gov/datasets/gene/id/23167 (accessed on 20 December 2024).
- Vijayakrishnan, N.; Phillips, S.E.; Broadie, K. Drosophila rolling blackout displays lipase domain-dependent and -independent endocytic functions downstream of dynamin. Traffic 2010, 11, 1567–1578. [Google Scholar] [CrossRef]
- Bojjireddy, N.; Guzman-Hernandez, M.L.; Reinhard, N.R.; Jovic, M.; Balla, T. EFR3s are palmitoylated plasma membrane proteins that control responsiveness to G-protein-coupled receptors. J. Cell Sci. 2015, 128, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Gironacci, M.M.; Bruna-Haupt, E. Unraveling the crosstalk between renin-angiotensin system receptors. Acta Physiol. 2024, 240, e14134. [Google Scholar] [CrossRef]
- Forrester, S.J.; Booz, G.W.; Sigmund, C.D.; Coffman, T.M.; Kawai, T.; Rizzo, V.; Scalia, R.; Eguchi, S. Angiotensin II Signal Transduction: An Update on Mechanisms of Physiology and Pathophysiology. Physiol. Rev. 2018, 98, 1627–1738. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.F.; Sun, Y.L.; Hamet, P.; Inagami, T. The angiotensin II type 1 receptor and receptor-associated proteins. Cell Res. 2001, 11, 165–180. [Google Scholar] [CrossRef]
- Nadel, G.; Yao, Z.; Hacohen-Lev-Ran, A.; Wainstein, E.; Maik-Rachline, G.; Ziv, T.; Naor, Z.; Admon, A.; Seger, R. Phosphorylation of PP2Ac by PKC is a key regulatory step in the PP2A-switch-dependent AKT dephosphorylation that leads to apoptosis. Cell Commun. Signal 2024, 22, 154. [Google Scholar] [CrossRef]
- Streb, H.; Irvine, R.F.; Berridge, M.J.; Schulz, I. Release of Ca2+ from a nonmitochondrial intracellular store in pancreatic acinar cells by inositol-1,4,5-trisphosphate. Nature 1983, 306, 67–69. [Google Scholar] [CrossRef]
- van Gerwen, J.; Shun-Shion, A.S.; Fazakerley, D.J. Insulin signalling and GLUT4 trafficking in insulin resistance. Biochem. Soc. Trans. 2023, 51, 1057–1069. [Google Scholar] [CrossRef]
- Bogan, J.S. Regulation of glucose transporter translocation in health and diabetes. Annu. Rev. Biochem. 2012, 81, 507–532. [Google Scholar] [CrossRef]
- Geiser, A.; Foylan, S.; Tinning, P.W.; Bryant, N.J.; Gould, G.W. GLUT4 dispersal at the plasma membrane of adipocytes: A super-resolved journey. Biosci. Rep. 2023, 43, BSR20230946. [Google Scholar] [CrossRef]
- Sun, Y.; Jaldin-Fincati, J.; Liu, Z.; Bilan, P.J.; Klip, A. A complex of Rab13 with MICAL-L2 and alpha-actinin-4 is essential for insulin-dependent GLUT4 exocytosis. Mol. Biol. Cell 2016, 27, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Stenkula, K.G.; Lizunov, V.A.; Cushman, S.W.; Zimmerberg, J. Insulin controls the spatial distribution of GLUT4 on the cell surface through regulation of its postfusion dispersal. Cell Metab. 2010, 12, 250–259. [Google Scholar] [CrossRef]
- Wieczorke, R.; Dlugai, S.; Krampe, S.; Boles, E. Characterisation of mammalian GLUT glucose transporters in a heterologous yeast expression system. Cell Physiol. Biochem. 2003, 13, 123–134. [Google Scholar] [CrossRef]
- Boles, E.; Oreb, M. A Growth-Based Screening System for Hexose Transporters in Yeast. Methods Mol. Biol. 2018, 1713, 123–135. [Google Scholar] [PubMed]
- Schmidl, S.; Tamayo Rojas, S.A.; Iancu, C.V.; Choe, J.Y.; Oreb, M. Functional Expression of the Human Glucose Transporters GLUT2 and GLUT3 in Yeast Offers Novel Screening Systems for GLUT-Targeting Drugs. Front. Mol. Biosci. 2020, 7, 598419. [Google Scholar] [CrossRef]
- Koester, A.M.; Geiser, A.; Laidlaw, K.M.E.; Morris, S.; Cutiongco, M.F.A.; Stirrat, L.; Gadegaard, N.; Boles, E.; Black, H.L.; Bryant, N.J.; et al. EFR3 and phosphatidylinositol 4-kinase IIIalpha regulate insulin-stimulated glucose transport and GLUT4 dispersal in 3T3-L1 adipocytes. Biosci. Rep. 2022, 42, BSR20221181. [Google Scholar] [CrossRef] [PubMed]
- Koester, A.M.; Geiser, A.; Bowman, P.R.T.; van de Linde, S.; Gadegaard, N.; Bryant, N.J.; Gould, G.W. GLUT4 translocation and dispersal operate in multiple cell types and are negatively correlated with cell size in adipocytes. Sci. Rep. 2022, 12, 20535. [Google Scholar] [CrossRef]
- Qian, Q.; Liu, Q.; Zhou, D.; Pan, H.; Liu, Z.; He, F.; Ji, S.; Wang, D.; Bao, W.; Liu, X.; et al. Brain-specific ablation of Efr3a promotes adult hippocampal neurogenesis via the brain-derived neurotrophic factor pathway. FASEB J. 2017, 31, 2104–2113. [Google Scholar] [CrossRef]
- Nie, C.; Hu, H.; Shen, C.; Ye, B.; Wu, H.; Xiang, M. Expression of EFR3A in the mouse cochlea during degeneration of spiral ganglion following hair cell loss. PLoS ONE 2015, 10, e0117345. [Google Scholar] [CrossRef]
- Hu, H.; Ma, Y.; Ye, B.; Wang, Q.; Yang, T.; Lv, J.; Shi, J.; Wu, H.; Xiang, M. The role of Efr3a in age-related hearing loss. Neuroscience 2017, 341, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Ye, B.; Zhang, L.; Wang, Q.; Liu, Z.; Ji, S.; Liu, Q.; Lv, J.; Ma, Y.; Xu, Y.; et al. Efr3a Insufficiency Attenuates the Degeneration of Spiral Ganglion Neurons after Hair Cell Loss. Front. Mol. Neurosci. 2017, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.R.; Pirruccello, M.; Cheng, F.; Kang, H.J.; Fernandez, T.V.; Baskin, J.M.; Choi, M.; Liu, L.; Ercan-Sencicek, A.G.; Murdoch, J.D.; et al. Rare deleterious mutations of the gene EFR3A in autism spectrum disorders. Mol. Autism 2014, 5, 31. [Google Scholar] [CrossRef]
- Zhao, K.; Bai, X.; Wang, X.; Cao, Y.; Zhang, L.; Li, W.; Wang, S. Insight on the hub gene associated signatures and potential therapeutic agents in epilepsy and glioma. Brain Res. Bull. 2023, 199, 110666. [Google Scholar] [CrossRef]
- He, Y.; Wei, M.; Wu, Y.; Qin, H.; Li, W.; Ma, X.; Cheng, J.; Ren, J.; Shen, Y.; Chen, Z.; et al. Amyloid beta oligomers suppress excitatory transmitter release via presynaptic depletion of phosphatidylinositol-4,5-bisphosphate. Nat. Commun. 2019, 10, 1193. [Google Scholar] [CrossRef]
- Wei, X.; Wang, J.; Yang, E.; Zhang, Y.; Qian, Q.; Li, X.; Huang, F.; Sun, B. Efr3b is essential for social recognition by modulating the excitability of CA2 pyramidal neurons. Proc. Natl. Acad. Sci. USA 2024, 121, e2314557121. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Shen, C.; Jin, X.; Li, X.; Wu, D. Mir-367 is downregulated in coronary artery disaese and its overexpression exerts anti-inflammatory effect via inhibition of the NF-κB-activated inflammatory pathway. Int. J. Clin. Exp. Patholol. 2017, 10, 4047–4057. [Google Scholar]
- Liu, X.H.; Xu, H.; Geng, Q.; Mak, W.-H.; Ling, F.; Su, Z.; Yang, F.; Zhang, T.; Chen, J.; Yang, H.; et al. New genetic variants associated with major adverse cardiovascular events in patients with acute coronary syndromes and treated with clopidogrel and aspirin. Pharmacogen. J. 2021, 21, 664–672. [Google Scholar] [CrossRef]
- Zhou, D.; Yang, L.; Zheng, L.; Ge, W.; Li, D.; Zhang, Y.; Hu, X.; Gao, Z.; Xu, J.; Huang, Y.; et al. Exome capture sequencing of adenoma reveals genetic alterations in multiple cellular pathways at the early stage of colorectal tumorigenesis. PLoS ONE 2013, 8, e53310. [Google Scholar] [CrossRef]
- Adhikari, H.; Kattan, W.E.; Kumar, S.; Zhou, P.; Hancock, J.F.; Counter, C.M. Oncogenic KRAS is dependent upon an EFR3A-PI4KA signaling axis for potent tumorigenic activity. Nat. Commun. 2021, 12, 5248. [Google Scholar] [CrossRef]
- Luo, J. KRAS mutation in pancreatic cancer. Semin. Oncol. 2021, 48, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Kattan, W.E.; Liu, J.; Montufar-Solis, D.; Liang, H.; Brahmendra Barathi, B.; van der Hoeven, R.; Zhou, Y.; Hancock, J.F. Components of the phosphatidylserine endoplasmic reticulum to plasma membrane transport mechanism as targets for KRAS inhibition in pancreatic cancer. Proc. Natl. Acad. Sci. USA 2021, 118, e2114126118. [Google Scholar] [CrossRef]
- Yang, J.; Gong, Y.; Jiang, Q.; Liu, L.; Li, S.; Zhou, Q.; Huang, F.; Liu, Z. Circular RNA Expression Profiles in Nasopharyngeal Carcinoma by Sequence Analysis. Front. Oncol. 2020, 10, 601. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Xia, B. Circular RNA EFR3A promotes nasopharyngeal carcinoma progression through modulating the miR-654-3p/EFR3A axis. Cell. Mol. Biol. 2023, 69, 111–117. [Google Scholar] [CrossRef]
- Conn, V.M.; Chinnaiyan, A.M.; Conn, S.J. Circular RNA in cancer. Nat. Rev. Cancer 2024. online ahead of print. [Google Scholar]
- Toledo, C.M.; Ding, Y.; Hoellerbauer, P.; Davis, R.J.; Basom, R.; Girard, E.J.; Lee, E.; Corrin, P.; Hart, T.; Bolouri, H.; et al. Genome-wide CRISPR-Cas9 Screens Reveal Loss of Redundancy between PKMYT1 and WEE1 in Glioblastoma Stem-like Cells. Cell Rep. 2015, 13, 2425–2439. [Google Scholar] [CrossRef]
- Wu, X.; Chi, R.J.; Baskin, J.M.; Lucast, L.; Burd, C.G.; De Camilli, P.; Reinisch, K.M. Structural insights into assembly and regulation of the plasma membrane phosphatidylinositol 4-kinase complex. Dev. Cell 2014, 28, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Misra, S.; Beach, B.M.; Hurley, J.H. Structure of the VHS domain of human Tom1 (target of myb 1): Insights into interactions with proteins and membranes. Biochemistry 2000, 39, 11282–11290. [Google Scholar] [CrossRef]
- Boal, F.; Mansour, R.; Gayral, M.; Saland, E.; Chicanne, G.; Xuereb, J.M.; Marcellin, M.; Burlet-Schiltz, O.; Sansonetti, P.J.; Payrastre, B.; et al. TOM1 is a PI5P effector involved in the regulation of endosomal maturation. J. Cell Sci. 2015, 128, 815–827. [Google Scholar] [CrossRef]
- Mao, Y.; Nickitenko, A.; Duan, X.; Lloyd, T.E.; Wu, M.N.; Bellen, H.; Quiocho, F.A. Crystal structure of the VHS and FYVE tandem domains of Hrs, a protein involved in membrane trafficking and signal transduction. Cell 2000, 100, 447–456. [Google Scholar] [CrossRef]
- Lohi, O.; Poussu, A.; Mao, Y.; Quiocho, F.; Lehto, V.P. VHS domain—A longshoreman of vesicle lines. FEBS Lett. 2002, 513, 19–23. [Google Scholar] [CrossRef]
- Andrade, M.A.; Petosa, C.; O’Donoghue, S.I.; Muller, C.W.; Bork, P. Comparison of ARM and HEAT protein repeats. J. Mol. Biol. 2001, 309, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Available online: http://pfam.xfam.org/ (accessed on 25 November 2024).
- Kippert, F.; Gerloff, D.L. Highly sensitive detection of individual HEAT and ARM repeats with HHpred and COACH. PLoS ONE 2009, 4, e7148. [Google Scholar] [CrossRef]
- RCSB Protein Data Bank. Available online: https://www.rcsb.org/ (accessed on 25 November 2024).
- Krieger, E.; Vriend, G. YASARA View—Molecular graphics for all devices—From smartphones to workstations. Bioinformatics 2014, 30, 2981–2982. [Google Scholar] [CrossRef] [PubMed]
- Baskin, J.M.; Wu, X.; Christiano, R.; Oh, M.S.; Schauder, C.M.; Gazzerro, E.; Messa, M.; Baldassari, S.; Assereto, S.; Biancheri, R.; et al. The leukodystrophy protein FAM126A (hyccin) regulates PtdIns(4)P synthesis at the plasma membrane. Nat. Cell Biol. 2016, 18, 132–138. [Google Scholar] [CrossRef]
- Lees, J.A.; Zhang, Y.; Oh, M.S.; Schauder, C.M.; Yu, X.; Baskin, J.M.; Dobbs, K.; Notarangelo, L.D.; De Camilli, P.; Walz, T.; et al. Architecture of the human PI4KIIIalpha lipid kinase complex. Proc. Natl. Acad. Sci. USA 2017, 114, 13720–13725. [Google Scholar] [CrossRef]
- Suresh, S.; Shaw, A.L.; Pemberton, J.G.; Scott, M.K.; Harris, N.J.; Parson, M.A.H.; Jenkins, M.L.; Rohilla, P.; Alvarez-Prats, A.; Balla, T.; et al. Molecular basis for plasma membrane recruitment of PI4KA by EFR3. Sci. Adv. 2024, 10, eadp6660. [Google Scholar] [CrossRef] [PubMed]
- Korycka, J.; Lach, A.; Heger, E.; Boguslawska, D.M.; Wolny, M.; Toporkiewicz, M.; Augoff, K.; Korzeniewski, J.; Sikorski, A.F. Human DHHC proteins: A spotlight on the hidden player of palmitoylation. Eur. J. Cell Biol. 2012, 91, 107–117. [Google Scholar] [CrossRef]
- Tabaczar, S.; Czogalla, A.; Podkalicka, J.; Biernatowska, A.; Sikorski, A.F. Protein palmitoylation: Palmitoyltransferases and their specificity. Exp. Biol. Med. 2017, 242, 1150–1157.Efr3. [Google Scholar] [CrossRef]
- Pei, S.; Piao, H.L. Exploring Protein S-Palmitoylation: Mechanisms, Detection, and Strategies for Inhibitor Discovery. ACS Chem. Biol. 2024, 19, 1868–1882. [Google Scholar] [CrossRef]
- Ren, J.; Wen, L.; Gao, X.; Jin, C.; Xue, Y.; Yao, X. CSS-Palm 2.0: An updated software for palmitoylation sites prediction. Protein Eng. Des. Sel. 2008, 21, 639–644. [Google Scholar] [CrossRef]
- Blanc, M.; David, F.P.A.; van der Goot, F.G. SwissPalm 2: Protein S-Palmitoylation Database. Methods Mol. Biol. 2019, 2009, 203–214. [Google Scholar] [PubMed]
- Batrouni, A.G.; Bag, N.; Phan, H.T.; Baird, B.A.; Baskin, J.M. A palmitoylation code controls PI4KIIIalpha complex formation and PI(4,5)P2 homeostasis at the plasma membrane. J. Cell Sci. 2022, 135, jcs259365. [Google Scholar] [CrossRef] [PubMed]
- Nakatsu, F.; Baskin, J.M.; Chung, J.; Tanner, L.B.; Shui, G.; Lee, S.Y.; Pirruccello, M.; Hao, M.; Ingolia, N.T.; Wenk, M.R.; et al. PtdIns4P synthesis by PI4KIIIalpha at the plasma membrane and its impact on plasma membrane identity. J. Cell Biol. 2012, 199, 1003–1016. [Google Scholar] [CrossRef] [PubMed]
- Hornbeck, P.V.; Zhang, B.; Murray, B.; Kornhauser, J.M.; Latham, V.; Skrzypek, E. PhosphoSitePlus, 2014: Mutations, PTMs and recalibrations. Nucleic Acids Res. 2015, 43, D512–D520. [Google Scholar] [CrossRef]
- Available online: https://string-db.org/ (accessed on 20 December 2024).
- Roux, K.J.; Kim, D.I.; Burke, B.; May, D.G. BioID: A Screen for Protein-Protein Interactions. Curr. Protoc. Protein Sci. 2018, 91, 19.23.1–19.23.15. [Google Scholar] [CrossRef]
- Dong, J.M.; Tay, F.P.; Swa, H.L.; Gunaratne, J.; Leung, T.; Burke, B.; Manser, E. Proximity biotinylation provides insight into the molecular composition of focal adhesions at the nanometer scale. Sci. Signal 2016, 9, rs4. [Google Scholar] [CrossRef]
- Falkenburger, B.H.; Jensen, J.B.; Dickson, E.J.; Suh, B.C.; Hille, B. Phosphoinositides: Lipid regulators of membrane proteins. J. Physiol. 2010, 588, 3179–3185. [Google Scholar] [CrossRef]
- Katan, M.; Cockcroft, S. Phosphatidylinositol(4,5)bisphosphate: Diverse functions at the plasma membrane. Essays Biochem. 2020, 64, 513–531. [Google Scholar]
- Audhya, A.; Emr, S.D. Stt4 PI 4-kinase localizes to the plasma membrane and functions in the Pkc1-mediated MAP kinase cascade. Dev. Cell 2002, 2, 593–605. [Google Scholar] [CrossRef]
- Posor, Y.; Jang, W.; Haucke, V. Phosphoinositides as membrane organizers. Nat. Rev. Mol. Cell Biol. 2022, 23, 797–816. [Google Scholar] [CrossRef]
- Ray, J.; Sapp, D.G.; Fairn, G.D. Phosphatidylinositol 3,4-bisphosphate: Out of the shadows and into the spotlight. Curr. Opin. Cell Biol. 2024, 88, 102372. [Google Scholar] [CrossRef] [PubMed]
- Hammond, G.R.; Machner, M.P.; Balla, T. A novel probe for phosphatidylinositol 4-phosphate reveals multiple pools beyond the Golgi. J. Cell Biol. 2014, 205, 113–126. [Google Scholar] [CrossRef]
- Salazar, A.N.; Gorter de Vries, A.R.; van den Broek, M.; Wijsman, M.; de la Torre Cortes, P.; Brickwedde, A.; Brouwers, N.; Daran, J.G.; Abeel, T. Nanopore sequencing enables near-complete de novo assembly of Saccharomyces cerevisiae reference strain CEN.PK113-7D. FEMS Yeast Res. 2017, 17, fox074. [Google Scholar] [CrossRef]
- Wong, K.; Cantley, L.C. Cloning and characterization of a human phosphatidylinositol 4-kinase. J. Biol. Chem. 1994, 269, 28878–28884. [Google Scholar] [CrossRef]
- Yoshida, S.; Ohya, Y.; Goebl, M.; Nakano, A.; Anraku, Y. A novel gene, STT4, encodes a phosphatidylinositol 4-kinase in the PKC1 protein kinase pathway of Saccharomyces cerevisiae. J. Biol. Chem. 1994, 269, 1166–1172. [Google Scholar] [CrossRef] [PubMed]
- Cutler, N.S.; Heitman, J.; Cardenas, M.E. STT4 is an essential phosphatidylinositol 4-kinase that is a target of wortmannin in Saccharomyces cerevisiae. J. Biol. Chem. 1997, 272, 27671–27677. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rodas, R.; Labbaoui, H.; Orange, F.; Solis, N.; Zaragoza, O.; Filler, S.G.; Bassilana, M.; Arkowitz, R.A. Plasma Membrane Phosphatidylinositol-4-Phosphate Is Not Necessary for Candida albicans Viability yet Is Key for Cell Wall Integrity and Systemic Infection. mBio 2021, 13, e0387321. [Google Scholar] [CrossRef]
- Scheufler, C.; Brinker, A.; Bourenkov, G.; Pegoraro, S.; Moroder, L.; Bartunik, H.; Hartl, F.U.; Moarefi, I. Structure of TPR domain-peptide complexes: Critical elements in the assembly of the Hsp70-Hsp90 multichaperone machine. Cell 2000, 101, 199–210. [Google Scholar] [CrossRef]
- Izert, M.A.; Szybowska, P.E.; Gorna, M.W.; Merski, M. The Effect of Mutations in the TPR and Ankyrin Families of Alpha Solenoid Repeat Proteins. Front. Bioinform. 2021, 1, 696368. [Google Scholar] [CrossRef]
- Baird, D.; Stefan, C.; Audhya, A.; Weys, S.; Emr, S.D. Assembly of the PtdIns 4-kinase Stt4 complex at the plasma membrane requires Ypp1 and Efr3. J. Cell Biol. 2008, 183, 1061–1074. [Google Scholar] [CrossRef]
- Agarwal, N.S.; Northrop, L.; Anyane-Yeboa, K.; Aggarwal, V.S.; Nagy, P.L.; Demirdag, Y.Y. Tetratricopeptide repeat domain 7A (TTC7A) mutation in a newborn with multiple intestinal atresia and combined immunodeficiency. J. Clin. Immunol. 2014, 34, 607–610. [Google Scholar] [CrossRef]
- Jardine, S.; Dhingani, N.; Muise, A.M. TTC7A: Steward of Intestinal Health. Cell Mol. Gastroenterol. Hepatol. 2019, 7, 555–570. [Google Scholar] [CrossRef] [PubMed]
- Sharafian, S.; Alimadadi, H.; Shahrooei, M.; Gharagozlou, M.; Aghamohammadi, A.; Parvaneh, N. A Novel TTC7A Deficiency Presenting With Combined Immunodeficiency and Chronic Gastrointestinal Problems. J. Investig. Allergol. Clin. Immunol. 2018, 28, 358–360. [Google Scholar] [CrossRef] [PubMed]
- Mou, W.; Yang, S.; Guo, R.; Fu, L.; Zhang, L.; Guo, W.; Du, J.; He, J.; Ren, Q.; Hao, C.; et al. A Novel Homozygous TTC7A Missense Mutation Results in Familial Multiple Intestinal Atresia and Combined Immunodeficiency. Front. Immunol. 2021, 12, 759308. [Google Scholar] [CrossRef]
- Dannheim, K.; Ouahed, J.; Field, M.; Snapper, S.; Raphael, B.P.; Glover, S.C.; Bishop, P.R.; Bhesania, N.; Kamin, D.; Thiagarajah, J.; et al. Pediatric Gastrointestinal Histopathology in Patients with Tetratricopeptide Repeat Domain 7A (TTC7A) Germline Mutations: A Rare Condition Leading to Multiple Intestinal Atresias, Severe Combined Immunodeficiency, and Congenital Enteropathy. Am. J. Surg. Pathol. 2022, 46, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Hypomyelinating Leukoencephalopathy. Available online: https://www.malacards.org/card/hypomyelinating_leukoencephalopathy (accessed on 10 January 2025).
- Gazzerro, E.; Baldassari, S.; Giacomini, C.; Musante, V.; Fruscione, F.; La Padula, V.; Biancheri, R.; Scarfi, S.; Prada, V.; Sotgia, F.; et al. Hyccin, the molecule mutated in the leukodystrophy hypomyelination and congenital cataract (HCC), is a neuronal protein. PLoS ONE 2012, 7, e32180. [Google Scholar] [CrossRef]
- Traverso, M.; Yuregir, O.O.; Mimouni-Bloch, A.; Rossi, A.; Aslan, H.; Gazzerro, E.; Baldassari, S.; Fruscione, F.; Minetti, C.; Zara, F.; et al. Hypomyelination and congenital cataract: Identification of novel mutations in two unrelated families. Eur. J. Paediatr. Neurol. 2013, 17, 108–111. [Google Scholar] [CrossRef]
- Li, S.; Han, T. Frequent loss of FAM126A expression in colorectal cancer results in selective FAM126B dependency. iScience 2024, 27, 109646. [Google Scholar] [CrossRef]
- Dornan, G.L.; Dalwadi, U.; Hamelin, D.J.; Hoffmann, R.M.; Yip, C.K.; Burke, J.E. Probing the Architecture, Dynamics, and Inhibition of the PI4KIIIalpha/TTC7/FAM126 Complex. J. Mol. Biol. 2018, 430, 3129–3142. [Google Scholar] [CrossRef]
- Chung, J.; Nakatsu, F.; Baskin, J.M.; De Camilli, P. Plasticity of PI4KIIIalpha interactions at the plasma membrane. EMBO Rep. 2015, 16, 312–320. [Google Scholar] [CrossRef]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef]
- Sezgin, E.; Levental, I.; Mayor, S.; Eggeling, C. The mystery of membrane organization: Composition, regulation and roles of lipid rafts. Nat. Rev. Mol. Cell Biol. 2017, 18, 361–374. [Google Scholar] [CrossRef]
- Yuan, Z.; Hansen, S.B. Cholesterol Regulation of Membrane Proteins Revealed by Two-Color Super-Resolution Imaging. Membranes 2023, 13, 250. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.G.N.; Kusumi, A. Refinement of Singer-Nicolson fluid-mosaic model by microscopy imaging: Lipid rafts and actin-induced membrane compartmentalization. Biochim. Biophys. Acta Biomembr. 2023, 1865, 184093. [Google Scholar] [CrossRef] [PubMed]
- Saldana-Villa, A.K.; Lara-Lemus, R. The Structural Proteins of Membrane Rafts, Caveolins and Flotillins, in Lung Cancer: More Than Just Scaffold Elements. Int. J. Med. Sci. 2023, 20, 1662–1670. [Google Scholar] [CrossRef]
- Neumann-Giesen, C.; Falkenbach, B.; Beicht, P.; Claasen, S.; Luers, G.; Stuermer, C.A.; Herzog, V.; Tikkanen, R. Membrane and raft association of reggie-1/flotillin-2: Role of myristoylation, palmitoylation and oligomerization and induction of filopodia by overexpression. Biochem. J. 2004, 378, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Fantini, J.; Barrantes, F.J. How cholesterol interacts with membrane proteins: An exploration of cholesterol-binding sites including CRAC, CARC, and tilted domains. Front. Physiol. 2013, 4, 31. [Google Scholar] [CrossRef]
- Suzuki, K.G.N. Single-Molecule Imaging of Ganglioside Probes in Living Cell Plasma Membranes. Methods Mol. Biol. 2023, 2613, 215–227. [Google Scholar]
- Komatsuya, K.; Kikuchi, N.; Hirabayashi, T.; Kasahara, K. The Regulatory Roles of Cerebellar Glycosphingolipid Microdomains/Lipid Rafts. Int. J. Mol. Sci. 2023, 24, 5566. [Google Scholar] [CrossRef]
- Ruzzi, F.; Cappello, C.; Semprini, M.S.; Scalambra, L.; Angelicola, S.; Pittino, O.M.; Landuzzi, L.; Palladini, A.; Nanni, P.; Lollini, P.L. Lipid rafts, caveolae, and epidermal growth factor receptor family: Friends or foes? Cell Commun. Signal 2024, 22, 489. [Google Scholar] [CrossRef]
- Isik, O.A.; Cizmecioglu, O. Rafting on the Plasma Membrane: Lipid Rafts in Signaling and Disease. Adv. Exp. Med. Biol. 2023, 1436, 87–108. [Google Scholar] [PubMed]
- Shouman, S.; El-Kholy, N.; Hussien, A.E.; El-Derby, A.M.; Magdy, S.; Abou-Shanab, A.M.; Elmehrath, A.O.; Abdelwaly, A.; Helal, M.; El-Badri, N. SARS-CoV-2-associated lymphopenia: Possible mechanisms and the role of CD147. Cell Commun. Signal 2024, 22, 349. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.; Temerozo, J.R.; Pereira-Dutra, F.S.; Ferreira, A.C.; Mattos, M.; Goncalves, B.S.; Sacramento, C.Q.; Palhinha, L.; Cunha-Fernandes, T.; Dias, S.S.G.; et al. Simvastatin Downregulates the SARS-CoV-2-Induced Inflammatory Response and Impairs Viral Infection Through Disruption of Lipid Rafts. Front. Immunol. 2022, 13, 820131. [Google Scholar] [CrossRef]
- Zhang, S.; Zhu, N.; Li, H.F.; Gu, J.; Zhang, C.J.; Liao, D.F.; Qin, L. The lipid rafts in cancer stem cell: A target to eradicate cancer. Stem Cell Res. Ther. 2022, 13, 432. [Google Scholar] [CrossRef] [PubMed]
- Erazo-Oliveras, A.; Munoz-Vega, M.; Salinas, M.L.; Wang, X.; Chapkin, R.S. Dysregulation of cellular membrane homeostasis as a crucial modulator of cancer risk. FEBS J. 2024, 291, 1299–1352. [Google Scholar] [CrossRef]
- Mollinedo, F.; Gajate, C. Lipid rafts as signaling hubs in cancer cell survival/death and invasion: Implications in tumor progression and therapy: Thematic Review Series: Biology of Lipid Rafts. J. Lipid Res. 2020, 61, 611–635. [Google Scholar] [CrossRef]
- Hryniewicz-Jankowska, A.; Augoff, K.; Biernatowska, A.; Podkalicka, J.; Sikorski, A.F. Membrane rafts as a novel target in cancer therapy. Biochim. Biophys. Acta 2014, 1845, 155–165. [Google Scholar] [CrossRef]
- Ferrer, I. Amyloid-beta Pathology Is the Common Nominator Proteinopathy of the Primate Brain Aging. J. Alzheimers Dis. 2024, 100, S153–S164. [Google Scholar] [CrossRef]
- Kwiatkowska, K.; Matveichuk, O.V.; Fronk, J.; Ciesielska, A. Flotillins: At the Intersection of Protein S-Palmitoylation and Lipid-Mediated Signaling. Int. J. Mol. Sci. 2020, 21, 2283. [Google Scholar] [CrossRef]
- Trybus, M.; Niemiec, L.; Biernatowska, A.; Hryniewicz-Jankowska, A.; Sikorski, A.F. MPP1-based mechanism of resting state raft organization in the plasma membrane. Is it a general or specialized mechanism in erythroid cells? Folia Histochem. Cytobiol. 2019, 57, 43–55. [Google Scholar] [CrossRef]
- Chytla, A.; Gajdzik-Nowak, W.; Olszewska, P.; Biernatowska, A.; Sikorski, A.F.; Czogalla, A. Not Just Another Scaffolding Protein Family: The Multifaceted MPPs. Molecules 2020, 25, 4954. [Google Scholar] [CrossRef] [PubMed]
- Biernatowska, A.; Podkalicka, J.; Majkowski, M.; Hryniewicz-Jankowska, A.; Augoff, K.; Kozak, K.; Korzeniewski, J.; Sikorski, A.F. The role of MPP1/p55 and its palmitoylation in resting state raft organization in HEL cells. Biochim. Biophys. Acta 2013, 1833, 1876–1884. [Google Scholar] [CrossRef]
- Podkalicka, J.; Biernatowska, A.; Majkowski, M.; Grzybek, M.; Sikorski, A.F. MPP1 as a Factor Regulating Phase Separation in Giant Plasma Membrane-Derived Vesicles. Biophys. J. 2015, 108, 2201–2211. [Google Scholar] [CrossRef]
- Biernatowska, A.; Wojtowicz, K.; Trombik, T.; Sikorski, A.F.; Czogalla, A. MPP1 Determines the Mobility of Flotillins and Controls the Confinement of Raft-Associated Molecules. Cells 2022, 11, 311. [Google Scholar] [CrossRef] [PubMed]
- Podkalicka, J.; Biernatowska, A.; Olszewska, P.; Tabaczar, S.; Sikorski, A.F. The microdomain-organizing protein MPP1 is required for insulin-stimulated activation of H-Ras. Oncotarget 2018, 9, 18410–18421. [Google Scholar] [CrossRef]
- Biernatowska, A.; Augoff, K.; Podkalicka, J.; Tabaczar, S.; Gajdzik-Nowak, W.; Czogalla, A.; Sikorski, A.F. MPP1 directly interacts with flotillins in erythrocyte membrane—Possible mechanism of raft domain formation. Biochim. Biophys. Acta Biomembr. 2017, 1859, 2203–2212. [Google Scholar] [CrossRef]
- Biernatowska, A.; Olszewska, P.; Grzymajlo, K.; Drabik, D.; Kraszewski, S.; Sikorski, A.F.; Czogalla, A. Molecular characterization of direct interactions between MPP1 and flotillins. Sci. Rep. 2021, 11, 14751. [Google Scholar] [CrossRef] [PubMed]
- Trybus, M.; Hryniewicz-Jankowska, A.; Wojtowicz, K.; Trombik, T.; Czogalla, A.; Sikorski, A.F. EFR3A: A new raft domain organizing protein? Cell Mol. Biol. Lett. 2023, 28, 86. [Google Scholar] [CrossRef]
- Brown, D.A. Analysis of raft affinity of membrane proteins by detergent-insolubility. Methods Mol. Biol. 2007, 398, 9–20. [Google Scholar]
- Sezgin, E.; Kaiser, H.J.; Baumgart, T.; Schwille, P.; Simons, K.; Levental, I. Elucidating membrane structure and protein behavior using giant plasma membrane vesicles. Nat. Protoc. 2012, 7, 1042–1051. [Google Scholar] [CrossRef]
- Simons, K.; Toomre, D. Lipid rafts and signal transduction. Nat. Rev. Mol. Cell Biol. 2000, 1, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; MacKinnon, R. Structure of the flotillin complex in a native membrane environment. Proc. Natl. Acad. Sci. USA 2024, 121, e2409334121. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trybus, M.; Hryniewicz-Jankowska, A.; Czogalla, A.; Sikorski, A.F. EFR3A, an Intriguing Gene, and Protein with a Scaffolding Function. Cells 2025, 14, 445. https://doi.org/10.3390/cells14060445
Trybus M, Hryniewicz-Jankowska A, Czogalla A, Sikorski AF. EFR3A, an Intriguing Gene, and Protein with a Scaffolding Function. Cells. 2025; 14(6):445. https://doi.org/10.3390/cells14060445
Chicago/Turabian StyleTrybus, Magdalena, Anita Hryniewicz-Jankowska, Aleksander Czogalla, and Aleksander F. Sikorski. 2025. "EFR3A, an Intriguing Gene, and Protein with a Scaffolding Function" Cells 14, no. 6: 445. https://doi.org/10.3390/cells14060445
APA StyleTrybus, M., Hryniewicz-Jankowska, A., Czogalla, A., & Sikorski, A. F. (2025). EFR3A, an Intriguing Gene, and Protein with a Scaffolding Function. Cells, 14(6), 445. https://doi.org/10.3390/cells14060445