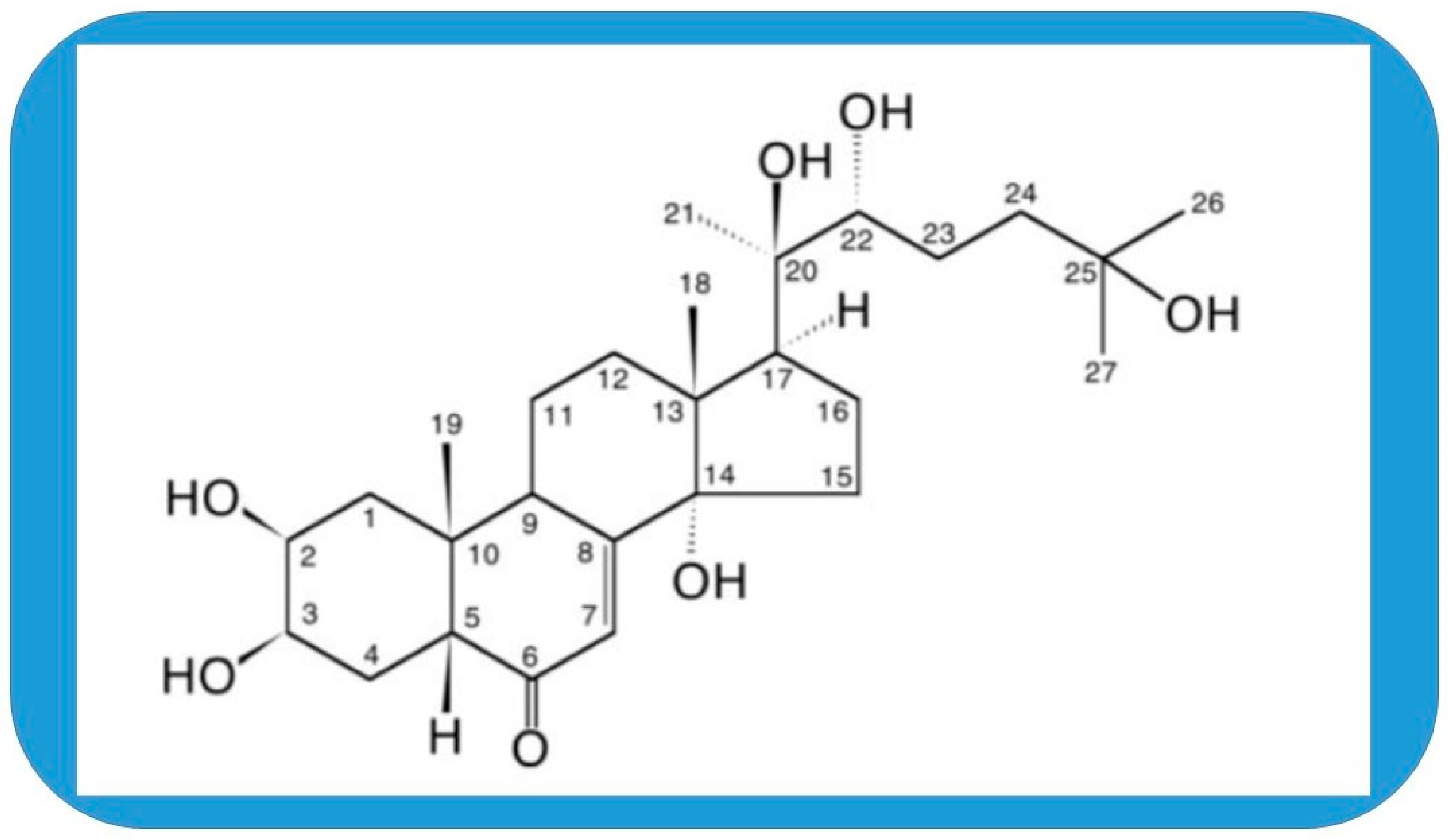
Molecular Targets of 20-Hydroxyecdysone in Mammals, Mechanism of Action: Is It a Calorie Restriction Mimetic and Anti-Aging Compound?


{kind=link}
{kind=link}
Abstract
1. Introduction
2. Cell, Tissue, and Systematic Action
2.1. Effects on Skeletal Muscle
2.2. Effects on Skin, Bone, and Cartilage
2.3. Influence on Nervous System
2.4. Actions on Inflammation and Apoptosis
2.5. Impact on Liver and Adipose Tissue
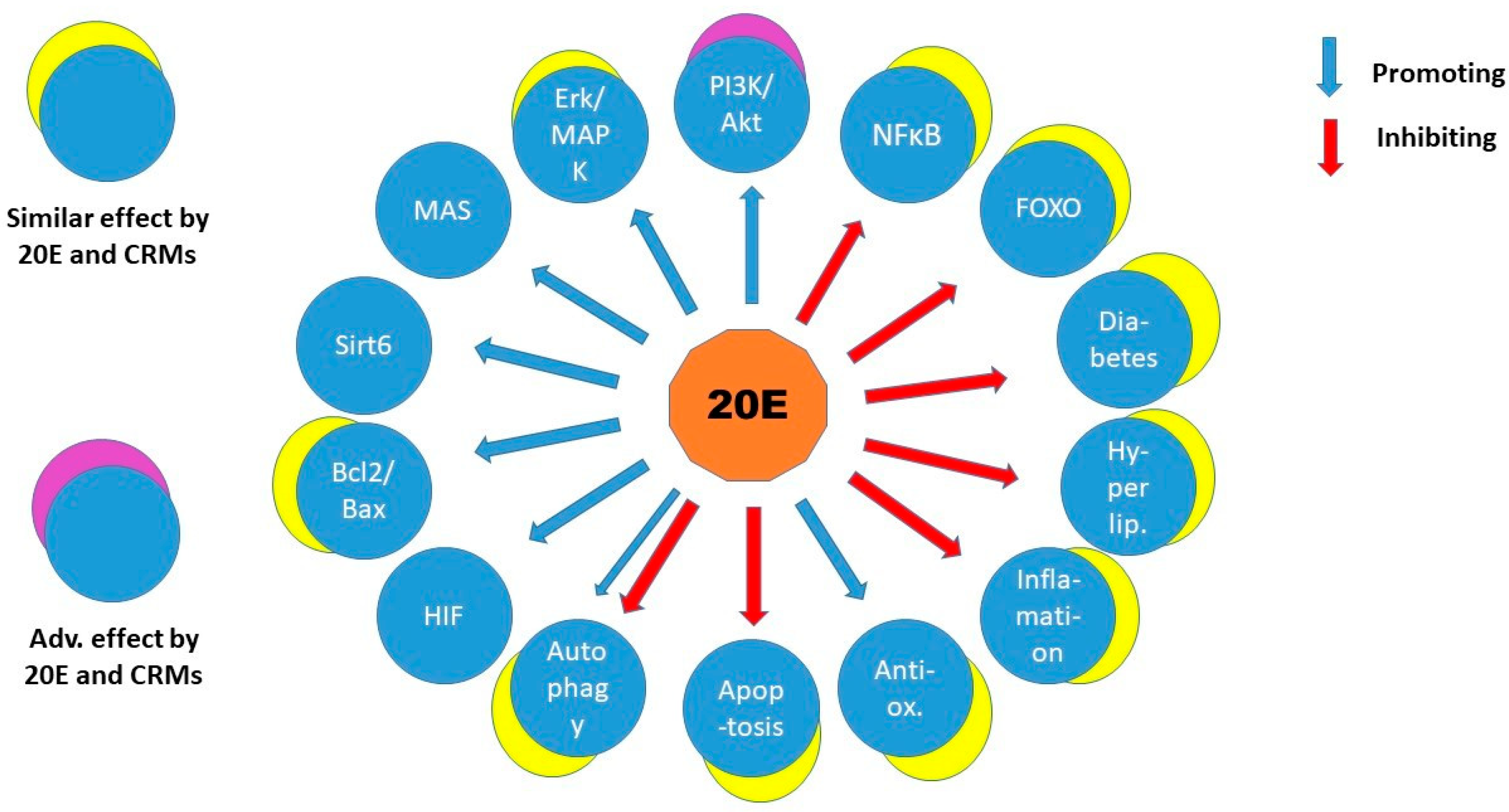
2.6. Calorie Restriction Mimetics and 20E
3. Discussion
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dinan, L.; Dioh, W.; Veillet, S.; Lafont, R. 20-Hydroxyecdysone, from Plant Extracts to Clinical Use: Therapeutic Potential for the Treatment of Neuromuscular, Cardio-Metabolic and Respiratory Diseases. Biomedicines 2021, 9, 492. [Google Scholar] [CrossRef] [PubMed]
- Dioh, W.; Tourette, C.; Del Signore, S.; Daudigny, L.; Dupont, P.; Balducci, C.; Dilda, P.J.; Lafont, R.; Veillet, S. A Phase 1 study for safety and pharmacokinetics of BIO101 (20-hydroxyecdysone) in healthy young and older adults. J. Cachexia Sarcopenia Muscle 2023, 14, 1259–1273. [Google Scholar] [CrossRef]
- Lobo, S.M.; Plantefève, G.; Nair, G.; Cavalcante, A.J.; de Moraes, N.F.; Nunes, E.; Barnum, O.; Stadnik, C.M.B.; Lima, M.P.; Lins, M.; et al. Efficacy of oral 20-hydroxyecdysone (BIO101), a MAS receptor activator, in adults with severe COVID-19 (COVA): A randomized, placebo-controlled, phase 2/3 trial. EClinicalMedicine 2024, 68, 102383. [Google Scholar] [CrossRef]
- Báthori, M.; Tóth, N.; Hunyadi, A.; Márki, A.; Zádor, E. Phytoecdysteroids and anabolic-androgenic steroids—Structure and effects on humans. Curr. Med. Chem. 2008, 15, 75–91. [Google Scholar] [CrossRef] [PubMed]
- Tóth, N.; Hunyadi, A.; Báthori, M.; Zádor, E. Phytoecdysteroids and vitamin D analogues—Similarities in structure and mode of action. Curr. Med. Chem. 2010, 17, 1974–1994. [Google Scholar] [CrossRef]
- Berger, K.; Schiefner, F.; Rudolf, M.; Awiszus, F.; Junne, F.; Vogel, M.; Lohmann, C.H. Long-term effects of doping with anabolic steroids during adolescence on physical and mental health. Orthopadie 2024, 53, 608–616. [Google Scholar] [CrossRef]
- Lafont, R.; Balducci, C.; Dinan, L. Ecdysteroids. Encyclopedia 2021, 1, 1267–1302. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Bajguz, A.; Hayat, S. Phytoecdysteroids: Distribution, Structural Diversity, Biosynthesis, Activity, and Crosstalk with Phytohormones. Int. J. Mol. Sci. 2022, 23, 8664. [Google Scholar] [CrossRef]
- Gorelick-Feldman, J.; Cohick, W.; Raskin, I. Ecdysteroids elicit a rapid Ca2+ flux leading to Akt activation and increased protein synthesis in skeletal muscle cells. Steroids 2010, 75, 632–637. [Google Scholar] [CrossRef]
- Zhao, X.F. G protein-coupled receptors function as cell membrane receptors for the steroid hormone 20-hydroxyecdysone. Cell Commun. Signal 2020, 18, 146. [Google Scholar] [CrossRef]
- Parr, M.K.; Zhao, P.; Haupt, O.; Ngueu, S.T.; Hengevoss, J.; Fritzemeier, K.H.; Piechotta, M.; Schlörer, N.; Muhn, P.; Zheng, W.; et al. Estrogen receptor beta is involved in skeletal muscle hypertrophy induced by the phytoecdysteroid ecdysterone. Mol. Nutr. Food Res. 2014, 58, 1861–1872. [Google Scholar] [CrossRef] [PubMed]
- Lafont, R.; Serova, M.; Didry-Barca, B.; Raynal, S.; Guibout, L.; Dinan, L.; Veillet, S.; Latil, M.; Dioh, W.; Dilda, P.J. 20-Hydroxyecdysone activates the protective arm of the RAAS via the MAS receptor. J. Mol. Endocrinol. 2021, 68, 77–87. [Google Scholar] [CrossRef]
- Jin, Z.; Wang, B.; Ren, L.; Yang, J.; Zheng, Z.; Yao, F.; Ding, R.; Wang, J.; He, J.; Wang, W.; et al. 20-Hydroxyecdysone inhibits inflammation via SIRT6-mediated NF-κB signaling in endothelial cells. Biochim. Biophys. Acta Mol. Cell Res. 2023, 1870, 119460. [Google Scholar] [CrossRef]
- Gorelick-Feldman, J.; Maclean, D.; Ilic, N.; Poulev, A.; Lila, M.A.; Cheng, D.; Raskin, I. Phytoecdysteroids increase protein synthesis in skeletal muscle cells. J. Agric. Food Chem. 2008, 56, 3532–3537. [Google Scholar] [CrossRef]
- Hirunsai, M.; Yimlamai, T.; Suksamrarn, A. Effect of 20-Hydroxyecdysone on Proteolytic Regulation in Skeletal Muscle Atrophy. In Vivo 2016, 30, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Anthony, T.G.; Mirek, E.T.; Bargoud, A.R.; Phillipson-Weiner, L.; DeOliveira, C.M.; Wetstein, B.; Graf, B.L.; Kuhn, P.E.; Raskin, I. Evaluating the effect of 20-hydroxyecdysone (20HE) on mechanistic target of rapamycin complex 1 (mTORC1) signaling in the skeletal muscle and liver of rats. Appl. Physiol. Nutr. Metab. 2015, 40, 1324–1328. [Google Scholar] [CrossRef] [PubMed]
- Tóth, N.; Szabó, A.; Kacsala, P.; Héger, J.; Zádor, E. 20-Hydroxyecdysone increases fibre size in a muscle-specific fashion in rat. Phytomedicine 2008, 15, 691–698. [Google Scholar] [CrossRef]
- Csábi, J.; Rafai, T.; Hunyadi, A.; Zádor, E. Poststerone increases muscle fibre size partly similar to its metabolically parent compound, 20-hydroxyecdysone. Fitoterapia 2019, 134, 459–464. [Google Scholar] [CrossRef]
- Isenmann, E.; Ambrosio, G.; Joseph, J.F.; Mazzarino, M.; de la Torre, X.; Zimmer, P.; Kazlauskas, R.; Goebel, C.; Botrè, F.; Diel, P.; et al. Ecdysteroids as non-conventional anabolic agent: Performance enhancement by ecdysterone supplementation in humans. Arch. Toxicol. 2019, 93, 1807–1816. [Google Scholar] [CrossRef]
- The WADA 2020 Monitoring Program. Available online: https://www.wada-ama.org/en/resources/scientific-research/ecdysterone-detection-anti-doping-control-restricted-use-or-regular (accessed on 12 March 2025).
- Zwetsloot, K.A.; Shanely, R.A.; Godwin, J.S.; Hodgman, C.F. Phytoecdysteroids Accelerate Recovery of Skeletal Muscle Function Following in vivo Eccentric Contraction-Induced Injury in Adult and Old Mice. Front. Rehabil. Sci. 2021, 2, 757789. [Google Scholar] [CrossRef]
- Lawrence, M.M.; Zwetsloot, K.A.; Arthur, S.T.; Sherman, C.A.; Huot, J.R.; Badmaev, V.; Grace, M.; Lila, M.A.; Nieman, D.C.; Shanely, R.A. Phytoecdysteroids Do Not Have Anabolic Effects in Skeletal Muscle in Sedentary Aging Mice. Int. J. Environ. Res. Public Health 2021, 18, 370. [Google Scholar] [CrossRef]
- Cheng, D.M.; Kutzler, L.W.; Boler, D.D.; Drnevich, J.; Killefer, J.; Lila, M.A. Continuous infusion of 20-hydroxyecdysone increased mass of triceps brachii in C57BL/6 mice. Phytother. Res. 2013, 27, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Serova, M.; Didry-Barca, B.; Deloux, R.; Foucault, A.; Veillet, S.; Lafont, R.; Dilda, P.J.; Latil, M. BIO101 stimulates myoblast differentiation and improves muscle function in adult and old mice. J. Cachexia Sarcopenia Muscle 2024, 15, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Todorova, V.; Ivanova, S.; Chakarov, D.; Kraev, K.; Ivanov, K. Ecdysterone and Turkesterone-Compounds with Prominent Potential in Sport and Healthy Nutrition. Nutrients 2024, 16, 1382. [Google Scholar] [CrossRef]
- Afsar, B.; Afsar, R.E.; Caliskan, Y.; Lentine, K.L.; Edwards, J.C. Renin angiotensin system-induced muscle wasting: Putative mechanisms and implications for clinicians. Mol. Cell Biochem. 2024. [Google Scholar] [CrossRef]
- Gao, L.; Cai, G.; Shi, X. Beta-ecdysterone induces osteogenic differentiation in mouse mesenchymal stem cells and relieves osteoporosis. Biol. Pharm. Bull. 2008, 31, 2245–2249. [Google Scholar] [CrossRef]
- Seidlova-Wuttke, D.; Christel, D.; Kapur, P.; Nguyen, B.T.; Jarry, H.; Wuttke, W. Beta-ecdysone has bone protective but no estrogenic effects in ovariectomized rats. Phytomedicine 2010, 17, 884–889. [Google Scholar] [CrossRef] [PubMed]
- Kapur, P.; Wuttke, W.; Jarry, H.; Seidlova-Wuttke, D. Beneficial effects of beta-Ecdysone on the joint, epiphyseal cartilage tissue and trabecular bone in ovariectomized rats. Phytomedicine 2010, 17, 350–355. [Google Scholar] [CrossRef]
- Ehrhardt, C.; Wessels, J.T.; Wuttke, W.; Seidlová-Wuttke, D. The effects of 20-hydroxyecdysone and 17β-estradiol on the skin of ovariectomized rats. Menopause 2011, 18, 323–327. [Google Scholar] [CrossRef]
- Jian, C.-X.; Liu, X.-F.; Hu, J.; Li, C.-J.; Zhang, G.; Li, Y.; Zhu, J.-W.; Tan, Y.-H. 20-Hydroxyecdysone-induced bone morphogenetic protein-2-dependent osteogenic differentiation through the ERK pathway in human periodontal ligament stem cells. Eur. J. Pharmacol. 2013, 698, 48–56. [Google Scholar] [CrossRef]
- Dai, W.; Jiang, L.; Lay, Y.-A.E.; Chen, H.; Jin, G.; Zhang, H.; Kot, A.; Ritchie, R.O.; Lane, N.E.; Yao, W. Prevention of glucocorticoid induced bone changes with beta-ecdysone. Bone 2015, 74, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Zhang, H.; Zhong, Z.A.; Jiang, L.; Chen, H.; Lay, Y.A.; Kot, A.; Ritchie, R.O.; Lane, N.E.; Yao, W. β-Ecdysone Augments Peak Bone Mass in Mice of Both Sexes. Clin. Orthop. Relat. Res. 2015, 473, 2495–2504. [Google Scholar] [CrossRef]
- Peschel, W.; Kump, A.; Prieto, J.M. Effects of 20-hydroxyecdysone, Leuzea carthamoides extracts, dexamethasone and their combinations on the NF-κB activation in HeLa cells. J. Pharm. Pharmacol. 2011, 63, 1483–1495. [Google Scholar] [CrossRef]
- Park, I.; Park, K.; Lee, H.-S.; Hong, S.-M.; Sriramulu, D.K.; Hwang, H.; Song, S.; Baek, J.G.; Kim, D.H.; Kim, S.Y.; et al. Ecdysteroids from the Korean Endemic Species Ajuga spectabilis with Activities against Glucocorticoid Receptors and 11β-Hydroxysteroid Dehydrogenase Type 1. ACS Omega 2023, 8, 26191–26200. [Google Scholar] [CrossRef]
- Lim, H.S.; Yoon, K.; Lee, D.H.; Lee, Y.S.; Chung, J.H.; Park, G. Effects of 20-hydroxyecdysone on UVB-induced photoaging in hairless mice. Biomed. Pharmacother. 2023, 164, 114899. [Google Scholar] [CrossRef]
- Dai, W.-W.; Wang, L.-B.; Jin, G.-Q.; Wu, H.-J.; Zhang, J.; Wang, C.-L.; Wei, Y.-J.; Lee, J.-H.; Lay, Y.-A.E.; Yao, W. Beta-Ecdysone Protects Mouse Osteoblasts from Glucocorticoid-Induced Apoptosis In Vitro. Planta Med. 2017, 83, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.H.; Yue, Z.S.; Xin, D.W.; Zeng, L.R.; Xiong, Z.F.; Hu, Z.Q.; Xu, C.D. β-Ecdysterone promotes autophagy and inhibits apoptosis in osteoporotic rats. Mol. Med. Rep. 2018, 17, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Sheu, S.Y.; Ho, S.R.; Sun, J.S.; Chen, C.Y.; Ke, C.J. Arthropod steroid hormone (20-Hydroxyecdysone) suppresses IL-1β-induced catabolic gene expression in cartilage. BMC Complement. Altern. Med. 2015, 15, 1. [Google Scholar] [CrossRef]
- You, W.L.; Xu, Z.L. β-ecdysone promotes osteogenic differentiation of bone marrow mesenchymal stem cells. J. Gene Med. 2020, 22, e3207. [Google Scholar] [CrossRef]
- Han, J.; Guan, J.; Zhu, X. β-Ecdysone attenuates cartilage damage in a mouse model of collagenase-induced osteoarthritis via mediating FOXO1/ADAMTS-4/5 signaling axis. Histol. Histopathol. 2021, 36, 785–794. [Google Scholar]
- Yan, C.P.; Wang, X.K.; Jiang, K.; Yin, C.; Xiang, C.; Wang, Y.; Pu, C.; Chen, L.; Li, Y.L. β-Ecdysterone Enhanced Bone Regeneration Through the BMP-2/SMAD/RUNX2/Osterix Signaling Pathway. Front. Cell Dev. Biol. 2022, 10, 883228. [Google Scholar] [CrossRef]
- Jun, J.H.; Yoon, W.J.; Seo, S.B.; Woo, K.M.; Kim, G.S.; Ryoo, H.M.; Baek, J.H. BMP2-activated Erk/MAP kinase stabilizes Runx2 by increasing p300 levels and histone acetyltransferase activity. J. Biol. Chem. 2010, 285, 36410–36419. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.; Chen, Z.; Liu, Z.; Zhang, J.H.; Xi, G.; Feng, H. The effect of ecdysterone on cerebral vasospasm following experimental subarachnoid hemorrhage in vitro and in vivo. Neurol. Res. 2008, 30, 571–580. [Google Scholar] [CrossRef]
- Hu, J.; Zhao, T.Z.; Chu, W.H.; Luo, C.X.; Tang, W.H.; Yi, L.; Feng, H. Protective effects of 20-hydroxyecdysone on CoCl(2)-induced cell injury in PC12 cells. J. Cell Biochem. 2010, 111, 1512–1521. [Google Scholar] [CrossRef]
- Luo, C.; Yi, B.; Fan, W.; Chen, K.; Gui, L.; Chen, Z.; Li, L.; Feng, H.; Chi, L. Enhanced angiogenesis and astrocyte activation by ecdysterone treatment in a focal cerebral ischemia rat model. Acta Neurochir. Suppl. 2011, 110, 151–155. [Google Scholar] [PubMed]
- Hu, J.; Luo, C.X.; Chu, W.H.; Shan, Y.A.; Qian, Z.M.; Zhu, G.; Yu, Y.B.; Feng, H. 20-Hydroxyecdysone protects against oxidative stress-induced neuronal injury by scavenging free radicals and modulating NF-κB and JNK pathways. PLoS ONE 2012, 7, e50764. [Google Scholar] [CrossRef]
- Xia, X.; Zhang, Q.; Liu, R.; Wang, Z.; Tang, N.; Liu, F.; Huang, G.; Jiang, X.; Gui, G.; Wang, L.; et al. Effects of 20-hydroxyecdysone on improving memory deficits in streptozotocin-induced type 1 diabetes mellitus in rat. Eur. J. Pharmacol. 2014, 740, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Trillo, L.; Das, D.; Hsieh, W.; Medina, B.; Moghadam, S.; Lin, B.; Dang, V.; Sanchez, M.M.; De Miguel, Z.; Ashford, J.W.; et al. Ascending monoaminergic systems alterations in Alzheimer’s disease. translating basic science into clinical care. Neurosci. Biobehav. Rev. 2013, 37, 1363–1379. [Google Scholar] [CrossRef]
- Lim, H.S.; Moon, B.C.; Lee, J.; Choi, G.; Park, G. The insect molting hormone 20-hydroxyecdysone protects dopaminergic neurons against MPTP-induced neurotoxicity in a mouse model of Parkinson’s disease. Free Radic. Biol. Med. 2020, 159, 23–36. [Google Scholar] [CrossRef]
- Zou, Y.; Wang, R.; Guo, H.; Dong, M. Phytoestrogen β-Ecdysterone Protects PC12 Cells Against MPP+-Induced Neurotoxicity In Vitro: Involvement of PI3K-Nrf2-Regulated Pathway. Toxicol. Sci. 2015, 147, 28–38. [Google Scholar] [CrossRef]
- Wu, J.; Gao, L.; Shang, L.; Wang, G.; Wei, N.; Chu, T.; Chen, S.; Zhang, Y.; Huang, J.; Wang, J.; et al. Ecdysterones from Rhaponticum carthamoides (Willd.) Iljin reduce hippocampal excitotoxic cell loss and upregulate mTOR signaling in rats. Fitoterapia 2017, 119, 158–167. [Google Scholar] [CrossRef]
- Franco, R.R.; Takata, L.d.A.; Chagas, K.; Justino, A.B.; Saraiva, A.L.; Goulart, L.R.; Ávila, V.d.M.R.; Otoni, W.C.; Espindola, F.S.; da Silva, C.R. A 20-hydroxyecdysone-enriched fraction from Pfaffia glomerata (Spreng.) pedersen roots alleviates stress, anxiety, and depression in mice. J. Ethnopharmacol. 2021, 267, 113599. [Google Scholar] [CrossRef] [PubMed]
- Gholipour, P.; Komaki, A.; Ramezani, M.; Parsa, H. Effects of the combination of high-intensity interval training and Ecdysterone on learning and memory abilities, antioxidant enzyme activities, and neuronal population in an Amyloid-beta-induced rat model of Alzheimer’s disease. Physiol. Behav. 2022, 251, 113817. [Google Scholar] [CrossRef]
- Todorova, V.; Ivanova, S.; Yotov, V.; Zaytseva, E.; Ardasheva, R.; Turiyski, V.; Prissadova, N.; Ivanov, K. Phytoecdysteroids: Quantification in Selected Plant Species and Evaluation of Some Effects on Gastric Smooth Muscles. Molecules 2024, 29, 5145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xu, X.; Xu, T.; Qin, S. β-Ecdysterone suppresses interleukin-1β-induced apoptosis and inflammation in rat chondrocytes via inhibition of NF-κB signaling pathway. Drug Dev. Res. 2014, 75, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Posey, K.L. Curcumin and Resveratrol: Nutraceuticals with so Much Potential for Pseudoachondroplasia and Other ER-Stress Conditions. Biomolecules 2024, 14, 154. [Google Scholar] [CrossRef]
- Nassar, K.; El-Mekawey, D.; Elmasry, A.E.; Refaey, M.S.; El-Sayed Ghoneim, M.; Elshaier, Y.A.M.M. The significance of caloric restriction mimetics as anti-aging drugs. Biochem. Biophys. Res. Commun. 2024, 692, 149354. [Google Scholar] [CrossRef]
- Pan, Z.; Niu, Y.; Liang, Y.; Zhang, X.; Dong, M. β-Ecdysterone Protects SH-SY5Y Cells Against 6-Hydroxydopamine-Induced Apoptosis via Mitochondria-Dependent Mechanism: Involvement of p38(MAPK)-p53 Signaling Pathway. Neurotox. Res. 2016, 30, 453–466. [Google Scholar] [CrossRef]
- Sun, Y.; Zhao, D.L.; Liu, Z.X.; Sun, X.H.; Li, Y. Beneficial effect of 20-hydroxyecdysone exerted by modulating antioxidants and inflammatory cytokine levels in collagen-induced arthritis: A model for rheumatoid arthritis. Mol. Med. Rep. 2017, 16, 6162–6169. [Google Scholar] [CrossRef]
- Yang, L.; Hui, A.; Jian, P. Ecdysterone Accelerates Healing of Radiation-Induced Oral Mucositis in Rats by Increasing Matrix Cell Proliferation. Radiat. Res. 2019, 191, 237–244. [Google Scholar]
- Yang, L.; Pan, J. Therapeutic Effect of Ecdysterone Combine Paeonol Oral Cavity Direct Administered on Radiation-Induced Oral Mucositis in Rats. Int. J. Mol. Sci. 2019, 20, 3800. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Friedemann, T.; Pan, J. Ecdysterone Attenuates the Development of Radiation-Induced Oral Mucositis in Rats at Early Stage. Radiat. Res. 2021, 196, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Romaniuk-Drapała, A.; Lisiak, N.; Totoń, E.; Matysiak, A.; Nawrot, J.; Nowak, G.; Kaczmarek, M.; Rybczyńska, M.; Rubiś, B. Proapoptotic and proautophagic activity of 20-hydroxyecdysone in breast cancer cells in vitro. Chem. Biol. Interact. 2021, 342, 109479. [Google Scholar] [CrossRef]
- Kizelsztein, P.; Govorko, D.; Komarnytsky, S.; Evans, A.; Wang, Z.; Cefalu, W.T.; Raskin, I. 20-Hydroxyecdysone decreases weight and hyperglycemia in a diet-induced obesity mice model. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E433–E439. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, R.; Naresh, R.; Shanthi, P.; Sachdanandam, P. Efficacy of 20-OH-ecdysone on hepatic key enzymes of carbohydrate metabolism in streptozotocin induced diabetic rats. Phytomedicine 2012, 19, 725–729. [Google Scholar] [CrossRef]
- Naresh Kumar, R.; Sundaram, R.; Shanthi, P.; Sachdanandam, P. Protective role of 20-OH ecdysone on lipid profile and tissue fatty acid changes in streptozotocin induced diabetic rats. Eur. J. Pharmacol. 2013, 698, 489–498. [Google Scholar] [CrossRef]
- Semiane, N.; Foufelle, F.; Ferré, P.; Hainault, I.; Ameddah, S.; Mallek, A.; Khalkhal, A.; Dahmani, Y. High carbohydrate diet induces nonalcoholic steato-hepatitis (NASH) in a desert gerbil. Comptes Rendus Biol. 2017, 340, 25–36. [Google Scholar] [CrossRef]
- Mallek, A.; Movassat, J.; Ameddah, S.; Liu, J.; Semiane, N.; Khalkhal, A.; Dahmani, Y. Experimental diabetes induced by streptozotocin in the desert gerbil, Gerbillus gerbillus, and the effects of short-term 20-hydroxyecdysone administration. Biomed. Pharmacother. 2018, 102, 354–361. [Google Scholar] [CrossRef]
- Agoun, H.; Semiane, N.; Mallek, A.; Bellahreche, Z.; Hammadi, S.; Madjerab, M.; Abdlalli, M.; Khalkhala, A.; Dahmani, Y. High-carbohydrate diet-induced metabolic disorders in Gerbillus tarabuli (a new model of non-alcoholic fatty-liver disease). Protective effects of 20-hydroxyecdysone. Arch. Physiol. Biochem. 2021, 127, 127–135. [Google Scholar] [CrossRef]
- Bellahreche, Z.; Dahmani, Y. 20-Hydroxyecdysone bioavailability and pharmacokinetics in Gerbillus tarabuli, a gerbil model for metabolic syndrome studies. Steroids 2023, 198, 109262. [Google Scholar] [CrossRef]
- Foucault, A.; Mathé, V.; Lafont, R.; Even, P.; Dioh, W.; Veillet, S.; Tomé, D.; Huneau, J.; Hermier, D.; Quignard-Boulangé, A. Quinoa extract enriched in 20-hydroxyecdysone protects mice from diet-induced obesity and modulates adipokines expression. Obesity 2012, 20, 270–277. [Google Scholar] [CrossRef]
- Graf, B.L.; Poulev, A.; Kuhn, P.; Grace, M.H.; Lila, M.A.; Raskin, I. Quinoa seeds leach phytoecdysteroids and other compounds with anti-diabetic properties. Food Chem. 2014, 163, 178–185. [Google Scholar] [CrossRef]
- Foucault, A.-S.; Even, P.; Lafont, R.; Dioh, W.; Veillet, S.; Tomé, D.; Huneau, J.-F.; Hermier, D.; Quignard-Boulangé, A. Quinoa extract enriched in 20-hydroxyecdysone affects energy homeostasis and intestinal fat absorption in mice fed a high-fat diet. Physiol. Behav. 2014, 128, 226–231. [Google Scholar] [CrossRef]
- Merenda, T.; Juszczak, F.; Ferier, E.; Duez, P.; Patris, S.; Declèves, A.É.; Nachtergael, A. Natural compounds proposed for the management of non-alcoholic fatty liver disease. Nat. Prod. Bioprospect. 2024, 14, 24. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Jin, H.; Zheng, S.L.; Xia, P.; Cai, Y.; Ni, X.J. Phytoecdysteroids from Ajuga iva act as potential antidiabetic agent against alloxan-induced diabetic male albino rats. Biomed. Pharmacother. 2017, 96, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.J.; Madrigal-Matute, J.; Scheibye-Knudsen, M.; Fang, E.; Aon, M.; González-Reyes, J.A.; Cortassa, S.; Kaushik, S.; Gonzalez-Freire, M.; Patel, B.; et al. Effects of sex, strain, and energy intake on hallmarks of aging in mice. Cell Metab. 2016, 23, 1093–1112. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Min, K.J. Caloric restriction and its mimetics. BMB Rep. 2013, 46, 181–187. [Google Scholar] [CrossRef]
- Mattison, J.A.; Colman, R.J.; Beasley, T.M.; Allison, D.B.; Kemnitz, J.W.; Roth, G.S.; Ingram, D.K.; Weindruch, R.; de Cabo, R.; Anderson, R.M. Caloric restriction improves health and survival of rhesus monkeys. Nat. Commun. 2017, 8, 14063. [Google Scholar] [CrossRef]
- Most, J.; Tosti, V.; Redman, L.M.; Fontana, L. Calorie restriction in humans: An update. Ageing Res. Rev. 2017, 39, 36–45. [Google Scholar] [CrossRef]
- Ravussin, E.; Redman, L.M.; Rochon, J.; Das, S.K.; Fontana, L.; Kraus, W.E.; Romashkan, S.; Williamson, D.A.; Meydani, S.N.; Villareal, D.T.; et al. A 2-year randomized controlled trial of human caloric restriction: Feasibility and effects on predictors of health span and longevity. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2015, 70, 1097–1104. [Google Scholar] [CrossRef]
- Das, S.K.; Balasubramanian, P.; Weerasekara, Y.K. Nutrition modulation of human aging: The calorie restriction paradigm. Mol. Cell. Endocrinol. 2017, 455, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; Silver, R.E.; Senior, A.; Gilhooly, C.H.; Bhapkar, M.; Le Couteur, D. Diet composition, adherence to calorie restriction, and cardiometabolic disease risk modification. Aging Cell 2023, 22, e14018. [Google Scholar] [CrossRef]
- Waziry, R.; Ryan, C.P.; Corcoran, D.L.; Huffman, K.M.; Kobor, M.S.; Kothari, M.; Graf, G.H.; Kraus, V.B.; Kraus, W.E.; Lin, D.T.S.; et al. Effect of long-term caloric restriction on DNA methylation measures of biological aging in healthy adults from the CALERIE trial. Nat. Aging 2023, 3, 248–257, Erratum in Nat. Aging 2023, 3, 753. [Google Scholar] [CrossRef]
- Hastings, W.J.; Ye, Q.; Wolf, S.E.; Ryan, C.P.; Das, S.K.; Huffman, K.M.; Kobor, M.S.; Kraus, W.E.; MacIsaac, J.L.; Martin, C.K.; et al. Effect of long-term caloric restriction on telomere length in healthy adults: CALERIE™ 2 trial analysis. Aging Cell 2024, 23, e14149. [Google Scholar] [CrossRef] [PubMed]
- Rubin, R. Cut Calories, Lengthen Life Span? Randomized Trial Uncovers Evidence That Calorie Restriction Might Slow Aging, but Questions Remain. JAMA 2023, 329, 1049–1050. [Google Scholar] [CrossRef]
- Ekmekcioglu, C. Nutrition and longevity—From mechanisms to uncertainties. Crit. Rev. Food Sci. Nutr. 2020, 60, 3063–3082. [Google Scholar] [CrossRef]
- Hofer, S.J.; Davinelli, S.; Bergmann, M.; Scapagnini, G.; Madeo, F. Caloric Restriction Mimetics in Nutrition and Clinical Trials. Front. Nutr. 2021, 8, 717343. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, I.; Hollenberg, M.D.; Ding, H.; Triggle, C.R. A Critical Review of the Evidence That Metformin Is a Putative Anti-Aging Drug That Enhances Healthspan and Extends Lifespan. Front. Endocrinol. 2021, 12, 718942. [Google Scholar] [CrossRef]
- Buniam, J.; Chansela, P.; Weerachayaphorn, J.; Saengsirisuwan, V. Dietary Supplementation with 20-Hydroxyecdysone Ameliorates Hepatic Steatosis and Reduces White Adipose Tissue Mass in Ovariectomized Rats Fed a High-Fat, High-Fructose Diet. Biomedicines 2023, 11, 2071. [Google Scholar] [CrossRef]
- Guru, B.; Tamrakar, A.K.; Manjula, S.N.; Prashantha Kumar, B.R. Novel dual PPARα/γ agonists protect against liver steatosis and improve insulin sensitivity while avoiding side effects. Eur. J. Pharmacol. 2022, 935, 175322. [Google Scholar] [CrossRef]
- Hu, J.; Feng, Y.; Li, B.; Wang, F.; Qian, Q.; Tian, W.; Niu, I.; Wang, X. Identification of quality markers for Cyanotis arachnoidea and analysis of its physiological mechanism based on chemical pattern recognition, network pharmacology, and experimental validation. Peer J. 2023, 11, e15948. [Google Scholar] [CrossRef] [PubMed]
- Mamadalieva, N.Z.; Hussain, H.; Mollica, A.; Zengin, G.; Mamadalieva, R.Z.; Elhady, S.S.; Fadil, S.A.; Ashour, M.L.; Youssef, F.S. Ecdysteroids as potent enzyme inhibitors and verification of their activity using in vitro and in silico docking studies. Life 2022, 12, 824. [Google Scholar] [CrossRef]
- Liu, Q.; Zhuang, S.; Li, S.; Nong, Y.; Zhang, Y.; Liang, J.; Zhang, Y.; Liu, C. Rapid screening, isolation, and activity evaluation of potential xanthine oxidase inhibitors in Polyporus umbellatus and mechanism of action in the treatment of gout. Phytochem. Anal. 2023, 35, 116–134. [Google Scholar] [CrossRef] [PubMed]
- Nambaaru, S.; Sahu, P.N.; Katta, S.; Ganapavarapu, V.R.S.; Kancharla, B.; Sen, A. Phytochemical analysis and anti-inflammatory potential of Floscopa splendes Lour. and exploring ecdysterone as promising therapeutic agent: An HR-LCMS screening and molecular docking analysis. ChemistrySelect 2024, 9, e202403146. [Google Scholar] [CrossRef]
- Shuvalov, O.; Kirdeeva, Y.; Fefilova, E.; Daks, A.; Fedorova, O.; Parfenyev, S.; Nazarov, A.; Vlasova, Y.; Krasnov, G.S.; Barlev, N.A. 20-Hydroxyecdysone Boosts Energy Production and Biosynthetic Processes in Non-Transformed Mouse Cells. Antioxidants 2024, 13, 1349. [Google Scholar] [CrossRef]
- Leogrande, P.; Jardines, D.; Brito, D.M.; de la Torre, X.; Botrè, F.; Luch, A.; Diel, P.; Parr, M.K. Investigation of the Physiological and Post-training Effects of Ecdysteroid Supplementation by Multivariate Analysis of the Human Serum Metabolome. J. Chemom. 2024, 38, e3594. [Google Scholar] [CrossRef]
- Das, S.; Morvan, F.; Jourde, B.; Meier, V.; Kahle, P.; Brebbia, P.; Toussaint, G.; Glass, D.J.; Fornaro, M. ATP citrate lyase improves mitochondrial function in skeletal muscle. Cell Metab. 2015, 21, 868–876. [Google Scholar] [CrossRef]
- Das, S.; Morvan, F.; Morozzi, G.; Jourde, B.; Minetti, G.C.; Kahle, P.; Rivet, H.; Brebbia, P.; Toussaint, G.; Glass, D.J.; et al. ATP Citrate Lyase Regulates Myofibre Differentiation and Increases Regeneration by Altering Histone Acetylation. Cell Rep. 2017, 21, 3003–3011. [Google Scholar] [CrossRef]
- Espino-Gonzalez, E.; Tickle, P.G.; Altara, R.; Gallagher, H.; Cheng, C.W.; Engman, V.; Wood, N.; da Silva, G.J.J.; Scalabrin, M.; Yu, X.; et al. Caloric restriction rejuvenates skeletal muscle growth in heart failure with preserved ejection fraction. J. Am. Coll. Cardiol. Basic Trans. Sci. 2024, 9, 223–240. [Google Scholar] [CrossRef]
- Zádor, E. Paradoxical Effect of Caloric Restriction on Overload-Induced Muscle Growth in Diastolic Heart Failure. J. Am. Coll. Cardiol. Basic Trans. Sci. 2024, 9, 241–243. [Google Scholar] [CrossRef]
- Ryu, K.W.; Fung, T.S.; Baker, D.C.; Saoi, M.; Park, J.; Febres-Aldana, C.A.; Aly, R.G.; Cui, R.; Sharma, A.; Fu, Y.; et al. Cellular ATP demand creates metabolically distinct subpopulations of mitochondria. Nature 2024, 635, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Tarkowská, D.; Strnad, M. Plant ecdysteroids: Plant sterols with intriguing distributions, biological effects and relations to plant hormones. Planta 2016, 244, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Popova, E.; Titova, M.; Tynykulov, M.; Zakirova, R.P.; Kulichenko, I.; Prudnikova, O.; Nosov, A. Sustainable Production of Ajuga Bioactive Metabolites Using Cell Culture Technologies: A Review. Nutrients 2023, 15, 1246. [Google Scholar] [CrossRef]
- Shuvalov, O.; Kirdeeva, Y.; Fefilova, E.; Netsvetay, S.; Zorin, M.; Vlasova, Y.; Fedorova, O.; Daks, A.; Parfenyev, S.; Barlev, N. 20-Hydroxyecdysone Confers Antioxidant and Antineoplastic Properties in Human Non-Small Cell Lung Cancer Cells. Metabolites 2023, 13, 656. [Google Scholar] [CrossRef]
- Baev, A.Y.; Charishnikova, O.S.; Khasanov, F.A.; Nebesnaya, K.S.; Makhmudov, A.R.; Rakhmedova, M.T.; Khushbaktova, Z.A.; Syrov, V.N.; Levitskaya, Y.V. Ecdysterone prevents negative effect of acute immobilization stress on energy metabolism of rat liver mitochondria. J. Steroid Biochem. Mol. Biol. 2022, 219, 106066. [Google Scholar] [CrossRef] [PubMed]
- Machado, S.A.; Pasquarelli-Do-Nascimento, G.; da Silva, D.S.; Farias, G.R.; Santos, I.d.O.; Baptista, L.B.; Magalhães, K.G. Browning of the white adipose tissue regulation: New insights into nutritional and metabolic relevance in health and diseases. Nutr. Metab. 2022, 19, 61. [Google Scholar] [CrossRef]
- Bellahreche, Z.; Sihali-Beloui, O.; Semiane, N.; Mallek, A.; Dahmani, Y. 20-Hydroxyecdysone induces brown adipose tissue glycogen accumulation in obese gerbil model: Histochemical study. Ann. Endocrinol. 2023, 84, 670. [Google Scholar] [CrossRef]
- Neves, C.S.; Gomes, S.S.; dos Santos, T.R.; de Almeida, M.M.; de Souza, Y.O.; Garcia, R.M. The phytoecdysteroid β-ecdysone is genotoxic in Rodent Bone Marrow Micronuclei and Allium cepa L. Assays. J. Ethnopharmacol. 2016, 177, 81–84. [Google Scholar] [CrossRef]
- Russell, S.J.; Kahn, C.R. Endocrine regulation of ageing. Nat. Rev. Mol. Cell Biol. 2007, 8, 681–691. [Google Scholar] [CrossRef]
- Suh, Y.; Atzmon, G.; Cho, M.-O.; Hwang, D.; Liu, B.; Leahy, D.J.; Barzilai, N.; Cohen, P. Functionally significant insulin-like growth factor I receptor mutations in centenarians. Proc. Natl. Acad. Sci. USA 2008, 105, 3438–3442. [Google Scholar] [CrossRef]
- Giacomozzi, C.; Martin, A.; Fernández, M.C.; Gutiérrez, M.; Iascone, M.; Domené, H.M.; Dominici, F.P.; Bergadá, I.; Cangiano, B.; Persani, L.; et al. Novel Insulin-Like Growth Factor 1 Gene Mutation: Broadening of the Phenotype and Implications for Insulin Resistance. J. Clin. Endocrinol. Metab. 2023, 108, 1355–1369. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zádor, E. Molecular Targets of 20-Hydroxyecdysone in Mammals, Mechanism of Action: Is It a Calorie Restriction Mimetic and Anti-Aging Compound? Cells 2025, 14, 431. https://doi.org/10.3390/cells14060431
Zádor E. Molecular Targets of 20-Hydroxyecdysone in Mammals, Mechanism of Action: Is It a Calorie Restriction Mimetic and Anti-Aging Compound? Cells. 2025; 14(6):431. https://doi.org/10.3390/cells14060431
Chicago/Turabian StyleZádor, Ernő. 2025. "Molecular Targets of 20-Hydroxyecdysone in Mammals, Mechanism of Action: Is It a Calorie Restriction Mimetic and Anti-Aging Compound?" Cells 14, no. 6: 431. https://doi.org/10.3390/cells14060431
APA StyleZádor, E. (2025). Molecular Targets of 20-Hydroxyecdysone in Mammals, Mechanism of Action: Is It a Calorie Restriction Mimetic and Anti-Aging Compound? Cells, 14(6), 431. https://doi.org/10.3390/cells14060431