Abstract
Astrocytes play a pivotal role in shaping synaptic function and in learning, memory, and emotion. Recent studies show that perisynaptic astrocytic processes form structured interactions with pre- and postsynaptic elements, which extends synaptic diversity beyond neuron–neuron connections. Accumulating evidence indicates that astrocytic Ca2+ signaling, gliotransmission, and local translation modulate synaptic efficacy and contribute to the formation and stabilization of memory traces. It is therefore essential to define how astrocytic microdomains, multisynaptic leaflet domains, and network-level ensembles cooperate to regulate circuit computation across space and time. Advances in super-resolution and volumetric in vivo imaging and spatial transcriptomics now enable detailed, cell-type- and compartment-specific analysis of astrocyte–synapse interactions in vivo. In this review, we highlight these approaches and synthesize classical and emerging mechanisms by which astrocytes read neuronal activity, write to synapses, and coordinate network states. We also discuss theoretical frameworks such as neuron–astrocyte associative memory models that formalize astrocytic calcium states as distributed substrates for storage and control. This integrated view provides new insight into the multicellular logic of memory and suggests paths toward understanding and treating neurological and psychiatric disorders.
1. Introduction
Astrocytes are now recognized as active partners in synaptic function rather than passive support cells. Perisynaptic astrocytic processes actively sense neurotransmitter release, generate intracellular Ca2+ signals [1], and regulate the synaptic milieu through gliotransmission, ion buffering, glutamate uptake, and metabolic coupling [2,3,4,5]. Within the tripartite organization that includes the presynaptic terminal, the postsynaptic spine, and the surrounding astrocyte, astrocytes modulate classical forms of plasticity such as long-term potentiation (LTP), long-term depression (LTD), and spike-timing-dependent plasticity (STDP) [6,7,8], which expands plasticity beyond a neuron-only phenomenon to a neuron–glia interaction [9,10,11]. Recent advances in super-resolution microscopy [12], genetic manipulation, and computational analysis indicate strong spatial and temporal compartmentalization. Nanoscale calcium microdomains (also called “hotspots” or “calcium nanodomains”) are very small areas where calcium levels rise inside the thin perisynaptic processes. These localized calcium signals allow astrocytes to affect each synapse in a specific way. Interconnected leaflet networks (also called peri-synaptic astrocytic processes, PAPs), which are the smallest terminal branches of astrocytes, seem to combine or integrate activity coming from many different synapses [13,14,15,16]. At larger scales, ensembles of co-activated astrocytes have been shown to participate directly in memory encoding and recall, indicating that astrocytes themselves may form functional components of the memory engram [17].
It is therefore timely to reassess the long-standing neuron-centric view of learning and memory. Since Hebb’s postulate, most models and experiments have framed plasticity as changes in synaptic weights within neuronal networks [18,19,20,21,22]. This framework has been foundational. However, classical neuron-only models show clear limitations. Memory capacity tends to scale only linearly with network size, intrinsic homeostatic safeguards remain underspecified, and millisecond electrical rules are difficult to reconcile with slower biochemical and metabolic processes that support durable behavioral change [23,24,25,26,27,28]. The tripartite synapse concept, together with accumulating evidence for astrocyte-dependent modulation, introduces additional state variables, time constants, and control mechanisms that can address these gaps [1,2,3,4,5,6,7,8,9,29,30].
Theoretical work has begun to formalize astrocytes as computational elements. Emerging frameworks, exemplified by the NAAM model, propose that information can be stored not only in neuronal synaptic weights but also in dynamic Ca2+ states within astrocytic processes [31,32,33,34]. These states may enhance capacity, temporal stability, and context-dependent modulation. Although these predictions require further experimental testing, they offer a principled bridge between biological computation and artificial architectures and motivate targeted experiments on astrocyte–neuron cooperation [35,36].
In this review, we integrate classical and emerging perspectives on astrocyte-mediated synaptic plasticity and computation across scales. We first revisit the foundational mechanisms within the tripartite synapse and clarify their limitations. We then move from nanoscale Ca2+ microdomains and leaflet networks to mesoscale astrocyte ensembles that are implicated in learning and recall. By synthesizing experimental and theoretical insights, we highlight how astrocytes contribute to memory storage, network coordination, and computational capacity, fundamentally redefining how the brain learns and remembers (Figure 1 and Table 1).
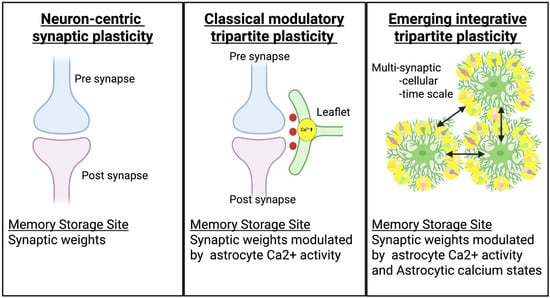
Figure 1.
Comparison of the key components and memory storage sites across three conceptual models of synaptic plasticity.

Table 1.
Comparison of three frameworks.
2. Literature Search Strategy and Inclusion Criteria
To guide a comprehensive and balanced narrative synthesis and to enhance the transparency of our literature scope, we conducted a structured search of studies published between January 1990 and October 2025. Searches were performed in PubMed, Scopus, and Web of Science using combinations of the terms “astrocyte”, “synaptic plasticity”, “calcium signaling”, “tripartite synapse”, “astrocyte ensemble”, and “learning and memory”. Additional studies were identified by examining the reference lists of key primary papers and recent reviews.
The focus of this narrative review is on mammalian central nervous system synapses, integrating mechanistic, physiological, and computational perspectives. We do not aim to provide exhaustive coverage of all glial synapses, nor do we systematically review peripheral tripartite synapses such as those at the neuromuscular junction, where glial modulation is also well established (reviewed in [37,38]).
In the neuron-centric model, plasticity arises from pre- and postsynaptic mechanisms such as NMDA receptor activation and AMPAR trafficking, with memory stored at individual neuronal synapses. In the classical tripartite model, plasticity involves coordinated activity among presynaptic terminals, postsynaptic neurons, and astrocytic leaflets, where astrocytic Ca2+ signaling and gliotransmission modulate synaptic strength and timing. In the emerging integrative model, astrocytes contribute to plasticity through multi-synaptic and cellular-scale calcium states that integrate network activity over time, making astrocyte microdomains an additional site for distributed memory encoding and storage within neural circuits.
3. Classical Core Mechanisms of Astrocyte-Mediated Synaptic Plasticity
Ben Barres’s pioneering studies established astrocytes as powerful drivers of synaptogenesis and synaptic efficacy [39], and on this foundation Araque et al. (1999) formalized the tripartite synapse [29], reframing plasticity to include astrocytes as active regulators rather than passive support cells. In this framework, perisynaptic astrocytes closely ensheath most excitatory synapses in the hippocampus [40] and cortex [41,42], enabling them to sense [30], integrate [30,43], and modulate [16,44] neuronal activity with remarkable precision [13]. Classical models propose that astrocytes may contribute to synaptic plasticity through several mechanisms, including (1) calcium-dependent release of neuroactive molecules, (2) extracellular homeostasis, and (3) metabolic support.
A central proposed mechanism is gliotransmission, the Ca2+-dependent release of neuroactive molecules from astrocytes [45,46]. Activation of metabotropic receptors such as mGluRs [47], GABA_B [48], and P2Y [49] triggers intracellular Ca2+ elevations, leading to the release of glutamate [50], ATP [51], D-serine [52], or GABA [53]. These gliotransmitters fine-tune synaptic efficacy: D-serine acts as an NMDA receptor co-agonist required for LTP induction [52,54], ATP-derived adenosine suppresses presynaptic release [55,56]. Astrocyte-derived glutamate or GABA [57] can, respectively, enhance or dampen neuronal excitability. Through these processes, astrocytes exert bidirectional control over synaptic strength and plasticity [58,59]. While these findings support a role for gliotransmission, the physiological relevance of gliotransmission in vivo remains actively debated [60,61]. Studies using IP3R2 knockout mice, where global astrocytic Ca2+ elevations are disrupted, show largely preserved LTP and complex behavior [62,63]. These observations raise questions about how frequently gliotransmission participates under physiological conditions, although they may also reflect compensatory Ca2+ signaling through alternative pathways such as IP3R1/3, store-operated calcium entry, or microdomain-restricted Ca2+ events that remain intact in IP3R2-deficient astrocytes [64]. Concerns have also been raised regarding the cellular origin of D-serine [65,66], and vesicular versus channel-mediated release pathways [67], indicating that gliotransmission is likely a context-dependent mechanism [64].
Astrocytes also maintain the extracellular milieu necessary for stable transmission. High-affinity glutamate transporters GLT-1 and GLAST rapidly clear glutamate from the cleft, limiting receptor overactivation and preventing excitotoxicity [68]. Inwardly rectifying potassium channels (Kir4.1) and extensive gap-junction coupling enable redistribution of K⁺ ions, preventing local hyperexcitability [69]. Moreover, astrocytes sustain neuronal metabolism via the astrocyte–neuron lactate shuttle, ensuring that energy supply matches the demands of activity-dependent plasticity [70,71]. Furthermore, astrocytes secrete matricellular proteins such as thrombospondins (TSP1/2), hevin (SPARCL1), and SPARC. Thrombospondins promote excitatory synapse formation [72], while hevin facilitates excitatory synaptogenesis and SPARC antagonizes this process by competing for shared binding partners [73,74]. In addition, central neuropeptides such as oxytocin, vasopressin, and CRH act as potent modulators of astrocytic calcium signaling, metabolism, and gene expression, linking astrocyte function to mood regulation and neurodegenerative disease mechanisms [75].
The induction and maintenance of classical forms of synaptic plasticity such as LTP, LTD, and STDP. The timing of gliotransmitter release can alter the polarity of spike-timing-dependent plasticity [6,8], while astrocytic p38α MAPK signaling has been shown to be essential for hippocampal LTD expression [76,77]. By regulating neurotransmitter availability, receptor occupancy, and ionic balance, astrocytes actively shape the rules of synaptic modification [30,45,78].
Nevertheless, the temporal mismatch between astrocytic Ca2+ elevations (seconds to minutes) and fast synaptic events (milliseconds) raises questions about how astrocytes interact with rapid forms of synaptic plasticity [79,80]. Similarly, synaptic specificity remains a major unresolved issue: broad astrocytic Ca2+ transients could impact many synapses simultaneously unless constrained by microdomain-level compartmentalization, which was not resolved by early imaging studies [81]. Moreover, the model treated astrocytes as homogeneous units, overlooking their regional diversity [82] and nanoscale structural specialization [83]. While classical mechanisms explain modulation of local synapses, they do not fully account for how astrocytes coordinate information across networks or sustain long-term plasticity over extended timescales [84,85].
These limitations have collectively prompted a conceptual expansion beyond the classical framework. Advances in high-resolution imaging, genetic manipulation, and modeling now reveal a hierarchically organized architecture of astrocytic signaling. Structures from nanoscale microdomains to network-level ensembles support integrative and dynamic roles in learning and memory, as discussed in the following sections.
4. Beyond the Classical Tripartite Model
Recent discoveries have expanded our understanding of astrocytes far beyond the boundaries of the classical tripartite synapse [78,86]. Rather than functioning as uniform modulators, astrocytes exhibit highly compartmentalized organization and operate across multiple spatial and temporal scales. At the nanoscale, fine perisynaptic processes form distinct Ca2+ microdomains that decode synapse-specific signaling [13,15,87]. At larger scales, astrocytic leaflets and process networks integrate activity across thousands of synapses, enabling context-dependent modulation of neuronal ensembles [14,83].
Astrocytes also display remarkable computational versatility. They can decode neuronal firing patterns [2,88], release gliotransmitters with temporal precision [67], and coordinate large-scale network oscillations [89,90]. Emerging evidence further indicates that subsets of astrocytes participate directly in memory encoding and recall, forming astrocytic ensembles in memory that influence behavioral output [17,91].
In the following sections, we examine how this hierarchical organization from microdomains to network-level assemblies [92] endows astrocytes with integrative and computational capabilities that extend well beyond the classical neuron-centric view of plasticity [93,94] (Figure 2 and Table 2).
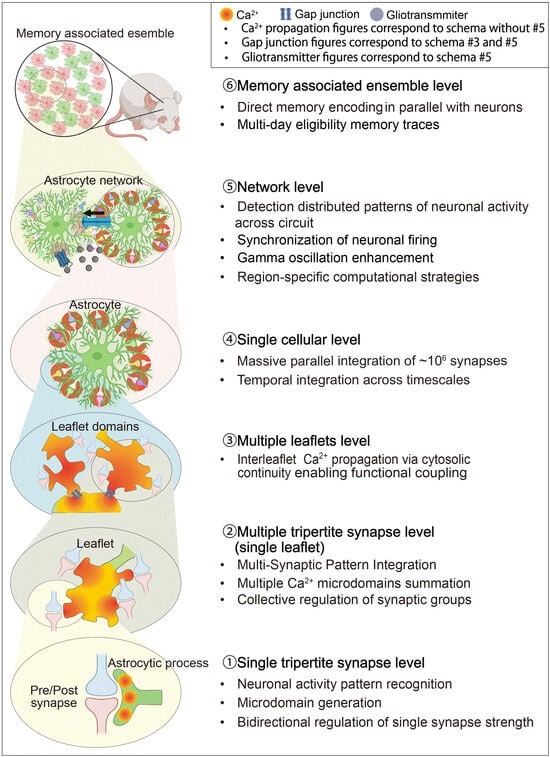
Figure 2.
Hierarchical Structure of the Astrocyte-Integrated Framework.
Table 2.
(A). Hierarchical Structure of the Astrocyte-Integrated Framework (MICRO-, MESO-, MACRO-SCALE). (B). Hierarchical Structure of the Astrocyte-Integrated Framework (NETWORK-, ENSEMBLE-SCALE).
Schematic illustration of the hierarchical organization of astrocyte signaling and integration across spatial and functional scales. At the single tripartite synapse level, individual tripartite synapses are modulated by localized Ca2+ transients and astrocytic read–write mechanisms. At the multi-synaptic level, a single astrocytic leaflet interacts with up to ten synapses, integrating local inputs into coherent microcircuit modulation. At the multi-leaflet level, neighboring leaflets form gap–junction-interconnected domains that coordinate calcium activity across a single astrocyte territory, thereby supporting domain-wide information processing. At the single-astrocyte level, one protoplasmic astrocyte contacts up to 106 synapses, integrating excitatory and inhibitory inputs through intracellular calcium signaling and metabolic coupling. At the network level, intercellular communication via Ca2+ waves through gap junction coupling, and gliotransmitter release synchronizes astrocytic and neuronal ensembles across brain regions. Finally, at the ensemble level, distributed astrocyte populations form functional networks that participate in learning, memory processing, and state-dependent coordination of neural circuits. Adapted from [14].
4.1. Microdomains and Leaflet Domains: Hierarchical Units of Astrocytic Integration
Recent studies revealed that astrocytic signaling is executed through a hierarchically organized nanoscale architecture that supports both local precision and mesoscale integration [80,83]. Within this framework, two complementary structural and functional units emerge: microdomains, which mediate Ca2+ signaling at single synapses [13], and leaflet domains, which integrate activity across multiple neighboring synapses [14]. Together, these levels of organization endow astrocytes with the capacity to decode, transform, and coordinate information within complex neuronal networks.
4.1.1. Microdomains: Synapse-Specific Read–Write Units
Super-resolution 3D-STED microscopy has shown that fine astrocytic processes form a spongiform meshwork composed of bulbous nodes connected by narrow shafts which are thin, elongated tubular structures. Each node, typically 300–400 nm in diameter, contacts dendritic spines and contains endoplasmic reticulum fragments enriched in IP3 receptors [13]. This node–shaft alternation imposes diffusion barriers, generating discrete biochemical compartments where Ca2+ transients remain spatially confined [87]. Within these microdomains, Ca2+ transients are triggered by synaptic glutamate spillover [118,119], ATP release, or neuromodulator action via metabotropic receptors [1]. These localized events are fast (hundreds of milliseconds to seconds) and asynchronous across the astrocyte, enabling the simultaneous monitoring of thousands of synapses with high spatial fidelity [15].
Astrocytes do not merely relay this synaptic information [111]. Covelo and Araque [2] demonstrated that single astrocytes can distinguish neuronal firing patterns and release distinct combinations of gliotransmitters. Short or low-frequency stimulation (~1 s or ≤20 Hz) elicits Ca2+-dependent glutamate release and transient potentiation, whereas high-frequency activity (30–90 s or >20 Hz) produces biphasic effects, with an initial potentiation followed by ATP/adenosine-mediated depression. Such pattern-specific responsiveness enables bidirectional regulation of synaptic strength [120], filtering transient versus sustained neuronal activity [45]. Microdomains thus perform synapse-specific read and write operations, converting electrical activity into biochemical modulation at the single-synapse level [96,121].
At the molecular level, astrocytic local translation represents an important molecular mechanism enabling calcium-dependent modulation of tripartite synapses. Recent studies using PAP-TRAP (peripheral astrocyte processes-translating ribosome affinity purification) and puromycin labeling [98] have revealed that astrocytes actively synthesize proteins within their perisynaptic processes, enabling rapid and spatially precise regulation of local proteomes. Ribosomes and mRNAs are abundant in these fine processes, and Ca2+ signals triggered by synaptic activity locally activate translation via mTOR [122] and eIF2α pathways [101]. This activity-dependent protein synthesis produces neurotransmitter transporters (GLT-1, GLAST), metabolic enzymes (Glul), and synapse-regulating factors (Thbs4, Hevin, SPARC, MERTK), allowing astrocytes to remodel individual microdomains in real time in response to neuronal activity. Recent evidence further demonstrates that astrocytic translation is essential for memory consolidation, as contextual fear conditioning modulates both the distribution and translation of ribosome-bound mRNAs within perisynaptic astrocytic processes [99]. Several PAP-enriched transcripts—such as Flt1, Fth1, Ccnd2, Mdm2, Gnb2l1, and Eef1a1—are directly linked to learning and memory [123,124,125], suggesting that local translation dynamically reconfigures perisynaptic proteomes to support long-term synaptic plasticity and memory stabilization.
4.1.2. Leaflet Domains: Structural Basis for Multi-Synaptic Integration
Beyond the single-synapse scale, astrocytic leaflet domains represent a newly recognized level of organization [107] that enables multi-synaptic integration. Recent volumetric electron microscopy and nanoscale calcium imaging [14] revealed that leaflets are ultra-thin lamellar extensions (<250 nm) emanating from distal processes. Quantitative reconstructions show that astrocytic leaflets ensheath approximately 90% of excitatory synapses in clusters and only ~10% individually, indicating that astrocytes regulate groups of synapses collectively rather than in isolation. Each leaflet contains minute endoplasmic reticulum (ER) saccules expressing IP3R1 [97,126], but notably lacks mitochondria, suggesting specialization for rapid, transient calcium signaling. Crucially, leaflets are interconnected via gap junction connexin43 [14], creating cytosolically continuous domains through which calcium and other signaling molecules propagate [104,127]. This organization allows multiple leaflets to function as integrated computational units capable of coordinating responses across clusters of synapses. In these domains, signals from several presynaptic neurons converge, permitting astrocytes to act as spatiotemporal integrators of network activity rather than passive modulators of single synapses.
Calcium signaling within leaflet domains follows threshold-dependent dynamics [92].Physiological increases in intracellular Ca2+ (~80–140 nM) are sufficient to trigger localized glutamate release [112], whereas larger or sustained elevations recruit additional gliotransmitter pathways such as ATP and D-serine release [128]. These graded responses convert variations in input strength and duration into specific biochemical outputs, enabling astrocytes to filter or amplify neuronal activity in a context-dependent manner. When multiple neighboring synapses are co-activated, calcium transients originating in individual leaflets can merge into prolonged, high-amplitude waves, reflecting an emergent property of cooperative integration within the leaflet network.
Functionally, leaflet domains serve as biochemical control hubs that bridge microdomain precision with mesoscale coordination. By pooling inputs from roughly ten or more presynaptic neurons, they stabilize excitation–inhibition balance [129] while preserving synapse-specific modulation. The architecture of leaflet domains thus provides a biophysical substrate for analog summation and threshold gating, analogous to nonlinear operations in computational systems [130]. This mechanism may underlie higher-order processes such as memory formation, decision-making, and contextual learning [85,116], in which selective activation and suppression of synaptic ensembles are required for adaptive behavior. Collectively, the integration of microdomain-level decoding and leaflet-level computation supports a unified view of astrocytes as hierarchical processors. Through localized calcium events and domain-level coupling, astrocytes transform distributed neuronal activity into coherent network-level modulation, linking microscopic signaling with macroscopic cognition. To provide quantitative context for these hierarchical structures, we summarize characteristic dimensions, calcium signal kinetics, and synaptic coverage ratios across microdomains, leaflets, single-cell territories, and network scales in Table 3. This table integrates values reported across super-resolution imaging, in vivo calcium imaging, and anatomical reconstruction studies.
Table 3.
Quantitative summary of astrocytic structural units and calcium dynamics.
4.2. Astrocytes as Network Coordinators
Beyond their nanoscale precision, astrocytes operate as large-scale integrators that coordinate neuronal activity across thousands of synapses [133]. Each protoplasmic astrocyte in the cortex or hippocampus occupies a distinct territorial domain encompassing tens to hundreds of thousands of synapses [108]. This organization enables astrocytes to detect distributed patterns of neuronal activity and transform them into coherent modulatory outputs, positioning them as essential computational hubs within neural circuits [134]. Astrocytes also exhibit regional and structural diversity—cortical astrocytes emphasize glutamatergic modulation [83,102], whereas white-matter astrocytes specialize in metabolic and axonal support [110,135]. Moreover, recent studies reveal that astrocytes regulate circuits not only through classical secreted factors but also via contact-mediated adhesion mechanisms [136,137], providing spatial precision and dynamic control of synaptic organization.
4.2.1. Network-Level Coordination
Astrocytic calcium signaling extends beyond local synaptic modulation to orchestrate large-scale network states. In vivo two-photon imaging demonstrates that astrocytic Ca2+ transients can synchronize previously asynchronous neuronal populations, generating coherent oscillations essential for attention, sensory binding, and memory consolidation [109,138]. Manipulation of astrocytic calcium activity alone is sufficient to induce transitions between synchronous and asynchronous cortical states [90,109], highlighting their causal role in governing network-level excitability and rhythmic organization.
Astrocytes also play a crucial role in maintaining the excitatory–inhibitory balance necessary for stable computation. Through temporally and spatially precise release of glutamate, ATP, and D-serine, they stabilize recurrent network activity and prevent runaway excitation [45,129]. Both experimental and computational studies indicate that astrocyte-modulated tripartite synapses can regulate oscillatory coherence [89], especially within recurrent feedback loops that sustain rhythmic network activity [30,139]. In this way, astrocytes function as adaptive homeostatic controllers [140,141] that balancing excitability and inhibition to maintain circuit stability while permitting experience-dependent plasticity.
4.2.2. Temporal Integration and Metaplasticity
Astrocytes supply a slow and integrative timescale that enriches neural computation. Their calcium waves can summate inputs over seconds to minutes, bridging transient synaptic activity with long-term modifications in circuit function [142]. This capacity allows astrocytes to convert rapid neuronal signals into persistent modulatory states that contribute to metaplasticity, the plasticity of plasticity itself [32,106]. Astrocyte-mediated gliotransmission regulates oscillatory coupling and temporal precision across cortical and subcortical circuits. Astrocytic glutamate release enhances gamma oscillation power (30–50 Hz) during active cognitive states [89], while ATP- and adenosine-dependent modulation of inhibitory interneurons improves oscillatory stability and synchrony [143,144,145]. These findings reveal that astrocytes act as temporal integrators, synchronizing neuronal ensembles across multiple timescales and linking fast synaptic signaling to slower, modulatory rhythms that underlie cognition and behavioral adaptation.
4.2.3. Region-Specific Coordination and Adaptive Plasticity
Astrocyte-mediated network coordination is further shaped by regional diversity in molecular signatures and circuit connectivity. Transcriptomic analyses show that astrocytes express distinct gene programs depending on their brain region, developmental origin, and local neuronal partners [146,147,148]. This regional specialization enables astrocytes to differentially regulate excitatory and inhibitory transmission [135,149], neuromodulator sensitivity [150], and oscillatory dynamics across cortical, hippocampal, and subcortical networks [89,109,146].Functionally, this diversity allows astrocytes to implement region-specific computational strategies. For example, astrocytes in the hippocampus contribute to theta–gamma coupling crucial for memory encoding [113], while those in the prefrontal cortex modulate sustained firing patterns associated with working memory and decision-making [151]. White matter astrocytes, in contrast, emphasize metabolic homeostasis and long-range axonal conduction rather than rapid synaptic modulation [110]. Under pathological conditions, this heterogeneity becomes more pronounced: Serrano-Pozo et al. [152] demonstrated region- and stage-specific transcriptomic remodeling of astrocytes in Alzheimer’s disease, while Clayton et al. [153] identified injury- and disease-specific reactive subtypes that influence circuit recovery and degeneration. Recognizing such diversity reframes astrocyte-mediated network control not as a uniform process but as a regionally tuned and functionally adaptive system. Through this multiscale organization, from local microdomain signaling to domain wide integration and global network synchronization, astrocytes enable the brain to maintain stability, flexibility, and computational depth across diverse behavioral contexts.
4.2.4. Adhesion-Based Mechanisms for Astrocytic Control of Neural Circuits
Astrocytes not only modulate neural activity through diffusible factors such as thrombospondins [72] and hevin [73] but also engage in direct contact-dependent adhesion mechanisms that shape synaptic connectivity and circuit function. These adhesion-based interactions represent an emerging paradigm [154] in which astrocytes use cell adhesion molecules (CAMs) to establish precise physical interfaces with neurons [136], enabling spatially restricted, bidirectional signaling [74] essential for synapse formation, maintenance, and plasticity [155].
Astrocytic neuroligins (NL1–3) and neurexins form trans-synaptic complexes that synchronize astrocyte morphological maturation with excitatory and inhibitory synaptogenesis during postnatal development [136]. NrCAM, acting through homophilic adhesion with neuronal NrCAM and coupling to the postsynaptic scaffold gephyrin, organizes GABAergic synapses and constrains astrocyte process overgrowth into the neuropil [156,157]. Integrin-based focal adhesion complexes (β-integrin, Talin, FAK, Tensin) further regulate astrocyte coverage of synapses and glutamate transporter expression, maintaining synaptic homeostasis under dynamic network conditions [158]. N-cadherin/δ-catenin complexes coordinate layer-specific astrocyte morphogenesis and neuronal dendrite stabilization [159] while Eph-ephrin signaling governs synapse remodeling, elimination, and astrocyte reactivity in development and disease [160,161,162]. Moreover, astrocytic phagocytic receptors MEGF10 and MERTK mediate selective synapse pruning, providing circuit-specific refinement of excitatory and inhibitory connectivity [163,164]. Collectively, these adhesion pathways operate in two modes. In a cooperative mode, where astrocyte–neuron adhesions promote synaptic maturation and stability, and in a competitive mode, where molecules like NrCAM function as “stop signals” to prevent excessive astrocytic infiltration into synaptic territories [156,157]. Dysregulation of these mechanisms can lead to excitatory–inhibitory imbalance and contribute to neurodevelopmental and psychiatric disorders such as autism and schizophrenia [136,165,166]. Thus, adhesion-based signaling establishes astrocytes as structural and molecular architects of neural circuits—integrating physical adhesion with biochemical communication to coordinate circuit assembly, balance, and long-term plasticity.
4.3. Astrocyte Ensembles in Memory Regulate Memory Processing
An emerging paradigm reshaping the neuroscience of memory is that astrocytes form functional ensembles that regulate the storage and recall of memories. These astrocyte ensembles in memory consist of spatially clustered astrocytes co-activated during memory processing [17,91]. Their coordinated activation suggests that astrocytes may contribute directly to the formation, maintenance, and retrieval of memory traces—beyond a purely supportive or modulatory role.
4.3.1. Early Evidence for Astrocytic Contributions to Memory
Initial studies demonstrated that activating astrocytes during learning enhances memory formation. Chemogenetic stimulation of hippocampal astrocytes during fear conditioning using Gq DREADDs strengthened synaptic potentiation and improved long-term memory [116]. Astrocyte-neuron lactate transport was shown to be required for long-term memory consolidation [167]. Subsequent calcium imaging studies revealed task-specific astrocytic activation patterns during learning and recall across multiple regions, including the hippocampus [116], amygdala [168], and prefrontal cortex [15,16]. Together, these findings established that astrocytes dynamically participate in memory-related network activity.
4.3.2. Astrocytic Ensembles as Memory Engram Components
A landmark study by Williamson, Kwon, and colleagues [17] provided direct evidence that astrocytes form engram-like ensembles essential for memory storage and recall. Learning experiences induced c-Fos expression in approximately 2–4% of hippocampal astrocytes, which were selectively reactivated during recall. These memory-related astrocytic ensembles, termed learning-associated astrocyte ensembles (LAAs), were spatially clustered near engram neurons, exhibited coordinated calcium dynamics, and enhanced local synaptic connectivity. Functional manipulations confirmed their causal role. Chemogenetic activation of astrocytic ensembles in memory, using a c-fos promoter–CreER–Lox-Stop-Lox intersectional genetic system, reactivated associated neuronal engrams and induced context-independent memory recall, whereas astrocyte-specific Fos deletion disrupted learning and recall. Transcriptomic analyses identified NFIA as a key transcriptional regulator of ensemble formation, with its loss impairing astrocyte-driven modulation of neuronal excitability. Recently, AstroLight technology, a calcium-dependent and light-activated gene expression system labeled with fluorescent markers, enables selective identification and manipulation of behaviorally relevant astrocytes in the nucleus accumbens [115]. Reactivation of these astrocytic ensembles drove reward-seeking behavior even in the absence of external cues. According to Dewa et al. [91], astrocytic ensembles stabilize long-term memory through sequential priming and activation. Learning induces adrenergic receptor upregulation that primes the ensembles. Subsequent recall drives cAMP and Ca2+ signaling and IGFBP2 release, which strengthens local engrams. These observations indicate that astrocytic ensembles may serve as multi-day eligibility traces for long-term stabilization. Collectively, these discoveries redefine the concept of the memory engram as a multicellular phenomenon. Astrocytic ensembles encode memory-relevant information in parallel with neurons, coordinate local plasticity, and regulate recall through calcium-dependent and transcriptional mechanisms. This neuron–astrocyte partnership represents a fundamental shift toward an integrated model of memory formation and stabilization in the mammalian brain.
4.4. Astrocytic Computational Frameworks and the Neuron–Astrocyte Associative Memory (NAAM) Model
Recent experimental discoveries have decisively shifted the perception of astrocytes from passive support cells to active computational partners in memory processing [78,117,169]. In parallel, theoretical neuroscience has begun to formalize this idea, positioning astrocytes as core elements within hybrid neuron–glia computational frameworks. Early conceptual models proposed that astrocytes could transiently store or integrate information through ionic or molecular states. Caudle [170] suggested that astrocytic ion-channel configurations might serve as a substrate for “glial memory,” while the “Memory Orchestra” framework [117,171] posited that astrocytes encode contextual aspects of experience such as spatial and temporal context through lasting biochemical states. Experimental support for these ideas came from Williamson et al. [17], who demonstrated that learning-associated astrocyte ensembles are reactivated during memory recall, providing direct evidence that astrocytic populations may participate in mnemonic encoding. Together, these findings inspired a new generation of computational models that extend beyond neuron-centric paradigms.
Building on these discoveries, the Neuron–Astrocyte Associative Memory (NAAM) model proposed by Kozachkov, Slotine, and Krotov [93] provides the first rigorous theoretical framework describing astrocytic computation. NAAM posits that memories are encoded not only in neuronal synaptic weights but also in the dynamic calcium signaling states between astrocytic processes. The state of each process is defined by its bidirectional interactions with neurons at tripartite synapses and its lateral communication with neighboring processes through intracellular calcium transport. Each process therefore acts as a semi-independent computational subunit, while coordinated calcium dynamics across multiple processes give rise to stable, distributed attractor states that could maintain information beyond transient neuronal firing.
Recent evidence from Lucas Benoit et al. [14] supports a plausible biological substrate for this interprocess communication. Their study revealed that astrocytic leaflets are interconnected by connexin43-mediated gap junctions, forming cytosolically continuous domains that permit calcium and IP3 to propagate between adjacent processes [172]. This organization allows localized calcium events to merge into broad, long-lasting elevations, consistent with the collective attractor dynamics envisioned in the NAAM model. However, connexin43 coupling alone is unlikely to fully account for intracellular calcium transport. Additional mechanisms may also support lateral propagation of signals between astrocytic processes. ER-based calcium diffusion [95], mitochondrial buffering [173,174], and local store-operated calcium entry (SOCE) [175] collectively enable coherent, memory-like calcium patterns [92,105]. Within this framework, astrocytes function not as simple modulators but as distributed computational arrays. Each process encodes a local calcium state that reflects recent neuronal activity [13], while process-to-process interactions integrate these local representations into a coherent astrocytic state [14]. The resulting calcium dynamics can persist for seconds to minutes [16,142], far longer than millisecond neuronal firing, and thus could provide a plausible mechanism for activity-silent memory, the temporary maintenance of information without sustained electrical activity. This property suggests that astrocytes may act as intermediate memory buffers that preserve contextual information across time gaps in neuronal signaling [34,176]. Mathematically, the NAAM model predicts supralinear memory capacity (Kmax ∝ N3), dramatically exceeding the linear scaling limit of classical Hopfield networks [177,178]. This enhancement arises from the astrocytic layer’s ability to couple large numbers of synapses through slow calcium-based feedback, effectively expanding the dimensionality of the representational space available for memory storage. A complementary theoretical study by Kozachkov et al. [179] further demonstrated that tripartite synapses can perform normalization operations analogous to the “self-attention” mechanism in Transformer architectures [180,181], linking astrocyte–neuron interactions to attention-like computation observed in artificial intelligence systems. These ideas remain theoretical and require substantial biological validation.
Functionally, this hybrid neuron–astrocyte computation may endow the brain with several key advantages. First, astrocytes provide temporal stability, bridging fast neural activity with slower biochemical integration. Second, they introduce nonlinear, context-dependent modulation, allowing networks to dynamically adjust learning rules according to ongoing physiological state. Third, they enable distributed redundancy, as astrocytic calcium states can maintain information even when neuronal activity ceases, ensuring robustness against transient disruptions. Together, these properties suggest that astrocytes play a fundamental role in maintaining the brain’s high-capacity, stable, and adaptive memory systems. However, whether astrocytic calcium states operate with sufficient specificity, speed, and reliability to support these computations in vivo remains an open question.
The NAAM model also generates clear experimental predictions. Disrupting calcium diffusion within astrocytes through pharmacological blockade of gap junctions [182] or genetic deletion of connexin43 [103] would be degrade memory consolidation and recall by fragmenting astrocytic attractor states. Indeed, astrocyte-specific Cx43 conditional knockout mice exhibit abolished barrel-cortex LTP and deficits in whisker-guided sensorimotor learning, demonstrating that loss of inter- and intra-astrocytic coupling impairs memory-relevant circuit plasticity [183]. Genetic deletion of IP3R2, which abolishes global astrocytic Ca2+ elevations, leads to selective impairments in remote memory recall, without affecting initial learning or short-term recall, in tasks such as contextual fear conditioning and spatial navigation [184]. Thus, astrocytic Ca2+ signaling via IP3R2 is required for memory consolidation under specific behavioral conditions, especially for the formation and maintenance of long-term memories. Conversely, enhancing process-to-process coupling may improve memory persistence or improve recovery after neuronal damage [93]. Moreover, neuromodulatory signals such as norepinephrine [185] or acetylcholine [7,186] may act as bias terms, adjusting astrocytic calcium thresholds and thereby tuning memory encoding efficiency according to behavioral context. Together, emerging computational and biological evidence support a unifying view of astrocytes as hybrid analog–digital processors [93], analog in their graded Ca2+ signaling and digital in their discrete synaptic connectivity. Earlier models emphasized their role in homeostasis and modulation [78], while the NAAM model extends this framework to include hypothesized roles in computation and memory storage. Testing its predictions by mapping calcium attractor dynamics or manipulating interprocess coupling will be crucial to understanding how neuron–astrocyte cooperation underlies the brain’s extraordinary capacity, efficiency, and resilience.
5. Future Directions
Recent advances have greatly expanded our understanding of astrocytes, revealing their roles in microdomain-level signaling, multisynaptic integration, cell-wide calcium dynamics, and network-scale ensemble activity. Despite this conceptual progress, fundamental questions remain unresolved. First, the precise coding logic of astrocytic calcium signaling is still poorly understood. How do distinct spatial and temporal calcium patterns correspond to specific forms of plasticity, synaptic modulation, or behavioral output? Second, the stability and persistence of astrocytic ensembles over time are unknown. Do learning-associated astrocytes form stable long-term networks, or are they dynamically reconfigured during memory updating and forgetting? Third, the molecular determinants of information storage remain to be defined, including calcium signaling mechanisms that trigger transcriptional or epigenetic changes to stabilize astrocytic states.
Technological limitations currently constrain progress. Super-resolution modalities such as STED microscopy offer nanoscale reconstructions of astrocytic leaflets but face severe photobleaching, phototoxicity, and limited axial resolution, restricting their applicability to live imaging and volumetric dynamics [187]. Calcium imaging similarly shows large discrepancies between in vitro and in vivo measurements: organic dyes such as Fluo-4AM underestimate microdomain activity by up to 85–90% due to poor labeling of fine perisynaptic processes, while widefield and confocal imaging obscure subcellular compartmentalization by integrating signals across optical planes. Emerging optical solutions, particularly two-photon microscopy, address many of these limitations by reducing photodamage, improving depth penetration, and enabling reliable detection of Ca2+ signals across somata, branches, and microdomains in intact circuits. Two-photon light-sheet approaches reduce photobleaching further while supporting rapid volumetric imaging. Coupling these methods with membrane-targeted GECIs, process-specific reporters, voltage or pH indicators, spatial transcriptomics, and proteomic tools will be essential for mapping astrocytic computation across microdomains, leaflets, and ensembles during behavior.
Future progress will require coordinated integration of multiple approaches. At the molecular level, multi-scale analyses such as advanced imaging, ribosome profiling, proximity labeling, and epigenomic mapping will be needed to achieve a mechanistic understanding of how intracellular calcium regulates transcriptional and translational programs. A single method is insufficient to delineate how calcium dynamics propagate to gene expression, metabolic coupling, and gliotransmission to convert transient activity into durable memory. We therefore integrate proteomics with transcriptomics in a spatiotemporally aligned manner to capture expression programs together with the concurrent rewiring of protein networks [157,188,189]. At the cellular and circuit levels, optogenetic and chemogenetic manipulation, including ensemble-tagging strategies, will allow selective interrogation of astrocyte subpopulations during behavior. Computational modeling remains crucial for linking subcellular processes to systems-level outcomes, enabling quantitative predictions about learning capacity, information flow, and metabolic efficiency. Finally, incorporating astrocytic dynamics into artificial neural-network architectures may inspire new paradigms in adaptive and energy-efficient computing.
6. Conclusions
Astrocytes have moved from the periphery of neuroscience to a central position in discussions of cognition, plasticity, and memory. Across nanoscale microdomains, multi-synaptic leaflets, single-cell territories, and network-level ensembles, they provide computational capabilities that complement and extend fast neuronal signaling. Their slow calcium dynamics, adhesion-based synaptic organization, local translation, and metabolic flexibility together offer a multi-scale mechanism that may stabilize, contextualize, and integrate information across time.
Key messages emerging from this review are as follows:
- (1)
- Astrocytes participate in memory formation through diverse mechanisms, including parallel processing and integration of inputs from multiple synapses, the structural and functional regulation of synapses, circuit-level neuronal regulation and the encoding of memory within astrocyte ensembles. These processes operate hierarchically across microdomains, multi-synaptic leaflets, single-cell territories, and network-level ensembles, enabling astrocytes to couple local plasticity with large-scale circuit adaptation.
- (2)
- Explaining the brain’s remarkable computational and mnemonic capabilities may require moving beyond neuron- and synapse-centric views to include astrocytic dynamics, as increasingly supported by emerging experimental and computational frameworks. These models propose that slow, integrative calcium states within astrocytes could complement fast neuronal signaling by providing temporally extended, activity-silent forms of information storage. Incorporating astrocytes into memory theory may therefore help reconcile how the brain achieves high capacity, stability, and robustness with limited metabolic cost. The stability and robustness potentially conferred by such astrocytic memory codes along with possible metabolic efficiency gains remain theoretically compelling but require further experimental validation.
Clinically, disruptions in these astrocytic processes ranging from impaired Ca2+ signaling and altered gliotransmission to defective local translation, metabolic dysfunction, and abnormal synaptic adhesion are increasingly implicated in disorders such as Alzheimer’s disease, Parkinson’s disease, autism spectrum disorder, depression, epilepsy, and traumatic brain injury. Understanding how astrocytes shape plasticity across scales may therefore reveal new therapeutic targets, including modulators of astrocytic calcium dynamics, metabolic pathways, adhesion molecules, and local translational control.
Future progress will require integrating high-resolution imaging, cell-type-specific manipulation, computational modeling, and behavioral ensemble tagging to critically test whether and how astrocytes encode, store, and retrieve information. By uncovering the multicellular logic of neuron–astrocyte cooperation, we may not only reshape fundamental theories of memory and learning but also accelerate the development of treatments for cognitive disorders and inspire new paradigms in adaptive, energy-efficient computing.
Author Contributions
M.Y.: Conceptualization, Visualization, Writing—original draft, Writing—review and editing. T.T.: Conceptualization, Funding acquisition, Visualization, Writing—original draft, Writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a Grant-in-Aid for Scientific Research B (21380936), PRESTO (21461219 and 24029397) from JST (T.T.), Brain Mind 2.0 from AMED (24019272, 24019528) (T.T.), The Ono Pharmaceutical Foundation for Oncology (T.T., T.M.), Immunology and Neurology (T.T.), the Memorial Foundation for Medical and Pharmaceutical Research (T.T.), a Research Grant from the JGC Saneyoshi Scholarship Foundation (T.T.), the Takeda Science Foundation (T.T.), and MEXT Grants-in-Aid for Scientific Research on Innovative Areas—“Cell type census of adaptive neuronal circuits: biological mechanisms of structural and functional organization (Adaptive Circuit Census, ACC)” (24984754) (T.T.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no competing financial interests.
References
- Bazargani, N.; Attwell, D. Astrocyte Calcium Signaling: The Third Wave. Nat. Neurosci. 2016, 19, 182–189. [Google Scholar] [CrossRef]
- Covelo, A.; Araque, A. Neuronal Activity Determines Distinct Gliotransmitter Release from a Single Astrocyte. Elife 2018, 7, e32237. [Google Scholar] [CrossRef]
- Panatier, A.; Vallée, J.; Haber, M.; Murai, K.K.; Lacaille, J.-C.; Robitaille, R. Astrocytes Are Endogenous Regulators of Basal Transmission at Central Synapses. Cell 2011, 146, 785–798. [Google Scholar] [CrossRef]
- Pascual, O.; Casper, K.B.; Kubera, C.; Zhang, J.; Revilla-Sanchez, R.; Sul, J.-Y.; Takano, H.; Moss, S.J.; McCarthy, K.; Haydon, P.G. Astrocytic Purinergic Signaling Coordinates Synaptic Networks. Science 2005, 310, 113–116. [Google Scholar] [CrossRef]
- Volterra, A.; Meldolesi, J. Astrocytes, from Brain Glue to Communication Elements: The Revolution Continues. Nat. Rev. Neurosci. 2005, 6, 626–640. [Google Scholar] [CrossRef]
- Min, R.; Nevian, T. Astrocyte Signaling Controls Spike Timing-Dependent Depression at Neocortical Synapses. Nat. Neurosci. 2012, 15, 746–753. [Google Scholar] [CrossRef]
- Navarrete, M.; Perea, G.; Fernandez de Sevilla, D.; Gómez-Gonzalo, M.; Núñez, A.; Martín, E.D.; Araque, A. Astrocytes Mediate in Vivo Cholinergic-Induced Synaptic Plasticity. PLoS Biol. 2012, 10, e1001259. [Google Scholar] [CrossRef]
- Falcón-Moya, R.; Pérez-Rodríguez, M.; Prius-Mengual, J.; Andrade-Talavera, Y.; Arroyo-García, L.E.; Pérez-Artés, R.; Mateos-Aparicio, P.; Guerra-Gomes, S.; Oliveira, J.F.; Flores, G.; et al. Astrocyte-Mediated Switch in Spike Timing-Dependent Plasticity during Hippocampal Development. Nat. Commun. 2020, 11, 4388. [Google Scholar] [CrossRef]
- Kofuji, P.; Newman, E.A. Potassium Buffering in the Central Nervous System. Neuroscience 2004, 129, 1045–1056. [Google Scholar] [CrossRef]
- Murphy-Royal, C.; Dupuis, J.; Groc, L.; Oliet, S.H.R. Astroglial Glutamate Transporters in the Brain: Regulating Neurotransmitter Homeostasis and Synaptic Transmission. J. Neurosci. Res. 2017, 95, 2140–2151. [Google Scholar] [CrossRef]
- Beard, E.; Lengacher, S.; Dias, S.; Magistretti, P.J.; Finsterwald, C. Astrocytes as Key Regulators of Brain Energy Metabolism: New Therapeutic Perspectives. Front. Physiol. 2021, 12, 825816. [Google Scholar] [CrossRef]
- Heller, J.P.; Rusakov, D.A. The Nanoworld of the Tripartite Synapse: Insights from Super-Resolution Microscopy. Front. Cell Neurosci. 2017, 11, 374. [Google Scholar] [CrossRef] [PubMed]
- Arizono, M.; Inavalli, V.V.G.K.; Panatier, A.; Pfeiffer, T.; Angibaud, J.; Levet, F.; Ter Veer, M.J.T.; Stobart, J.; Bellocchio, L.; Mikoshiba, K.; et al. Structural Basis of Astrocytic Ca2+ Signals at Tripartite Synapses. Nat. Commun. 2020, 11, 1906. [Google Scholar] [CrossRef] [PubMed]
- Benoit, L.; Hristovska, I.; Liaudet, N.; Jouneau, P.-H.; Fertin, A.; de Ceglia, R.; Litvin, D.; Di Castro, M.A.; Jevtic, M.; Zalachoras, I.; et al. Astrocytes Functionally Integrate Multiple Synapses via Specialized Leaflet Domains. Cell 2025, 188, 6453–6472.e16. [Google Scholar] [CrossRef] [PubMed]
- Bindocci, E.; Savtchouk, I.; Liaudet, N.; Becker, D.; Carriero, G.; Volterra, A. Three-Dimensional Ca2+ Imaging Advances Understanding of Astrocyte Biology. Science 2017, 356, eaai8185. [Google Scholar] [CrossRef]
- Stobart, J.L.; Ferrari, K.D.; Barrett, M.J.P.; Glück, C.; Stobart, M.J.; Zuend, M.; Weber, B. Cortical Circuit Activity Evokes Rapid Astrocyte Calcium Signals on a Similar Timescale to Neurons. Neuron 2018, 98, 726–735.e4. [Google Scholar] [CrossRef]
- Williamson, M.R.; Kwon, W.; Woo, J.; Ko, Y.; Maleki, E.; Yu, K.; Murali, S.; Sardar, D.; Deneen, B. Learning-Associated Astrocyte Ensembles Regulate Memory Recall. Nature 2025, 637, 478–486. [Google Scholar] [CrossRef]
- Citri, A.; Malenka, R.C. Synaptic Plasticity: Multiple Forms, Functions, and Mechanisms. Neuropsychopharmacology 2008, 33, 18–41. [Google Scholar] [CrossRef]
- Magee, J.C.; Grienberger, C. Synaptic Plasticity Forms and Functions. Annu. Rev. Neurosci. 2020, 43, 95–117. [Google Scholar] [CrossRef]
- Langille, J.J.; Brown, R.E. The Synaptic Theory of Memory: A Historical Survey and Reconciliation of Recent Opposition. Front. Syst. Neurosci. 2018, 12, 52. [Google Scholar] [CrossRef]
- Morris, R.G.D.O. Hebb: The Organization of Behavior, Wiley: New York; 1949. Brain Res. Bull. 1999, 50, 437. [Google Scholar] [CrossRef]
- Mateos-Aparicio, P.; Rodríguez-Moreno, A. The Impact of Studying Brain Plasticity. Front. Cell Neurosci. 2019, 13, 66. [Google Scholar]
- Martin, S.J.; Grimwood, P.D.; Morris, R.G. Synaptic Plasticity and Memory: An Evaluation of the Hypothesis. Annu. Rev. Neurosci. 2000, 23, 649–711. [Google Scholar] [CrossRef]
- Fusi, S. Memory Capacity of Neural Network Models. In The Oxford Handbook of Human Memory, Two Volume Pack; Oxford University Press: Oxford, UK, 2024; pp. 740–764. ISBN 9780190917982. [Google Scholar]
- Turrigiano, G. Homeostatic Synaptic Plasticity: Local and Global Mechanisms for Stabilizing Neuronal Function. Cold Spring Harb. Perspect. Biol. 2012, 4, a005736. [Google Scholar]
- Abraham, W.C.; Jones, O.D.; Glanzman, D.L. Is Plasticity of Synapses the Mechanism of Long-Term Memory Storage? NPJ Sci. Learn. 2019, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; McLaughlin, D.W.; Peskin, C.S. A Biochemical Description of Postsynaptic Plasticity-with Timescales Ranging from Milliseconds to Seconds. Proc. Natl. Acad. Sci. USA 2024, 121, e2311709121. [Google Scholar]
- Bittner, K.C.; Milstein, A.D.; Grienberger, C.; Romani, S.; Magee, J.C. Behavioral Time Scale Synaptic Plasticity Underlies CA1 Place Fields. Science 2017, 357, 1033–1036. [Google Scholar] [CrossRef]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Tripartite Synapses: Glia, the Unacknowledged Partner. Trends Neurosci. 1999, 22, 208–215. [Google Scholar] [CrossRef]
- Perea, G.; Navarrete, M.; Araque, A. Tripartite Synapses: Astrocytes Process and Control Synaptic Information. Trends Neurosci. 2009, 32, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Porto-Pazos, A.B.; Veiguela, N.; Mesejo, P.; Navarrete, M.; Alvarellos, A.; Ibáñez, O.; Pazos, A.; Araque, A. Artificial Astrocytes Improve Neural Network Performance. PLoS ONE 2011, 6, e19109. [Google Scholar] [CrossRef]
- Min, R.; Santello, M.; Nevian, T. The Computational Power of Astrocyte Mediated Synaptic Plasticity. Front. Comput. Neurosci. 2012, 6, 93. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Pan, L.; Liu, X. AstroNet: When Astrocyte Meets Artificial Neural Network. In Proceedings of the 2023 IEEE/CVF Conference on Computer Vision and Pattern Recognition (CVPR), Vancouver, BC, Canada, 18–22 June 2023; IEEE: New York, NY, USA, 2023. [Google Scholar]
- Gordleeva, S.Y.; Tsybina, Y.A.; Krivonosov, M.I.; Ivanchenko, M.V.; Zaikin, A.A.; Kazantsev, V.B.; Gorban, A.N. Modeling Working Memory in a Spiking Neuron Network Accompanied by Astrocytes. Front. Cell Neurosci. 2021, 15, 631485. [Google Scholar] [CrossRef]
- Yunusoglu, A.; Le, D.; Isik, M.; Dikmen, I.C.; Karadag, T. Neuromorphic Circuits with Spiking Astrocytes for Increased Energy Efficiency, Fault Tolerance, and Memory Capacitance. arXiv 2025. [Google Scholar] [CrossRef]
- Alvarellos-González, A.; Pazos, A.; Porto-Pazos, A.B. Computational Models of Neuron-Astrocyte Interactions Lead to Improved Efficacy in the Performance of Neural Networks. Comput. Math. Methods Med. 2012, 2012, 476324. [Google Scholar] [CrossRef]
- Sancho, L.; Contreras, M.; Allen, N.J. Glia as Sculptors of Synaptic Plasticity. Neurosci. Res. 2021, 167, 17–29. [Google Scholar]
- Todd, K.J.; Robitaille, R. Purinergic Modulation of Synaptic Signalling at the Neuromuscular Junction. Pflug. Arch. 2006, 452, 608–614. [Google Scholar]
- Pfrieger, F.W.; Barres, B.A. Synaptic Efficacy Enhanced by Glial Cells in Vitro. Science 1997, 277, 1684–1687. [Google Scholar] [CrossRef] [PubMed]
- Ventura, R.; Harris, K.M. Three-Dimensional Relationships between Hippocampal Synapses and Astrocytes. J. Neurosci. 1999, 19, 6897–6906. [Google Scholar] [CrossRef]
- Thomas, C.I.; Ryan, M.A.; McNabb, M.C.; Kamasawa, N.; Scholl, B. Astrocyte Coverage of Excitatory Synapses Correlates to Measures of Synapse Structure and Function in Ferret Primary Visual Cortex. Glia 2024, 72, 1785–1800. [Google Scholar] [CrossRef] [PubMed]
- Genoud, C.; Quairiaux, C.; Steiner, P.; Hirling, H.; Welker, E.; Knott, G.W. Plasticity of Astrocytic Coverage and Glutamate Transporter Expression in Adult Mouse Cortex. PLoS Biol. 2006, 4, e343. [Google Scholar]
- Deemyad, T.; Lüthi, J.; Spruston, N. Astrocytes Integrate and Drive Action Potential Firing in Inhibitory Subnetworks. Nat. Commun. 2018, 9, 4336. [Google Scholar] [CrossRef]
- Kol, A.; Adamsky, A.; Groysman, M.; Kreisel, T.; London, M.; Goshen, I. Astrocytes Contribute to Remote Memory Formation by Modulating Hippocampal-Cortical Communication during Learning. Nat. Neurosci. 2020, 23, 1229–1239. [Google Scholar] [CrossRef]
- Araque, A.; Carmignoto, G.; Haydon, P.G.; Oliet, S.H.R.; Robitaille, R.; Volterra, A. Gliotransmitters Travel in Time and Space. Neuron 2014, 81, 728–739. [Google Scholar] [CrossRef] [PubMed]
- Goenaga, J.; Araque, A.; Kofuji, P.; Herrera Moro Chao, D. Calcium Signaling in Astrocytes and Gliotransmitter Release. Front. Synaptic Neurosci. 2023, 15, 1138577. [Google Scholar] [CrossRef]
- Ding, S.; Fellin, T.; Zhu, Y.; Lee, S.-Y.; Auberson, Y.P.; Meaney, D.F.; Coulter, D.A.; Carmignoto, G.; Haydon, P.G. Enhanced Astrocytic Ca2+ Signals Contribute to Neuronal Excitotoxicity after Status Epilepticus. J. Neurosci. 2007, 27, 10674–10684. [Google Scholar] [CrossRef]
- Kang, J.; Jiang, L.; Goldman, S.A.; Nedergaard, M. Astrocyte-Mediated Potentiation of Inhibitory Synaptic Transmission. Nat. Neurosci. 1998, 1, 683–692. [Google Scholar] [CrossRef]
- Shigetomi, E.; Suzuki, H.; Hirayama, Y.J.; Sano, F.; Nagai, Y.; Yoshihara, K.; Koga, K.; Tateoka, T.; Yoshioka, H.; Shinozaki, Y.; et al. Disease-Relevant Upregulation of P2Y1 Receptor in Astrocytes Enhances Neuronal Excitability via IGFBP2. Nat. Commun. 2024, 15, 6525. [Google Scholar] [CrossRef] [PubMed]
- Parpura, V.; Basarsky, T.A.; Liu, F.; Jeftinija, K.; Jeftinija, S.; Haydon, P.G. Glutamate-Mediated Astrocyte-Neuron Signalling. Nature 1994, 369, 744–747. [Google Scholar] [CrossRef] [PubMed]
- Lalo, U.; Palygin, O.; Rasooli-Nejad, S.; Andrew, J.; Haydon, P.G.; Pankratov, Y. Exocytosis of ATP from Astrocytes Modulates Phasic and Tonic Inhibition in the Neocortex. PLoS Biol. 2014, 12, e1001747. [Google Scholar] [CrossRef]
- Henneberger, C.; Papouin, T.; Oliet, S.H.R.; Rusakov, D.A. Long-Term Potentiation Depends on Release of d-Serine from Astrocytes. Nature 2010, 463, 232–236. [Google Scholar]
- Yoon, B.-E.; Lee, C.J. GABA as a Rising Gliotransmitter. Front. Neural. Circuits 2014, 8, 141. [Google Scholar] [CrossRef]
- Coyle, J.T.; Balu, D.; Wolosker, H. D-Serine, the Shape-Shifting NMDA Receptor Co-Agonist. Neurochem. Res. 2020, 45, 1344–1353. [Google Scholar] [PubMed]
- Lovatt, D.; Xu, Q.; Liu, W.; Takano, T.; Smith, N.A.; Schnermann, J.; Tieu, K.; Nedergaard, M. Neuronal Adenosine Release, and Not Astrocytic ATP Release, Mediates Feedback Inhibition of Excitatory Activity. Proc. Natl. Acad. Sci. USA 2012, 109, 6265–6270. [Google Scholar] [CrossRef] [PubMed]
- Florian, C.; Vecsey, C.G.; Halassa, M.M.; Haydon, P.G.; Abel, T. Astrocyte-Derived Adenosine and A1 Receptor Activity Contribute to Sleep Loss-Induced Deficits in Hippocampal Synaptic Plasticity and Memory in Mice. J. Neurosci. 2011, 31, 6956–6962. [Google Scholar] [CrossRef]
- Woo, J.; Min, J.O.; Kang, D.-S.; Kim, Y.S.; Jung, G.H.; Park, H.J.; Kim, S.; An, H.; Kwon, J.; Kim, J.; et al. Control of Motor Coordination by Astrocytic Tonic GABA Release through Modulation of Excitation/Inhibition Balance in Cerebellum. Proc. Natl. Acad. Sci. USA 2018, 115, 5004–5009. [Google Scholar] [CrossRef]
- Maltsev, A.; Roshchin, M.; Bezprozvanny, I.; Smirnov, I.; Vlasova, O.; Balaban, P.; Borodinova, A. Bidirectional Regulation by “Star Forces”: Ionotropic Astrocyte’s Optical Stimulation Suppresses Synaptic Plasticity, Metabotropic One Strikes Back. Hippocampus 2023, 33, 18–36. [Google Scholar] [PubMed]
- Li, L.; Zhou, J.; Sun, H.; Liu, J.; Wang, H.; Liu, X.; Wang, C. A Computational Model to Investigate GABA-Activated Astrocyte Modulation of Neuronal Excitation. Comput. Math. Methods Med. 2020, 2020, 8750167. [Google Scholar] [CrossRef]
- Fiacco, T.A.; McCarthy, K.D. Multiple Lines of Evidence Indicate That Gliotransmission Does Not Occur under Physiological Conditions. J. Neurosci. 2018, 38, 3–13. [Google Scholar] [CrossRef]
- Sloan, S.A.; Barres, B.A. Looks Can Be Deceiving: Reconsidering the Evidence for Gliotransmission. Neuron 2014, 84, 1112–1115. [Google Scholar] [CrossRef]
- Gonçalves-Ribeiro, J.; Vaz, S.H. The IP3R2 Knockout Mice in Behavior: A Blessing or a Curse? J. Neurochem. 2025, 169, e70062. [Google Scholar] [CrossRef]
- Agulhon, C.; Fiacco, T.A.; McCarthy, K.D. Hippocampal Short- and Long-Term Plasticity Are Not Modulated by Astrocyte Ca2+ Signaling. Science 2010, 327, 1250–1254. [Google Scholar] [CrossRef]
- Savtchouk, I.; Volterra, A. Gliotransmission: Beyond Black-and-White. J. Neurosci. 2018, 38, 14–25. [Google Scholar] [CrossRef]
- Wolosker, H.; Balu, D.T.; Coyle, J.T. Astroglial versus Neuronal D-Serine: Check Your Controls! Trends Neurosci. 2017, 40, 520–522. [Google Scholar] [CrossRef]
- Papouin, T.; Henneberger, C.; Rusakov, D.A.; Oliet, S.H.R. Astroglial versus Neuronal D-Serine: Fact Checking. Trends Neurosci. 2017, 40, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Sahlender, D.A.; Savtchouk, I.; Volterra, A. What Do We Know about Gliotransmitter Release from Astrocytes? Philos. Trans. R Soc. Lond. B Biol. Sci. 2014, 369, 20130592. [Google Scholar] [CrossRef]
- Pajarillo, E.; Rizor, A.; Lee, J.; Aschner, M.; Lee, E. The Role of Astrocytic Glutamate Transporters GLT-1 and GLAST in Neurological Disorders: Potential Targets for Neurotherapeutics. Neuropharmacology 2019, 161, 107559. [Google Scholar] [CrossRef]
- Bataveljic, D.; Pivonkova, H.; de Concini, V.; Hébert, B.; Ezan, P.; Briault, S.; Bemelmans, A.-P.; Pichon, J.; Menuet, A.; Rouach, N. Astroglial Kir4.1 Potassium Channel Deficit Drives Neuronal Hyperexcitability and Behavioral Defects in Fragile X Syndrome Mouse Model. Nat. Commun. 2024, 15, 3583. [Google Scholar] [CrossRef]
- Mason, S. Lactate Shuttles in Neuroenergetics-Homeostasis, Allostasis and Beyond. Front. Neurosci. 2017, 11, 43. [Google Scholar] [CrossRef]
- Kim, Y.; Dube, S.E.; Park, C.B. Brain Energy Homeostasis: The Evolution of the Astrocyte-Neuron Lactate Shuttle Hypothesis. Korean J. Physiol. Pharmacol. 2025, 29, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Christopherson, K.S.; Ullian, E.M.; Stokes, C.C.A.; Mullowney, C.E.; Hell, J.W.; Agah, A.; Lawler, J.; Mosher, D.F.; Bornstein, P.; Barres, B.A. Thrombospondins Are Astrocyte-Secreted Proteins That Promote CNS Synaptogenesis. Cell 2005, 120, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Kucukdereli, H.; Allen, N.J.; Lee, A.T.; Feng, A.; Ozlu, M.I.; Conatser, L.M.; Chakraborty, C.; Workman, G.; Weaver, M.; Sage, E.H.; et al. Control of Excitatory CNS Synaptogenesis by Astrocyte-Secreted Proteins Hevin and SPARC. Proc. Natl. Acad. Sci. USA 2011, 108, E440-9. [Google Scholar]
- Singh, S.K.; Stogsdill, J.A.; Pulimood, N.S.; Dingsdale, H.; Kim, Y.H.; Pilaz, L.-J.; Kim, I.H.; Manhaes, A.C.; Rodrigues, W.S., Jr.; Pamukcu, A.; et al. Astrocytes Assemble Thalamocortical Synapses by Bridging NRX1α and NL1 via Hevin. Cell 2016, 164, 183–196. [Google Scholar] [CrossRef]
- Yang, M.-J.; Jia, M.; Cai, M.; Feng, X.; Huang, L.-N.; Yang, J.-J. Central Neuropeptides as Key Modulators of Astrocyte Function in Neurodegenerative and Neuropsychiatric Disorders. Psychopharmacology 2025, 242, 2353–2371. [Google Scholar] [CrossRef]
- Navarrete, M.; Cuartero, M.I.; Palenzuela, R.; Draffin, J.E.; Konomi, A.; Serra, I.; Colié, S.; Castaño-Castaño, S.; Hasan, M.T.; Nebreda, Á.R.; et al. Astrocytic P38α MAPK Drives NMDA Receptor-Dependent Long-Term Depression and Modulates Long-Term Memory. Nat. Commun. 2019, 10, 2968. [Google Scholar] [PubMed]
- Durkee, C.; Kofuji, P.; Navarrete, M.; Araque, A. Astrocyte and Neuron Cooperation in Long-Term Depression. Trends Neurosci. 2021, 44, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Santello, M.; Toni, N.; Volterra, A. Astrocyte Function from Information Processing to Cognition and Cognitive Impairment. Nat. Neurosci. 2019, 22, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Volterra, A.; Liaudet, N.; Savtchouk, I. Astrocyte Ca2+ Signalling: An Unexpected Complexity. Nat. Rev. Neurosci. 2014, 15, 327–335. [Google Scholar]
- Semyanov, A.; Henneberger, C.; Agarwal, A. Making Sense of Astrocytic Calcium Signals—From Acquisition to Interpretation. Nat. Rev. Neurosci. 2020, 21, 551–564. [Google Scholar]
- Lia, A.; Henriques, V.J.; Zonta, M.; Chiavegato, A.; Carmignoto, G.; Gómez-Gonzalo, M.; Losi, G. Calcium Signals in Astrocyte Microdomains, a Decade of Great Advances. Front. Cell Neurosci. 2021, 15, 673433. [Google Scholar]
- Pestana, F.; Edwards-Faret, G.; Belgard, T.G.; Martirosyan, A.; Holt, M.G. No Longer Underappreciated: The Emerging Concept of Astrocyte Heterogeneity in Neuroscience. Brain Sci. 2020, 10, 168. [Google Scholar] [CrossRef]
- Khakh, B.S.; Deneen, B. The Emerging Nature of Astrocyte Diversity. Annu. Rev. Neurosci. 2019, 42, 187–207. [Google Scholar] [CrossRef]
- Rupprecht, P.; Duss, S.N.; Becker, D.; Lewis, C.M.; Bohacek, J.; Helmchen, F. Centripetal Integration of Past Events in Hippocampal Astrocytes Regulated by Locus Coeruleus. Nat. Neurosci. 2024, 27, 927–939. [Google Scholar] [CrossRef]
- Sun, W.; Liu, Z.; Jiang, X.; Chen, M.B.; Dong, H.; Liu, J.; Südhof, T.C.; Quake, S.R. Spatial Transcriptomics Reveal Neuron-Astrocyte Synergy in Long-Term Memory. Nature 2024, 627, 374–381. [Google Scholar] [CrossRef]
- Murphy-Royal, C.; Ching, S.; Papouin, T. A Conceptual Framework for Astrocyte Function. Nat. Neurosci. 2023, 26, 1848–1856. [Google Scholar] [CrossRef] [PubMed]
- Denizot, A.; Arizono, M.; Nägerl, U.V.; Berry, H.; De Schutter, E. Control of Ca2+ Signals by Astrocyte Nanoscale Morphology at Tripartite Synapses. Glia 2022, 70, 2378–2391. [Google Scholar] [CrossRef] [PubMed]
- Kastanenka, K.V.; Moreno-Bote, R.; De Pittà, M.; Perea, G.; Eraso-Pichot, A.; Masgrau, R.; Poskanzer, K.E.; Galea, E. A Roadmap to Integrate Astrocytes into Systems Neuroscience. Glia 2020, 68, 5–26. [Google Scholar] [PubMed]
- Lee, H.S.; Ghetti, A.; Pinto-Duarte, A.; Wang, X.; Dziewczapolski, G.; Galimi, F.; Huitron-Resendiz, S.; Piña-Crespo, J.C.; Roberts, A.J.; Verma, I.M.; et al. Astrocytes Contribute to Gamma Oscillations and Recognition Memory. Proc. Natl. Acad. Sci. USA 2014, 111, E3343-52. [Google Scholar] [CrossRef]
- Fellin, T.; Pascual, O.; Gobbo, S.; Pozzan, T.; Haydon, P.G.; Carmignoto, G. Neuronal Synchrony Mediated by Astrocytic Glutamate through Activation of Extrasynaptic NMDA Receptors. Neuron 2004, 43, 729–743. [Google Scholar] [CrossRef]
- Dewa, K.-I.; Kaseda, K.; Kuwahara, A.; Kubotera, H.; Yamasaki, A.; Awata, N.; Komori, A.; Holtz, M.A.; Kasai, A.; Skibbe, H.; et al. The Astrocytic Ensemble Acts as a Multiday Trace to Stabilize Memory. Nature 2025, 648, 1–11. [Google Scholar] [CrossRef]
- Lines, J.; Baraibar, A.; Nanclares, C.; Martin, E.D.; Aguilar, J.; Kofuji, P.; Navarrete, M.; Araque, A. A Spatial Threshold for Astrocyte Calcium Surge. Elife 2024, 12, RP90046. [Google Scholar] [CrossRef]
- Kozachkov, L.; Slotine, J.-J.; Krotov, D. Neuron-Astrocyte Associative Memory. Proc. Natl. Acad. Sci. USA 2025, 122, e2417788122. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Nedergaard, M. Physiology of Astroglia. Physiol. Rev. 2018, 98, 239–389. [Google Scholar] [CrossRef] [PubMed]
- Arizono, M.; Bannai, H.; Nakamura, K.; Niwa, F.; Enomoto, M.; Matsu-Ura, T.; Miyamoto, A.; Sherwood, M.W.; Nakamura, T.; Mikoshiba, K. Receptor-Selective Diffusion Barrier Enhances Sensitivity of Astrocytic Processes to Metabotropic Glutamate Receptor Stimulation. Sci. Signal. 2012, 5, ra27. [Google Scholar] [CrossRef]
- Di Castro, M.A.; Chuquet, J.; Liaudet, N.; Bhaukaurally, K.; Santello, M.; Bouvier, D.; Tiret, P.; Volterra, A. Local Ca2+ Detection and Modulation of Synaptic Release by Astrocytes. Nat. Neurosci. 2011, 14, 1276–1284. [Google Scholar] [CrossRef]
- Sherwood, M.W.; Arizono, M.; Panatier, A.; Mikoshiba, K.; Oliet, S.H.R. Astrocytic IP3Rs: Beyond IP3R2. Front. Cell Neurosci. 2021, 15, 695817. [Google Scholar] [CrossRef]
- Sakers, K.; Lake, A.M.; Khazanchi, R.; Ouwenga, R.; Vasek, M.J.; Dani, A.; Dougherty, J.D. Astrocytes Locally Translate Transcripts in Their Peripheral Processes. Proc. Natl. Acad. Sci. USA 2017, 114, E3830–E3838. [Google Scholar] [CrossRef]
- Mazaré, N.; Oudart, M.; Moulard, J.; Cheung, G.; Tortuyaux, R.; Mailly, P.; Mazaud, D.; Bemelmans, A.-P.; Boulay, A.-C.; Blugeon, C.; et al. Local Translation in Perisynaptic Astrocytic Processes Is Specific and Changes after Fear Conditioning. Cell Rep. 2020, 32, 108076. [Google Scholar] [CrossRef]
- de la Cruz-Gambra, A.; Baleriola, J. Astrocyte-Secreted Factors Modulate Synaptic Protein Synthesis as Revealed by Puromycin Labeling of Isolated Synaptosomes. Front. Mol. Neurosci. 2025, 18, 1427036. [Google Scholar] [CrossRef]
- Costa-Mattioli, M.; Gobert, D.; Stern, E.; Gamache, K.; Colina, R.; Cuello, C.; Sossin, W.; Kaufman, R.; Pelletier, J.; Rosenblum, K.; et al. EIF2alpha Phosphorylation Bidirectionally Regulates the Switch from Short- to Long-Term Synaptic Plasticity and Memory. Cell 2007, 129, 195–206. [Google Scholar] [CrossRef]
- de Ceglia, R.; Ledonne, A.; Litvin, D.G.; Lind, B.L.; Carriero, G.; Latagliata, E.C.; Bindocci, E.; Di Castro, M.A.; Savtchouk, I.; Vitali, I.; et al. Specialized Astrocytes Mediate Glutamatergic Gliotransmission in the CNS. Nature 2023, 622, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Giaume, C.; Naus, C.C.; Sáez, J.C.; Leybaert, L. Glial Connexins and Pannexins in the Healthy and Diseased Brain. Physiol. Rev. 2021, 101, 93–145. [Google Scholar] [CrossRef] [PubMed]
- Blomstrand, F.; Aberg, N.D.; Eriksson, P.S.; Hansson, E.; Rönnbäck, L. Extent of Intercellular Calcium Wave Propagation Is Related to Gap Junction Permeability and Level of Connexin-43 Expression in Astrocytes in Primary Cultures from Four Brain Regions. Neuroscience 1999, 92, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Scemes, E.; Giaume, C. Astrocyte Calcium Waves: What They Are and What They Do. Glia 2006, 54, 716–725. [Google Scholar] [CrossRef]
- Hulme, S.R.; Jones, O.D.; Raymond, C.R.; Sah, P.; Abraham, W.C. Mechanisms of Heterosynaptic Metaplasticity. Philos. Trans. R Soc. Lond. B Biol. Sci. 2014, 369, 20130148. [Google Scholar] [CrossRef] [PubMed]
- Aten, S.; Kiyoshi, C.M.; Arzola, E.P.; Patterson, J.A.; Taylor, A.T.; Du, Y.; Guiher, A.M.; Philip, M.; Camacho, E.G.; Mediratta, D.; et al. Ultrastructural View of Astrocyte Arborization, Astrocyte-Astrocyte and Astrocyte-Synapse Contacts, Intracellular Vesicle-like Structures, and Mitochondrial Network. Prog. Neurobiol. 2022, 213, 102264. [Google Scholar] [CrossRef]
- Bushong, E.A.; Martone, M.E.; Jones, Y.Z.; Ellisman, M.H. Protoplasmic Astrocytes in CA1 Stratum Radiatum Occupy Separate Anatomical Domains. J. Neurosci. 2002, 22, 183–192. [Google Scholar] [CrossRef]
- Poskanzer, K.E.; Yuste, R. Astrocytes Regulate Cortical State Switching in Vivo. Proc. Natl. Acad. Sci. USA 2016, 113, E2675-84. [Google Scholar]
- Bocchi, R.; Thorwirth, M.; Simon-Ebert, T.; Koupourtidou, C.; Clavreul, S.; Kolf, K.; Della Vecchia, P.; Bottes, S.; Jessberger, S.; Zhou, J.; et al. Astrocyte Heterogeneity Reveals Region-Specific Astrogenesis in the White Matter. Nat. Neurosci. 2025, 28, 457–469. [Google Scholar] [CrossRef]
- Durkee, C.A.; Araque, A. Diversity and Specificity of Astrocyte-Neuron Communication. Neuroscience 2019, 396, 73–78. [Google Scholar]
- Agulhon, C.; Petravicz, J.; McMullen, A.B.; Sweger, E.J.; Minton, S.K.; Taves, S.R.; Casper, K.B.; Fiacco, T.A.; McCarthy, K.D. What Is the Role of Astrocyte Calcium in Neurophysiology? Neuron 2008, 59, 932–946. [Google Scholar] [CrossRef]
- Shirvalkar, P.R.; Rapp, P.R.; Shapiro, M.L. Bidirectional Changes to Hippocampal Theta-Gamma Comodulation Predict Memory for Recent Spatial Episodes. Proc. Natl. Acad. Sci. USA 2010, 107, 7054–7059. [Google Scholar] [CrossRef]
- Zenke, F.; Gerstner, W. Hebbian Plasticity Requires Compensatory Processes on Multiple Timescales. Philos. Trans. R Soc. Lond. B Biol. Sci. 2017, 372, 20160259. [Google Scholar] [CrossRef]
- Serra, I.; Martín-Monteagudo, C.; Sánchez Romero, J.; Quintanilla, J.P.; Ganchala, D.; Arevalo, M.-A.; García-Marqués, J.; Navarrete, M. Astrocyte Ensembles Manipulated with AstroLight Tune Cue-Motivated Behavior. Nat. Neurosci. 2025, 28, 616–626. [Google Scholar] [CrossRef]
- Adamsky, A.; Kol, A.; Kreisel, T.; Doron, A.; Ozeri-Engelhard, N.; Melcer, T.; Refaeli, R.; Horn, H.; Regev, L.; Groysman, M.; et al. Astrocytic Activation Generates DE Novo Neuronal Potentiation and Memory Enhancement. Cell 2018, 174, 59–71.e14. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Jin, S.-Y.; Yang, J.-M.; Gao, T.-M. The Memory Orchestra: Contribution of Astrocytes. Neurosci. Bull. 2023, 39, 409–424. [Google Scholar] [CrossRef]
- Trpevski, D.; Khodadadi, Z.; Carannante, I.; Hellgren Kotaleski, J. Glutamate Spillover Drives Robust All-or-None Dendritic Plateau Potentials-an in Silico Investigation Using Models of Striatal Projection Neurons. Front. Cell Neurosci. 2023, 17, 1196182. [Google Scholar] [CrossRef]
- Hires, S.A.; Zhu, Y.; Tsien, R.Y. Optical Measurement of Synaptic Glutamate Spillover and Reuptake by Linker Optimized Glutamate-Sensitive Fluorescent Reporters. Proc. Natl. Acad. Sci. USA 2008, 105, 4411–4416. [Google Scholar] [CrossRef] [PubMed]
- Gordleeva, S.Y.; Stasenko, S.V.; Semyanov, A.V.; Dityatev, A.E.; Kazantsev, V.B. Bi-Directional Astrocytic Regulation of Neuronal Activity within a Network. Front. Comput. Neurosci. 2012, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Papouin, T.; Dunphy, J.; Tolman, M.; Foley, J.C.; Haydon, P.G. Astrocytic Control of Synaptic Function. Philos. Trans. R Soc. Lond. B Biol. Sci. 2017, 372, 20160154. [Google Scholar] [CrossRef] [PubMed]
- Hochmuth, L.; Hirrlinger, J. Physiological and Pathological Role of MTOR Signaling in Astrocytes. Neurochem. Res. 2024, 50, 53. [Google Scholar] [CrossRef]
- Winfree, R.L.; Nolan, E.; Blennow, K.; Zetterberg, H.; Gifford, K.A.; Pechman, K.R.; Schneider, J.; Bennett, D.A.; Petyuk, V.A.; Jefferson, A.L.; et al. Vascular Endothelial Growth Factor Receptor-1 (FLT1) Interactions with Amyloid-Beta in Alzheimer’s Disease: A Putative Biomarker of Amyloid-Induced Vascular Damage. Neurobiol. Aging 2025, 147, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Chiou, B.; Neely, E.B.; Mcdevitt, D.S.; Simpson, I.A.; Connor, J.R. Transferrin and H-Ferritin Involvement in Brain Iron Acquisition during Postnatal Development: Impact of Sex and Genotype. J. Neurochem. 2020, 152, 381–396. [Google Scholar] [CrossRef]
- Grimm, C.M.; Aksamaz, S.; Schulz, S.; Teutsch, J.; Sicinski, P.; Liss, B.; Kätzel, D. Schizophrenia-Related Cognitive Dysfunction in the Cyclin-D2 Knockout Mouse Model of Ventral Hippocampal Hyperactivity. Transl. Psychiatry 2018, 8, 212. [Google Scholar] [CrossRef]
- Sharp, A.H.; Nucifora, F.C., Jr.; Blondel, O.; Sheppard, C.A.; Zhang, C.; Snyder, S.H.; Russell, J.T.; Ryugo, D.K.; Ross, C.A. Differential Cellular Expression of Isoforms of Inositol 1,4,5-Triphosphate Receptors in Neurons and Glia in Brain. J. Comp. Neurol. 1999, 406, 207–220. [Google Scholar] [CrossRef]
- Fujii, Y.; Maekawa, S.; Morita, M. Astrocyte Calcium Waves Propagate Proximally by Gap Junction and Distally by Extracellular Diffusion of ATP Released from Volume-Regulated Anion Channels. Sci. Rep. 2017, 7, 13115. [Google Scholar] [CrossRef]
- Mothet, J.-P.; Pollegioni, L.; Ouanounou, G.; Martineau, M.; Fossier, P.; Baux, G. Glutamate Receptor Activation Triggers a Calcium-Dependent and SNARE Protein-Dependent Release of the Gliotransmitter D-Serine. Proc. Natl. Acad. Sci. USA 2005, 102, 5606–5611. [Google Scholar] [CrossRef] [PubMed]
- Miguel-Quesada, C.; Zaforas, M.; Herrera-Pérez, S.; Lines, J.; Fernández-López, E.; Alonso-Calviño, E.; Ardaya, M.; Soria, F.N.; Araque, A.; Aguilar, J.; et al. Astrocytes Adjust the Dynamic Range of Cortical Network Activity to Control Modality-Specific Sensory Information Processing. Cell Rep. 2023, 42, 112950. [Google Scholar] [CrossRef] [PubMed]
- Gordleeva, S.Y.; Ermolaeva, A.V.; Kastalskiy, I.A.; Kazantsev, V.B. Astrocyte as Spatiotemporal Integrating Detector of Neuronal Activity. Front. Physiol. 2019, 10, 294. [Google Scholar] [CrossRef]
- Henneberger, C.; Bard, L.; Panatier, A.; Reynolds, J.P.; Kopach, O.; Medvedev, N.I.; Minge, D.; Herde, M.K.; Anders, S.; Kraev, I.; et al. LTP Induction Boosts Glutamate Spillover by Driving Withdrawal of Perisynaptic Astroglia. Neuron 2020, 108, 919–936.e11. [Google Scholar] [CrossRef]
- Halassa, M.M.; Fellin, T.; Takano, H.; Dong, J.-H.; Haydon, P.G. Synaptic Islands Defined by the Territory of a Single Astrocyte. J. Neurosci. 2007, 27, 6473–6477. [Google Scholar] [CrossRef]
- Cahill, M.K.; Collard, M.; Tse, V.; Reitman, M.E.; Etchenique, R.; Kirst, C.; Poskanzer, K.E. Network-Level Encoding of Local Neurotransmitters in Cortical Astrocytes. Nature 2024, 629, 146–153. [Google Scholar] [CrossRef]
- Allen, N.J.; Lyons, D.A. Glia as Architects of Central Nervous System Formation and Function. Science 2018, 362, 181–185. [Google Scholar] [CrossRef]
- Endo, F.; Kasai, A.; Soto, J.S.; Yu, X.; Qu, Z.; Hashimoto, H.; Gradinaru, V.; Kawaguchi, R.; Khakh, B.S. Molecular Basis of Astrocyte Diversity and Morphology across the CNS in Health and Disease. Science 2022, 378, eadc9020. [Google Scholar] [CrossRef]
- Stogsdill, J.A.; Ramirez, J.; Liu, D.; Kim, Y.H.; Baldwin, K.T.; Enustun, E.; Ejikeme, T.; Ji, R.-R.; Eroglu, C. Astrocytic Neuroligins Control Astrocyte Morphogenesis and Synaptogenesis. Nature 2017, 551, 192–197. [Google Scholar] [CrossRef]
- Farmer, W.T.; Abrahamsson, T.; Chierzi, S.; Lui, C.; Zaelzer, C.; Jones, E.V.; Bally, B.P.; Chen, G.G.; Théroux, J.-F.; Peng, J.; et al. Neurons Diversify Astrocytes in the Adult Brain through Sonic Hedgehog Signaling. Science 2016, 351, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Szabó, Z.; Héja, L.; Szalay, G.; Kékesi, O.; Füredi, A.; Szebényi, K.; Dobolyi, Á.; Orbán, T.I.; Kolacsek, O.; Tompa, T.; et al. Extensive Astrocyte Synchronization Advances Neuronal Coupling in Slow Wave Activity in Vivo. Sci. Rep. 2017, 7, 6018. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.-J.; Aćimović, J.; Manninen, T.; Ahokainen, I.; Stapmanns, J.; Lehtimäki, M.; Diesmann, M.; van Albada, S.J.; Plesser, H.E.; Linne, M.-L. Modeling Neuron-Astrocyte Interactions in Neural Networks Using Distributed Simulation. PLoS Comput. Biol. 2025, 21, e1013503. [Google Scholar] [CrossRef] [PubMed]
- Kofuji, P.; Araque, A. Astrocytes and Behavior. Annu. Rev. Neurosci. 2021, 44, 49–67. [Google Scholar] [CrossRef]
- Guerra-Gomes, S.; Sousa, N.; Pinto, L.; Oliveira, J.F. Functional Roles of Astrocyte Calcium Elevations: From Synapses to Behavior. Front. Cell Neurosci. 2017, 11, 427. [Google Scholar] [CrossRef]
- Haydon, P.G.; Nedergaard, M. How Do Astrocytes Participate in Neural Plasticity? Cold Spring Harb. Perspect. Biol. 2014, 7, a020438. [Google Scholar] [CrossRef]
- Koizumi, S.; Fujishita, K.; Tsuda, M.; Shigemoto-Mogami, Y.; Inoue, K. Dynamic Inhibition of Excitatory Synaptic Transmission by Astrocyte-Derived ATP in Hippocampal Cultures. Proc. Natl. Acad. Sci. USA 2003, 100, 11023–11028. [Google Scholar] [CrossRef]
- Untiet, V.; Beinlich, F.R.M.; Kusk, P.; Kang, N.; Ladrón-de-Guevara, A.; Song, W.; Kjaerby, C.; Andersen, M.; Hauglund, N.; Bojarowska, Z.; et al. Astrocytic Chloride Is Brain State Dependent and Modulates Inhibitory Neurotransmission in Mice. Nat. Commun. 2023, 14, 1871. [Google Scholar] [CrossRef]
- Theparambil, S.M.; Kopach, O.; Braga, A.; Nizari, S.; Hosford, P.S.; Sagi-Kiss, V.; Hadjihambi, A.; Konstantinou, C.; Esteras, N.; Gutierrez Del Arroyo, A.; et al. Adenosine Signalling to Astrocytes Coordinates Brain Metabolism and Function. Nature 2024, 632, 139–146. [Google Scholar] [CrossRef]
- Batiuk, M.Y.; Martirosyan, A.; Wahis, J.; de Vin, F.; Marneffe, C.; Kusserow, C.; Koeppen, J.; Viana, J.F.; Oliveira, J.F.; Voet, T.; et al. Identification of Region-Specific Astrocyte Subtypes at Single Cell Resolution. Nat. Commun. 2020, 11, 1220. [Google Scholar] [CrossRef]
- Schober, A.L.; Wicki-Stordeur, L.E.; Murai, K.K.; Swayne, L.A. Foundations and Implications of Astrocyte Heterogeneity during Brain Development and Disease. Trends Neurosci. 2022, 45, 692–703. [Google Scholar] [CrossRef] [PubMed]
- Herrero-Navarro, Á.; Puche-Aroca, L.; Moreno-Juan, V.; Sempere-Ferràndez, A.; Espinosa, A.; Susín, R.; Torres-Masjoan, L.; Leyva-Díaz, E.; Karow, M.; Figueres-Oñate, M.; et al. Astrocytes and Neurons Share Region-Specific Transcriptional Signatures That Confer Regional Identity to Neuronal Reprogramming. Sci. Adv. 2021, 7, eabe8978. [Google Scholar] [CrossRef] [PubMed]
- Matos, M.; Bosson, A.; Riebe, I.; Reynell, C.; Vallée, J.; Laplante, I.; Panatier, A.; Robitaille, R.; Lacaille, J.-C. Astrocytes Detect and Upregulate Transmission at Inhibitory Synapses of Somatostatin Interneurons onto Pyramidal Cells. Nat. Commun. 2018, 9, 4254. [Google Scholar] [CrossRef] [PubMed]
- Pittolo, S.; Yokoyama, S.; Willoughby, D.D.; Taylor, C.R.; Reitman, M.E.; Tse, V.; Wu, Z.; Etchenique, R.; Li, Y.; Poskanzer, K.E. Dopamine Activates Astrocytes in Prefrontal Cortex via A1-Adrenergic Receptors. Cell Rep. 2022, 40, 111426. [Google Scholar] [CrossRef]
- Mederos, S.; Sánchez-Puelles, C.; Esparza, J.; Valero, M.; Ponomarenko, A.; Perea, G. GABAergic Signaling to Astrocytes in the Prefrontal Cortex Sustains Goal-Directed Behaviors. Nat. Neurosci. 2021, 24, 82–92. [Google Scholar] [CrossRef]
- Serrano-Pozo, A.; Li, H.; Li, Z.; Muñoz-Castro, C.; Jaisa-Aad, M.; Healey, M.A.; Welikovitch, L.A.; Jayakumar, R.; Bryant, A.G.; Noori, A.; et al. Astrocyte Transcriptomic Changes along the Spatiotemporal Progression of Alzheimer’s Disease. Nat. Neurosci. 2024, 27, 2384–2400. [Google Scholar] [CrossRef]
- Clayton, B.L.L.; Liddelow, S.A. Heterogeneity of Astrocyte Reactivity. Annu. Rev. Neurosci. 2025, 48, 231–249. [Google Scholar] [CrossRef]
- Allen, N.J.; Eroglu, C. Cell Biology of Astrocyte-Synapse Interactions. Neuron 2017, 96, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.-S.; Allen, N.J.; Eroglu, C. Astrocytes Control Synapse Formation, Function, and Elimination. Cold Spring Harb. Perspect. Biol. 2015, 7, a020370. [Google Scholar] [CrossRef]
- Takano, T.; Wallace, J.T.; Baldwin, K.T.; Purkey, A.M.; Uezu, A.; Courtland, J.L.; Soderblom, E.J.; Shimogori, T.; Maness, P.F.; Eroglu, C.; et al. Chemico-Genetic Discovery of Astrocytic Control of Inhibition in Vivo. Nature 2020, 588, 296–302. [Google Scholar] [CrossRef]
- Takano, T.; Soderling, S.H. Tripartite Synaptomics: Cell-Surface Proximity Labeling in Vivo. Neurosci. Res. 2021, 173, 14–21. [Google Scholar] [CrossRef]
- Cho, S.; Muthukumar, A.K.; Stork, T.; Coutinho-Budd, J.C.; Freeman, M.R. Focal Adhesion Molecules Regulate Astrocyte Morphology and Glutamate Transporters to Suppress Seizure-like Behavior. Proc. Natl. Acad. Sci. USA 2018, 115, 11316–11321. [Google Scholar] [CrossRef]
- Tan, C.X.; Bindu, D.S.; Hardin, E.J.; Sakers, K.; Baumert, R.; Ramirez, J.J.; Savage, J.T.; Eroglu, C. δ-Catenin Controls Astrocyte Morphogenesis via Layer-Specific Astrocyte-Neuron Cadherin Interactions. J. Cell Biol. 2023, 222, e202303138. [Google Scholar] [CrossRef]
- Nikolakopoulou, A.M.; Koeppen, J.; Garcia, M.; Leish, J.; Obenaus, A.; Ethell, I.M. Astrocytic Ephrin-B1 Regulates Synapse Remodeling Following Traumatic Brain Injury. ASN Neuro. 2016, 8, 1759091416630220. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, Y.; Qiao, Y.; Lin, J.; Lau, J.K.Y.; Fu, W.-Y.; Fu, A.K.Y.; Ip, N.Y. Astrocytic EphA4 Signaling Is Important for the Elimination of Excitatory Synapses in Alzheimer’s Disease. Proc. Natl. Acad. Sci. USA 2025, 122, e2420324122. [Google Scholar] [CrossRef]
- Tyzack, G.E.; Hall, C.E.; Sibley, C.R.; Cymes, T.; Forostyak, S.; Carlino, G.; Meyer, I.F.; Schiavo, G.; Zhang, S.-C.; Gibbons, G.M.; et al. A Neuroprotective Astrocyte State Is Induced by Neuronal Signal EphB1 but Fails in ALS Models. Nat. Commun. 2017, 8, 1164. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.-S.; Clarke, L.E.; Wang, G.X.; Stafford, B.K.; Sher, A.; Chakraborty, C.; Joung, J.; Foo, L.C.; Thompson, A.; Chen, C.; et al. Astrocytes Mediate Synapse Elimination through MEGF10 and MERTK Pathways. Nature 2013, 504, 394–400. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Kim, H.; Chung, W.-S.; Park, H. Selective Regulation of Corticostriatal Synapses by Astrocytic Phagocytosis. Nat. Commun. 2025, 16, 2504. [Google Scholar] [CrossRef]
- Pietiläinen, O.; Trehan, A.; Meyer, D.; Mitchell, J.; Tegtmeyer, M.; Valakh, V.; Gebre, H.; Chen, T.; Vartiainen, E.; Farhi, S.L.; et al. Astrocytic Cell Adhesion Genes Linked to Schizophrenia Correlate with Synaptic Programs in Neurons. Cell Rep. 2023, 42, 111988. [Google Scholar] [CrossRef] [PubMed]
- Ling, E.; Nemesh, J.; Goldman, M.; Kamitaki, N.; Reed, N.; Handsaker, R.E.; Genovese, G.; Vogelgsang, J.S.; Gerges, S.; Kashin, S.; et al. A Concerted Neuron-Astrocyte Program Declines in Ageing and Schizophrenia. Nature 2024, 627, 604–611. [Google Scholar] [CrossRef]
- Suzuki, A.; Stern, S.A.; Bozdagi, O.; Huntley, G.W.; Walker, R.H.; Magistretti, P.J.; Alberini, C.M. Astrocyte-Neuron Lactate Transport Is Required for Long-Term Memory Formation. Cell 2011, 144, 810–823. [Google Scholar] [CrossRef]
- Martin-Fernandez, M.; Jamison, S.; Robin, L.M.; Zhao, Z.; Martin, E.D.; Aguilar, J.; Benneyworth, M.A.; Marsicano, G.; Araque, A. Synapse-Specific Astrocyte Gating of Amygdala-Related Behavior. Nat. Neurosci. 2017, 20, 1540–1548. [Google Scholar] [CrossRef] [PubMed]
- Brazhe, A.; Verisokin, A.; Verveyko, D.; Postnov, D. Astrocytes: New Evidence, New Models, New Roles. Biophys. Rev. 2023, 15, 1303–1333. [Google Scholar] [CrossRef] [PubMed]
- Caudle, R.M. Memory in Astrocytes: A Hypothesis. Theor. Biol. Med. Model. 2006, 3, 2. [Google Scholar] [CrossRef]
- Kol, A.; Goshen, I. The Memory Orchestra: The Role of Astrocytes and Oligodendrocytes in Parallel to Neurons. Curr. Opin. Neurobiol. 2021, 67, 131–137. [Google Scholar] [CrossRef]
- Giaume, C.; Koulakoff, A.; Roux, L.; Holcman, D.; Rouach, N. Astroglial Networks: A Step Further in Neuroglial and Gliovascular Interactions. Nat. Rev. Neurosci. 2010, 11, 87–99. [Google Scholar] [CrossRef]
- Parnis, J.; Montana, V.; Delgado-Martinez, I.; Matyash, V.; Parpura, V.; Kettenmann, H.; Sekler, I.; Nolte, C. Mitochondrial Exchanger NCLX Plays a Major Role in the Intracellular Ca2+ Signaling, Gliotransmission, and Proliferation of Astrocytes. J. Neurosci. 2013, 33, 7206–7219. [Google Scholar]
- Jackson, J.G.; Robinson, M.B. Reciprocal Regulation of Mitochondrial Dynamics and Calcium Signaling in Astrocyte Processes. J. Neurosci. 2015, 35, 15199–15213. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Rodríguez, J.J.; Parpura, V. Calcium Signalling in Astroglia. Mol. Cell Endocrinol. 2012, 353, 45–56. [Google Scholar] [CrossRef]
- De Pittà, M.; Brunel, N. Multiple Forms of Working Memory Emerge from Synapse-Astrocyte Interactions in a Neuron-Glia Network Model. Proc. Natl. Acad. Sci. USA 2022, 119, e2207912119. [Google Scholar]
- Krotov, D.; Hopfield, J. Large Associative Memory Problem in Neurobiology and Machine Learning. arXiv 2020. [Google Scholar] [CrossRef]
- Krotov, D.; Hopfield, J.J. Dense Associative Memory for Pattern Recognition. arXiv 2016. [Google Scholar] [CrossRef]
- Kozachkov, L.; Kastanenka, K.V.; Krotov, D. Building Transformers from Neurons and Astrocytes. Proc. Natl. Acad. Sci. USA 2023, 120, e2219150120. [Google Scholar] [CrossRef]
- Vaswani, A.; Shazeer, N.; Parmar, N.; Uszkoreit, J.; Jones, L.; Gomez, A.N.; Kaiser, L.; Polosukhin, I. Attention Is All You Need. arXiv 2017. [Google Scholar] [CrossRef]
- Ramsauer, H.; Schäfl, B.; Lehner, J.; Seidl, P.; Widrich, M.; Adler, T.; Gruber, L.; Holzleitner, M.; Pavlović, M.; Sandve, G.K.; et al. Hopfield Networks Is All You Need. arXiv 2020. [Google Scholar] [CrossRef]
- Orellana, J.A.; Sáez, P.J.; Shoji, K.F.; Schalper, K.A.; Palacios-Prado, N.; Velarde, V.; Giaume, C.; Bennett, M.V.L.; Sáez, J.C. Modulation of Brain Hemichannels and Gap Junction Channels by Pro-Inflammatory Agents and Their Possible Role in Neurodegeneration. Antioxid. Redox Signal. 2009, 11, 369–399. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Yu, H.-X.; Sun, M.-L.; Wang, Y.; Xi, W.; Yu, Y.-Q. Astrocyte-Restricted Disruption of Connexin-43 Impairs Neuronal Plasticity in Mouse Barrel Cortex. Eur. J. Neurosci. 2014, 39, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Duarte, A.; Roberts, A.J.; Ouyang, K.; Sejnowski, T.J. Impairments in Remote Memory Caused by the Lack of Type 2 IP3 Receptors. Glia 2019, 67, 1976–1989. [Google Scholar] [CrossRef]
- Paukert, M.; Agarwal, A.; Cha, J.; Doze, V.A.; Kang, J.U.; Bergles, D.E. Norepinephrine Controls Astroglial Responsiveness to Local Circuit Activity. Neuron 2014, 82, 1263–1270. [Google Scholar] [CrossRef]
- Araque, A.; Martín, E.D.; Perea, G.; Arellano, J.I.; Buño, W. Synaptically Released Acetylcholine Evokes Ca2+ Elevations in Astrocytes in Hippocampal Slices. J. Neurosci. 2002, 22, 2443–2450. [Google Scholar] [CrossRef] [PubMed]
- Werner, C.; Sauer, M.; Geis, C. Super-Resolving Microscopy in Neuroscience. Chem. Rev. 2021, 121, 11971–12015. [Google Scholar] [CrossRef]
- Ito, Y.; Nagamoto, S.; Takano, T. Synaptic Proteomics Decode Novel Molecular Landscape in the Brain. Front. Mol. Neurosci. 2024, 17, 1361956. [Google Scholar] [CrossRef] [PubMed]
- Matsubayashi, J.; Takano, T. Proximity Labeling Uncovers the Synaptic Proteome under Physiological and Pathological Conditions. Front. Cell Neurosci. 2025, 19, 1638627. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).