
Comprehensive Analysis Identifies THEMIS2 as a Potential Prognostic and Immunological Biomarker in Glioblastoma

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Gene Expression Analysis
2.3. Functional Enrichment Analysis
2.4. Immune Characteristics Analysis
2.5. Single-Cell Transcriptome Analysis
2.6. Macrophage Subtype Analysis
2.7. Intercellular Communication Analysis
2.8. Statistical Analysis
3. Results
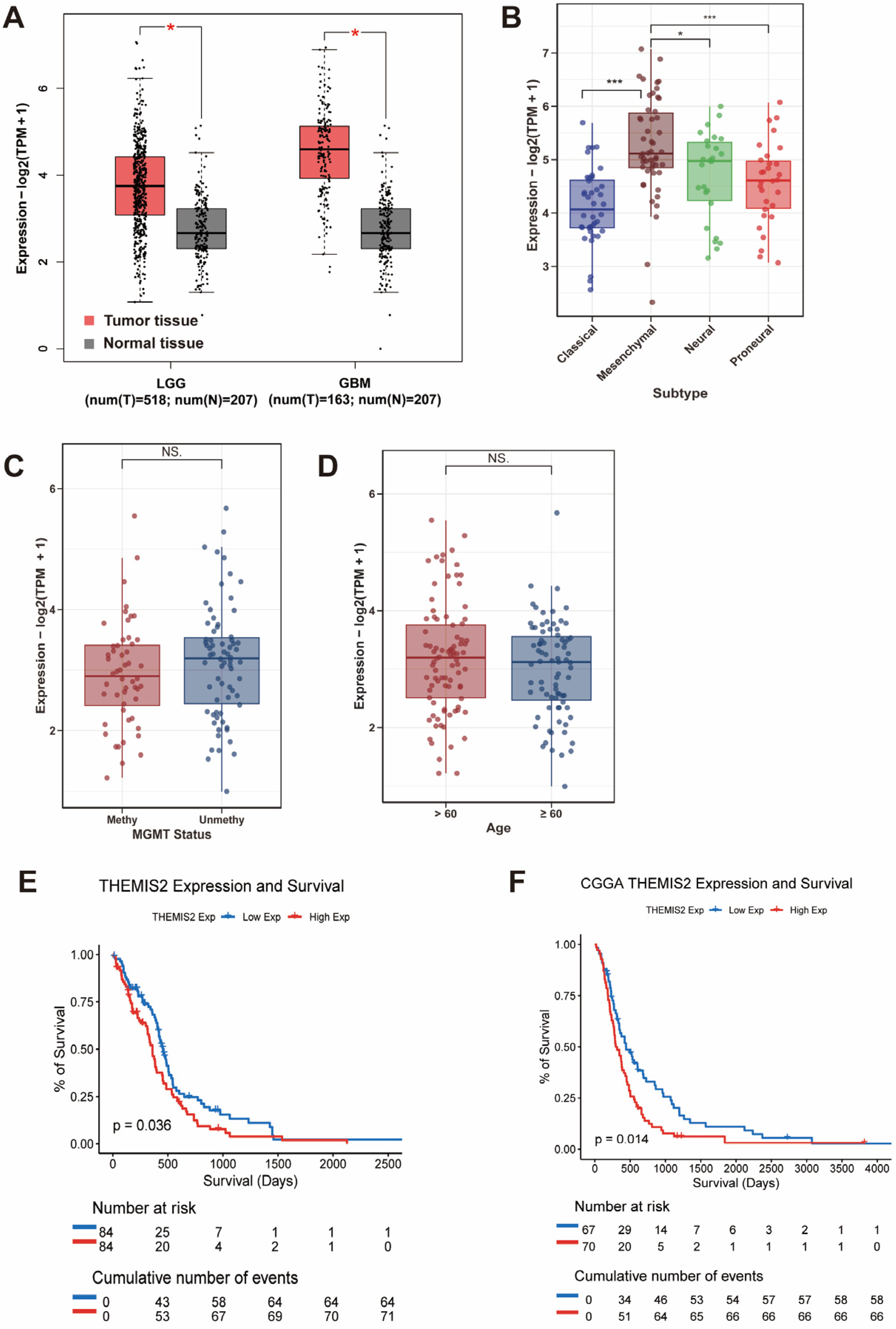
3.1. THEMIS2 Expression in GBM Clinical Subgroups and Its Prognostic Association with Survival
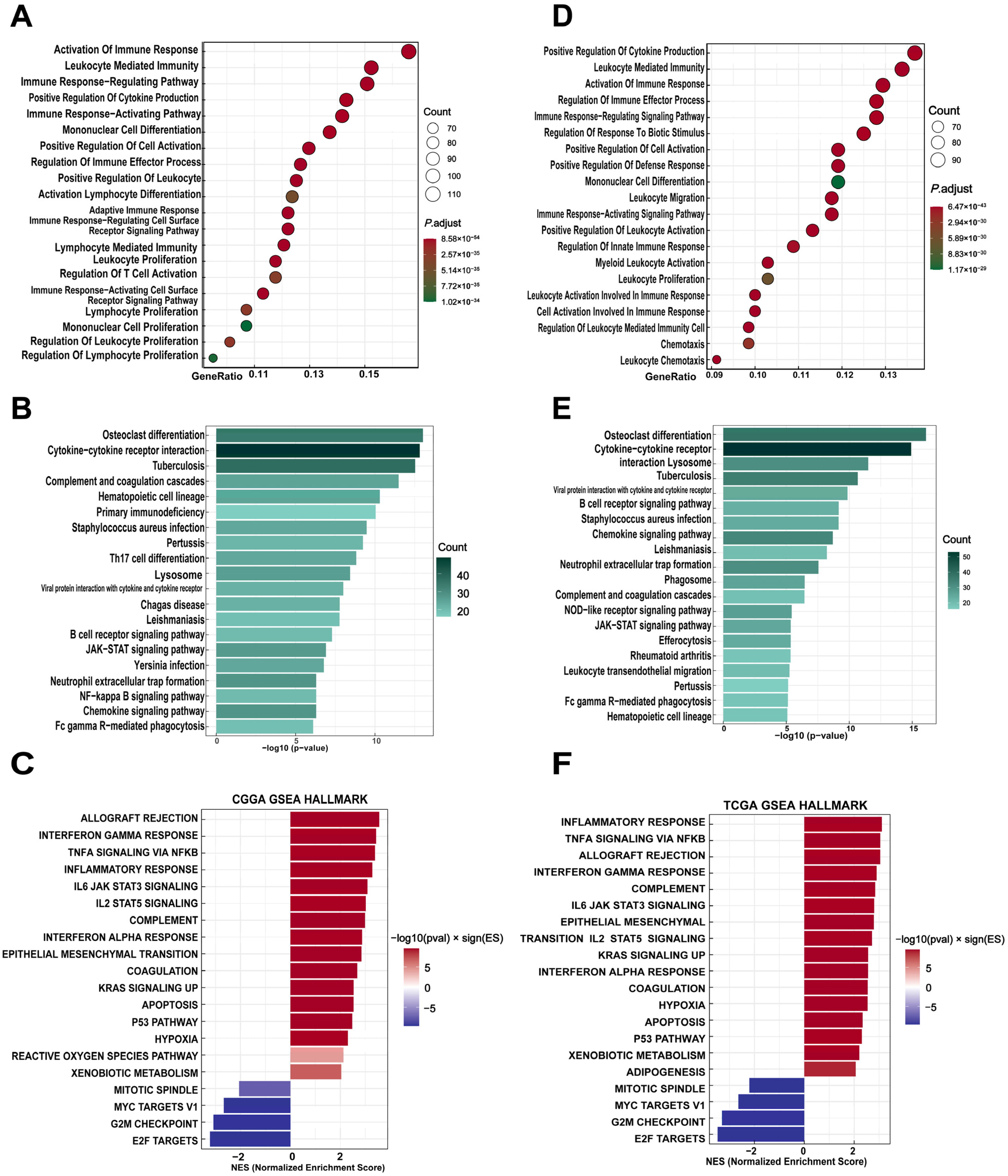
3.2. Functional Enrichment of THEMIS2-Associated Genes
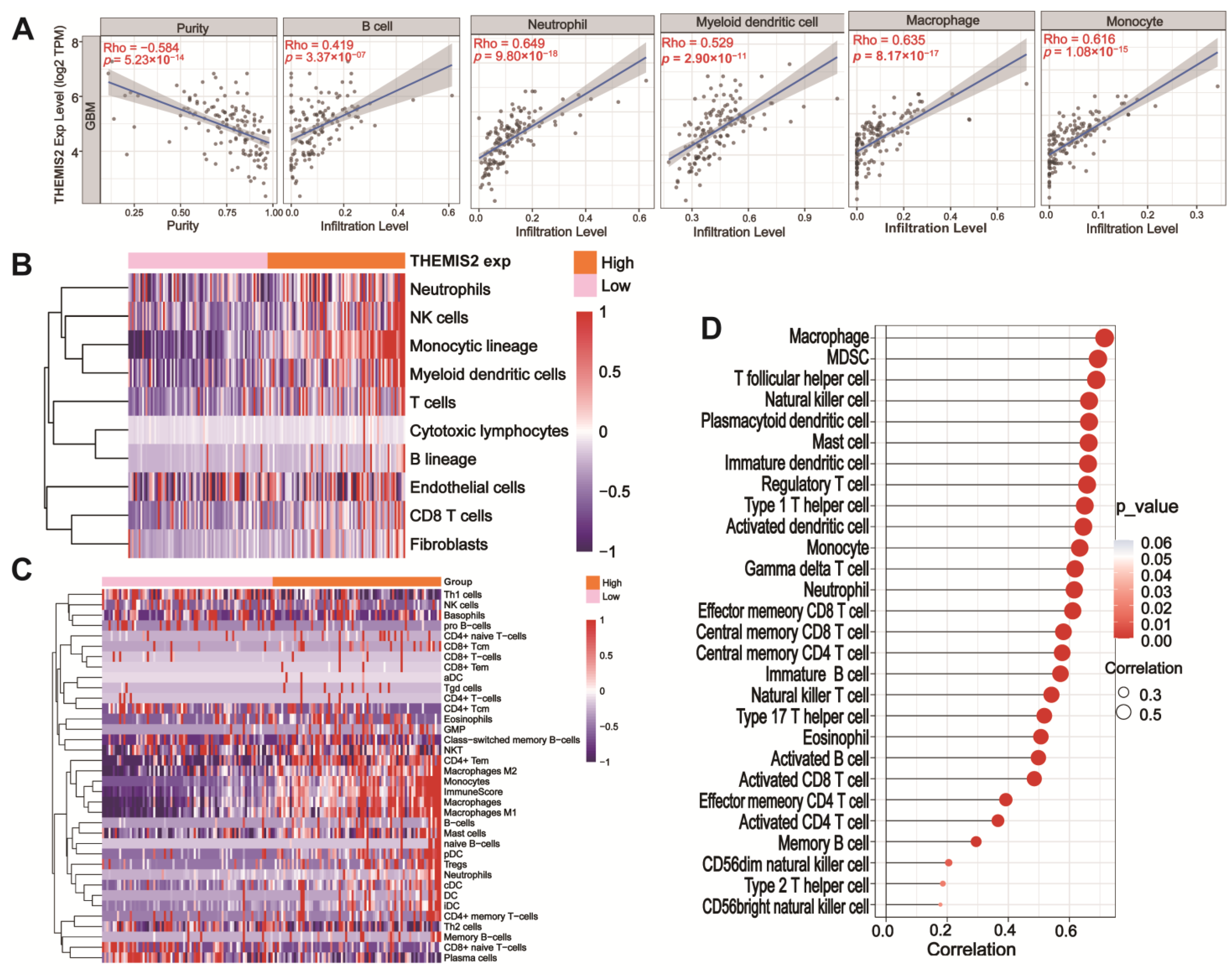
3.3. Immune Infiltration and THEMIS2 Correlation
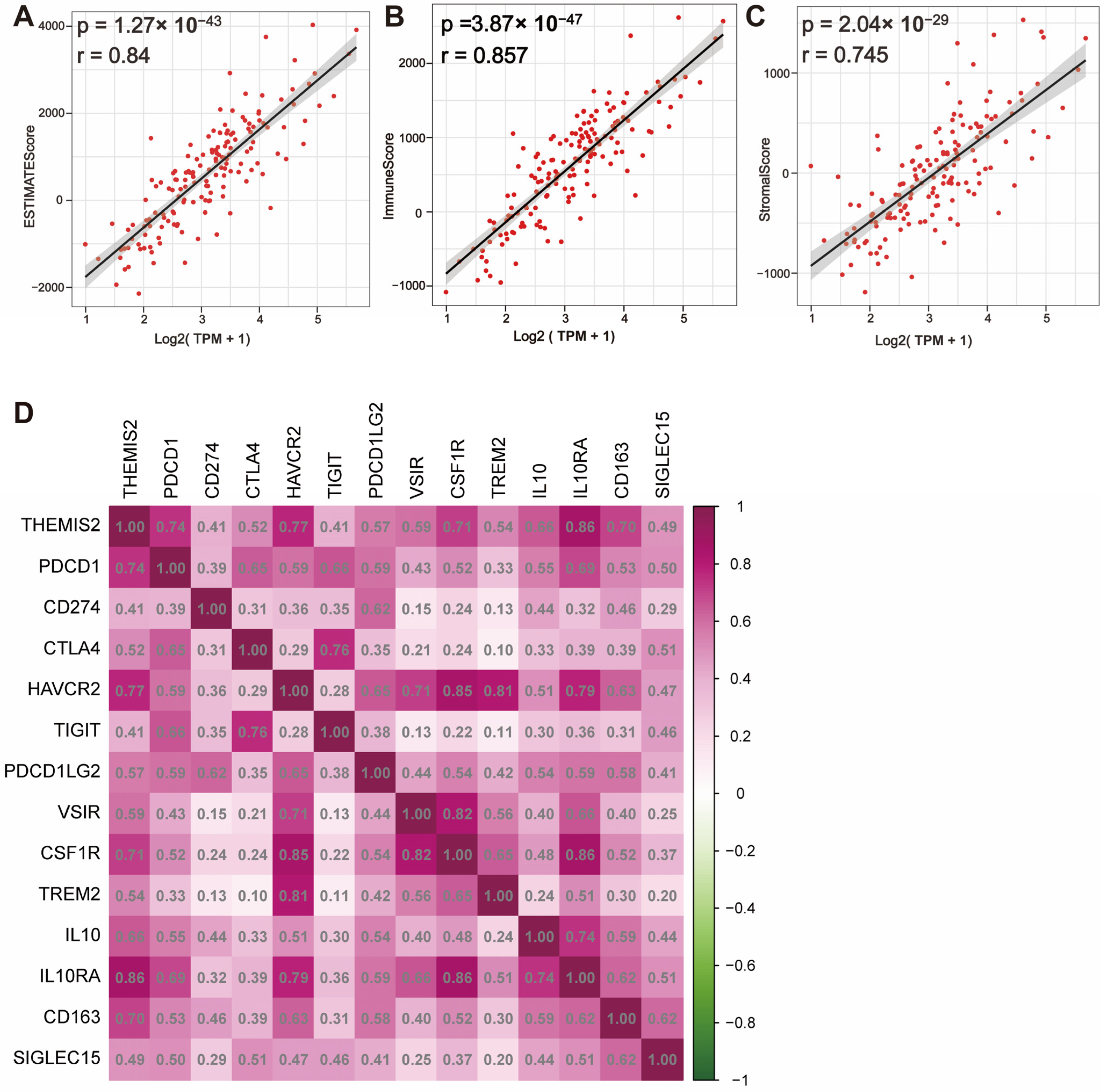
3.4. Correlation Between THEMIS2 Expression, Immune Microenvironment, and Immune Checkpoint Molecules in GBM
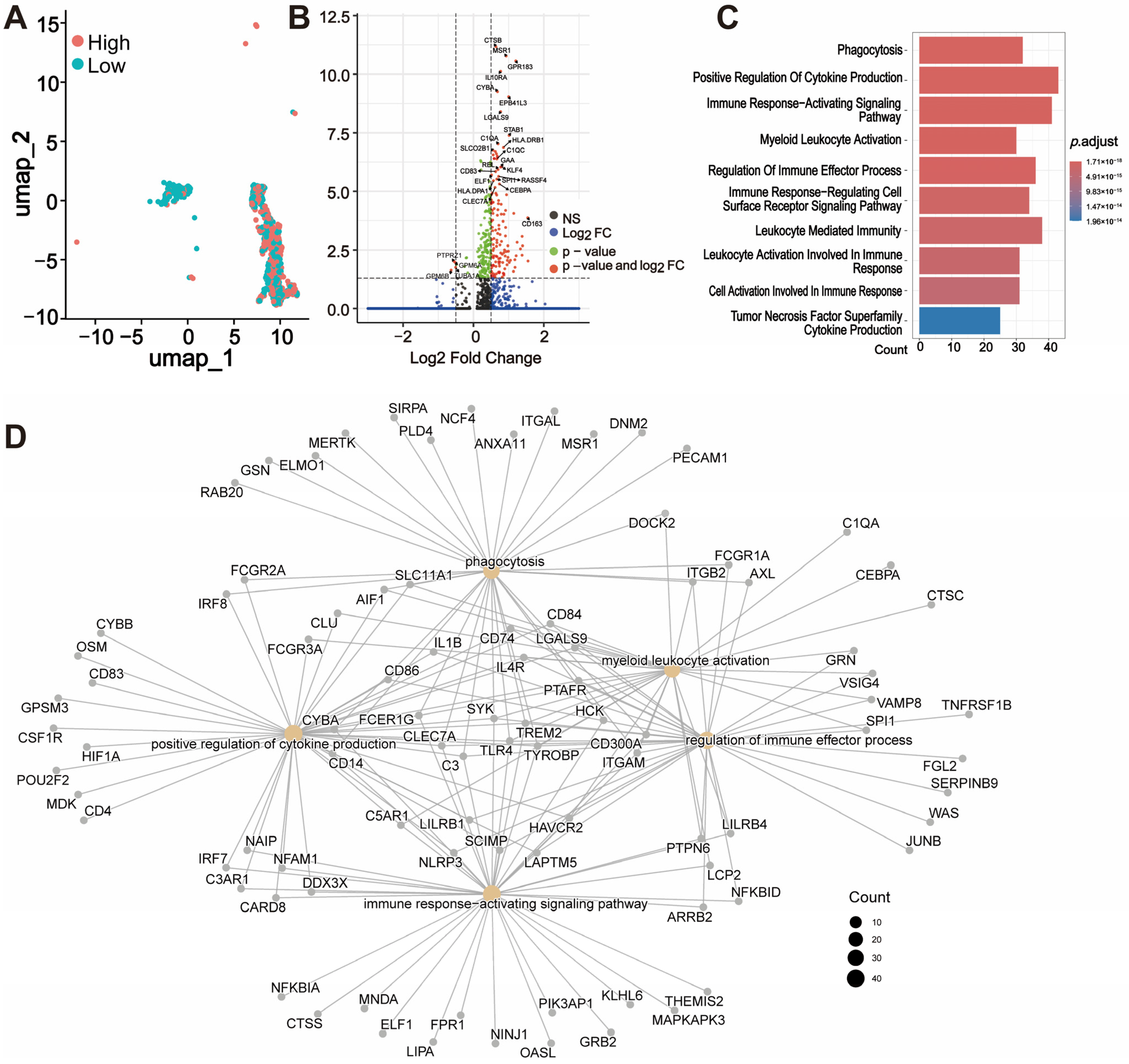
3.5. Single-Cell Analysis of THEMIS2 Expression in GBM Microenvironment
3.6. Macrophage Subtype Analysis Based on THEMIS2 Expression
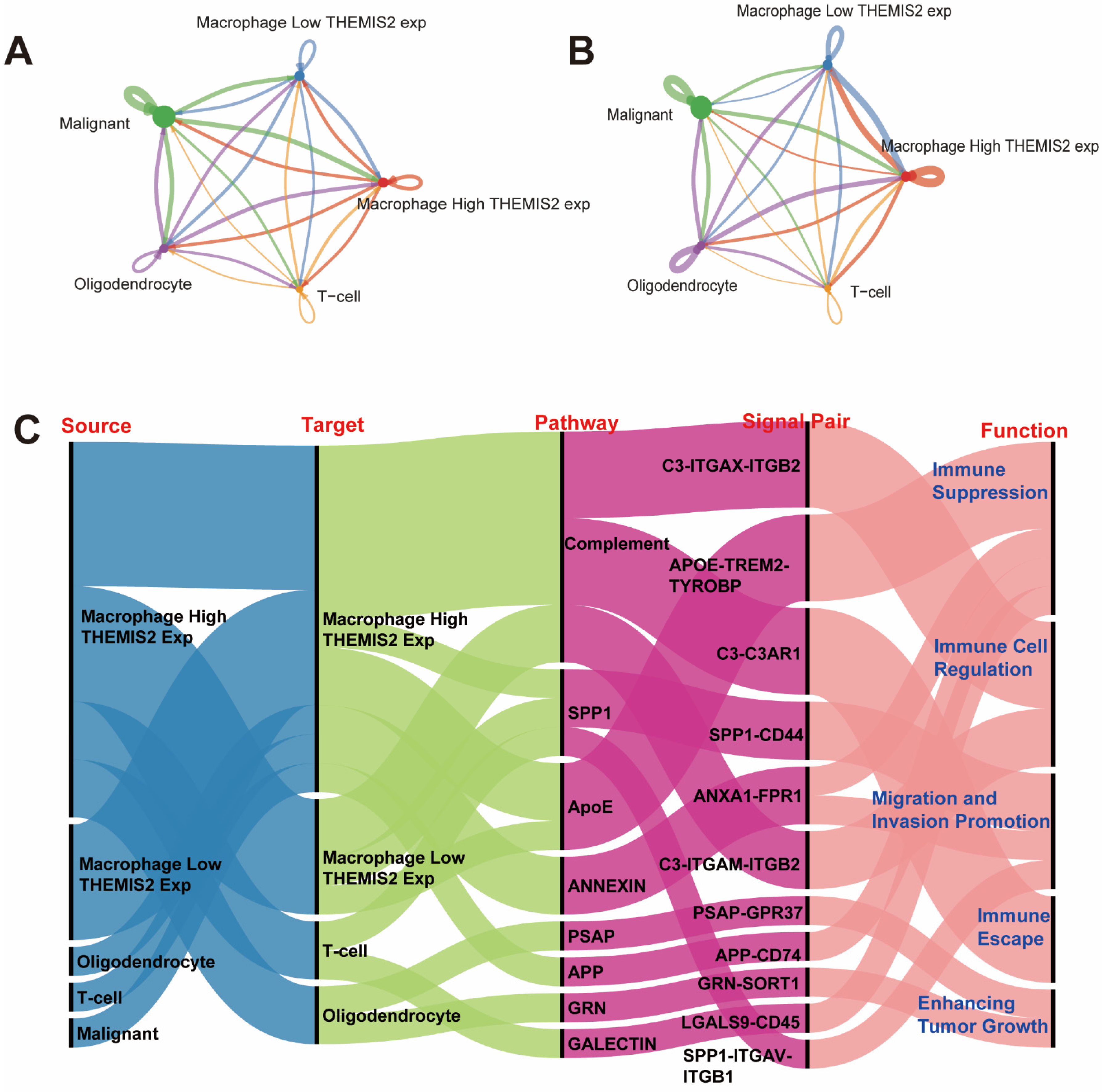
3.7. Intercellular Communication
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schaff, L.R.; Mellinghoff, I.K. Glioblastoma and Other Primary Brain Malignancies in Adults: A Review. JAMA 2023, 329, 574–587. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Berglund, A.E.; Etame, A.B. The Impact of Epigenetic Modifications on Adaptive Resistance Evolution in Glioblastoma. Int. J. Mol. Sci. 2021, 22, 8324. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.C.; Ashley, D.M.; López, G.Y.; Malinzak, M.; Friedman, H.S.; Khasraw, M. Management of glioblastoma: State of the art and future directions. CA Cancer J. Clin. 2020, 70, 299–312. [Google Scholar] [CrossRef]
- Omuro, A.; DeAngelis, L.M. Glioblastoma and other malignant gliomas: A clinical review. JAMA 2013, 310, 1842–1850. [Google Scholar] [CrossRef]
- Kloosterman, D.J.; Erbani, J.; Boon, M.; Farber, M.; Handgraaf, S.M.; Ando-Kuri, M.; Sánchez-López, E.; Fontein, B.; Mertz, M.; Nieuwland, M.; et al. Macrophage-mediated myelin recycling fuels brain cancer malignancy. Cell 2024, 187, 5336–5356. [Google Scholar] [CrossRef]
- Karimi, E.; Yu, M.W.; Maritan, S.M.; Perus, L.J.; Rezanejad, M.; Sorin, M.; Dankner, M.; Fallah, P.; Doré, S.; Zuo, D.; et al. Single-cell spatial immune landscapes of primary and metastatic brain tumours. Nature 2023, 614, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Q.; Liu, Y.; Wang, H.; Lin, Z.; Sun, L.; Liu, Y.; Lyu, Y.; Chen, L.; Yang, H.; Mao, Y. LILRB3 suppresses immunity in glioma and is associated with poor prognosis. Clin. Transl. Med. 2023, 13, e1396. [Google Scholar] [CrossRef] [PubMed]
- Marron, T.U.; Guerriero, J.L. SIGLEC9 tips the myeloid balance in glioblastoma. Nat. Cancer 2023, 4, 1217–1219. [Google Scholar] [CrossRef] [PubMed]
- Amin, T.; Hossain, A.; Jerin, N.; Mahmud, I.; Rahman, M.A.; Rafiqul Islam, S.M.; Islam, S.B.U. Immunoediting Dynamics in Glioblastoma: Implications for Immunotherapy Approaches. Cancer Control 2024, 31, 10732748241290067. [Google Scholar] [CrossRef]
- Ijaz, M.; Ullah, Z.; Aslam, B.; Khrshid, M.; Chen, P.; Guo, B. From promise to progress: The dynamic landscape of glioblastoma immunotherapy. Drug Discov. Today 2024, 29, 104188. [Google Scholar] [CrossRef]
- Ott, M.; Prins, R.M.; Heimberger, A.B. The immune landscape of common CNS malignancies: Implications for immunotherapy. Nat. Rev. Clin. Oncol. 2021, 18, 729–744. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Yen, J.H.; Sung, Y.W.; Tung, S.L.; Chen, P.M.; Chu, P.Y.; Shih, Y.C.; Chi, H.C.; Huang, Y.C.; Huang, S.J.; et al. Novel function of THEMIS2 in the enhancement of cancer stemness and chemoresistance by releasing PTP1B from MET. Oncogene 2022, 41, 997–1010. [Google Scholar] [CrossRef]
- Deborah, E.A.; Nabekura, T.; Shibuya, K.; Shibuya, A. THEMIS2 Impairs Antitumor Activity of NK Cells by Suppressing Activating NK Receptor Signaling. J. Immunol. 2024, 212, 1819–1828. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, K.N.; Wang, Q.; Li, G.; Zeng, F.; Zhang, Y.; Wu, F.; Chai, R.; Wang, Z.; Zhang, C.; et al. Chinese Glioma Genome Atlas (CGGA): A Comprehensive Resource with Functional Genomic Data from Chinese Glioma Patients. Genom. Proteom. Bioinform. 2021, 19, 1–12. [Google Scholar] [CrossRef]
- Li, G.; Li, L.; Li, Y.; Qian, Z.; Wu, F.; He, Y.; Jiang, H.; Li, R.; Wang, D.; Zhai, Y.; et al. An MRI radiomics approach to predict survival and tumour-infiltrating macrophages in gliomas. Brain 2022, 145, 1151–1161. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Becht, E.; Giraldo, N.A.; Lacroix, L.; Buttard, B.; Elarouci, N.; Petitprez, F.; Selves, J.; Laurent-Puig, P.; Sautès-Fridman, C.; Fridman, W.H.; et al. Estimating the population abundance of tissue-infiltrating immune and stromal cell populations using gene expression. Genome Biol. 2016, 17, 218. [Google Scholar]
- Aran, D.; Hu, Z.; Butte, A.J. xCell: Digitally portraying the tissue cellular heterogeneity landscape. Genome Biol. 2017, 18, 220. [Google Scholar] [CrossRef] [PubMed]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, K.; Shahmoradgoli, M.; Martínez, E.; Vegesna, R.; Kim, H.; Torres-Garcia, W.; Treviño, V.; Shen, H.; Laird, P.W.; Levine, D.A.; et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat. Commun. 2013, 4, 2612. [Google Scholar] [CrossRef]
- Hao, Y.; Stuart, T.; Kowalski, M.H.; Choudhary, S.; Hoffman, P.; Hartman, A.; Srivastava, A.; Molla, G.; Madad, S.; Fernandez-Granda, C.; et al. Dictionary learning for integrative, multimodal and scalable single-cell analysis. Nat. Biotechnol. 2024, 42, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Neftel, C.; Laffy, J.; Filbin, M.G.; Hara, T.; Shore, M.E.; Rahme, G.J.; Richman, A.R.; Silverbush, D.; Shaw, M.L.; Hebert, C.M.; et al. An Integrative Model of Cellular States, Plasticity, and Genetics for Glioblastoma. Cell 2019, 178, 835–849.e21. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [PubMed]
- Varn, F.S.; Johnson, K.C.; Martinek, J.; Huse, J.T.; Nasrallah, M.P.; Wesseling, P.; Cooper, L.A.; Malta, T.M.; Wade, T.E.; Sabedot, T.S.; et al. Glioma progression is shaped by genetic evolution and microenvironment interactions. Cell 2022, 185, 2184–2199. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Berglund, A.E.; Macaulay, R.J.; Etame, A.B. The Role of Mesenchymal Reprogramming in Malignant Clonal Evolution and Intra-Tumoral Heterogeneity in Glioblastoma. Cells 2024, 13, 942. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Hsieh, K.; Cherry, D.R.; Nehlsen, A.D.; Resende Salgado, L.; Lazarev, S.; Sindhu, K.K. Immune Escape in Glioblastoma: Mechanisms of Action and Implications for Immune Checkpoint Inhibitors and CAR T-Cell Therapy. Biology 2023, 12, 1528. [Google Scholar] [CrossRef]
- Jackson, C.M.; Choi, J.; Lim, M. Mechanisms of immunotherapy resistance: Lessons from glioblastoma. Nat. Immunol. 2019, 20, 1100–1109. [Google Scholar] [CrossRef] [PubMed]
- Sa, J.K.; Chang, N.; Lee, H.W.; Cho, H.J.; Ceccarelli, M.; Cerulo, L.; Yin, J.; Kim, S.S.; Caruso, F.P.; Lee, M.; et al. Transcriptional regulatory networks of tumor-associated macrophages that drive malignancy in mesenchymal glioblastoma. Genome Biol. 2020, 21, 216. [Google Scholar] [CrossRef]
- Sevenich, L. Turning “Cold” Into “Hot” Tumors-Opportunities and Challenges for Radio-Immunotherapy Against Primary and Metastatic Brain Cancers. Front. Oncol. 2019, 9, 163. [Google Scholar] [CrossRef]
- Tomaszewski, W.; Sanchez-Perez, L.; Gajewski, T.F.; Sampson, J.H. Brain Tumor Microenvironment and Host State: Implications for Immunotherapy. Clin. Cancer Res. 2019, 25, 4202–4210. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, P.; Wang, L.; Wu, J.; Tian, Y.; Chen, C.; Li, D.; Yao, Z.; Chen, R.; Xiang, G.; Gong, J.; et al. Overcoming cold tumors: A combination strategy of immune checkpoint inhibitors. Front. Immunol. 2024, 15, 1344272. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Wu, Q.; Berglund, A.E.; Macaulay, R.J.; Etame, A.B. Comprehensive Analysis Identifies THEMIS2 as a Potential Prognostic and Immunological Biomarker in Glioblastoma. Cells 2025, 14, 66. https://doi.org/10.3390/cells14020066
Chen J, Wu Q, Berglund AE, Macaulay RJ, Etame AB. Comprehensive Analysis Identifies THEMIS2 as a Potential Prognostic and Immunological Biomarker in Glioblastoma. Cells. 2025; 14(2):66. https://doi.org/10.3390/cells14020066
Chicago/Turabian StyleChen, Jianan, Qiong Wu, Anders E. Berglund, Robert J. Macaulay, and Arnold B. Etame. 2025. "Comprehensive Analysis Identifies THEMIS2 as a Potential Prognostic and Immunological Biomarker in Glioblastoma" Cells 14, no. 2: 66. https://doi.org/10.3390/cells14020066
APA StyleChen, J., Wu, Q., Berglund, A. E., Macaulay, R. J., & Etame, A. B. (2025). Comprehensive Analysis Identifies THEMIS2 as a Potential Prognostic and Immunological Biomarker in Glioblastoma. Cells, 14(2), 66. https://doi.org/10.3390/cells14020066