
The Antitumor Role of Incomptine A in a Breast Cancer Murine Model: Impairment of Hexokinase II Expression and Apoptosis Induction

 ,
,  ,
,  , ,
, ,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Incomptine A
2.2. Cell Culture
2.3. Laboratory Animals
2.4. Acute Oral Toxicity
2.5. Murine Model of Breast Cancer
2.6. Antitumor Activity
2.7. Toxicity and Safety Profile of IA in the Murine BC Model
2.8. Histopathological Analysis
2.9. Western Blot
2.10. Annexin V/Ghost Red Staining
2.11. Mitochondrial Respiratory Complex I Activity Assay
2.12. Statistical Analysis
3. Results
3.1. Acute Oral Toxicity of Incomptine A
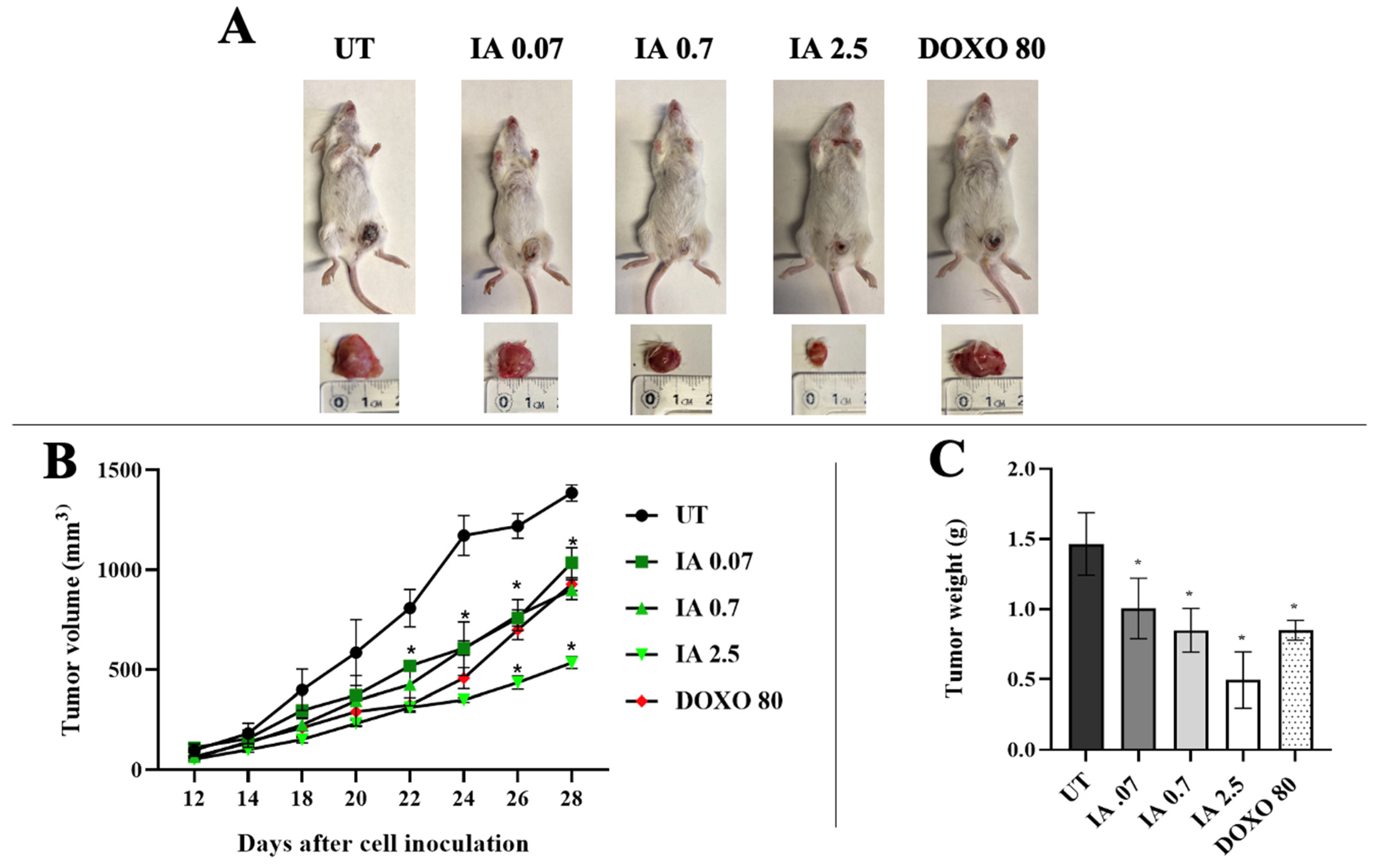
3.2. Antitumor Activity of Incomptine A
3.3. Histopathology Analysis of Tumors
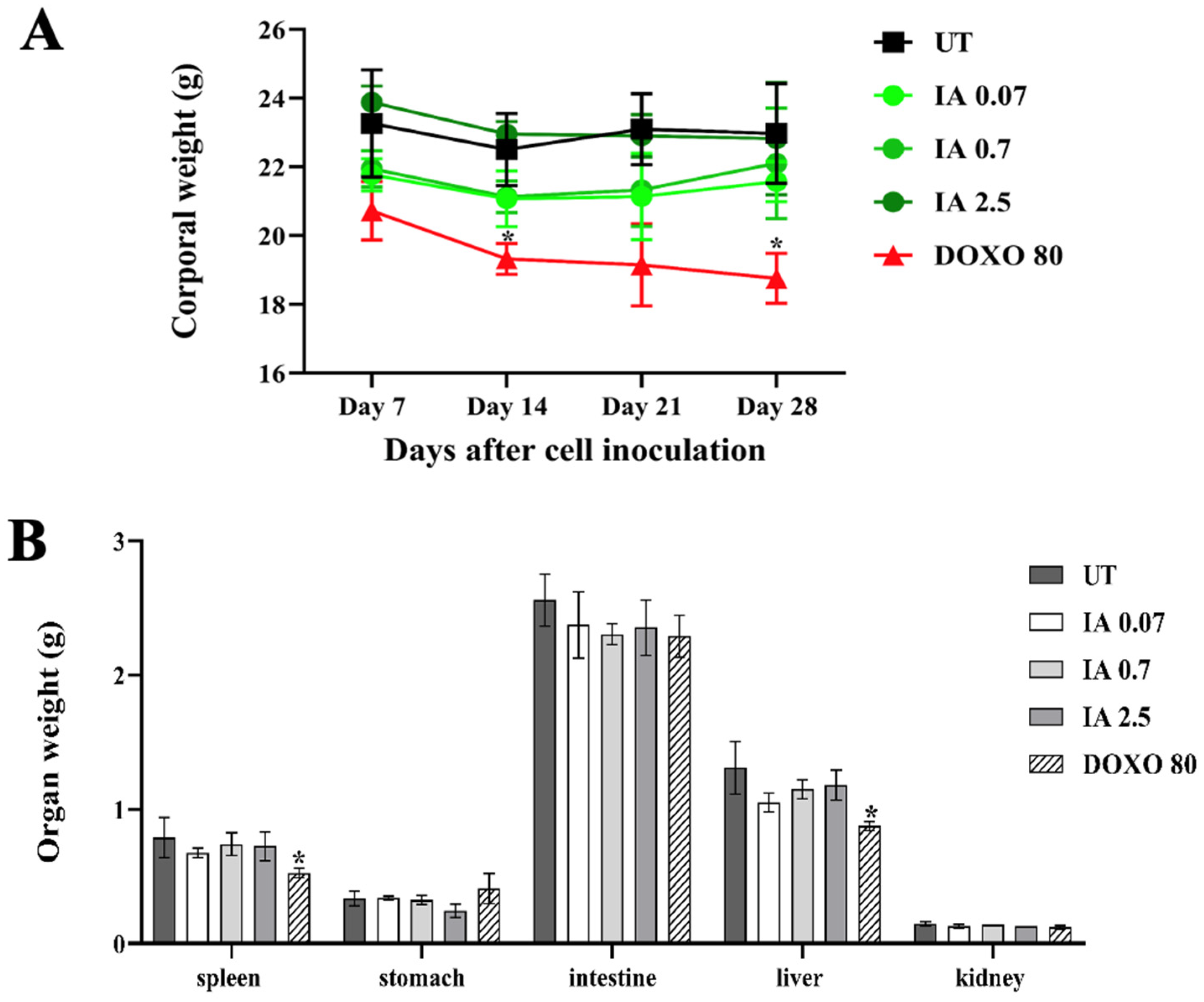
3.4. Toxicity of Incomptine A Treatment in an In Vivo Mouse Model
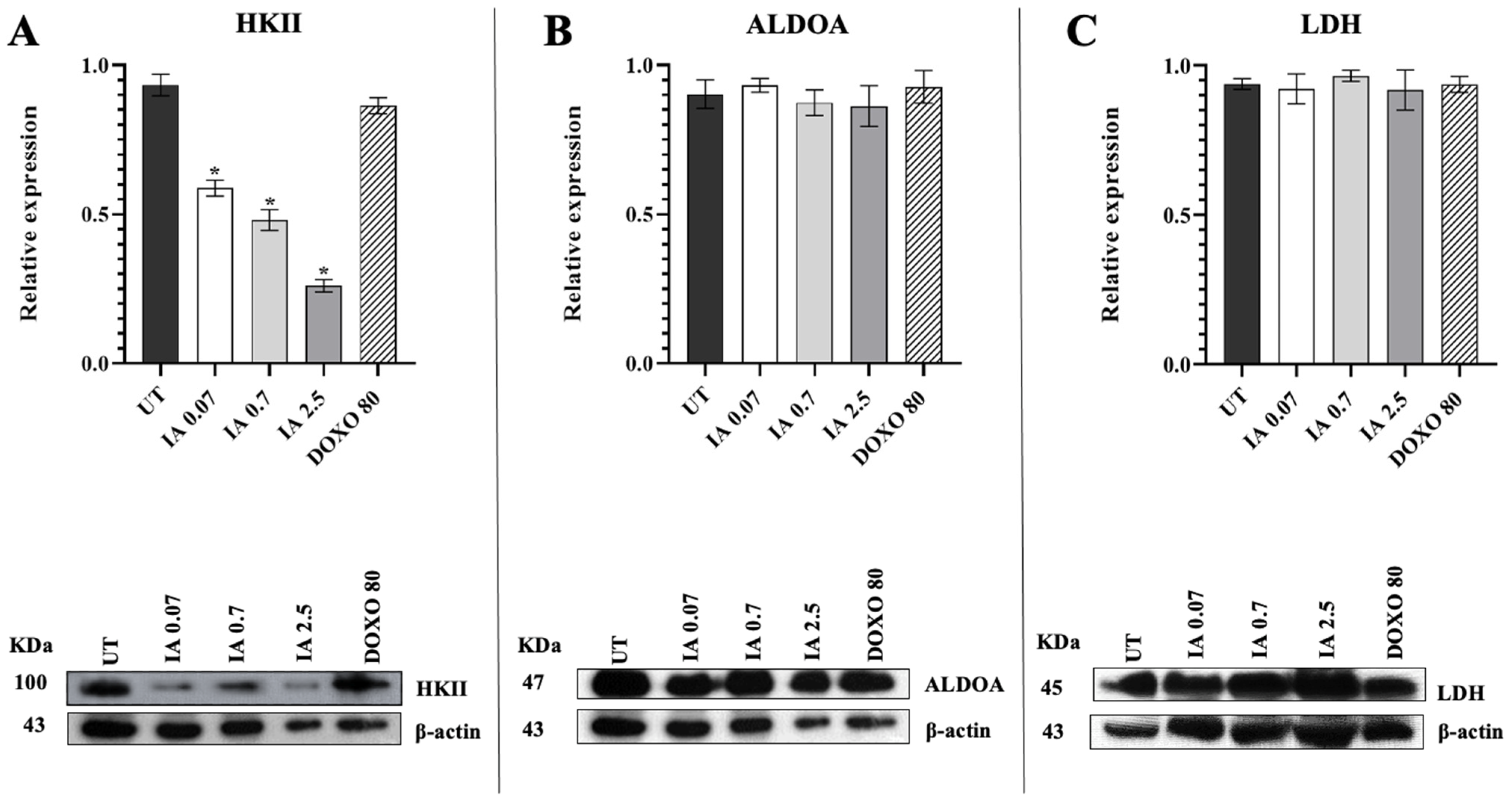
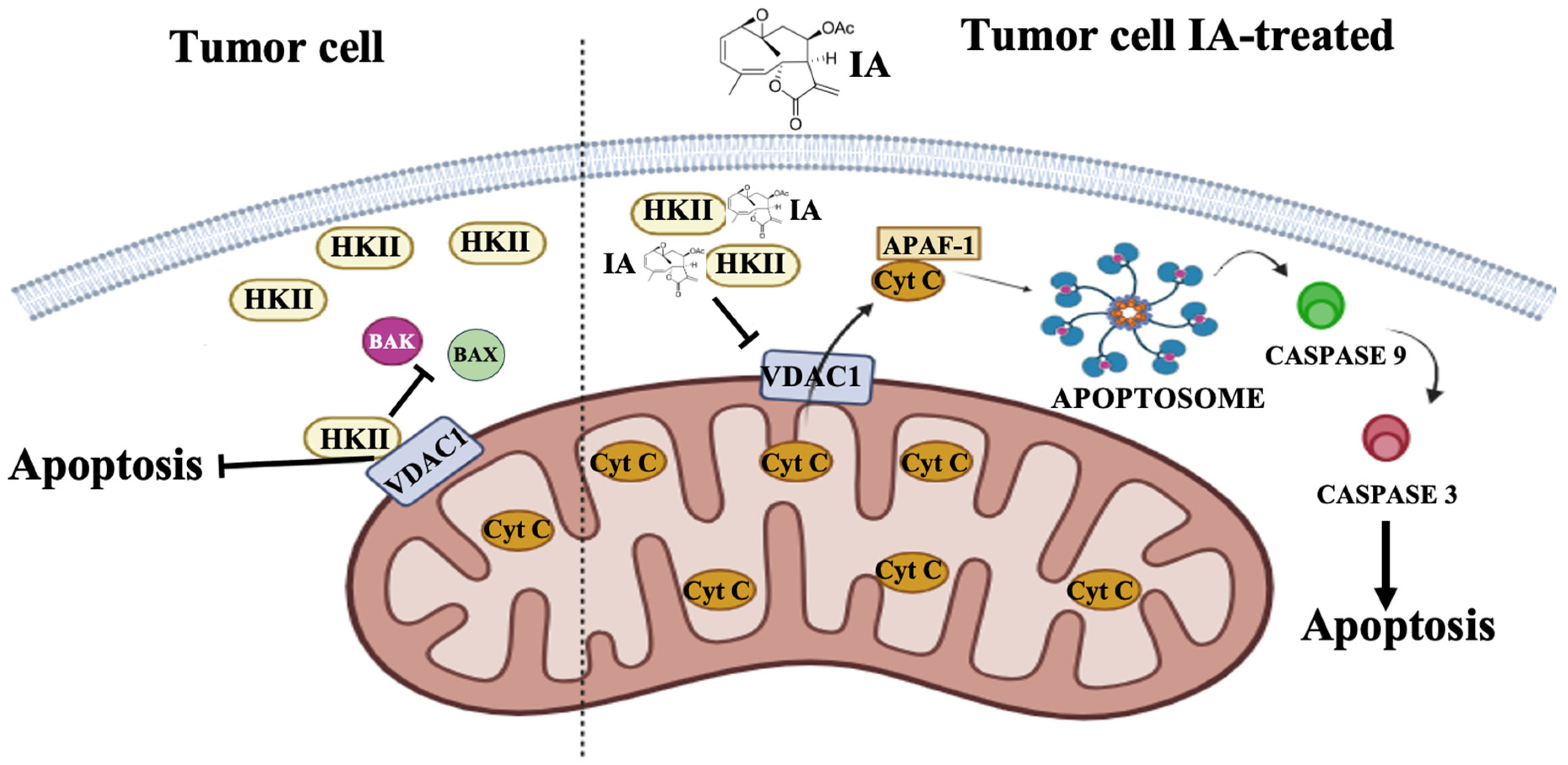
3.5. Treatment with Incomptine A Decreases HKII Expression in Mammary Tumors of Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mice | Treatment (mg/kg) | SGOT | SGPT |
|---|---|---|---|
| Healthy tumor-free | untreated | 250.2 ± 11 | 49 ± 5.80 |
| Mammary tumor | untreated | 970.5 ± 29 * | 445 ± 17 * |
| 0.07 | 213.6 ± 10 | 41.6 ± 6 | |
| IA | 0.70 | 244.3 ± 4 | 43.3 ± 9 |
| 2.50 | 264.6 ± 4 | 44.6 ± 6 | |
| DOXO | 80.00 | 478.6 ± 4 * | 54.6 ± 9 |
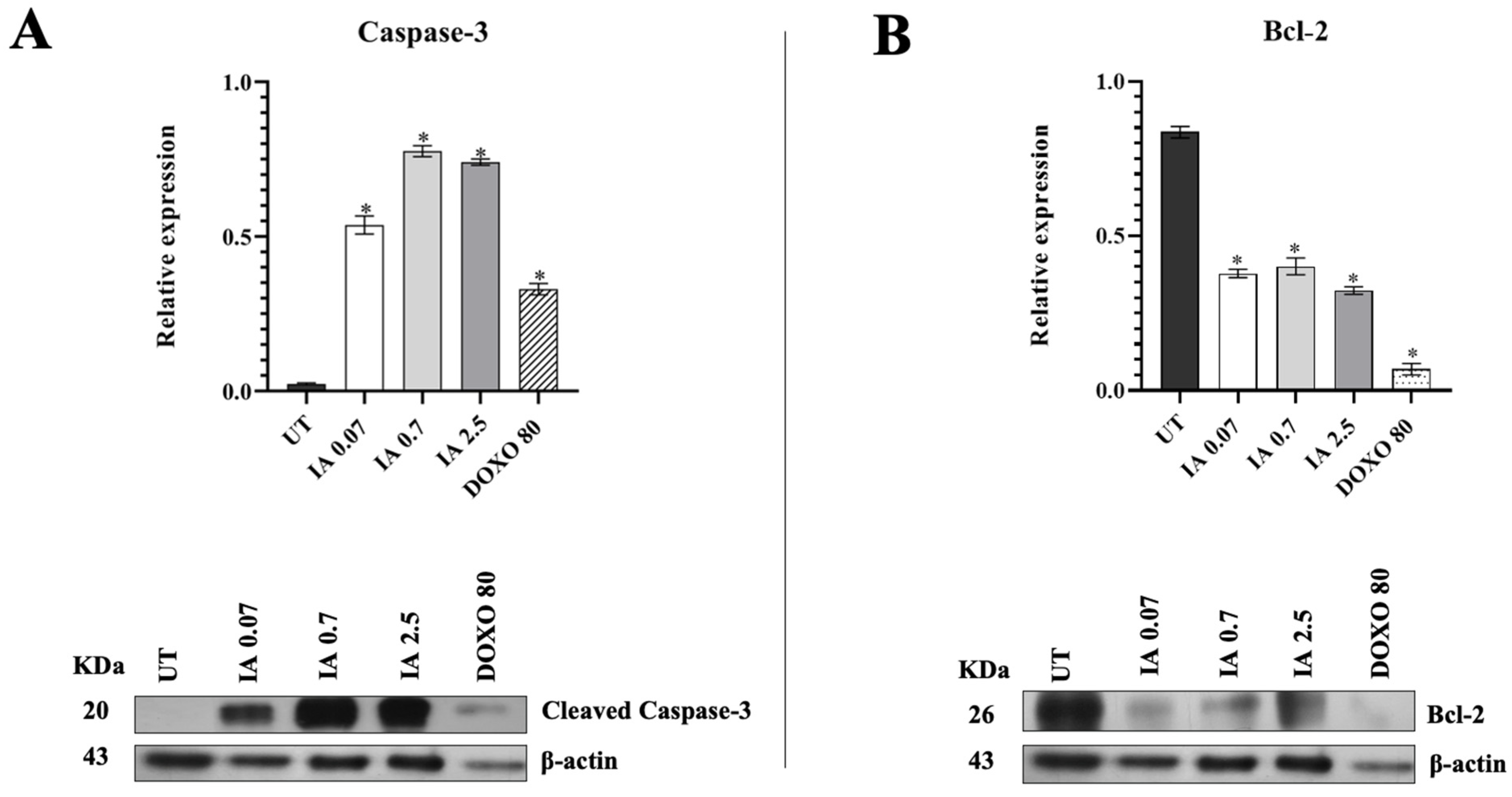
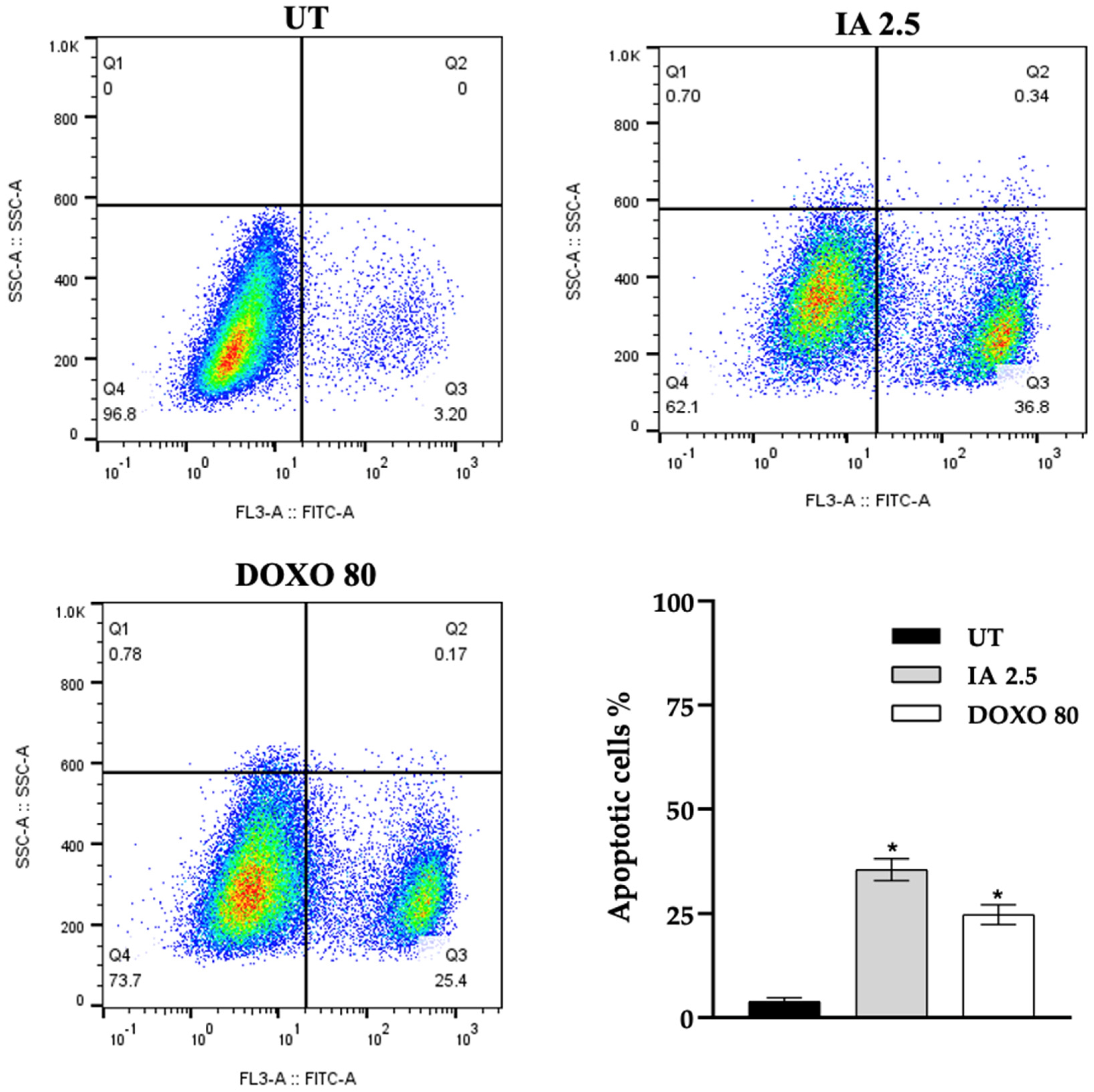
3.6. Incomptine A Induces Apoptosis in Breast Cancer Tumor Cells
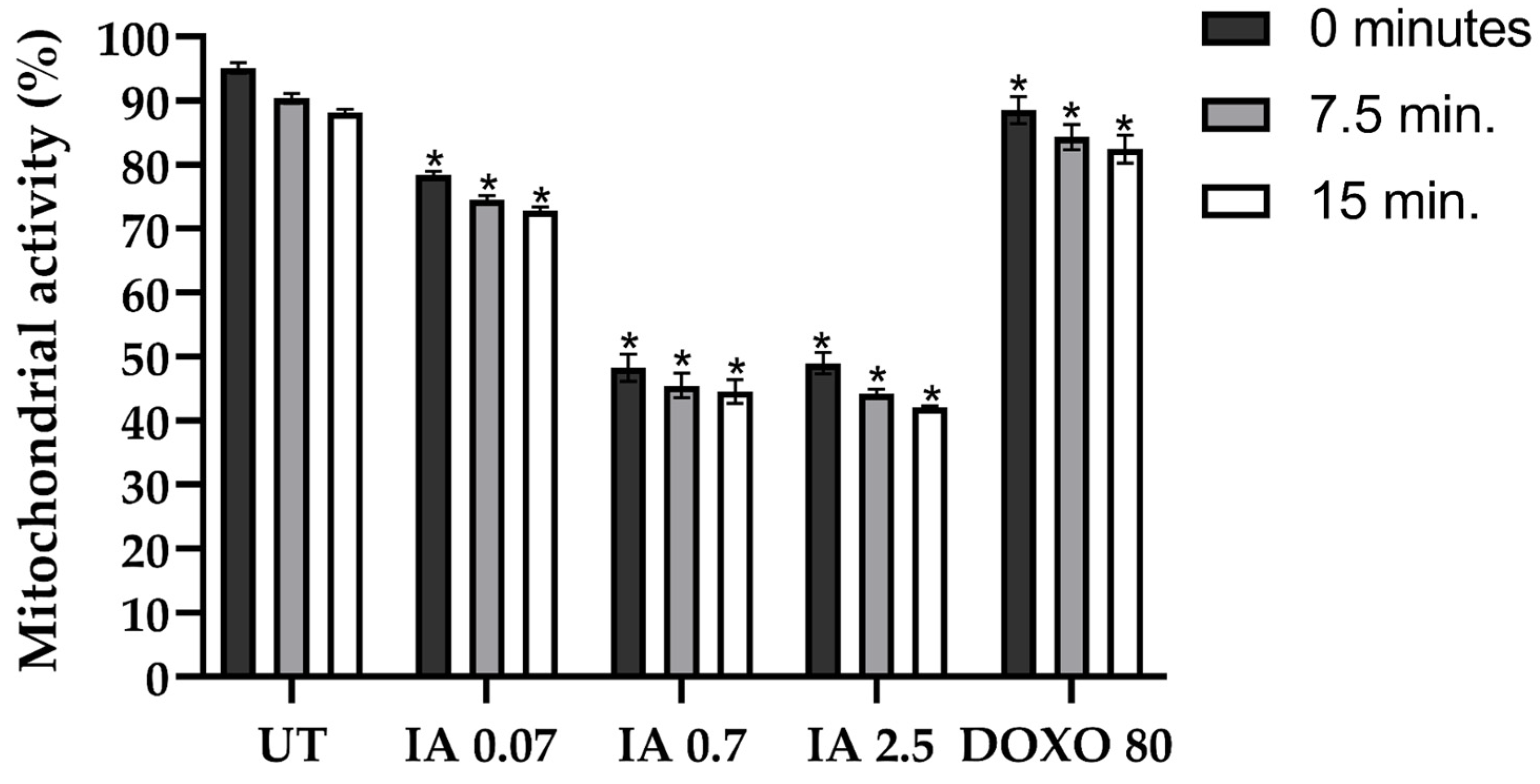
3.7. Incomptine A Decreases the Mitochondrial Activity of Breast Cancer Tumor Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BC | Breast cancer |
| HER-2+ | Human epidermal growth factor receptor 2 |
| IA | Incomptine A |
| HKII | Hexokinase II |
| VDAC1 | Voltage-dependent anion channel 1 |
| MOM | Mitochondrial outer membrane |
| ATP | Adenosine triphosphate |
| Cyt c | Cytochrome c |
| DOXO | Doxorubicin |
| UT | Untreated |
| CC50 | Cytotoxic concentration 50 |
| ED50 | Effective dose of 50 |
| LD50 | Lethal dose 50 |
| TI | Therapeutic index |
| ALDOA | Aldolase A |
| LDH | Lactate dehydrogenase |
| Bcl-2 | B-cell lymphoma 2 |
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef]
- INEGI (Instituto Nacional de Estadística, Geografía e Informática). Estadísticas a Propósito del día Internacional de la Lucha Contra el Cáncer de mama (19 de Octubre) Comunicado de Prensa número 595/23. 17 October 2023. Available online: https://www.inegi.org.mx/contenidos/saladeprensa/aproposito/2023/EAP_CMAMA23.pdf (accessed on 18 March 2025).
- Johnson, K.; Conant, E.; Soo, M. Molecular Subtypes of Breast Cancer: A Review for Breast Radiologists. J. Breast Imaging 2021, 3, 12–24. [Google Scholar] [CrossRef]
- Chun, K.; Park, J.; Fan, S. Predicting and Overcoming Chemotherapeutic Resistance in Breast Cancer. Adv. Exp. Med. Biol. 2017, 1026, 59–104. [Google Scholar]
- Schmitz, K.; DiSipio, T.; Gordon, L.; Hayes, S. Adverse breast cancer treatment effects: The economic case for making rehabilitative programs standard of care. Support. Care Cancer 2015, 23, 1807–1817. [Google Scholar] [CrossRef]
- Chaachouay, N.; Zidane, L. Plant-Derived Natural Products: A Source for Drug Discovery and Development. Drugs Drug Candidates 2024, 3, 184–207. [Google Scholar] [CrossRef]
- Dhyani, P.; Sati, P.; Sharma, E.; Attri, D.; Bahukhandi, A.; Tynybekov, B.; Szopa, A.; Sharifi-Rad, J.; Calina, D.; Suleria, H.A.; et al. Sesquiterpenoid lactones as potential anticancer agents: An update on molecular mechanisms and recent studies. Cancer Cell Int. 2022, 22, 305. [Google Scholar] [CrossRef]
- Pina-Jimenez, E.; Calzada, F.; Bautista, E.; Ordonez-Razo, R.; Velazquez, C.; Barbosa, E.; Garcia-Hernandez, N. Incomptine A Induces Apoptosis, ROS Production and a Differential Protein Expression on Non-Hodgkin’s Lymphoma Cells. Int. J. Mol. Sci. 2021, 22, 10516. [Google Scholar] [CrossRef] [PubMed]
- Calzada, F.; Bautista, E.; Hidalgo-Figueroa, S.; Garcia-Hernandez, N.; Barbosa, E.; Velazquez, C.; Ordonez-Razo, R.; Arietta-Garcia, A. Antilymphoma Effect of Incomptine A: In Vivo, In Silico, and Toxicological Studies. Molecules 2021, 26, 6646. [Google Scholar] [CrossRef] [PubMed]
- Arietta-Garcia, A.; Calzada, F.; Ramirez-Sanchez, I.; Bautista, E.; Garcia-Hernandez, N.; Ordonez-Razo, R. Advances in the Properties of Incomptine A: Cytotoxic Activity and Downregulation of Hexokinase II in Breast Cancer Cell Lines. Int. J. Mol. Sci. 2023, 24, 12406. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.; Koo, J.S. Glucose Metabolism and Glucose Transporters in Breast Cancer. Front. Cell Dev. Biol. 2021, 9, 728759. [Google Scholar] [CrossRef]
- Sato-Tadano, A.; Suzuki, T.; Amari, M.; Takagi, K.; Miki, Y.; Tamaki, K.; Watanabe, M.; Ishida, T.; Sasano, H.; Ohuchi, N. Hexokinase II in breast carcinoma: A potent prognostic factor associated with hypoxia-inducible factor-1alpha and Ki-67. Cancer Sci. 2013, 104, 1380–1388. [Google Scholar] [CrossRef]
- Schoeniger, A.; Wolf, P.; Edlich, F. How Do Hexokinases Inhibit Receptor-Mediated Apoptosis? Biology 2022, 11, 412. [Google Scholar] [CrossRef]
- Haloi, N.; Wen, P.; Cheng, Q.; Yang, M.; Natarajan, G.; Camara, A.; Kwok, W.; Tajkhorshid, E. Structural basis of complex formation between mitochondrial anion channel VDAC1 and Hexokinase-II. Commun. Biol. 2021, 4, 667. [Google Scholar] [CrossRef] [PubMed]
- Magri, A.; Reina, S.; De Pinto, V. VDAC1 as Pharmacological Target in Cancer and Neurodegeneration: Focus on Its Role in Apoptosis. Front. Chem. 2018, 6, 108. [Google Scholar] [CrossRef]
- Bautista, E.; Calzada, F.; Yepez-Mulia, L.; Chavez-Soto, M.; Ortega, A. Incomptines C and D, two heliangolides from Decachaeta. incompta. and their antiprotozoal activity. Planta Med. 2012, 78, 1698–1701. [Google Scholar] [CrossRef]
- Norma Oficial Mexicana. NOM-062-ZOO-1999: Especificaciones Técnicas Para la Producción, Cuidado y Uso de Los Animales de Laboratorio. 1999. Available online: https://www.gob.mx/cms/uploads/attachment/file/203498/NOM-062-ZOO-1999_220801.pdf (accessed on 18 March 2025).
- OECD. Guideline for Testing of Chemicals 423. Acute Oral Toxicity-Acute Toxic Class Method. Organización para la cooperación y el desarrollo económico, OECD/OCDE. 2001. Available online: https://read.oecd-ilibrary.org/environment/test-no-423-acute-oral-toxicity-acute-toxic-class-method_9789264071001-en#page1 (accessed on 18 March 2025).
- Pulaski, B.; Ostrand-Rosenberg, S. Mouse 4T1 breast tumor model. Curr. Protoc. Immunol. 2001, 20, 20–22. [Google Scholar] [CrossRef]
- Schlie-Guzman, M.; Garcia-Carranca, A.; Gonzalez-Esquinca, A. In vitro and in vivo antiproliferative activity of laherradurin and cherimolin-2 of Annona diversifolia Saff. Phytother. Res. 2009, 23, 1128–1133. [Google Scholar] [CrossRef]
- Muller, P.; Milton, M. The determination and interpretation of the therapeutic index in drug development. Nat. Rev. Drug Discov. 2012, 11, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Trang, K.; Kim, S.; Kim, I.; Lee, S.; Lee, S.; Kim, D.; Kim, S. Parthenolide suppresses tumor growth in a xenograft model of colorectal cancer cells by inducing mitochondrial dysfunction and apoptosis. Int. J. Oncol. 2012, 41, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Park, S.; Zhang, H.; Park, S.; Kwon, W.; Kim, E.; Zhang, X.; Jang, S.; Yoon, D.; Choi, S.; et al. Targeting AKT with costunolide suppresses the growth of colorectal cancer cells and induces apoptosis in vitro and in vivo. J. Exp. Clin. Cancer Res. 2021, 40, 114. [Google Scholar] [CrossRef]
- Thorn, C.; Oshiro, C.; Marsh, S.; Hernandez-Boussard, T.; McLeod, H.; Klein, T.; Altman, R. Doxorubicin pathways: Pharmacodynamics and adverse effects. Pharmacogenet. Genom. 2011, 21, 440–446. [Google Scholar] [CrossRef]
- Hiensch, A.E.; Bolam, K.; Mijwel, S.; Jeneson, J.; Huitema, A.; Kranenburg, O.; van der Wall, E.; Rundqvist, H.; Wengstrom, Y.; May, A. Doxorubicin-induced skeletal muscle atrophy: Elucidating the underlying molecular pathways. Acta Physiol. 2020, 229, e13400. [Google Scholar] [CrossRef] [PubMed]
- Jadapalli, J.; Wright, G.; Kain, V.; Sherwani, M.; Sonkar, R.; Yusuf, N.; Halade, G. Doxorubicin triggers splenic contraction and irreversible dysregulation of COX and LOX that alters the inflammation-resolution program in the myocardium. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1091–H1100. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, P.; Renu, K.; Valsala Gopalakrishnan, A. New molecular and biochemical insights of doxorubicin-induced hepatotoxicity. Life Sci. 2020, 250, 117599. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, S.; Wang, X. The Metabolic Mechanisms of Breast Cancer Metastasis. Front. Oncol. 2020, 10, 602416. [Google Scholar] [CrossRef]
- Navarro, C.; Ortega, A.; Santeliz, R.; Garrido, B.; Chacin, M.; Galban, N.; Vera, I.; De Sanctis, J.; Bermudez, V. Metabolic Reprogramming in Cancer Cells: Emerging Molecular Mechanisms and Novel Therapeutic Approaches. Pharmaceutics 2022, 14, 1301. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Ma, H.; Wang, J.; Huang, M.; Fu, D.; Qin, L.; Yin, Q. Energy metabolism pathways in breast cancer progression: The reprogramming, crosstalk, and potential therapeutic targets. Transl. Oncol. 2022, 26, 101534. [Google Scholar] [CrossRef]
- Li, R.; Mei, S.; Ding, Q.; Wang, Q.; Yu, L.; Zi, F. A pan-cancer analysis of the role of hexokinase II (HK2) in human tumors. Sci. Rep. 2022, 12, 18807. [Google Scholar] [CrossRef]
- Zhang, D.; Yip, Y.; Li, L. In silico construction of HK2-VDAC1 complex and investigating the HK2 binding-induced molecular gating mechanism of VDAC1. Mitochondrion 2016, 30, 222–228. [Google Scholar] [CrossRef]
- Bachmann, M.; Costa, R.; Peruzzo, R.; Prosdocimi, E.; Checchetto, V.; Leanza, L. Targeting Mitochondrial Ion Channels to Fight Cancer. Int. J. Mol. Sci. 2018, 19, 2060. [Google Scholar] [CrossRef]
- Stein, M.; Lin, H.; Jeyamohan, C.; Dvorzhinski, D.; Gounder, M.; Bray, K.; Eddy, S.; Goodin, S.; White, E.; DiPaola, R. Targeting tumor metabolism with 2-deoxyglucose in patients with castrate-resistant prostate cancer and advanced malignancies. Prostate 2010, 70, 1388–1394. [Google Scholar] [CrossRef]
- Muley, P.; Olinger, A.; Tummala, H. 2-Deoxyglucose induces cell cycle arrest and apoptosis in colorectal cancer cells independent of its glycolysis inhibition. Nutr. Cancer 2015, 67, 514–522. [Google Scholar] [CrossRef]
- Raez, L.; Papadopoulos, K.; Ricart, A.; Chiorean, E.; DiPaola, R.; Stein, M.; Rocha Lima, C.; Schlesselman, J.; Tolba, K.; Langmuir, V.; et al. A phase I dose-escalation trial of 2-deoxy-d-glucose alone or combined with docetaxel in patients with advanced solid tumors. Cancer Chemother. Pharmacol. 2013, 71, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; O’Donnell, A. ‘Sugarcoating’ 2-deoxyglucose: Mechanisms that suppress its toxic effects. Curr. Genet. 2021, 67, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Yin, Y.; Zhang, Y.; Zhao, Q.; Yang, Y.; Zhang, Z.; Zou, X. Parthenolide as a potential therapeutic agent for gastric cancer: Exploring oxidative stress and DNA damage. Pharmacol. Res. Mod. Chin. Med. 2024, 10, 100365. [Google Scholar] [CrossRef]
- Rotondo, R.; Oliva, M.; Arcella, A. The Sesquiterpene Lactone Cynaropicrin Manifests Strong Cytotoxicity in Glioblastoma Cells U-87 MG by Induction of Oxidative Stress. Biomedicines 2022, 10, 1583. [Google Scholar] [CrossRef]
- Chen, J.; Yan, Q.; Bai, M.; Liu, Q.; Song, S.; Yao, G. Deoxyelephantopin, a germacrane-type sesquiterpene lactone from Elephantopus scaber, induces mitochondrial apoptosis of hepatocarcinoma cells by targeting Hsp90alpha in vitro and in vivo. Phytother. Res. 2023, 37, 702–716. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arietta-García, A.G.; Calzada, F.; Franco-Vadillo, A.; Barrientos-Buendía, I.M.; Alarcón-Aguilar, F.J.; Bautista, E.; Santana-Sánchez, P.; Ramírez-Sánchez, I.; Ordoñez-Razo, R.M. The Antitumor Role of Incomptine A in a Breast Cancer Murine Model: Impairment of Hexokinase II Expression and Apoptosis Induction. Cells 2025, 14, 1192. https://doi.org/10.3390/cells14151192
Arietta-García AG, Calzada F, Franco-Vadillo A, Barrientos-Buendía IM, Alarcón-Aguilar FJ, Bautista E, Santana-Sánchez P, Ramírez-Sánchez I, Ordoñez-Razo RM. The Antitumor Role of Incomptine A in a Breast Cancer Murine Model: Impairment of Hexokinase II Expression and Apoptosis Induction. Cells. 2025; 14(15):1192. https://doi.org/10.3390/cells14151192
Chicago/Turabian StyleArietta-García, Angel Giovanni, Fernando Calzada, Antonio Franco-Vadillo, Irais Monserrat Barrientos-Buendía, Francisco Javier Alarcón-Aguilar, Elihú Bautista, Paola Santana-Sánchez, Israel Ramírez-Sánchez, and Rosa María Ordoñez-Razo. 2025. "The Antitumor Role of Incomptine A in a Breast Cancer Murine Model: Impairment of Hexokinase II Expression and Apoptosis Induction" Cells 14, no. 15: 1192. https://doi.org/10.3390/cells14151192
APA StyleArietta-García, A. G., Calzada, F., Franco-Vadillo, A., Barrientos-Buendía, I. M., Alarcón-Aguilar, F. J., Bautista, E., Santana-Sánchez, P., Ramírez-Sánchez, I., & Ordoñez-Razo, R. M. (2025). The Antitumor Role of Incomptine A in a Breast Cancer Murine Model: Impairment of Hexokinase II Expression and Apoptosis Induction. Cells, 14(15), 1192. https://doi.org/10.3390/cells14151192