
Gene Expression Analysis of HPRT-Deficient Cells Maintained with Physiological Levels of Folic Acid

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Generation of Pluripotent HPRT-Deficient Cells
2.2. Cell Culture Medium and Differentiation of NT2/D1
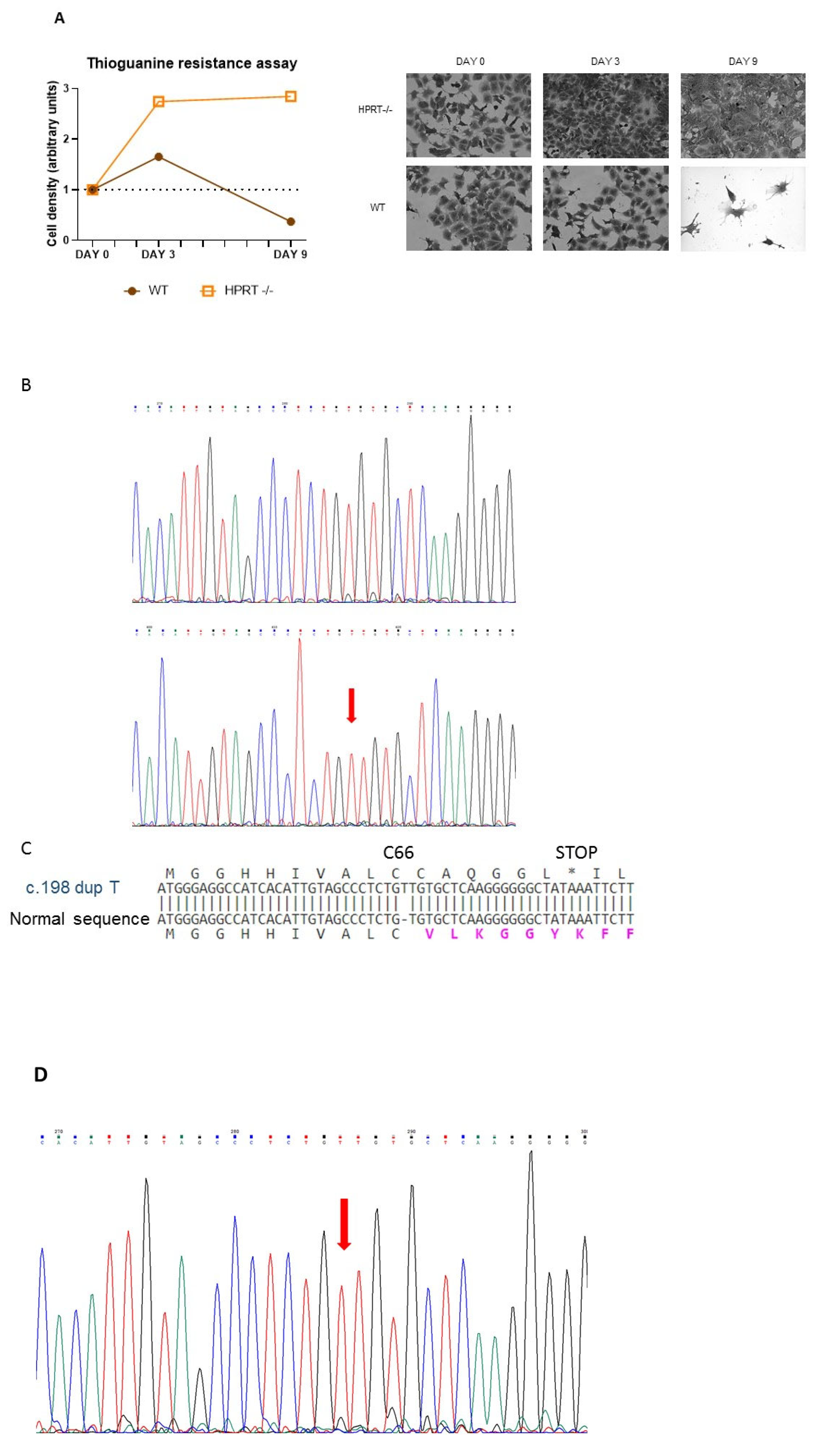
2.3. 6-Thioguanine Toxicity Assay
2.4. DNA Sequencing
2.5. HPRT1 Expression Levels and HPRT Activity
2.6. Hypoxanthine and Xanthine Determination
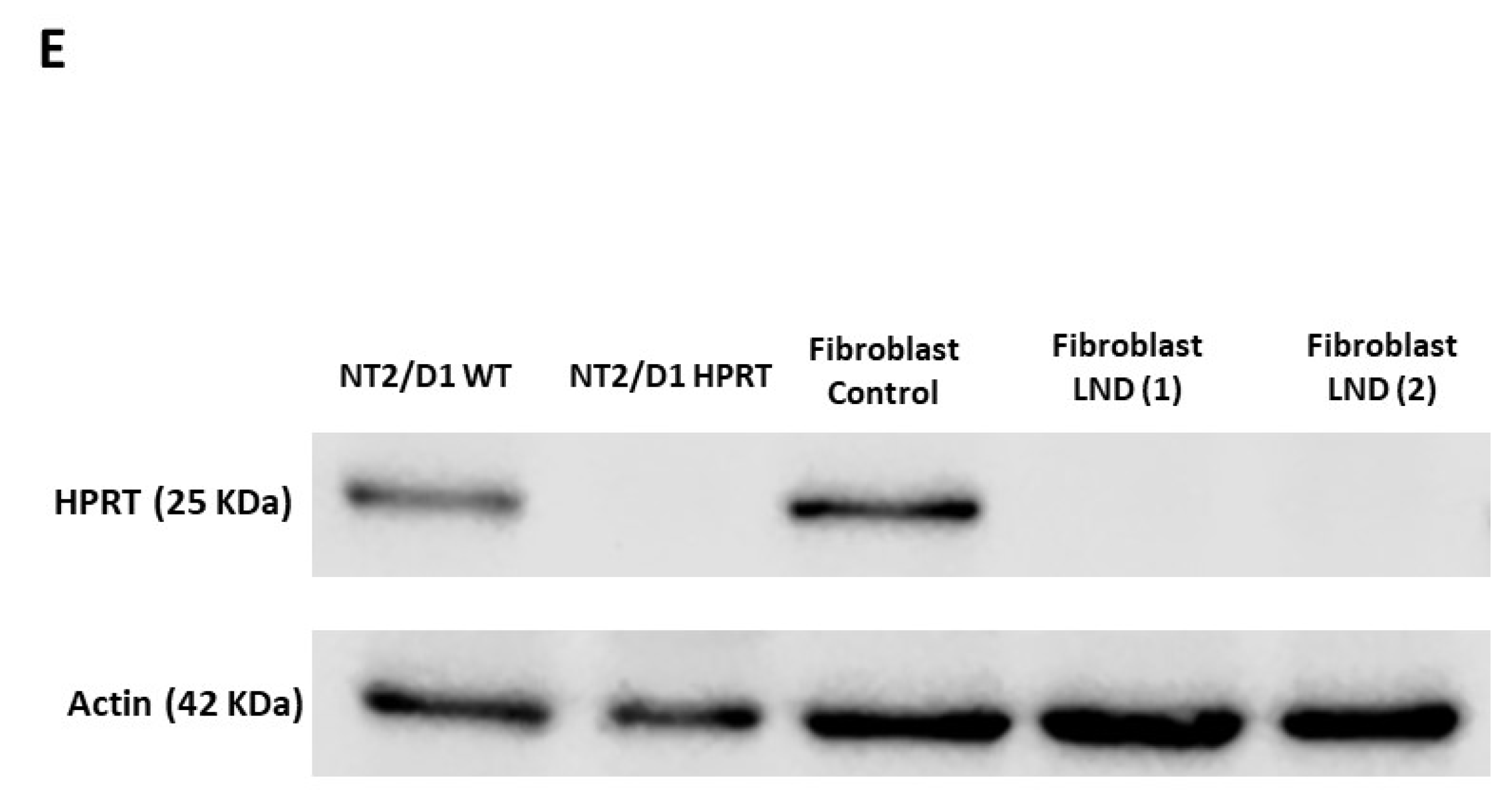
2.7. Western Blotting
2.8. Real-Time Quantitative PCR Array of Selected Genes Related to Neuronal Differentiation in HPRT-Deficient and Wild-Type NTD2/D1 Cells
2.9. Differential Global Gene Expression by RNAseq in HPRT-Deficient and Wild-Type NTD2/D1 Cells
2.10. Gene Ontology Analysis
3. Results
3.1. Generation of Pluripotent HPRT-Deficient Cells
3.2. Real-Time Quantitative PCR Array of Selected Genes Related to Neuronal Differentiation
3.2.1. Wild-Type Versus HPRT-Deficient Cells
3.2.2. Differentiated Wild-Type Versus Differentiated HPRT-Deficient Cells
3.3. Differential Global Gene Expression by RNAseq
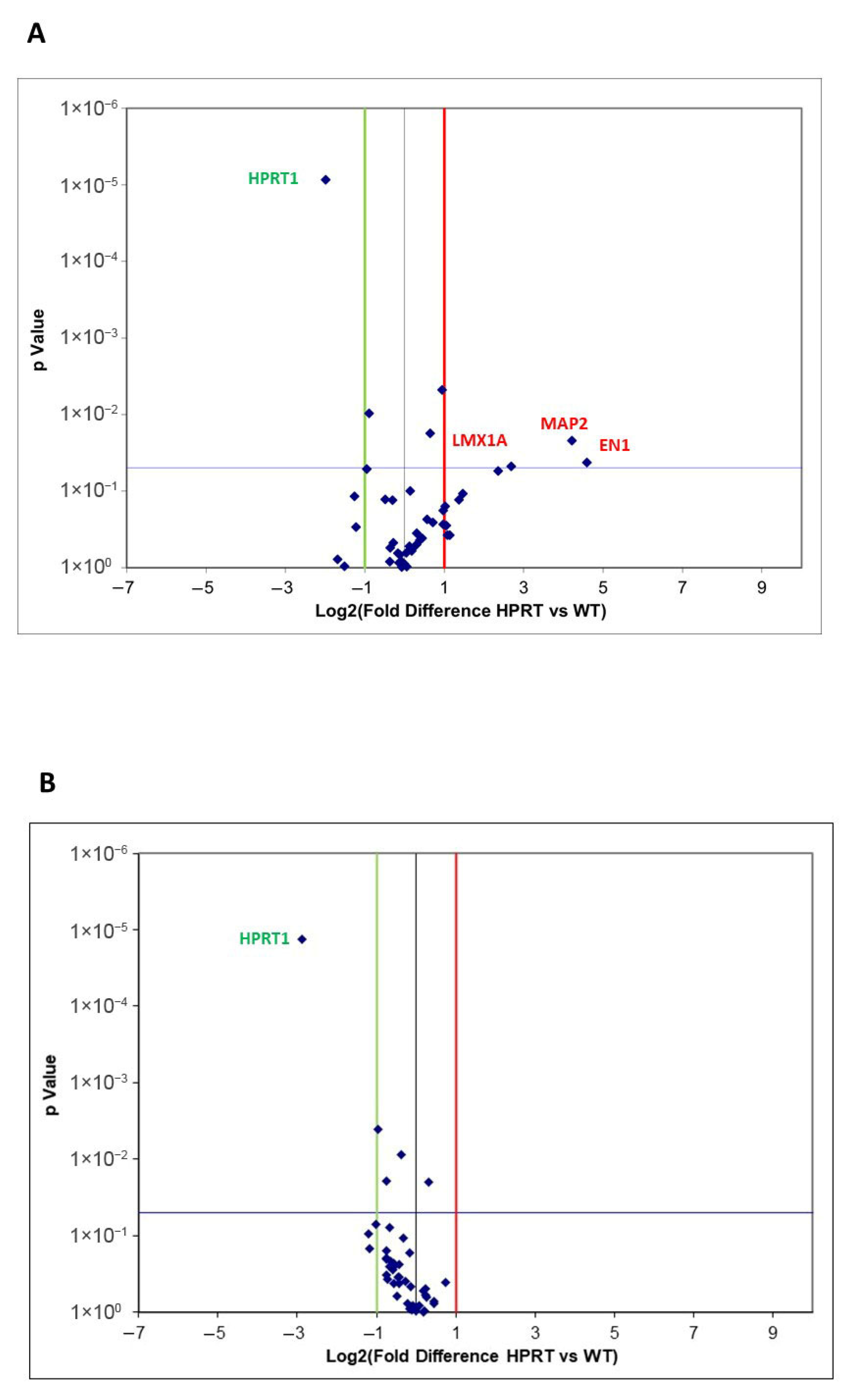
3.3.1. Wild-Type Versus HPRT-Deficient Cells
3.3.2. Differentiated Wild-Type Versus Differentiated HPRT-Deficient Cells
3.4. Gene Ontology Enrichment Analysis
3.4.1. Wild-Type Versus HPRT-Deficient Cells
3.4.2. Differentiated Wild-Type Versus Differentiated HPRT-Deficient Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lesch Lesch, M.; Nyhan, W.L. A familial disorder of uric acid metabolism and central nervous system function. Am. J. Med. 1964, 36, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Jinnah, H.A.; Visser, J.E.; Harris, J.C.; Verdu, A.; Larovere, L.; Ceballos-Picot, I.; Gonzalez-Alegre, P.; Neychev, V.; Torres, R.J.; Dulac, O.; et al. Lesch-Nyhan Disease International Study Group. Delineation of the motor disorder of Lesch-Nyhan disease. Brain 2006, 129, 1201–1217. [Google Scholar] [CrossRef] [PubMed]
- Seegmiller, J.E.; Rosenbloom, F.M.; Kelley, W.N. Enzyme defect associated with a sex-linked human neurological disorder and excessive purine synthesis. Science 1967, 31, 1682–1684. [Google Scholar] [CrossRef]
- Edwards, A.; Voss, H.; Rice, P.; Civitello, A.; Stegemann, J.; Schwager, C.; Zimmermann, J.; Erfle, H.; Caskey, C.T.; Ansorge, W. Automated DNA sequencing of the human HPRT locus. Genomics 1990, 6, 593–608. [Google Scholar] [CrossRef]
- Kelley, W.N.; Rosenbloom, F.M.; Henderson, J.F.; Seegmiller, J.E. A specific enzyme defect in gout associated with overproduction of uric acid. Proc. Natl. Acad. Sci. USA 1967, 57, 1735–1739. [Google Scholar] [CrossRef]
- Jinnah, H.A.; Ceballos-Picot, I.; Torres, R.J.; Visser, J.E.; Schretlen, D.J.; Verdu, A.; Laróvere, L.E.; Chen, C.J.; Cossu, A.; Wu, C.H.; et al. Lesch-Nyhan Disease International Study Group. Attenuated variants of Lesch-Nyhan disease. Brain 2010, 133, 671–689. [Google Scholar] [CrossRef]
- Rosenbloom, F.M.; Henderson, J.F.; Caldwell, I.C.; Kelley, W.N.; Seegmiller, J.E. Biochemical bases of accelerated purine biosynthesis de novo in human fibroblasts lacking hypoxanthine-guanine phosphoribosyltransferase. J. Biol. Chem. 1968, 243, 1166–1173. [Google Scholar] [CrossRef] [PubMed]
- López, J.M.; Outtrim, E.L.; Fu, R.; Sutcliffe, D.J.; Torres, R.J.; Jinnah, H.A. Physiological levels of folic acid reveal purine alterations in Lesch-Nyhan disease. Proc. Natl. Acad. Sci. USA 2020, 117, 12071–12079. [Google Scholar] [CrossRef]
- Torres, R.J.; Puig, J.G. Hypoxanthine deregulates genes involved in early neuronal development. Implications in Lesch-Nyhan disease pathogenesis. J. Inherit. Metab. Dis. 2015, 38, 1109–1118. [Google Scholar] [CrossRef]
- López, J.M. Is ZMP the toxic metabolite in Lesch-Nyhan disease? Med. Hypotheses 2008, 71, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Ernst, M.; Zametkin, A.J.; Matochik, J.A.; Pascualvaca, D.; Jons, P.H.; Hardy, K.; Hankerson, J.G.; Doudet, D.J.; Cohen, R.M. Presynaptic dopaminergic deficits in Lesch-Nyhan disease. N. Engl. J. Med. 1996, 334, 1568–1572. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.F.; Harris, J.C.; Naidu, S.; Yokoi, F.; Marenco, S.; Dannals, R.F.; Ravert, H.T.; Yaster, M.; Evans, A.; Rousset, O.; et al. Dopamine transporters are markedly reduced in Lesch-Nyhan disease in vivo. Proc. Natl. Acad. Sci. USA 1996, 93, 5539–5543. [Google Scholar] [CrossRef] [PubMed]
- Jinnah, H.A.; Wojcik, B.E.; Hunt, M.; Narang, N.; Lee, K.Y.; Goldstein, M.; Wamsley, J.K.; Langlais, P.J.; Friedmann, T. Dopamine deficiency in a genetic mouse model of Lesch-Nyhan disease. J. Neurosci. 1994, 14, 1164–1175. [Google Scholar] [CrossRef]
- Witteveen, J.S.; Loopstok, S.R.; Ballesteros, L.L.; Boonstra, A.; van Bakel, N.H.M.; van Boekel, W.H.P.; Martens, G.J.M.; Visser, J.E.; Kolk, S.M. HGprt deficiency disrupts dopaminergic circuit development in a genetic mouse model of Lesch-Nyhan disease. Cell Mol. Life Sci. 2022, 79, 341. [Google Scholar] [CrossRef]
- Schretlen, D.J.; Varvaris, M.; Ho, T.E.; Vannorsdall, T.D.; Gordon, B.; Harris, J.C.; Jinnah, H.A. Regional brain volume abnormalities in Lesch-Nyhan disease and its variants: A cross-sectional study. Lancet Neurol. 2013, 12, 1151–1158. [Google Scholar] [CrossRef]
- Schretlen, D.J.; Varvaris, M.; Vannorsdall, T.D.; Gordon, B.; Harris, J.C.; Jinnah, H.A. Brain white matter volume abnormalities in Lesch-Nyhan disease and its variants. Neurology 2015, 84, 190–196. [Google Scholar] [CrossRef]
- Dinasarapu, A.R.; Sutcliffe, D.J.; Seifar, F.; Visser, J.E.; Jinnah, H.A. Abnormalities of neural stem cells in Lesch-Nyhan disease. J. Neurogenet. 2022, 36, 81–87. [Google Scholar] [CrossRef]
- Sutcliffe, D.J.; Dinasarapu, A.R.; Visser, J.E.; Hoed, J.D.; Seifar, F.; Joshi, P.; Ceballos-Picot, I.; Sardar, T.; Hess, E.J.; Sun, Y.V.; et al. Induced pluripotent stem cells from subjects with Lesch-Nyhan disease. Sci. Rep. 2021, 11, 8523. [Google Scholar] [CrossRef]
- Ruillier, V.; Tournois, J.; Boissart, C.; Lasbareilles, M.; Mahé, G.; Chatrousse, L.; Cailleret, M.; Peschanski, M.; Benchoua, A. Rescuing compounds for Lesch-Nyhan disease identified using stem cell-based phenotypic screening. JCI Insight 2020, 5, e132094. [Google Scholar] [CrossRef]
- Mekhoubad, S.; Bock, C.; de Boer, A.S.; Kiskinis, E.; Meissner, A.; Eggan, K. Erosion of dosage compensation impacts human iPSC disease modeling. Cell Stem Cell 2012, 10, 595–609. [Google Scholar] [CrossRef]
- Cristini, S.; Navone, S.; Canzi, L.; Acerbi, F.; Ciusani, E.; Hladnik, U.; de Gemmis, P.; Alessandri, G.; Colombo, A.; Parati, E.; et al. Human neural stem cells: A model system for the study of Lesch-Nyhan disease neurological aspects. Hum. Mol. Genet. 2010, 19, 1939–1950. [Google Scholar] [CrossRef]
- Ceballos-Picot, I.; Mockel, L.; Potier, M.C.; Dauphinot, L.; Shirley, T.L.; Torero-Ibad, R.; Fuchs, J.; Jinnah, H.A. Hypoxanthine-guanine phosphoribosyl transferase regulates early developmental programming of dopamine neurons: Implications for Lesch-Nyhan disease pathogenesis. Hum. Mol. Genet. 2009, 18, 2317–2327. [Google Scholar] [CrossRef] [PubMed]
- Guibinga, G.H.; Hsu, S.; Friedmann, T. Deficiency of the housekeeping gene hypoxanthine-guanine phosphoribosyltransferase (HPRT) dysregulates neurogenesis. Mol. Ther. 2010, 18, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.H.; Guibinga, G.H.; Jinnah, H.A.; Friedmann, T. HPRT deficiency coordinately dysregulates canonical Wnt and presenilin-1 signaling: A neuro-developmental regulatory role for a housekeeping gene? PLoS ONE 2011, 6, e16572. [Google Scholar] [CrossRef]
- Guibinga, G.H.; Murray, F.; Barron, N. HPRT-deficiency dysregulates cAMP-PKA signaling and phosphodiesterase 10A expression: Mechanistic insight and potential target for Lesch-Nyhan Disease? PLoS ONE 2013, 8, e63333. [Google Scholar] [CrossRef]
- Connolly, G.P. Hypoxanthine-guanine phosphoribosyltransferase-deficiency produces aberrant neurite outgrowth of rodent neuroblastoma used to model the neurological disorder Lesch Nyhan syndrome. Neurosci. Lett. 2001, 314, 61–64. [Google Scholar] [CrossRef]
- Boer, P.; Brosh, S.; Wasserman, L.; Hammel, I.; Zoref-Shani, E.; Sperling, O. Decelerated rate of dendrite outgrowth from dopaminergic neurons in primary cultures from brains of hypoxanthine phosphoribosyltransferase-deficient knockout mice. Neurosci. Lett. 2001, 303, 45–48. [Google Scholar] [CrossRef]
- Lewers, J.C.; Ceballos-Picot, I.; Shirley, T.L.; Mockel, L.; Egami, K.; Jinnah, H.A. Consequences of impaired purine recycling in dopaminergic neurons. Neuroscience 2008, 152, 761–772. [Google Scholar] [CrossRef]
- Escudero-Ferruz, P.; Ontiveros, N.; Cano-Estrada, C.; Sutcliffe, D.J.; Jinnah, H.A.; Torres, R.J.; López, J.M. A new physiological medium uncovers biochemical and cellular alterations in Lesch-Nyhan disease fibroblasts. Mol. Med. 2024, 30, 3. [Google Scholar] [CrossRef]
- Pleasure, S.J.; Page, C.; Lee, V.M. Pure, postmitotic, polarized human neurons derived from NTera 2 cells provide a system for expressing exogenous proteins in terminally differentiated neurons. J. Neurosci. 1992, 12, 1802–1815. [Google Scholar] [CrossRef]
- Andrews, P.W. Retinoic acid induces neuronal differentiation of a cloned human embryonal carcinoma cell line in vitro. Dev. Biol. 1984, 103, 285–293. [Google Scholar] [CrossRef] [PubMed]
- García, M.G.; Torres, R.J.; Prior, C.; Puig, J.G. Normal HPRT coding region in complete and partial HPRT deficiency. Mol. Genet. Metab. 2008, 94, 167–172. [Google Scholar] [CrossRef]
- Rylance, H.J.; Wallace, R.C.; Nuki, G. Hypoxanthine-guanine phosphoribosyl transferase: Assay using high performance liquid chromatography. Clin. Chim. Acta 1982, 121, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Puig, J.G.; Torres, R.J.; Mateos, F.A.; Ramos, T.H.; Arcas, J.M.; Buño, A.S.; O’Neill, P. The spectrum of hypoxanthine-guanine phosphoribosyltransferase (HPRT) deficiency. Clinical experience based on 22 patients from 18 Spanish families. Medicine 2001, 80, 102–112. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 2 June 2025).
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Frazee, A.C.; Pertea, G.; Jaffe, A.E.; Langmead, B.; Salzberg, S.L.; Leek, J.T. Ballgown bridges the gap between transcriptome assembly and expression analysis. Nat. Biotechnol. 2015, 33, 243–246. [Google Scholar] [CrossRef]
- The Gene Ontology Consortium. The Gene Ontology knowledgebase in 2023. Genetics 2023, 224, iyad031. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.D.; Ebert, D.; Muruganujan, A.; Mushayahama, T.; Albou, L.P.; Mi, H. PANTHER: Making genome-scale phylogenetics accessible to all. Protein Sci. 2022, 31, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Öz, S.; Maercker, C.; Breiling, A. Embryonic carcinoma cells show specific dielectric resistance profiles during induced differentiation. PLoS ONE 2013, 8, e59895. [Google Scholar] [CrossRef] [PubMed]
- Torres, R.J.; Puig, J.G. Aicar effect in early neuronal development. Nucleosides Nucleotides Nucleic Acids 2018, 37, 261–272. [Google Scholar] [CrossRef]
- Wongtrakoongate, P.; Jones, M.; Gokhale, P.J.; Andrews, P.W. STELLA facilitates differentiation of germ cell and endodermal lineages of human embryonic stem cells. PLoS ONE 2013, 8, e56893. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, C.; Xu, J.; Liu, S.; Yu, L.; Chen, S.; Wen, H.; Li, Z.; Liu, N. Dppa3 facilitates self-renewal of embryonic stem cells by stabilization of pluripotent factors. Stem Cell Res. Ther. 2022, 13, 169. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Chen, X.; Rodgers, B.; Miura, F.; Bashtrykov, P.; Bonhomme, F.; Salinas-Luypaert, C.; Haxholli, D.; Gutekunst, N.; Aygenli, B.Ö.; et al. Non-canonical functions of UHRF1 maintain DNA methylation homeostasis in cancer cells. Nat. Commun. 2024, 15, 2960. [Google Scholar] [CrossRef]
- Zhou, S.; Liu, Y.; Ma, Y.; Zhang, X.; Li, Y.; Wen, J. C9ORF135 encodes a membrane protein whose expression is related to pluripotency in human embryonic stem cells. Sci. Rep. 2017, 7, 45311. [Google Scholar] [CrossRef]
- Stacey, N.C.; Ma, M.H.; Duley, J.A.; Connolly, G.P. Abnormalities in cellular adhesion of neuroblastoma and fibroblast models of Lesch Nyhan syndrome. Neuroscience 2000, 98, 397–401. [Google Scholar] [CrossRef]
- Bottiglieri, T. S-Adenosyl-L-methionine (SAMe): From the bench to the bedside--molecular basis of a pleiotrophic molecule. Am. J. Clin. Nutr. 2002, 76, 1151S–1157S. [Google Scholar] [CrossRef]
- Dolcetta, D.; Parmigiani, P.; Salmaso, L.; Bernardelle, R.; Cesari, U.; Andrighetto, G.; Baschirotto, G.; Nyhan, W.L.; Hladnik, U. Quantitative evaluation of the clinical effects of S-adenosylmethionine on mood and behavior in Lesch-Nyhan patients. Nucleosides Nucleotides Nucleic Acids 2013, 32, 174–188. [Google Scholar] [CrossRef]
- Gonçalves, C.S.; Le Boiteux, E.; Arnaud, P.; Costa, B.M. HOX gene cluster (de)regulation in brain: From neurodevelopment to malignant glial tumours. Cell Mol. Life Sci. 2020, 77, 3797–3821. [Google Scholar] [CrossRef] [PubMed]
- Quinonez, S.C.; Innis, J.W. Human HOX gene disorders. Mol. Genet. Metab. 2014, 111, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Kala, K.; Haugas, M.; Lilleväli, K.; Guimera, J.; Wurst, W.; Salminen, M.; Partanen, J. Gata2 is a tissue-specific post-mitotic selector gene for midbrain GABAergic neurons. Development 2009, 136, 253–262. [Google Scholar] [CrossRef] [PubMed]
- French, J.B.; Zhao, H.; An, S.; Niessen, S.; Deng, Y.; Cravatt, B.F.; Benkovic, S.J. Hsp70/Hsp90 chaperone machinery is involved in the assembly of the purinosome. Proc. Natl. Acad. Sci. USA 2013, 110, 2528–2533. [Google Scholar] [CrossRef]
- Fu, R.; Sutcliffe, D.; Zhao, H.; Huang, X.; Schretlen, D.J.; Benkovic, S.; Jinnah, H.A. Clinical severity in Lesch-Nyhan disease: The role of residual enzyme and compensatory pathways. Mol. Genet. Metab. 2015, 114, 55–61. [Google Scholar] [CrossRef]
- Chan, C.Y.; Pedley, A.M.; Kim, D.; Xia, C.; Zhuang, X.; Benkovic, S.J. Microtubule-directed transport of purine metabolons drives their cytosolic transit to mitochondria. Proc. Natl. Acad. Sci. USA 2018, 115, 13009–13014. [Google Scholar] [CrossRef]
- Zhao, H.; Chiaro, C.R.; Zhang, L.; Smith, P.B.; Chan, C.Y.; Pedley, A.M.; Pugh, R.J.; French, J.B.; Patterson, A.D.; Benkovic, S.J. Quantitative analysis of purine nucleotides indicates that purinosomes increase de novo purine biosynthesis. J. Biol. Chem. 2015, 290, 6705–6713. [Google Scholar] [CrossRef]
- O’Neill, A.G.; Burrell, A.L.; Zech, M.; Elpeleg, O.; Harel, T.; Edvardson, S.; Mor-Shaked, H.; Rippert, A.L.; Nomakuchi, T.; Izumi, K.; et al. Neurodevelopmental disorder mutations in the purine biosynthetic enzyme IMPDH2 disrupt its allosteric regulation. J. Biol. Chem. 2023, 299, 105012. [Google Scholar] [CrossRef]
- Mortimer, S.E.; Xu, D.; McGrew, D.; Hamaguchi, N.; Lim, H.C.; Bowne, S.J.; Daiger, S.P.; Hedstrom, L. IMP dehydrogenase type 1 associates with polyribosomes translating rhodopsin mRNA. J. Biol. Chem. 2008, 283, 36354–36360. [Google Scholar] [CrossRef]
- Kozhevnikova, E.N.; van der Knaap, J.A.; Pindyurin, A.V.; Ozgur, Z.; van Ijcken, W.F.; Moshkin, Y.M.; Verrijzer, C.P. Metabolic enzyme IMPDH is also a transcription factor regulated by cellular state. Mol. Cell. 2012, 47, 133–139. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, X.; Hu, J.; Li, T.; Chen, H.Z.; Zhang, A.; Wang, H.; Yu, J.; Zhang, L. PRPS2 enhances RNA m6A methylation by stimulating SAM synthesis through enzyme-dependent and independent mechanisms. Nat. Commun. 2025, 16, 3966. [Google Scholar] [CrossRef]
- Srivastava, S.; Samaniego-Castruita, D.; Khurana, S.; Shukla, V.; Ben-Sahra, I.; Foltz, D.R. Rate Limiting Enzymes in Nucleotide Metabolism Synchronize Nucleotide Biosynthesis and Chromatin Formation. bioRxiv 2025. [Google Scholar] [CrossRef] [PubMed]
- Hershfield, M.; Tarrant, T. Adenosine Deaminase Deficiency. 2006 Oct 3 [Updated 2024 Mar 7]. In GeneReviews® [Internet]; Adam, M.P., Feldman, J., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 2024. Available online: https://www.ncbi.nlm.nih.gov/books/NBK1483/ (accessed on 2 June 2025).
- Schoggins, J.W. Interferon-Stimulated Genes: What Do They All Do? Annu. Rev. Virol. 2019, 6, 567–584. [Google Scholar] [CrossRef]
- Allison, A.C.; Hovi, T.; Watts, R.W.; Webster, A.D. Immunological observations on patients with Lesch-Nyhan syndrome, and on the role of de-novo purine synthesis in lymphocyte transformation. Lancet 1975, 2, 1179–1183. [Google Scholar] [CrossRef] [PubMed]
- Gelfand, E.W.; Fox, I.H.; Stuckey, M.; Dosch, H.M. Normal B-lymphocyte function in patients with Lesch-Nyhan syndrome and HGPRT deficiency. Clin. Exp. Immunol. 1978, 31, 205–208. [Google Scholar] [PubMed]
- Bell, S.; McCarty, V.; Peng, H.; Jefri, M.; Hettige, N.; Antonyan, L.; Crapper, L.; O’Leary, L.A.; Zhang, X.; Zhang, Y.; et al. Lesch-Nyhan disease causes impaired energy metabolism and reduced developmental potential in midbrain dopaminergic cells. Stem Cell Rep. 2021, 16, 1749–1762. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Track id | Gene Name | log2FC | Fold Change | p Value | q Value |
|---|---|---|---|---|---|
| ENSG00000187569.2_2 | DPPA3 | −5.560093447 | 0.02119557 | 0.000419802 | 0.040596923 |
| ENSG00000118523.5_2 | CCN2 | −3.244903704 | 0.105484015 | 0.000278163 | 0.036303633 |
| ENSG00000142871.16_3 | CCN1 | −3.079243133 | 0.118319262 | 0.000260377 | 0.03573791 |
| ENSG00000165704.14_2 | HPRT1 | −2.786982045 | 0.144888797 | 2.73957 × 10−6 | 0.012297513 |
| ENSG00000148677.6_2 | ANKRD1 | −2.313122967 | 0.201224382 | 0.00093275 | 0.050325601 |
| ENSG00000120937.8_2 | NPPB | −2.212740872 | 0.21572408 | 0.001104595 | 0.052000195 |
| ENSG00000197614.10_2 | MFAP5 | −2.149300526 | 0.225421882 | 0.002006182 | 0.061695869 |
| ENSG00000135046.13_3 | ANXA1 | −1.987181661 | 0.252231146 | 0.000521378 | 0.043820278 |
| ENSG00000166741.7_3 | NNMT | −1.978824709 | 0.25369646 | 0.013596552 | 0.12292049 |
| ENSG00000106366.8_2 | SERPINE1 | −1.9413966 | 0.260364273 | 0.011572679 | 0.116032592 |
| ENSG00000156265.15_2 | MAP3K7CL | −1.938052915 | 0.26096841 | 0.01034748 | 0.111481945 |
| ENSG00000106823.12_3 | ECM2 | −1.932553968 | 0.26196501 | 0.001852116 | 0.060764564 |
| ENSG00000188015.9_3 | S100A3 | −1.900994117 | 0.267758798 | 0.003546679 | 0.072840089 |
| ENSG00000101460.12_2 | MAP1LC3A | −1.899122452 | 0.268106397 | 0.000409214 | 0.040303113 |
| ENSG00000163347.5_2 | CLDN1 | −1.857091876 | 0.276032133 | 0.000494154 | 0.042868547 |
| ENSG00000100345.20_2 | MYH9 | −1.76131462 | 0.29497925 | 0.000256178 | 0.03573791 |
| ENSG00000204711.8_2 | CFAP95 | −1.727468258 | 0.30198143 | 0.002632984 | 0.065817254 |
| ENSG00000135932.10_2 | CAB39 | −1.717650144 | 0.304043543 | 0.001235067 | 0.053730422 |
| ENSG00000198380.12_2 | GFPT1 | −1.631076319 | 0.322847258 | 0.037602012 | 0.180752644 |
| ENSG00000105971.14_2 | CAV2 | −1.600604271 | 0.329738838 | 0.024674901 | 0.150097079 |
| ENSG00000134531.9_3 | EMP1 | −1.589696922 | 0.332241243 | 0.003645433 | 0.073941233 |
| Track id | Gene Name | log2FC | Fold Change | p Value | q Value |
|---|---|---|---|---|---|
| ENSG00000168542.14_3 | COL3A1 | 4.247956394 | 19.00038035 | 0.000379474 | 0.039430012 |
| ENSG00000124208.16_3 | TMEM189-UBE2V1 | 3.408428068 | 10.61791114 | 0.028791507 | 0.161419507 |
| ENSG00000139219.17_3 | COL2A1 | 2.640248493 | 6.234390369 | 0.03364278 | 0.172691892 |
| ENSG00000125872.7_2 | LRRN4 | 2.293045787 | 4.900896867 | 0.000923861 | 0.050325601 |
| ENSG00000185664.14_4 | PMEL | 2.101598092 | 4.291845351 | 0.041762031 | 0.189378261 |
| ENSG00000138080.13_2 | EMILIN1 | 2.086381193 | 4.246814794 | 0.00021368 | 0.033201528 |
| ENSG00000185551.14_3 | NR2F2 | 2.049419436 | 4.139393602 | 0.015808124 | 0.128550994 |
| ENSG00000122691.12_3 | TWIST1 | 1.93767715 | 3.830883493 | 0.001135438 | 0.052593016 |
| ENSG00000116132.11_3 | PRRX1 | 1.87077789 | 3.657297255 | 0.001296592 | 0.0538418 |
| ENSG00000178252.17_3 | WDR6 | 1.840542095 | 3.581445765 | 0.034485201 | 0.17438591 |
| ENSG00000106333.12_2 | PCOLCE | 1.800690899 | 3.483870262 | 0.001220195 | 0.053665168 |
| ENSG00000163349.21_3 | HIPK1 | 1.79732213 | 3.475744731 | 0.002155682 | 0.063582685 |
| ENSG00000158270.11_3 | COLEC12 | 1.783032855 | 3.441488896 | 0.048515744 | 0.201942638 |
| ENSG00000171161.12_2 | ZNF672 | 1.753629373 | 3.372058052 | 0.034188418 | 0.173644775 |
| ENSG00000022556.15_3 | NLRP2 | 1.711161075 | 3.27424227 | 0.001155976 | 0.052593016 |
| ENSG00000255690.2_2 | TRIL | 1.63287963 | 3.101314059 | 0.007051968 | 0.096262957 |
| ENSG00000136383.6_2 | ALPK3 | 1.590057032 | 3.010612508 | 0.000466886 | 0.041915559 |
| ENSG00000135903.18_2 | PAX3 | 1.577988651 | 2.98553328 | 0.036641362 | 0.178731018 |
| ENSG00000136110.12_2 | CNMD | 1.449604509 | 2.731331661 | 6.13992 × 10−5 | 0.022921908 |
| ENSG00000117114.19_3 | ADGRL2 | 1.439041207 | 2.711406098 | 0.031070439 | 0.165870885 |
| ENSG00000113196.2_2 | HAND1 | 1.418099884 | 2.672333167 | 0.008240986 | 0.102888602 |
| Track id | Gene Name | log2FC | Fold Change | p Value | q Value |
|---|---|---|---|---|---|
| ENSG00000126709.14_2 | IFI6 | −5.878845546 | 0.016993825 | 0.000233723 | 0.014261355 |
| ENSG00000137959.15_3 | IFI44L | −5.608066648 | 0.020502354 | 3.82253 × 10−7 | 0.002122525 |
| ENSG00000185745.9_2 | IFIT1 | −5.254460452 | 0.026196893 | 6.57308 × 10−5 | 0.00935849 |
| ENSG00000187608.8_4 | ISG15 | −5.097947283 | 0.029198797 | 0.000133271 | 0.011568582 |
| ENSG00000111335.12_2 | OAS2 | −4.980723077 | 0.031670357 | 6.11792 × 10−7 | 0.002547809 |
| ENSG00000157601.13_3 | MX1 | −4.796144819 | 0.035992876 | 2.11695 × 10−6 | 0.00320583 |
| ENSG00000137965.10_2 | IFI44 | −4.795653196 | 0.036005143 | 0.00088056 | 0.02125452 |
| ENSG00000111331.12_2 | OAS3 | −4.672670264 | 0.039209029 | 3.48892 × 10−6 | 0.00339028 |
| ENSG00000119917.13_2 | IFIT3 | −4.576066362 | 0.041924391 | 6.18918 × 10−5 | 0.009288229 |
| ENSG00000169429.10_2 | CXCL8 | −4.495950675 | 0.044318391 | 9.12997 × 10−6 | 0.005849499 |
| ENSG00000089127.12_3 | OAS1 | −4.442799771 | 0.045981592 | 8.61343 × 10−5 | 0.01017607 |
| ENSG00000205413.7_3 | SAMD9 | −4.439354828 | 0.046091521 | 1.7325 × 10−5 | 0.007195438 |
| ENSG00000197632.8_3 | SERPINB2 | −4.215735621 | 0.053819186 | 3.44386 × 10−6 | 0.00339028 |
| ENSG00000169245.5_2 | CXCL10 | −4.091454118 | 0.058661017 | 7.72479 × 10−5 | 0.009748454 |
| ENSG00000173391.8_3 | OLR1 | −4.085638109 | 0.058897977 | 0.002476522 | 0.034021507 |
| ENSG00000185885.15_3 | IFITM1 | −4.045497159 | 0.060559741 | 0.006731331 | 0.05470077 |
| ENSG00000166741.7_3 | NNMT | −3.934604298 | 0.065398244 | 8.57421 × 10−5 | 0.01017607 |
| ENSG00000163739.4_2 | CXCL1 | −3.93287656 | 0.06547661 | 0.000384192 | 0.016613744 |
| ENSG00000184979.9_2 | USP18 | −3.849155456 | 0.0693887 | 3.94644 × 10−5 | 0.008176704 |
| ENSG00000125730.16_3 | C3 | −3.733039094 | 0.075204401 | 0.00197069 | 0.030365633 |
| ENSG00000165704.14_2 | HPRT1 | −2.039546227 | 0.243240232 | 8.9983 × 10−5 | 0.010363433 |
| Track id | Gene Name | log2FC | Fold Change | p Value | q Value |
|---|---|---|---|---|---|
| ENSG00000187634.11_4 | SAMD11 | 3.317151755 | 9.966947676 | 0.000104619 | 0.010578182 |
| ENSG00000173110.7_3 | HSPA6 | 3.089883973 | 8.514276679 | 0.004120717 | 0.042794832 |
| ENSG00000179348.11_3 | GATA2 | 2.793936011 | 6.935192898 | 0.001796619 | 0.028994304 |
| ENSG00000159184.7_2 | HOXB13 | 2.63230707 | 6.200166978 | 0.044819433 | 0.145877709 |
| ENSG00000118432.12_3 | CNR1 | 2.389582626 | 5.24005744 | 0.01782143 | 0.088564849 |
| ENSG00000170689.9_3 | HOXB9 | 2.343247752 | 5.074436947 | 0.020976437 | 0.09589378 |
| ENSG00000158164.6_2 | TMSB15A | 2.307040628 | 4.948669273 | 0.015467662 | 0.082319589 |
| ENSG00000159182.4_2 | PRAC1 | 2.197080343 | 4.585504101 | 0.007224322 | 0.056393045 |
| ENSG00000111341.9_2 | MGP | 2.182372513 | 4.538993785 | 0.007587234 | 0.057716822 |
| ENSG00000196361.9_3 | ELAVL3 | 2.162321548 | 4.47634598 | 0.002707677 | 0.035265434 |
| ENSG00000124194.16_3 | GDAP1L1 | 2.132951579 | 4.386139141 | 0.028791242 | 0.114518746 |
| ENSG00000120068.6_3 | HOXB8 | 2.107315751 | 4.308888453 | 0.004464332 | 0.044718486 |
| ENSG00000120075.5_3 | HOXB5 | 2.087021759 | 4.248700824 | 0.007689973 | 0.058020496 |
| ENSG00000162188.5_2 | GNG3 | 2.045937 | 4.129413815 | 0.047246979 | 0.150407077 |
| ENSG00000185559.13_2 | DLK1 | 1.998043763 | 3.994579836 | 0.011308426 | 0.069531367 |
| ENSG00000015592.16_2 | STMN4 | 1.986280174 | 3.962140869 | 0.034155545 | 0.125871793 |
| ENSG00000244242.1_3 | IFITM10 | 1.975606839 | 3.932936354 | 0.024756046 | 0.1053043 |
| ENSG00000142694.6_2 | EVA1B | 1.952401525 | 3.870182302 | 0.004311137 | 0.043861073 |
| ENSG00000108511.9_3 | HOXB6 | 1.933700212 | 3.820337809 | 0.006540761 | 0.053992073 |
| ENSG00000102924.11_2 | CBLN1 | 1.903898543 | 3.742230812 | 0.018988379 | 0.091299955 |
| Differentiated Cells | |||||
|---|---|---|---|---|---|
| Track id | Gene Name | log2FC | Fold Change | p Value | q Value |
| ENSG00000159184.7_2 | HOXB13 | 2.63230707 | 6.200166978 | 0.044819433 | 0.145877709 |
| ENSG00000170689.9_3 | HOXB9 | 2.343247752 | 5.074436947 | 0.020976437 | 0.09589378 |
| ENSG00000120068.6_3 | HOXB8 | 2.107315751 | 4.308888453 | 0.004464332 | 0.044718486 |
| ENSG00000120075.5_3 | HOXB5 | 2.087021759 | 4.248700824 | 0.007689973 | 0.058020496 |
| ENSG00000108511.9_3 | HOXB6 | 1.933700212 | 3.820337809 | 0.006540761 | 0.053992073 |
| ENSG00000128645.14_3 | HOXD1 | 1.696198455 | 3.240459616 | 0.00085944 | 0.021008689 |
| ENSG00000162761.14_2 | LMX1A | 1.575401297 | 2.980183774 | 0.001391974 | 0.025863001 |
| ENSG00000128709.12_3 | HOXD9 | 1.285888058 | 2.438320987 | 0.002395687 | 0.03350744 |
| ENSG00000164853.8_2 | UNCX | 1.268550707 | 2.409194227 | 0.020622765 | 0.094951359 |
| ENSG00000185668.7_3 | POU3F1 | 1.262708621 | 2.399458102 | 0.000188344 | 0.013335661 |
| ENSG00000173917.10_2 | HOXB2 | 1.240553358 | 2.362891456 | 0.01081035 | 0.068082724 |
| ENSG00000198353.7_3 | HOXC4 | 1.189794616 | 2.281202654 | 0.001179911 | 0.024068111 |
| ENSG00000111249.13_2 | CUX2 | 1.18968407 | 2.281027863 | 0.012392041 | 0.072717219 |
| ENSG00000120093.11_3 | HOXB3 | 1.15246945 | 2.22294068 | 0.002591134 | 0.034893382 |
| ENSG00000128710.5_3 | HOXD10 | 1.088479998 | 2.126498737 | 0.000640547 | 0.019493284 |
| ENSG00000105996.6_2 | HOXA2 | 1.047701401 | 2.067233567 | 0.002503953 | 0.034301685 |
| ENSG00000182742.5_3 | HOXB4 | 0.965568029 | 1.952832254 | 0.001836083 | 0.029163542 |
| ENSG00000197576.13_3 | HOXA4 | 0.931164381 | 1.906814341 | 0.005248627 | 0.048384965 |
| ENSG00000170166.5_3 | HOXD4 | 0.911411045 | 1.88088422 | 0.045232538 | 0.14662067 |
| ENSG00000215612.7_4 | HMX1 | 0.887549812 | 1.850031469 | 0.034230417 | 0.126013322 |
| ENSG00000121297.6_2 | TSHZ3 | 0.848984055 | 1.801232052 | 0.016044223 | 0.083608498 |
| ENSG00000177045.7_2 | SIX5 | 0.814854123 | 1.759120271 | 0.031145206 | 0.119736175 |
| ENSG00000128714.5_2 | HOXD13 | 0.770195003 | 1.705500293 | 0.025529381 | 0.107248486 |
| ENSG00000128652.11_3 | HOXD3 | 0.748395572 | 1.679923541 | 0.005374364 | 0.048887126 |
| ENSG00000138136.6_2 | LBX1 | 0.679070809 | 1.601108203 | 0.009589715 | 0.064232198 |
| ENSG00000007372.21_4 | PAX6 | 0.597865349 | 1.513475529 | 0.013698394 | 0.077090491 |
| ENSG00000174306.21_3 | ZHX3 | −0.722305605 | 0.606128 | 0.017358037 | 0.08736421 |
| ENSG00000016082.14_2 | ISL1 | −0.728438166 | 0.60355696 | 0.00146414 | 0.026395725 |
| ENSG00000163064.6_2 | EN1 | −0.86515322 | 0.548988099 | 0.00161515 | 0.027390246 |
| ENSG00000168779.19_3 | SOX2 | −0.958672156 | 0.514530265 | 0.000571957 | 0.01890408 |
| ENSG00000167034.9_2 | NKX3-1 | −1.164739985 | 0.446044643 | 0.00321428 | 0.03831649 |
| ENSG00000167157.10_2 | PRRX2 | −1.184994519 | 0.439826209 | 0.014110056 | 0.078479236 |
| ENSG00000204531.17_3 | POU5F1 | −1.601000772 | 0.329648227 | 0.011198978 | 0.069170405 |
| Undifferentiated cells | |||||
| Track id | Gene Name | log2FC | Fold Change | pValue | q Value |
| ENSG00000116132.11_3 | PRRX1 | 1.87077789 | 3.657297255 | 0.001296592 | 0.0538418 |
| ENSG00000135903.18_2 | PAX3 | 1.577988651 | 2.98553328 | 0.036641362 | 0.178731018 |
| ENSG00000173917.10_2 | HOXB2 | 1.144447461 | 2.2106145 | 0.011703642 | 0.116641064 |
| ENSG00000105996.6_2 | HOXA2 | 0.842597953 | 1.793276503 | 0.030772937 | 0.165161471 |
| ENSG00000170577.7_2 | SIX2 | 0.820143793 | 1.765581959 | 0.003329635 | 0.071309576 |
| ENSG00000016082.14_2 | ISL1 | 0.816106138 | 1.760647552 | 0.008983186 | 0.106504497 |
| ENSG00000128645.14_3 | HOXD1 | 0.77142207 | 1.706951503 | 0.001941455 | 0.061046111 |
| ENSG00000128714.5_2 | HOXD13 | 0.693889659 | 1.617638971 | 0.00149567 | 0.057054801 |
| ENSG00000185668.7_3 | POU3F1 | 0.686390153 | 1.609251878 | 0.034548416 | 0.17438591 |
| ENSG00000179981.9_3 | TSHZ1 | 0.680204656 | 1.602367046 | 0.033564051 | 0.172691892 |
| ENSG00000128709.12_3 | HOXD9 | 0.640661693 | 1.559044052 | 0.003497545 | 0.072360604 |
| ENSG00000229544.8_2 | NKX1-2 | 0.627938656 | 1.545355389 | 0.014494861 | 0.125234663 |
| ENSG00000215612.7_4 | HMX1 | 0.591368116 | 1.506674858 | 0.000547214 | 0.044818464 |
| ENSG00000125816.4_2 | NKX2-4 | −0.687731407 | 0.620829317 | 0.002069811 | 0.061914637 |
| BP GO Terms Associated with Upregulated Genes | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| ID | Term | Count | Size | NumInt | NumTotal | p Value | Enrichment Score | Fold Enrichment | Gene Ratio |
| GO:0007275 | Multicellular organism development | 134 | 4012 | 328 | 34,863 | 2.41257 × 10−42 | 41.61751927 | 3.550052586 | 0.408536585 |
| GO:0048731 | System development | 124 | 3584 | 328 | 34,863 | 3.35951 × 10−40 | 39.47372364 | 3.677431539 | 0.37804878 |
| GO:0048856 | Anatomical structure development | 147 | 5284 | 328 | 34,863 | 7.75032 × 10−38 | 37.11068015 | 2.95695992 | 0.448170732 |
| GO:0009653 | Anatomical structure morphogenesis | 96 | 2241 | 328 | 34,863 | 1.48542 × 10−37 | 36.82814949 | 4.55323734 | 0.292682927 |
| GO:0032502 | Developmental process | 152 | 5896 | 328 | 34,863 | 1.67503 × 10−35 | 34.77597846 | 2.740166959 | 0.463414634 |
| GO:0048513 | Animal organ development | 105 | 2902 | 328 | 34,863 | 6.25251 × 10−35 | 34.20394575 | 3.845765536 | 0.320121951 |
| GO:0009790 | Embryo development | 58 | 1048 | 328 | 34,863 | 8.56326 × 10−28 | 27.06736094 | 5.882441584 | 0.176829268 |
| GO:0050896 | Response to stimulus | 169 | 8433 | 328 | 34,863 | 7.98053 × 10−27 | 26.09796836 | 2.130078047 | 0.515243902 |
| GO:0009888 | Tissue development | 72 | 1770 | 328 | 34,863 | 2.49522 × 10−26 | 25.60289146 | 4.323646135 | 0.219512195 |
| BP GO Terms Associated with Downregulated Genes | |||||||||
| ID | Term | Count | Size | NumInt | NumTotal | pvalue | Enrichment Score | Fold Enrichment | Gene Ratio |
| GO:0050896 | Response to stimulus | 196 | 8433 | 325 | 34,863 | 5.9275 × 10−44 | 43.22712839 | 2.493189941 | 0.603076923 |
| GO:0032502 | Developmental process | 146 | 5896 | 325 | 34,863 | 2.6317 × 10−32 | 31.57976388 | 2.656297881 | 0.449230769 |
| GO:0048856 | Anatomical structure development | 131 | 5284 | 325 | 34,863 | 2.72826 × 10−28 | 27.56411377 | 2.659438071 | 0.403076923 |
| GO:0023051 | Regulation of signaling | 100 | 3633 | 325 | 34,863 | 4.72001 × 10−24 | 23.32605739 | 2.95267738 | 0.307692308 |
| GO:0010033 | Response to organic substance | 83 | 2578 | 325 | 34,863 | 5.51134 × 10−24 | 23.25874279 | 3.453636092 | 0.255384615 |
| GO:0048583 | Regulation of response to stimulus | 109 | 4250 | 325 | 34,863 | 5.59901 × 10−24 | 23.25188883 | 2.751179729 | 0.335384615 |
| GO:0010646 | Regulation of cell communication | 99 | 3639 | 325 | 34,863 | 2.08179 × 10−23 | 22.68156228 | 2.918330902 | 0.304615385 |
| GO:0007154 | Cell communication | 123 | 5357 | 325 | 34,863 | 3.06338 × 10−23 | 22.51379851 | 2.463002542 | 0.378461538 |
| GO:0023052 | Signaling | 119 | 5200 | 325 | 34,863 | 3.14735 × 10−22 | 21.50205514 | 2.454850296 | 0.366153846 |
| BP GO Terms Associated with Upregulated Genes | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| ID | Term | Count | Size | NumInt | NumTotal | p Value | Enrichment Score | Fold Enrichment | Gene Ratio |
| GO:0002181 | Cytoplasmic translation | 82 | 124 | 763 | 37,582 | 2.0816 × 10−108 | 107.6815974 | 32.57223185 | 0.107470511 |
| GO:0051171 | Regulation of nitrogen compound metabolic process | 307 | 6046 | 763 | 37,582 | 1.2740 × 10−58 | 57,89482511 | 2.501068479 | 0.402359109 |
| GO:0032502 | Developmental process | 284 | 5851 | 763 | 37,582 | 3.48369 × 10−49 | 48.45796012 | 2.390801899 | 0.372214941 |
| GO:0007275 | Multicellular organism development | 227 | 3984 | 763 | 37,582 | 4.84291 × 10−49 | 48.31489359 | 2.806479522 | 0.29750983 |
| GO:0048856 | Anatomical structure development | 260 | 5201 | 763 | 37,582 | 1.55391 × 10−46 | 45.80857371 | 2.462304986 | 0.340760157 |
| GO:0051252 | Regulation of RNA metabolic process | 213 | 3761 | 763 | 37,582 | 3.35767 × 10−45 | 44.4739615 | 2.78953375 | 0.279161206 |
| GO:0048731 | System development | 204 | 3556 | 763 | 37,582 | 6.62305 × 10−44 | 43.17894171 | 2.825685125 | 0.267365662 |
| GO:0006355 | Regulation of DNA-templated transcription | 197 | 3399 | 763 | 37,582 | 7.00027 × 10−43 | 42.15488536 | 2.854765317 | 0.25819135 |
| GO:1903506 | Regulation of nucleic acid template transcription | 197 | 3401 | 763 | 37,582 | 7.63059 × 10−43 | 42,11744152 | 2.853086537 | 0.25819134 |
| BP GO Terms Associated with Downregulated Genes | |||||||||
| ID | Term | Count | Size | NumInt | NumTotal | pvalue | Enrichment Score | Fold Enrichment | Gene Ratio |
| GO:0050896 | Response to stimulus | 732 | 8346 | 1265 | 37,582 | 6.4624 × 10−172 | 171.189604 | 2.605685903 | 0.578656126 |
| GO:0010033 | Response to organic substance | 342 | 2712 | 1265 | 37,582 | 7.5572 × 10−108 | 107.1216417 | 3.746500402 | 0.270355731 |
| GO:0048583 | Regulation of response to stimulus | 422 | 4191 | 1265 | 37,582 | 1.3389 × 10−102 | 101.8732672 | 2.991466563 | 0.333596838 |
| GO:0032502 | Developmental process | 499 | 5851 | 1265 | 37,582 | 2.55294 × 10−97 | 96.59295944 | 2.533726946 | 0.394466403 |
| GO:0002376 | Immune system process | 293 | 2206 | 1265 | 37,582 | 1.58977 × 10−96 | 95.79866673 | 3.945949065 | 0.231620553 |
| GO:0006952 | Defense response | 234 | 1406 | 1265 | 37,582 | 3.95178 × 10−96 | 95.40320743 | 4.944471744 | 0.184980237 |
| GO:0044419 | Biological process involved in Interspecies interaction between organisms | 243 | 1567 | 1265 | 37,582 | 3.00479 × 10−93 | 92.52218606 | 4.607089401 | 0.192094862 |
| GO:0048856 | Anatomical structure development | 458 | 5201 | 1265 | 37,582 | 3.04307 × 10−92 | 91.51668855 | 2.616182203 | 0.362055336 |
| GO:0007154 | Cell communication | 460 | 5246 | 1265 | 37,582 | 4.31433 × 10−92 | 91.36508615 | 2.605067064 | 0.363636364 |
| BP GO Terms Associated with Upregulated Genes | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| ID | Term | Count | Size | NumInt | NumTotal | p Value | Enrichment Score | Fold Enrichment | Gene Ratio |
| GO:0007399 | Nervous system development | 136 | 2219 | 763 | 37,582 | 2.91618 × 10−31 | 30.53518564 | 3.018818178 | 0.178243775 |
| GO:0022008 | Neurogenesis | 88 | 1259 | 763 | 37,582 | 5.7406 × 10−24 | 23.24104242 | 3.442803948 | 0.115334207 |
| GO:0048699 | Generation of neurons | 79 | 1097 | 763 | 37,582 | 2.28169 × 10−22 | 21.64174417 | 3.547119452 | 0.103538663 |
| GO:0030182 | Neuron differentiation | 76 | 1027 | 763 | 37,582 | 3.00494 × 10−22 | 21.52216429 | 3.64500811 | 0.099606815 |
| GO:0007417 | Central nervous system development | 75 | 1091 | 763 | 37,582 | 4.18264 × 10−20 | 19.37854917 | 3.386038276 | 0.098296199 |
| GO:0007420 | Brain development | 60 | 832 | 763 | 37,582 | 3.11326 × 10−17 | 16.50678452 | 3.552084384 | 0.078636959 |
| GO:0048666 | Neuron development | 58 | 813 | 763 | 37,582 | 1.74529 × 10−16 | 15.75813215 | 3.513927511 | 0.076015727 |
| GO:0061564 | Axón development | 37 | 388 | 763 | 37,582 | 9.47529 × 10−15 | 14.0234076 | 4.69705179 | 0.048492792 |
| GO:0031175 | Neuron projection development | 48 | 648 | 763 | 37,582 | 1.87305 × 10−14 | 13.72745142 | 3.648560749 | 0.062909567 |
| BP GO Terms Associated with Downregulated Genes | |||||||||
| ID | Term | Count | Size | NumInt | NumTotal | pvalue | Enrichment Score | Fold Enrichment | Gene Ratio |
| GO:0007399 | Nervous system development | 173 | 2219 | 1265 | 37,582 | 2.83609 × 10−25 | 24.54727957 | 2.316211234 | 0.136758893 |
| GO:0022008 | Neurogenesis | 117 | 1259 | 1265 | 37,582 | 3.29975 × 10−23 | 22.48151933 | 2.760892483 | 0.092490119 |
| GO:0030182 | Neuron differentiation | 90 | 1027 | 1265 | 37,582 | 1.80993 × 10−16 | 15.74233731 | 2.603523059 | 0.071146245 |
| GO:0048699 | Generation of neurons | 93 | 1097 | 1265 | 37,582 | 4.41822 × 10−16 | 15.35475299 | 2.518637607 | 0.073517787 |
| GO:0048666 | Neuron development | 75 | 813 | 1265 | 37,582 | 4.53421 × 10−15 | 14.34349797 | 2.740691043 | 0.059288538 |
| GO:1901214 | Regulation of neuron death | 42 | 330 | 1265 | 37,582 | 1.76991 × 10−13 | 12.75204903 | 3.781157025 | 0.033201581 |
| GO:0007610 | Behavior | 60 | 611 | 1265 | 37,582 | 1.8082 × 10−13 | 12.74275385 | 2.917423003 | 0.04743083 |
| GO:0031175 | Neuron projection development | 57 | 648 | 1265 | 37,582 | 5.9181 × 10−11 | 10.22781749 | 2.613299663 | 0.045059289 |
| GO:0051960 | Regulation of nervous system development | 45 | 461 | 1265 | 37,582 | 2.32408 × 10−10 | 9.633748122 | 2.90001972 | 0.035573123 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres, R.J.; Valentines-Casas, G.; Cano-Estrada, C.; Ontiveros, N.; López, J.M. Gene Expression Analysis of HPRT-Deficient Cells Maintained with Physiological Levels of Folic Acid. Cells 2025, 14, 1105. https://doi.org/10.3390/cells14141105
Torres RJ, Valentines-Casas G, Cano-Estrada C, Ontiveros N, López JM. Gene Expression Analysis of HPRT-Deficient Cells Maintained with Physiological Levels of Folic Acid. Cells. 2025; 14(14):1105. https://doi.org/10.3390/cells14141105
Chicago/Turabian StyleTorres, Rosa J., Gerard Valentines-Casas, Claudia Cano-Estrada, Neus Ontiveros, and José M. López. 2025. "Gene Expression Analysis of HPRT-Deficient Cells Maintained with Physiological Levels of Folic Acid" Cells 14, no. 14: 1105. https://doi.org/10.3390/cells14141105
APA StyleTorres, R. J., Valentines-Casas, G., Cano-Estrada, C., Ontiveros, N., & López, J. M. (2025). Gene Expression Analysis of HPRT-Deficient Cells Maintained with Physiological Levels of Folic Acid. Cells, 14(14), 1105. https://doi.org/10.3390/cells14141105