
9-cis-Retinoic Acid Improves Disease Modelling in iPSC-Derived Liver Organoids

 , , , ,
, , , , 

Abstract

1. Introduction
2. Materials and Methods
2.1. iPSC Culture Maintenance
2.2. Differentiation of iPSCs Towards Hepatic Stellate Cells (iHSCs)
2.3. Differentiation of iPSCs Towards Hepatocytes (iHepatocytes)
2.4. iPSC-Derived Liver Organoids
2.5. Isolation of pHSCs and Hepatocytes
2.6. pHSC/HepaRG Spheroids
2.7. Organoid Exposure to TGFβ and APAP
2.8. Cell Viability Assay
2.9. RNA Extraction and RT-qPCR
2.10. Immunofluorescence Stainings
2.11. Picrosirius Staining
2.12. Flow Cytometry Analysis
2.13. CYP3A4 Activity Assay
2.14. RNA Sequencing and Analysis
2.15. Statistical Analysis
3. Results
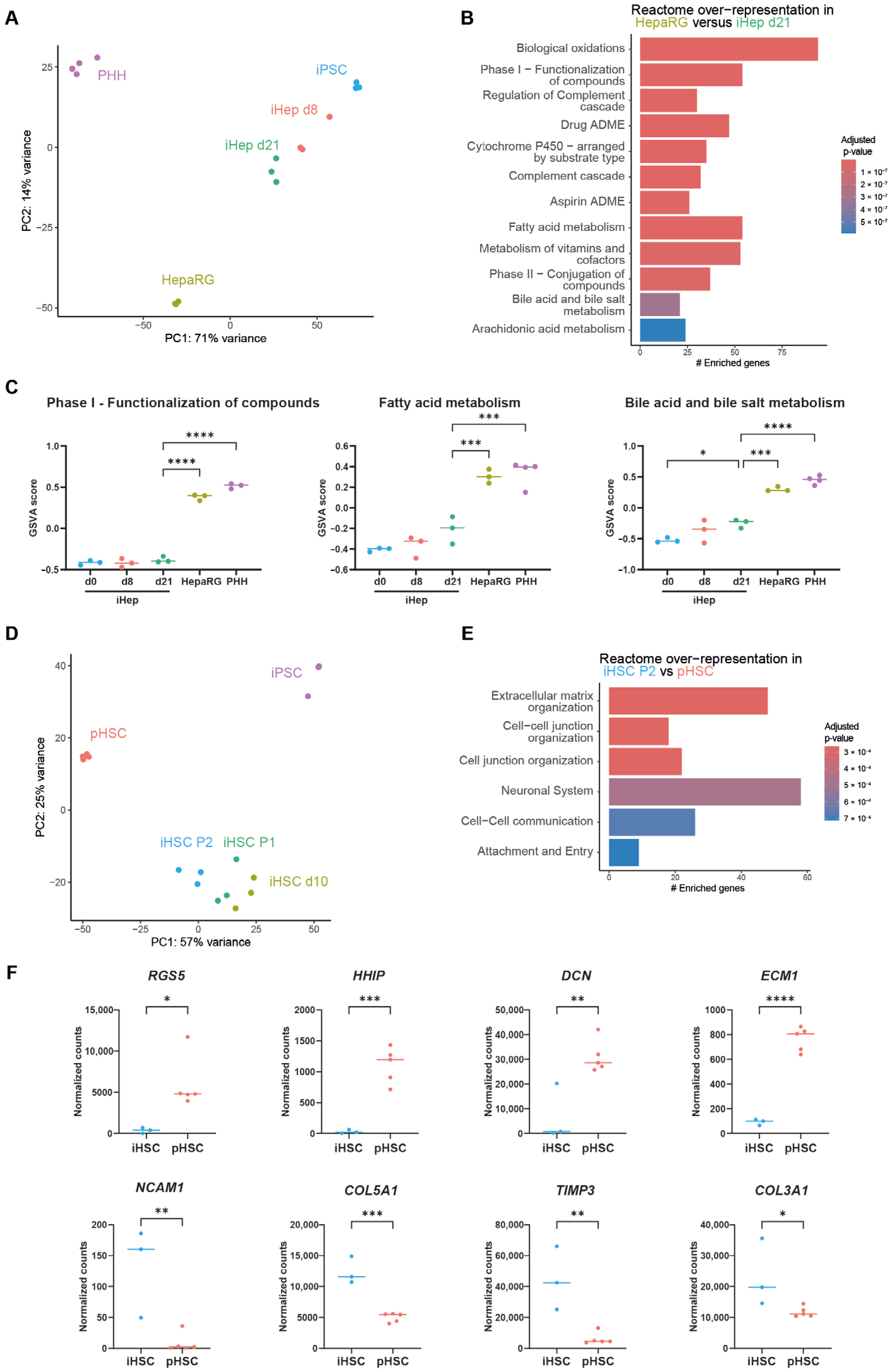
3.1. iPSC-Derived Liver Organoids Are More Fibrotic than HepaRG/pHSC Organoids
3.2. Immaturity of iPSC-Derived Liver Cells
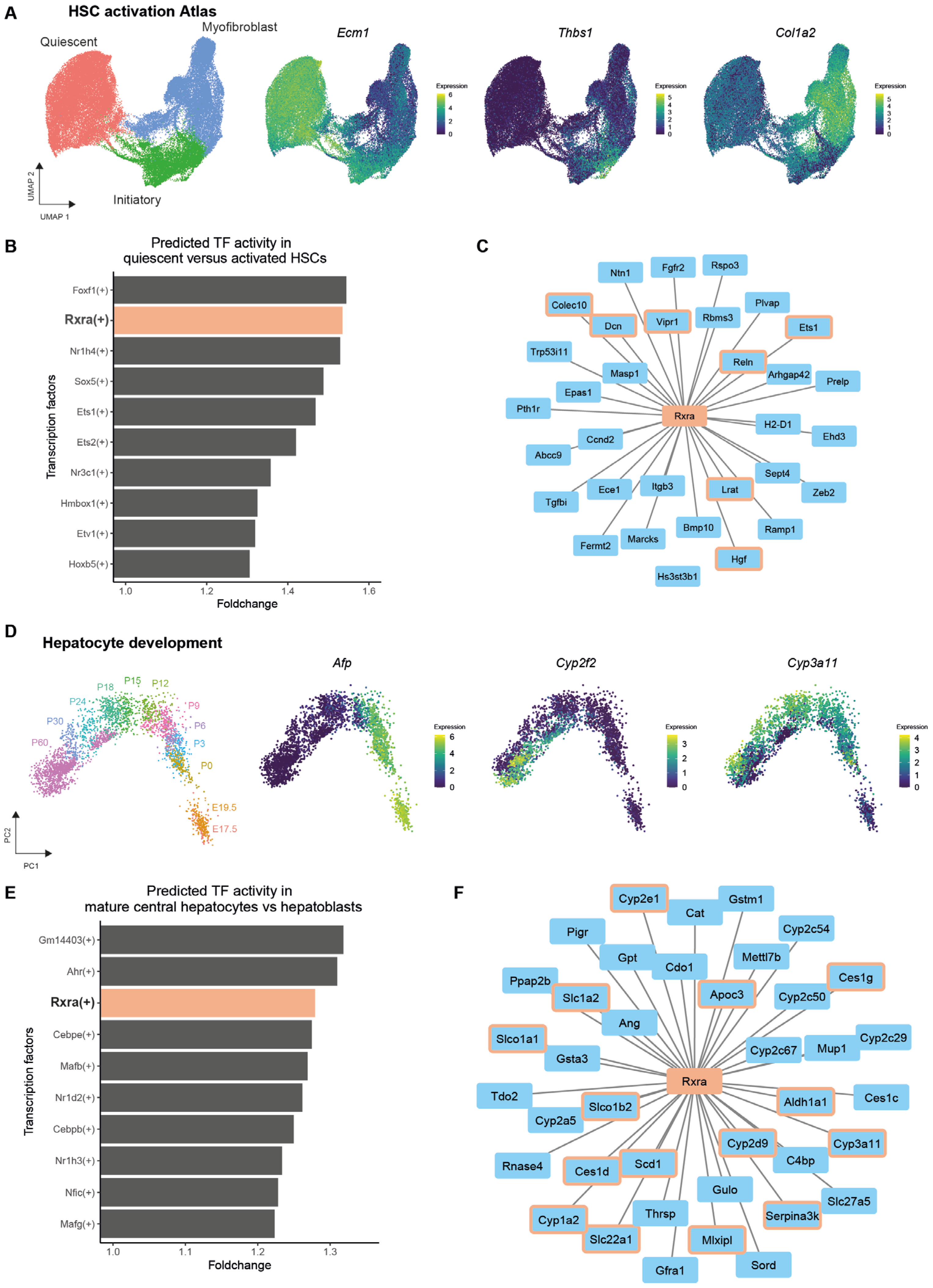
3.3. RXRA as a Target for Improving iPSC-Derived Liver Cells
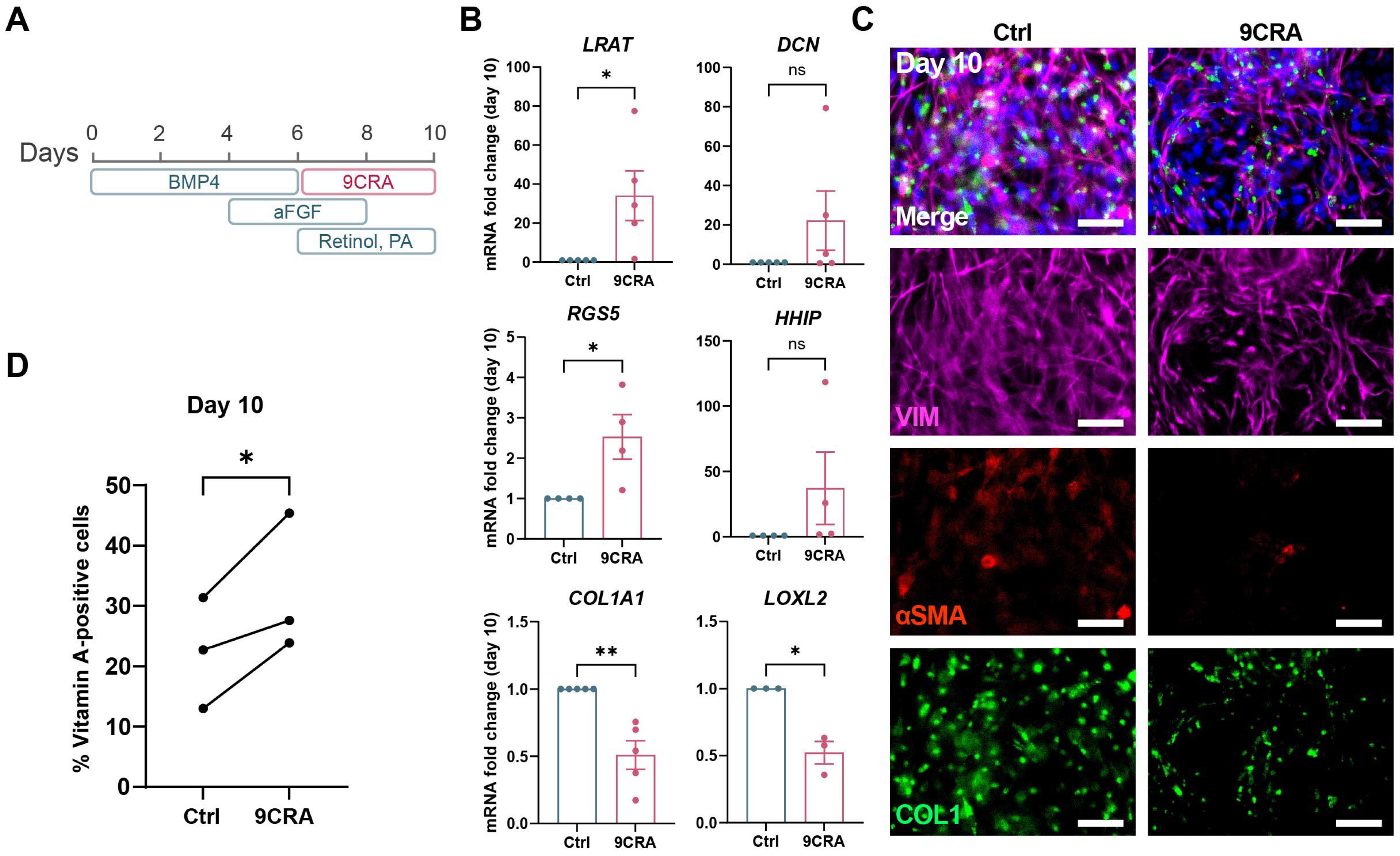
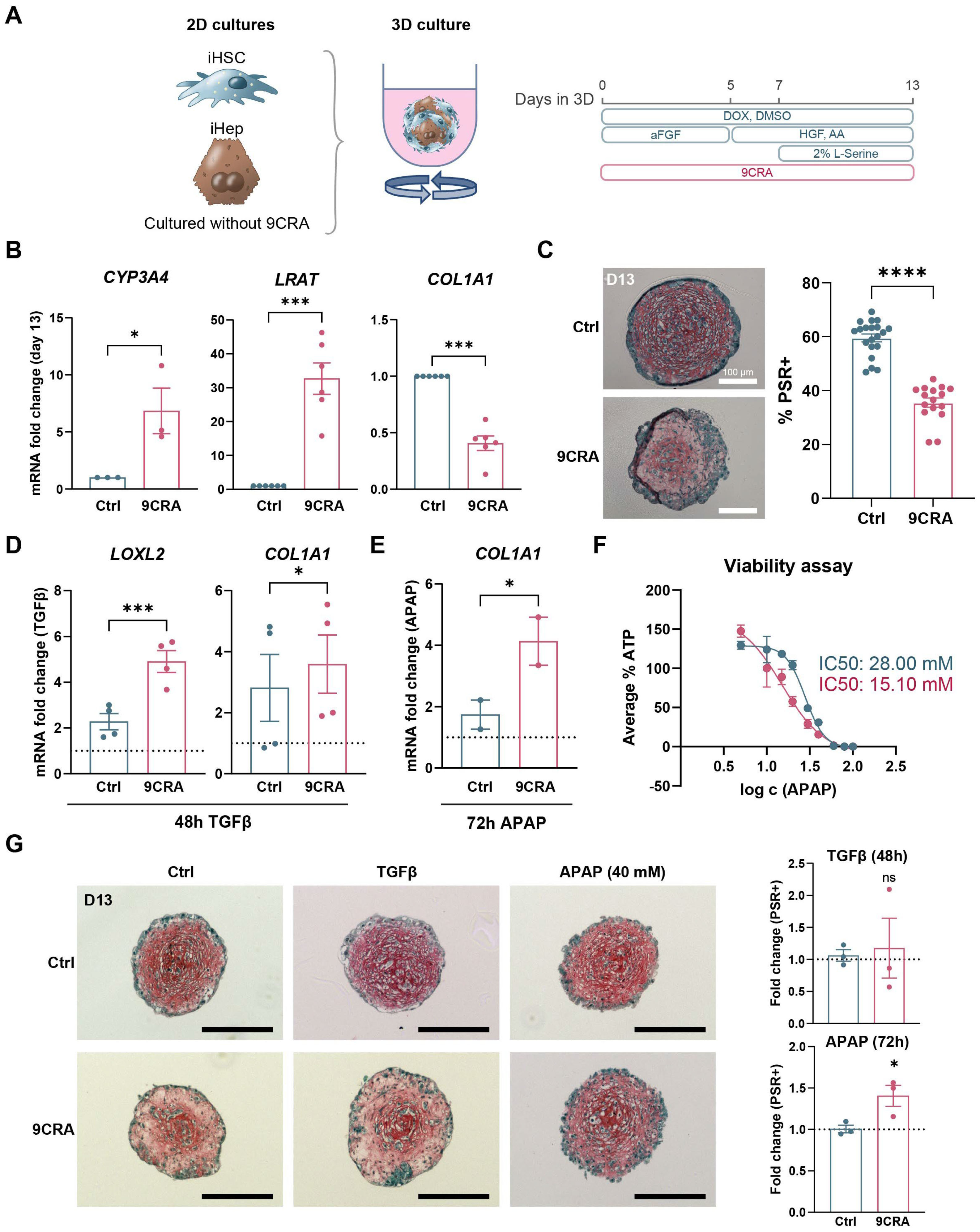
3.4. 9-cis-Retinoic Acid Treatment Improves HSC Differentiation from iPSCs
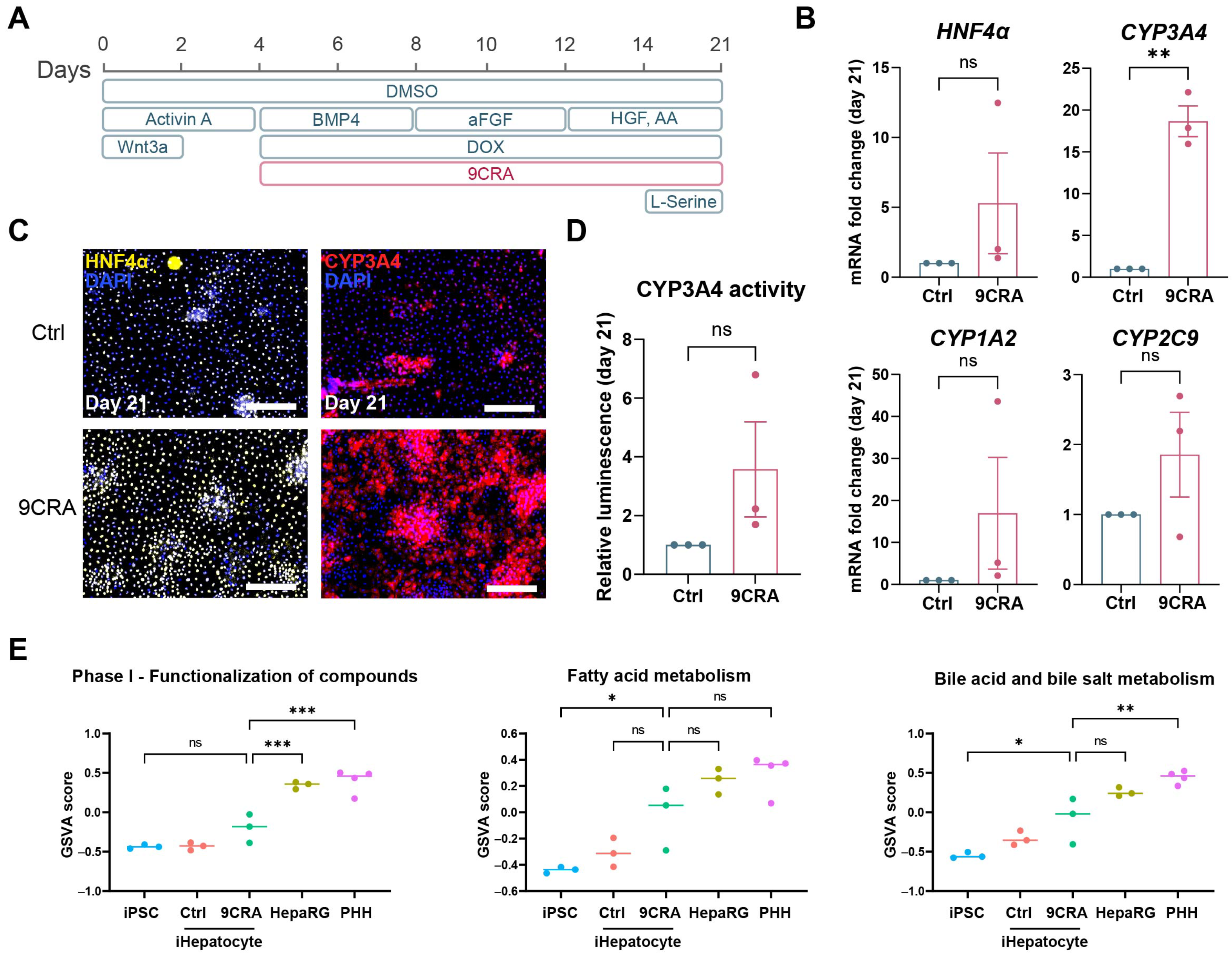
3.5. 9-cis-Retinoic Acid Makes iHepatocytes More Mature
3.6. 9-cis-Retinoic Acid Improves iPSC-Derived Liver Fibrosis Modelling
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HSC | Hepatic stellate cell |
| iPSC | Induced pluripotent stem cell |
| iHep | Induced pluripotent stem cell-derived hepatoblast |
| pHSC | Primary human hepatic stellate cell |
| iHSC | Induced pluripotent stem cell-derived hepatic stellate cell |
| iHepatocyte | Induced pluripotent stem cell-derived hepatocyte |
| 9CRA | 9-cis-retinoic acid |
| LDM | Liver differentiation medium |
| DMSO | Dimethyl sulfoxide |
| DOX | Doxycycline hyclate |
| TGFβ | Transforming growth factor beta |
| APAP | Acetaminophen |
| BSA | Bovine serum albumin |
| FBS | Foetal bovine serum |
| GSVA | Gene set variation analysis |
| RXRA | Retinoid X receptor alpha |
References
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of liver diseases in the world. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef]
- Tsuchida, T.; Friedman, S.L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Van Grunsven, L.A. 3D in vitro models of liver fibrosis. Adv. Drug Deliv. Rev. 2017, 121, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Boon, R.; Kumar, M.; Tricot, T.; Elia, I.; Ordovas, L.; Jacobs, F.; One, J.; De Smedt, J.; Eelen, G.; Bird, M.; et al. Amino acid levels determine metabolism and CYP450 function of hepatocytes and hepatoma cell lines. Nat. Commun. 2020, 11, 1393. [Google Scholar] [CrossRef] [PubMed]
- Coll, M.; Perea, L.; Boon, R.; Leite, S.B.; Vallverdu, J.; Mannaerts, I.; Smout, A.; El Taghdouini, A.; Blaya, D.; Rodrigo-Torres, D.; et al. Generation of Hepatic Stellate Cells from Human Pluripotent Stem Cells Enables In Vitro Modeling of Liver Fibrosis. Cell Stem Cell 2018, 23, 101–113.E7. [Google Scholar] [CrossRef]
- Wu, X.; Jiang, D.; Yang, Y.; Li, S.; Ding, Q. Modeling drug-induced liver injury and screening for anti-hepatofibrotic compounds using human PSC-derived organoids. Cell Regen. 2023, 12, 6. [Google Scholar] [CrossRef]
- Cools, L.; Dastjerd, M.K.; Smout, A.; Merens, V.; Yang, Y.; Reynaert, H.; Messaoudi, N.; Smet, V.; Kumar, M.; Verhulst, S.; et al. Human iPSC-derived liver co-culture spheroids to model liver fibrosis. Biofabrication 2024, 16, 3. [Google Scholar] [CrossRef]
- Ouchi, R.; Togo, S.; Kimura, M.; Shinozawa, T.; Koido, M.; Koike, H.; Thompson, W.; Karns, R.A.; Mayhew, C.N.; McGrath, P.S.; et al. Modeling Steatohepatitis in Humans with Pluripotent Stem Cell-Derived Organoids. Cell Metab. 2019, 30, 374–384.E6. [Google Scholar] [CrossRef]
- Leite, S.B.; Roosens, T.; El Taghdouini, A.; Mannaerts, I.; Smout, A.J.; Najimi, M.; Sokal, E.; Noor, F.; Chesne, C.; van Grunsven, L.A. Novel human hepatic organoid model enables testing of drug-induced liver fibrosis in vitro. Biomaterials 2016, 78, 1–10. [Google Scholar] [CrossRef]
- Bushnell, B. BBMap: A Fast, Accurate, Splice-Aware Aligner; Lawrence Berkeley National Laboratory, LBNL Report #, LBNL-7065E; LBL Publications: Berkeley, CA, USA, 2014. [Google Scholar]
- Pouzat, C.; Chaffiol, A. Automatic spike train analysis and report generation. An implementation with R, R2HTML and STAR. J. Neurosci. Methods 2009, 181, 119–144. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Hanzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef]
- Hafemeister, C.; Satija, R. Normalization and variance stabilization of single-cell RNA-seq data using regularized negative binomial regression. Genome Biol. 2019, 20, 296. [Google Scholar] [CrossRef]
- Merens, V.; Knetemann, E.; Gurbuz, E.; De Smet, V.; Messaoudi, N.; Reynaert, H.; Verhulst, S.; van Grunsven, L.A. Hepatic stellate cell single cell atlas reveals a highly similar activation process across liver disease aetiologies. JHEP Rep. 2025, 7, 101223. [Google Scholar] [CrossRef]
- Yang, L.; Wang, X.; Zheng, J.X.; Xu, Z.R.; Li, L.C.; Xiong, Y.L.; Zhou, B.C.; Gao, J.; Xu, C.R. Determination of key events in mouse hepatocyte maturation at the single-cell level. Dev. Cell 2023, 58, 1996–2010.E6. [Google Scholar] [CrossRef]
- Van de Sande, B.; Flerin, C.; Davie, K.; De Waegeneer, M.; Hulselmans, G.; Aibar, S.; Seurinck, R.; Saelens, W.; Cannoodt, R.; Rouchon, Q.; et al. A scalable SCENIC workflow for single-cell gene regulatory network analysis. Nat. Protoc. 2020, 15, 2247–2276. [Google Scholar] [CrossRef]
- Sison-Young, R.L.; Lauschke, V.M.; Johann, E.; Alexandre, E.; Antherieu, S.; Aerts, H.; Gerets, H.H.J.; Labbe, G.; Hoet, D.; Dorau, M.; et al. A multicenter assessment of single-cell models aligned to standard measures of cell health for prediction of acute hepatotoxicity. Arch. Toxicol. 2017, 91, 1385–1400. [Google Scholar] [CrossRef]
- McGill, M.R.; Yan, H.M.; Ramachandran, A.; Murray, G.J.; Rollins, D.E.; Jaeschke, H. HepaRG cells: A human model to study mechanisms of acetaminophen hepatotoxicity. Hepatology 2011, 53, 974–982. [Google Scholar] [CrossRef]
- De Smet, V.; Gurbuz, E.; Eysackers, N.; Dewyse, L.; Smout, A.; Kazemzadeh Dastjerd, M.; Lefesvre, P.; Messaoudi, N.; Reynaert, H.; Verhulst, S.; et al. Orphan receptor GPR176 in hepatic stellate cells exerts a profibrotic role in chronic liver disease. JHEP Rep. 2024, 6, 101036. [Google Scholar] [CrossRef] [PubMed]
- Maepa, S.W.; Ndlovu, H. Advances in generating liver cells from pluripotent stem cells as a tool for modeling liver diseases. Stem Cells 2020, 38, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Schrooders, Y.; Hauser, D.; van Herwijnen, M.; Albrecht, W.; Ter Braak, B.; Brecklinghaus, T.; Castell, J.V.; Elenschneider, L.; Escher, S.; et al. Comparing in vitro human liver models to in vivo human liver using RNA-Seq. Arch. Toxicol. 2021, 95, 573–589. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Liu, T.; Chen, W.; Hammad, S.; Longerich, T.; Hausser, I.; Fu, Y.; Li, N.; He, Y.; Liu, C.; et al. ECM1 Prevents Activation of Transforming Growth Factor beta, Hepatic Stellate Cells, and Fibrogenesis in Mice. Gastroenterology 2019, 157, 1352–1367.e13. [Google Scholar] [CrossRef]
- Lian, N.; Jiang, Y.; Zhang, F.; Jin, H.; Lu, C.; Wu, X.; Lu, Y.; Zheng, S. Curcumin regulates cell fate and metabolism by inhibiting hedgehog signaling in hepatic stellate cells. Lab. Investig. 2015, 95, 790–803. [Google Scholar] [CrossRef]
- Baghy, K.; Iozzo, R.V.; Kovalszky, I. Decorin-TGFbeta axis in hepatic fibrosis and cirrhosis. J. Histochem. Cytochem. 2012, 60, 262–268. [Google Scholar] [CrossRef]
- Bahrami, A.J.; Gunaje, J.J.; Hayes, B.J.; Riehle, K.J.; Kenerson, H.L.; Yeung, R.S.; Stempien-Otero, A.S.; Campbell, J.S.; Mahoney, W.M., Jr. Regulator of G-protein signaling-5 is a marker of hepatic stellate cells and expression mediates response to liver injury. PLoS ONE 2014, 9, e108505. [Google Scholar] [CrossRef]
- Haaker, M.W.; Vaandrager, A.B.; Helms, J.B. Retinoids in health and disease: A role for hepatic stellate cells in affecting retinoid levels. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158674. [Google Scholar] [CrossRef]
- Kotulkar, M.; Robarts, D.R.; Apte, U. HNF4alpha in Hepatocyte Health and Disease. Semin. Liver Dis. 2023, 43, 234–244. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Graffmann, N.; Scherer, B.; Adjaye, J. In vitro differentiation of pluripotent stem cells into hepatocyte like cells—Basic principles and current progress. Stem Cell Res. 2022, 61, 102763. [Google Scholar] [CrossRef] [PubMed]
- Tate, B.F.; Levin, A.A.; Grippo, J.F. The discovery of 9-cis retinoic acid: A hormone that binds the retinoid-X receptor. Trends Endocrinol. Metab. 1994, 5, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Fadel, L.; Reho, B.; Volko, J.; Bojcsuk, D.; Kolostyak, Z.; Nagy, G.; Muller, G.; Simandi, Z.; Hegedus, E.; Szabo, G.; et al. Agonist binding directs dynamic competition among nuclear receptors for heterodimerization with retinoid X receptor. J. Biol. Chem. 2020, 295, 10045–10061. [Google Scholar] [CrossRef]
- Nagy, L.; Schwabe, J.W. Mechanism of the nuclear receptor molecular switch. Trends Biochem. Sci. 2004, 29, 317–324. [Google Scholar] [CrossRef]
- Gantier, M.; Rispal, R.; Fourrier, A.; Menoret, S.; Delbos, F.; Anegon, I.; Nguyen, T.H. Cryopreserved cGMP-compliant human pluripotent stem cell-derived hepatic progenitors rescue mice from acute liver failure through rapid paracrine effects on liver cells. Stem Cell Res. Ther. 2024, 15, 71. [Google Scholar] [CrossRef]
- Fourrier, A.; Delbos, F.; Menoret, S.; Collet, C.; Thi Thuy, L.T.; Myara, A.; Petit, F.; Tolosa, L.; Laplanche, S.; Gomez-Lechon, M.J.; et al. Regenerative cell therapy for the treatment of hyperbilirubinemic Gunn rats with fresh and frozen human induced pluripotent stem cells-derived hepatic stem cells. Xenotransplantation 2020, 27, e12544. [Google Scholar] [CrossRef]
- Shapiro, A.M.J.; Thompson, D.; Donner, T.W.; Bellin, M.D.; Hsueh, W.; Pettus, J.; Wilensky, J.; Daniels, M.; Wang, R.M.; Brandon, E.P.; et al. Insulin expression and C-peptide in type 1 diabetes subjects implanted with stem cell-derived pancreatic endoderm cells in an encapsulation device. Cell Rep. Med. 2021, 2, 100466. [Google Scholar] [CrossRef]
- Heyman, R.A.; Mangelsdorf, D.J.; Dyck, J.A.; Stein, R.B.; Eichele, G.; Evans, R.M.; Thaller, C. 9-cis retinoic acid is a high affinity ligand for the retinoid X receptor. Cell 1992, 68, 397–406. [Google Scholar] [CrossRef]
- Kane, M.A. Analysis, occurrence, and function of 9-cis-retinoic acid. Biochim. Biophys. Acta 2012, 1821, 10–20. [Google Scholar] [CrossRef]
- Yoo, H.S.; Moss, K.O.; Cockrum, M.A.; Woo, W.; Napoli, J.L. Energy status regulates levels of the RAR/RXR ligand 9-cis-retinoic acid in mammalian tissues: Glucose reduces its synthesis in beta-cells. J. Biol. Chem. 2023, 299, 105255. [Google Scholar] [CrossRef]
- Govaere, O.; Cockell, S.; Tiniakos, D.; Queen, R.; Younes, R.; Vacca, M.; Alexander, L.; Ravaioli, F.; Palmer, J.; Petta, S.; et al. Transcriptomic profiling across the nonalcoholic fatty liver disease spectrum reveals gene signatures for steatohepatitis and fibrosis. Sci. Transl. Med. 2020, 12, eaba4448. [Google Scholar] [CrossRef] [PubMed]
- Philpott, J.; Kazimierczyk, S.; Korgaonkar, P.; Bordt, E.; Zois, J.; Vasudevan, C.; Meng, D.; Bhatia, I.; Lu, N.; Jimena, B.; et al. RXRalpha Regulates the Development of Resident Tissue Macrophages. Immunohorizons 2022, 6, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Den Braver-Sewradj, S.P.; den Braver, M.W.; Vermeulen, N.P.; Commandeur, J.N.; Richert, L.; Vos, J.C. Inter-donor variability of phase I/phase II metabolism of three reference drugs in cryopreserved primary human hepatocytes in suspension and monolayer. Toxicol. In Vitro 2016, 33, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Le, T.V.; Diep, T.T.N.; Nhan, V.T.; Uyen, N.L.T.; Thanh, D.M.; Truong, N.H. A simple and effective scaffold for mouse hepatic stellate cell primary culture. Am. J. Physiol. Cell Physiol. 2023, 324, C1213–C1222. [Google Scholar] [CrossRef]
- Li, H.; Zhang, J.; Huang, G.; Zhang, N.; Chen, Q.; Zhang, X. Effect of retinoid kappa receptor alpha (RXRalpha) transfection on the proliferation and phenotype of rat hepatic stellate cells in vitro. Chin. Med. J. 2002, 115, 928–932. [Google Scholar]
- Sharvit, E.; Abramovitch, S.; Reif, S.; Bruck, R. Amplified inhibition of stellate cell activation pathways by PPAR-gamma, RAR and RXR agonists. PLoS ONE 2013, 8, e76541. [Google Scholar] [CrossRef]
- Panebianco, C.; Oben, J.A.; Vinciguerra, M.; Pazienza, V. Senescence in hepatic stellate cells as a mechanism of liver fibrosis reversal: A putative synergy between retinoic acid and PPAR-gamma signalings. Clin. Exp. Med. 2017, 17, 269–280. [Google Scholar] [CrossRef]
- Guan, Y.; Enejder, A.; Wang, M.; Fang, Z.; Cui, L.; Chen, S.Y.; Wang, J.; Tan, Y.; Wu, M.; Chen, X.; et al. A human multi-lineage hepatic organoid model for liver fibrosis. Nat. Commun. 2021, 12, 6138. [Google Scholar] [CrossRef]
- Tsang, H.Y.; Yi Lo, P.H.; Ho Lee, K.K. Generation of Liver Organoids from Human Induced Pluripotent Stem Cells as Liver Fibrosis and Steatosis Models. BioRxiv. 2021. Available online: https://www.researchgate.net/publication/352869786_Generation_of_liver_organoids_from_human_induced_pluripotent_stem_cells_as_liver_fibrosis_and_steatosis_models (accessed on 22 June 2025).
- Buechler, M.B.; Pradhan, R.N.; Krishnamurty, A.T.; Cox, C.; Calviello, A.K.; Wang, A.W.; Yang, Y.A.; Tam, L.; Caothien, R.; Roose-Girma, M.; et al. Cross-tissue organization of the fibroblast lineage. Nature 2021, 593, 575–579. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Antibodies | ||
| HNF4α (1/200) | Santa Cruz Biotechnology (Dallas, TX, USA) | Cat# Sc-8987 |
| CYP3A4 (1/200) | Cypex limited UK (Dundee, UK) | Cat# MA5-17064 |
| αSMA-Cy3 (1/100) | Merck (Darmstadt, Germany) | Cat# C6198 |
| VIM (1/100) | Dako (Glostrup, Denmark) | Cat# M0725 |
| COL1 (1/100) | SouthernBiotech (Birmingham, AL, USA) | Cat# 1310-01 |
| PDGFRβ (1/100) | Abcam (Cambridge, UK) | Cat# ab32570 |
| Donkey anti-Rabbit IgG (H + L) Alexa Fluor 647 (1/200) | Invitrogen (Waltham, MA, USA) | Cat# A31573 |
| Goat anti-Mouse IgG (H + L) Alexa Fluor 488 (1/200) | Invitrogen | Cat# A11029 |
| Goat anti-Rabbit IgG (H + L) Alexa Fluor 488 (1/200) | Invitrogen | Cat# A11008 |
| Donkey anti-Mouse IgG (H + L) Alexa Fluor 647 (1/200) | Thermo Fisher Scientific (Waltham, MA, USA) | Cat# A31571 |
| Donkey anti-Goat IgG (H + L) Alexa Fluor 488 (1/200) | Thermo Fisher Scientific | Cat# A11055 |
| Chemicals, peptides, and recombinant proteins | ||
| Biolaminin | Biolamina (Sundbyberg, Sweden) | Cat# LN521-05 |
| Essential 8 flex medium | Thermo Fisher Scientific | Cat# A2858501 |
| DPBS with calcium and magnesium | Thermo Fisher Scientific | Cat# 14040117 |
| StemPro Accutase Cell Dissociation Reagent | Thermo Fisher Scientific | Cat# A11105-01 |
| RevitaCell Supplement 100X | Gibco (Waltham, MA, USA) | Cat# A2644501 |
| mTeSR1 complete medium | Stemcell Technologies (Vancouver, BC, Canada) | Cat# 85850 |
| BMP | PeproTech (Cranbury, NJ, USA) | Cat# 120-05 |
| aFGF | PeproTech | Cat# 100-17A |
| Retinol | Merck Group | Cat# R7632 |
| Palmitic acid | Sigma-Aldrich (St. Louis, MO, USA) | Cat# P5585 |
| 9-cis-retinoic acid | Sigma-Aldrich | Cat# R4643 |
| Dimethyl sulfoxide | Merck Group | Cat# D2650 |
| Trypsin-EDTA 0.05% | Invitrogen | Cat# 25300-054 |
| Dulbecco’s modified medium | Capricorn (Ebsdorfergrund, Germany) | Cat# CA DMEM-HPA |
| HEPES Buffer solution | Capricorn | Cat# CA HEP-B |
| Penicillin-Streptomycin | Gibco | Cat# 15140122 |
| Foetal Bovine Serum | TICO Europe (Amstelveen, Netherlands) | Cat# FBSEU500 |
| Doxycycline hyclate | Merck Group | Cat# D9891 |
| Activin A | PeproTech | Cat# AF-120-14E |
| Wnt3a | PeproTech | Cat# 315-20 |
| HGF | PeproTech | Cat# 100-39 |
| Essential amino acids | Thermo Fisher Scientific | Cat# 11130036 |
| Non-essential amino acids | Thermo Fisher Scientific | Cat# 11140035 |
| L-Serine | Thermo Fisher Scientific | Cat# A11179.22 |
| Transforming growth factor beta | Peprotech | Cat# 100-21C |
| Acetaminophen | Merck Group | Cat# A7085 |
| IGL-1 organ preservation solution | Institut Georges Lopez (Lissieu, France) | Cat# IGU.IGL-1 REV07 |
| Pronase E | Merck | Cat# 1074330005 |
| Collagenase P | Roche (Basel, Switzerland) | Cat# 11 213873001 |
| Nycodenz gradient | Axis Shield Diagnostics (Dundee, UK) | Cat# 1002424 |
| DNase I | Roche | Cat# 10104159001 |
| Percoll gradient | GE Healthcare (Chicago, IL, USA) | Cat# GE17-0891-02 |
| Williams E medium | Gibco | Cat# A12176-01 |
| Glutamax supplement (200mM) | Thermo Fisher Scientific | Cat# 35050038 |
| EGF | PeproTech | Cat# 315-09 |
| Formaldehyde solution | Merck Group | Cat# 1004965000 |
| Triton-X100 | Merck Group | Cat# T8787 |
| Bovine serum albumin | Sigma-Aldrich | Cat# A2153 |
| Fluorescence Mounting Medium | Dako | Cat# S302380-2 |
| DAPI dilactate | Merck Group | Cat# D9564 |
| Xylene | VWR International (Radnor, PA, USA) | Cat# 28973.363 |
| 2-Propanol | Merck Group | Cat# 59300 |
| Methanol | VWR International | Cat# 20903.368 |
| Sirius Red | Sigma-Aldrich | Cat# 365548 |
| Fast Green FCF | Merck Group | Cat# F7258 |
| Picric acid solution | VWR International | Cat# 84512.260 |
| DPX mounting medium for histology | Merck Group | Cat# 06522 |
| Fixable viability stain 780 | BD Biosciences (Franklin Lakes, NJ, USA) | Cat# 565388 |
| Critical commercial assays | ||
| CellTiter-Glo Luminescent Cell Viability assay | Promega (Madison, WI, USA) | Cat# G7571 |
| ReliaPrep RNA Cell Miniprep System | Promega | Cat# Z6012 |
| M-MLV Reverse Transcriptase, RNase H Minus and Point Mutant | Promega | Cat# M5301 |
| GoTaq qPCR Master Mix with BRYT Green Dye | Promega | Cat# A6002 |
| P450-Glo CYP3A4 activity assay | Promega | Cat# V9001 |
| QuantSeq 3’ mRNA-Seq V2 Library Prep Kit FWD with UDI 12 nt set B1 | Lexogen (Vienna, Austria) | Cat# 192.96 |
| Deposited data | ||
| All RNAseq SRA files are available at NCBI under Password for reviewers onolugasdncbnyn This includes Normalized_counts and Raw_counts files | GEO ID: GSE292959 | |
| All other data files are deposited at Zenodo | 10.5281/zenodo.15095698 | |
| Experimental models: cell lines | ||
| iPSCs | Sigma-Aldrich | IPSC0028-1VL |
| HepaRG | Biopredic (Saint-Grégoire, France) | Cat# HPR116 |
| Oligonucleotides | ||
| DCN forward (CCAATATCACCAGCATTCCTC) | Integrated DNA Technologies (IDT, Coralville, IA, USA) | NA |
| DCN reverse (CTGCTGATTTTGTTGCCATC) | IDT | NA |
| LRAT forward (GCTGGGCTTTACCCCCTA) | IDT | NA |
| LRAT reverse (CGAATAATTATCTTCACAGTCTCACAA) | IDT | NA |
| COL3A1 forward (GGAGCTGGCTACTTCTCGC) | IDT | NA |
| COL3A1 reverse (GGGAACATCCTCCTTCAACAG) | IDT | NA |
| COL1A1 forward (CCGGCTCCTGCTCCTCTTAGCG) | IDT | NA |
| COL1A1 reverse (CGTTCTGTACGCAGGTGATTGGTGG) | IDT | NA |
| LOXL2 forward (GGAGAGGACATACAATACCAAAGTG) | IDT | NA |
| LOXL2 reverse (CCATGGAGAATGGCCAGTAG) | IDT | NA |
| ALB forward (TGGCACAATGAAGTGGGTAA) | IDT | NA |
| ALB reverse (CTGAGCAAAGGCAATCAACA) | IDT | NA |
| CYP3A4 forward (TTCCTCCCTGAAAGATTCAGC) | IDT | NA |
| CY3A4 reverse (GTTGAAGAAGTCCTCCTAAGCT) | IDT | NA |
| CYP1A2 forward (CTGGGCACTTCGACCCTTAC) | IDT | NA |
| CYP1A2 reverse (TCTCATCGCTACTCTCAGGGA) | IDT | NA |
| CYP2C9 forward (CAGTCCCTGCAGCTCTCTTT) | IDT | NA |
| CYP2C9 reverse (TGCACAGTGAAACATAGGAAACTC) | IDT | NA |
| HNF4α forward (CACGGGCAAACACTACGGT) | IDT | NA |
| HNF4α reverse (TTGACCTTCGAGTGCTGATCC) | IDT | NA |
| GAPDH forward (AGCCACATCGCTCAGACAC) | IDT | NA |
| GAPDH reverse (GCCCAATACGACCAAATCC) | IDT | NA |
| RPL19 forward (ATTGGTCTCATTGGGGTCTAAC) | IDT | NA |
| RPL19 reverse (AGTATGCTCAGGCTTCAGAAGA) | IDT | NA |
| RGS5 forward (TTGCATGTGCCAGAAAGCAG) | IDT | NA |
| RGS5 reverse (TACCCCTTGGGTTTCGATGC) | IDT | NA |
| HHIP forward (CCGAGGCCATATTCCAGGTTT) | IDT | NA |
| HHIP reverse (TGGAAAGCACAACCCACCAT) | IDT | NA |
| Software and algorithms | ||
| Graphpad | 10.3.1 | https://www.graphpad.com/ |
| Flowjo | 10 | https://www.flowjo.com/ |
| Qupath | 0.4.4 | https://qupath.github.io/ |
| R | 4.3.1 | www.r-project.org |
| DESeq2 | 1.40.2 | https://bioconductor.org/packages/release/bioc/html/DESeq2.html |
| clusterProfiler | 4.8.3 | https://bioconductor.org/packages/release/bioc/html/clusterProfiler.html |
| GSVA | 1.48.3 | https://bioconductor.org/packages/release/bioc/html/GSVA.html |
| Seurat | 4.1.0 | https://satijalab.org/seurat/ |
| Python | 3.7.7 | https://www.python.org/ |
| pySCENIC | 0.11.0 | https://pyscenic.readthedocs.io/en/latest/ |
| Other | ||
| 96-well ultra-low attachment U-bottom plates | Greiner Bio-One (Kremsmünster, Austria) | Cat# 650970 |
| ROTILABO round sieve, 75, 0.5 mm | Roth (Karlsruhe, Germany) | Cat# A624.1 |
| Glomax | Promega | Cat# GM3000 |
| Nanodrop 2000 Spectrophotometer | Thermo Fisher Scientific | Cat# G870 |
| QuantStudio 3 Real-Time PCR System | Applied Biosystems (Foster City, CA, USA) | Cat# 272310385 |
| EVOS | Thermo Fisher Scientific | Cat# AMF7000 |
| 40 µm cell strainer | Fisher Scientific (Geel, Belgium) | Cat# 11587522 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kazemzadeh Dastjerd, M.; Merens, V.; Smout, A.; De Wolf, R.; Chesné, C.; Verfaillie, C.; Verhulst, S.; van Grunsven, L.A. 9-cis-Retinoic Acid Improves Disease Modelling in iPSC-Derived Liver Organoids. Cells 2025, 14, 983. https://doi.org/10.3390/cells14130983
Kazemzadeh Dastjerd M, Merens V, Smout A, De Wolf R, Chesné C, Verfaillie C, Verhulst S, van Grunsven LA. 9-cis-Retinoic Acid Improves Disease Modelling in iPSC-Derived Liver Organoids. Cells. 2025; 14(13):983. https://doi.org/10.3390/cells14130983
Chicago/Turabian StyleKazemzadeh Dastjerd, Mina, Vincent Merens, Ayla Smout, Rebeca De Wolf, Christophe Chesné, Catherine Verfaillie, Stefaan Verhulst, and Leo A. van Grunsven. 2025. "9-cis-Retinoic Acid Improves Disease Modelling in iPSC-Derived Liver Organoids" Cells 14, no. 13: 983. https://doi.org/10.3390/cells14130983
APA StyleKazemzadeh Dastjerd, M., Merens, V., Smout, A., De Wolf, R., Chesné, C., Verfaillie, C., Verhulst, S., & van Grunsven, L. A. (2025). 9-cis-Retinoic Acid Improves Disease Modelling in iPSC-Derived Liver Organoids. Cells, 14(13), 983. https://doi.org/10.3390/cells14130983