
Anti-Inflammatory Peptide Prevents Aβ25–35-Induced Inflammation in Rats via Lipoxygenase Inhibition

 , , and
, , and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Synthesis of Peptides—Aβ25–35 and YWCS
2.2. Mass Spectrometric Analysis of Aβ25–35 and YWCS Peptides
2.3. Experimental Rats and Groups
2.4. Intracerebroventricular (i.c.v.) Injection of Aggregated Aβ25–35 Peptide
2.5. Intranasal Administration of the Peptide YWCS
2.6. Behavior Study
2.6.1. Open Field (OF)
2.6.2. Elevated Plus Maze (EPM)
2.6.3. Novel Object Recognition (NOR)
2.6.4. Morris Water Maze (MWM)
2.7. MRI of Brain Volumetric Analysis
2.8. Isolation of Brain Tissue and Processing
Tissue Isolation and Lysate Preparation
2.9. Immunoblotting
2.10. Tissue Processing for Cryosectioning
2.11. Immunofluorescence Staining
2.12. Quantification of LOX-5 and LOX-12 in Rat Serum by Surface Plasmon Resonance (SPR)
2.13. Statistical Analysis
3. Results
3.1. Synthesis and Mass Spectrometric Analysis of Aβ25–35 and YWCS Peptides
3.2. AD-Rat Model and Intranasal Treatment of Peptide Inhibitor of LOX
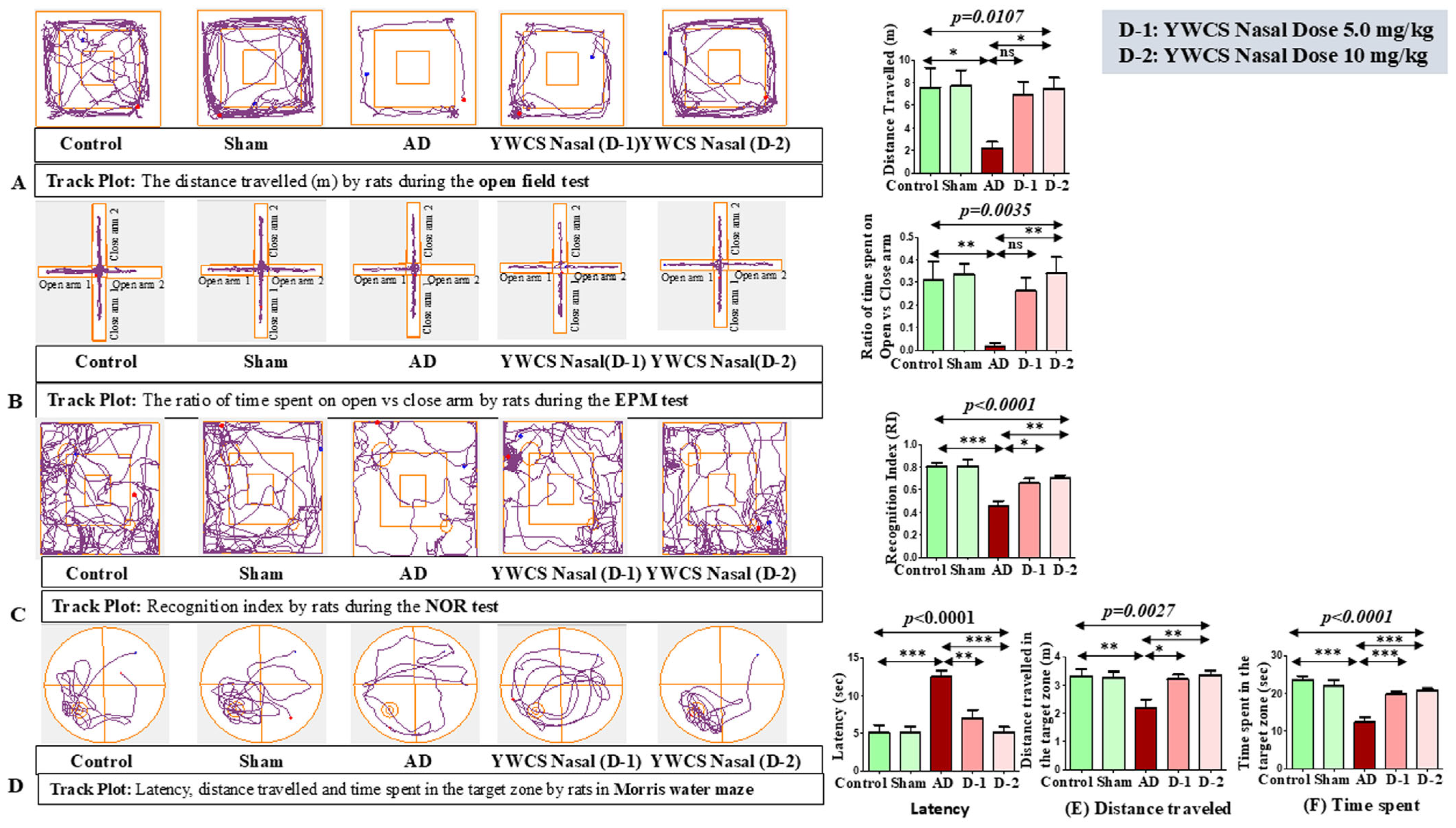
3.3. Behaviors Study
3.3.1. Open Field (OF)
3.3.2. Elevated Plus Maze (EPM)
3.3.3. Novel Object Recognition (NOR)
3.3.4. Morris Water Maze (MWM)
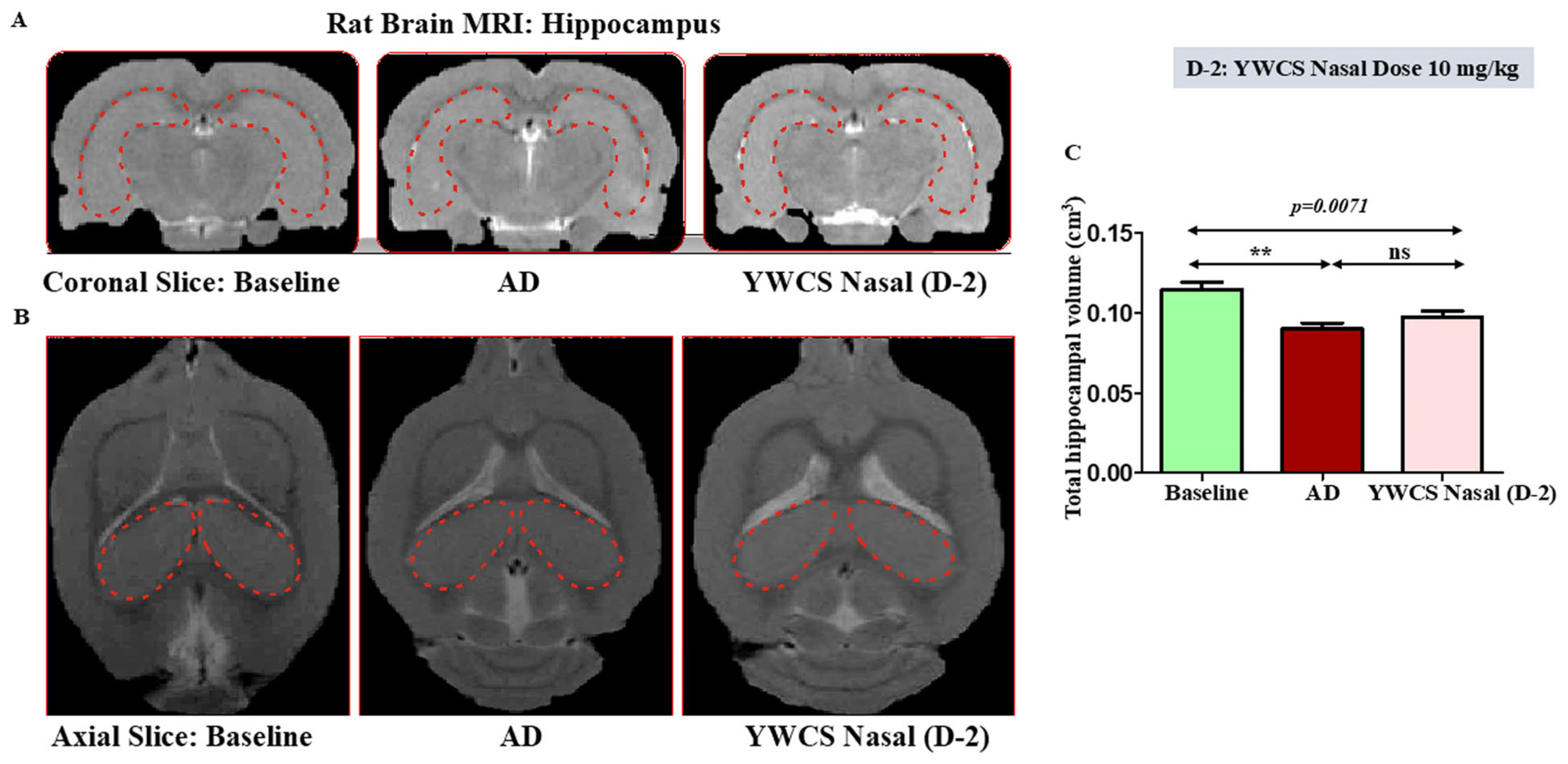
3.4. MRI Result
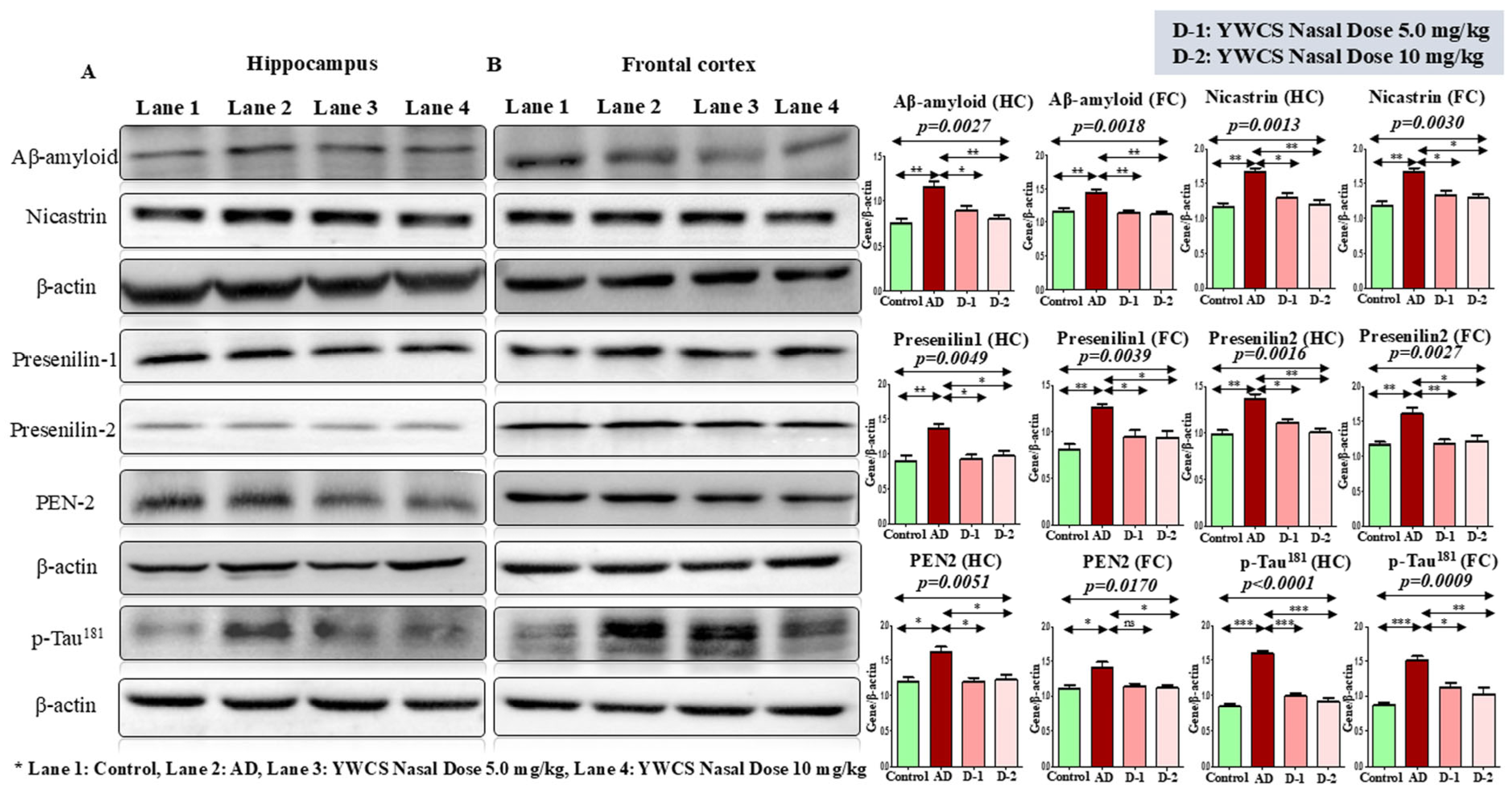
3.5. Protein Expression Level Regulated by the Inhibitor YWCS by Immunoblotting
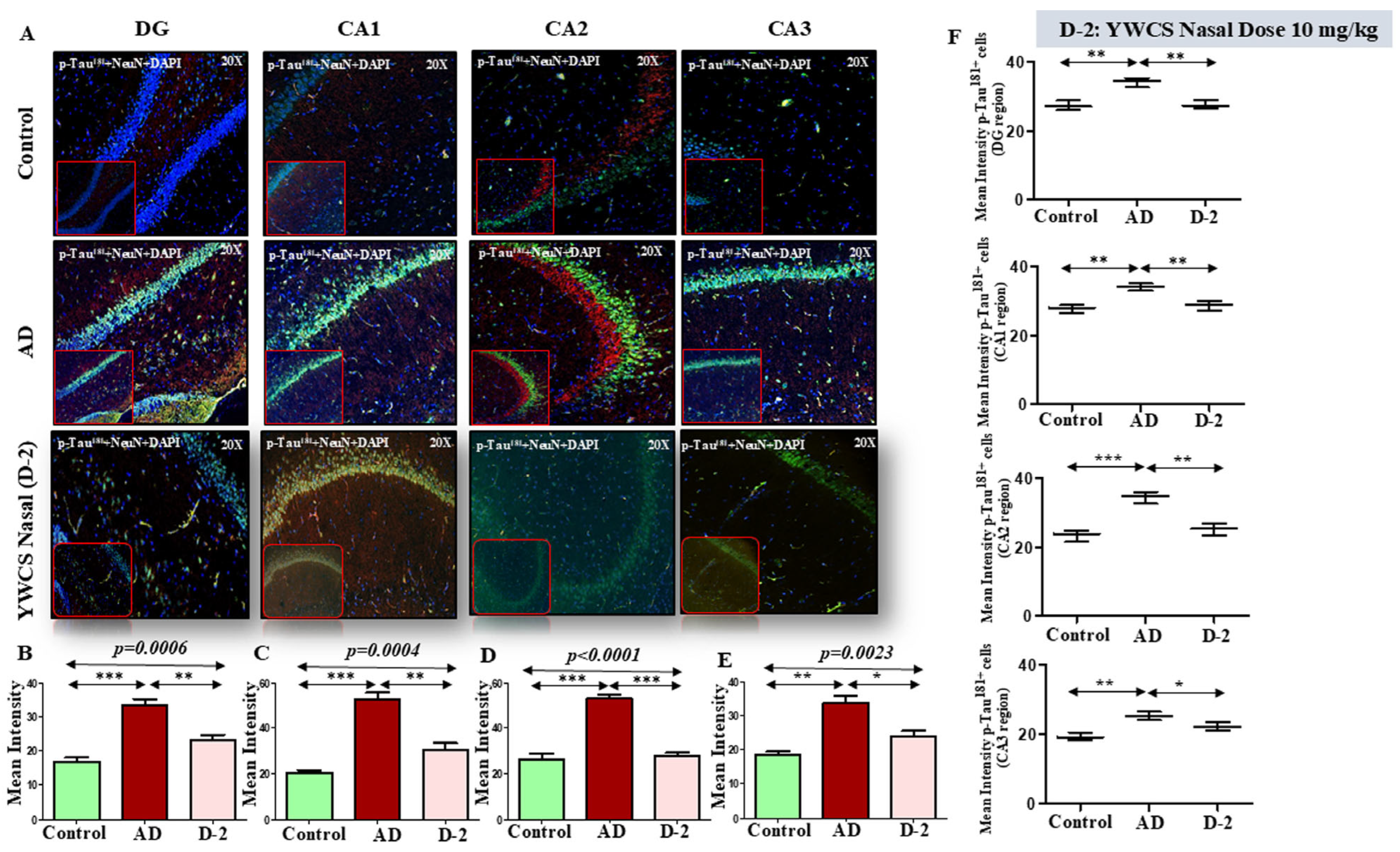
3.5.1. Expression Analysis of Aβ-Amyloid, γ-Secretase Components, and Phosphorylation of p-Tau181 by Western Blot
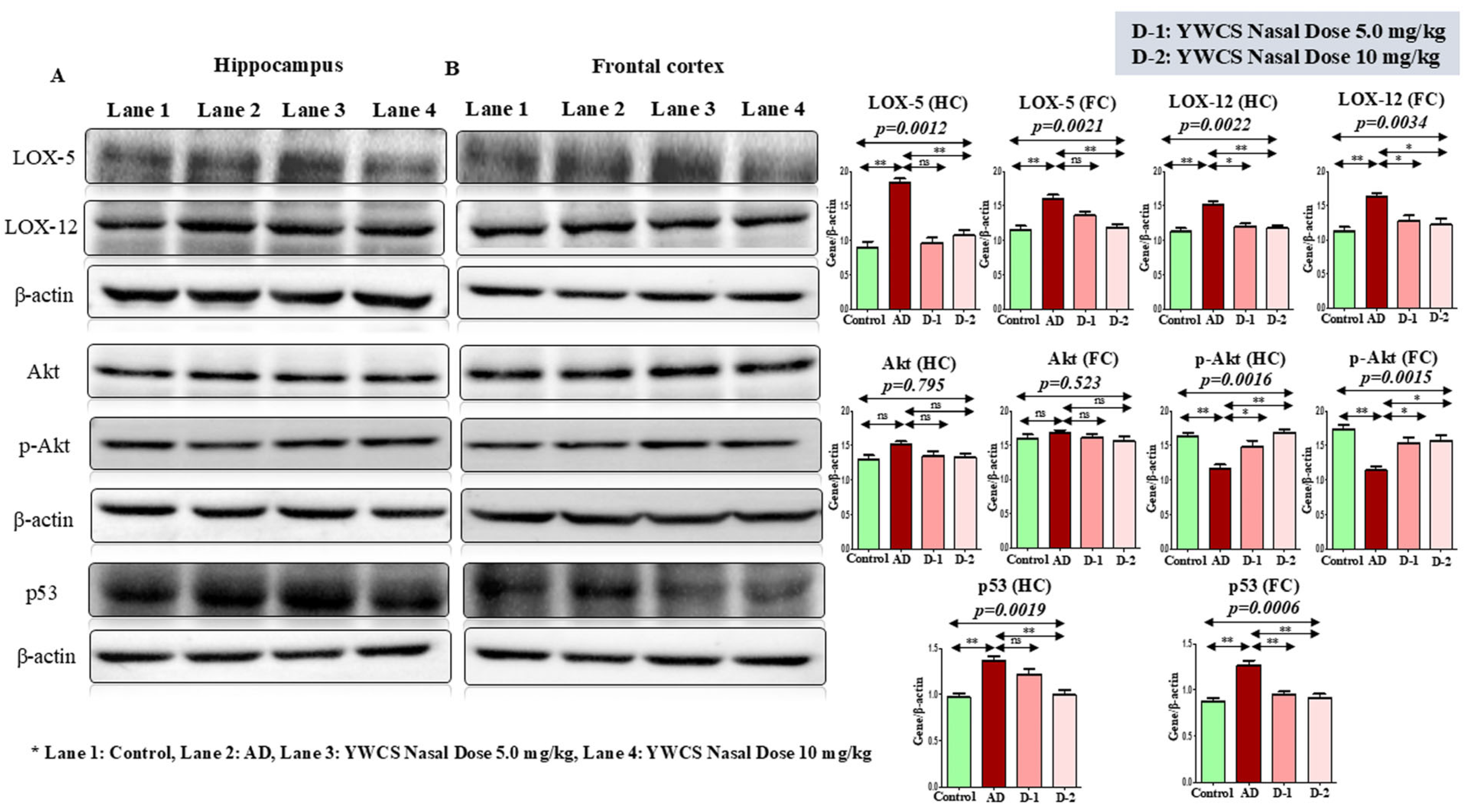
3.5.2. Effect of YWCS on Expression of LOX-5 and LOX-12
3.5.3. Effect of YWCS on Akt/p-Akt
AKT/p-AKT
3.5.4. Effect of YWCS on the Expression of p53
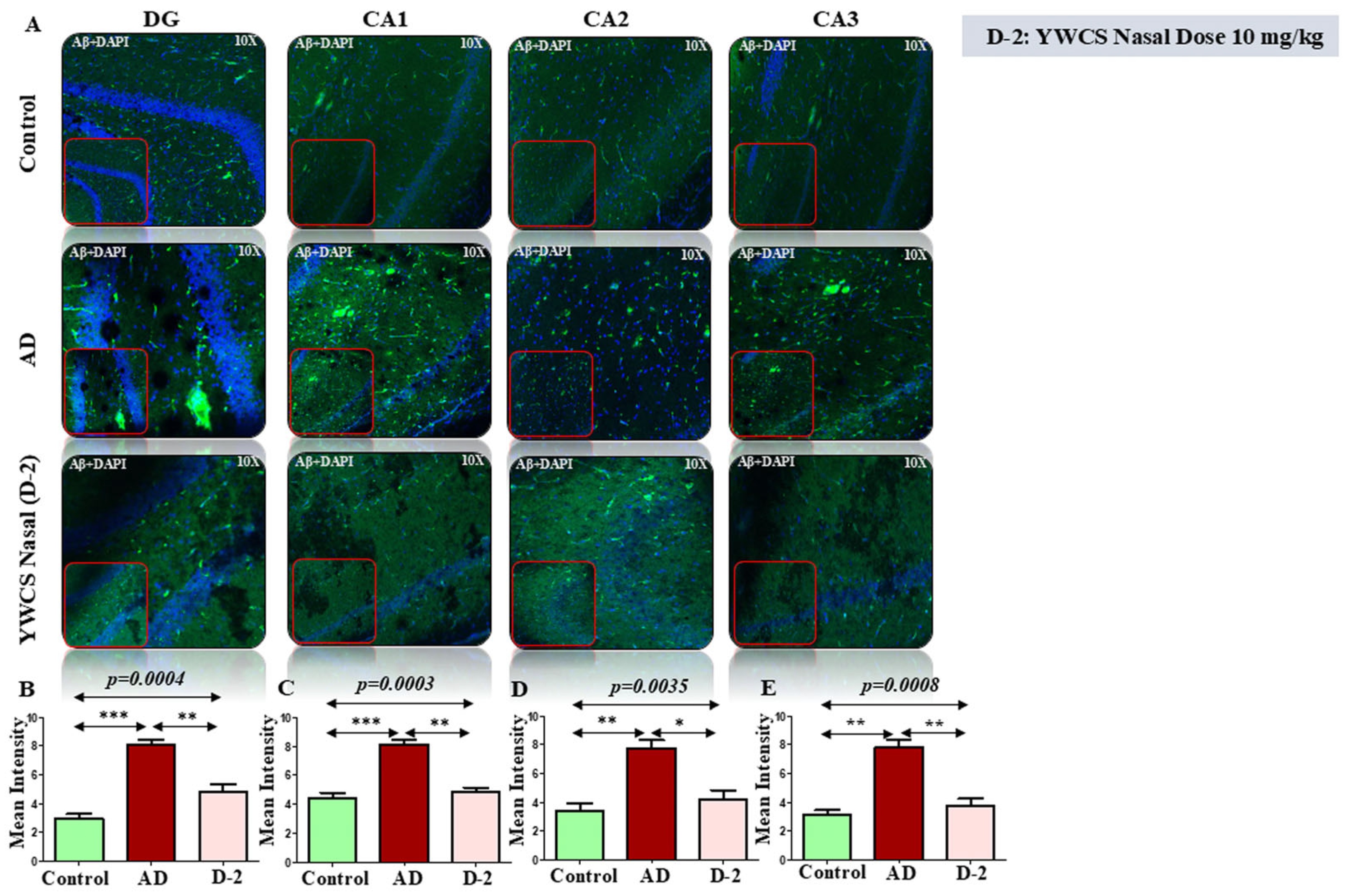
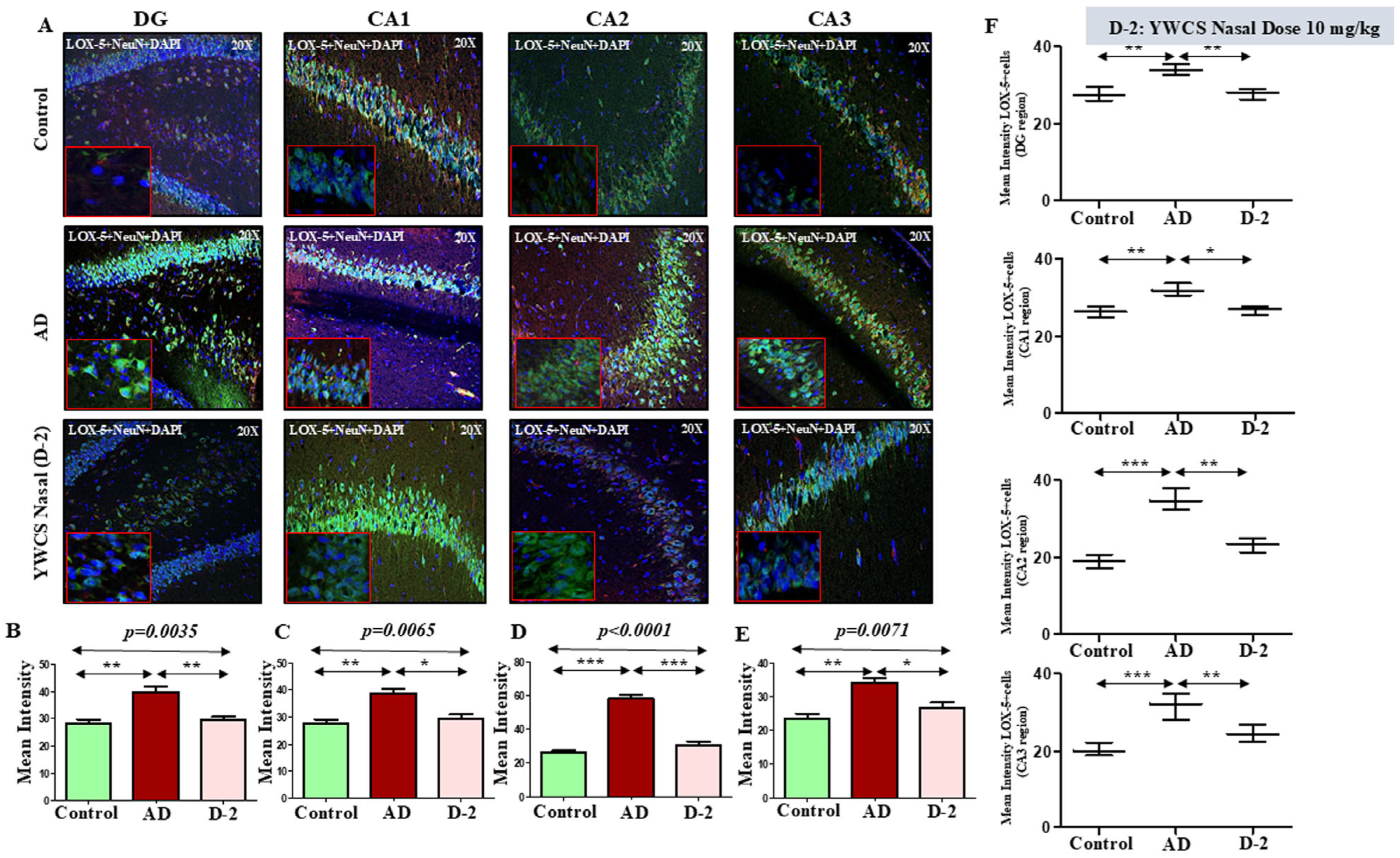
3.6. Immunofluorescence (IF) Assay
3.6.1. Amyloid βeta Oligomers and p-Tau181 Diffusion and Accumulation in Rat Model
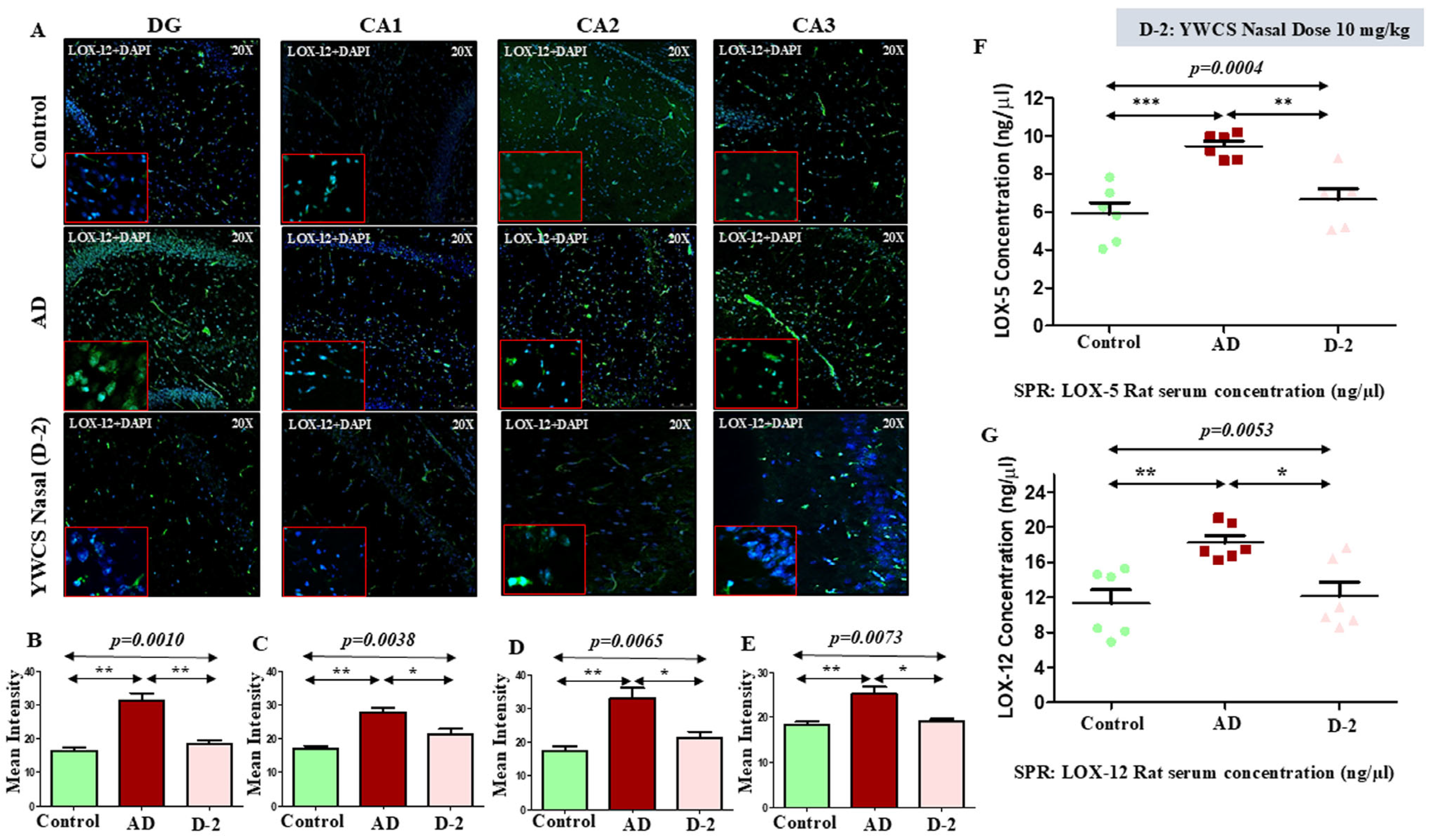
3.6.2. Lipoxygenases (LOX-5 and LOX-12)
LOX-5
LOX-12
3.7. Quantitative Analysis of Rat Serum LOX-5 and LOX-12 by SPR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Czapski, G.A.; Czubowicz, K.; Strosznajder, J.B.; Strosznajder, R.P. The Lipoxygenases: Their Regulation and Implication in Alzheimer’s Disease. Neurochem. Res. 2016, 41, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Hensley, K. Neuroinflammation in Alzheimer’s Disease: Mechanisms, Pathologic Consequences, and Potential for Therapeutic Manipulation. J. Alzheimer’s Dis. 2010, 21, 1–14. [Google Scholar] [CrossRef]
- Mathis, S.P.; Bodduluri, S.R.; Haribabu, B. Interrelationship between the 5-lipoxygenase pathway and microbial dysbiosis in the progression of Alzheimer’s disease. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 158982. [Google Scholar] [CrossRef]
- Yao, Y.; Clark, C.M.; Trojanowski, J.Q.; Lee, V.M.Y.; Praticò, D. Elevation of 12/15 lipoxygenase products in AD and mild cognitive impairment. Ann. Neurol. 2005, 58, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Giannopoulos, P.F.; Chiu, J.; Praticò, D. Antileukotriene therapy by reducing tau phosphorylation improves synaptic integrity and cognition of P301S transgenic mice. Aging Cell 2018, 17, e12759. [Google Scholar] [CrossRef]
- Vagnozzi, A.N.; Giannopoulos, P.F.; Praticò, D. Brain 5-lipoxygenase over-expression worsens memory, synaptic integrity, and tau pathology in the P301S mice. Aging Cell 2018, 17, e12695. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.; Praticò, D. The 5-Lipoxygenase as modulator of Alzheimer’s γ-secretase and therapeutic target. Brain Res. Bull. 2016, 126 Pt 2, 207–212. [Google Scholar] [CrossRef]
- Chu, J.; Praticò, D. 5-Lipoxygenase pharmacological blockade decreases tau phosphorylation in vivo: Involvement of the cyclin-dependent kinase-5. Neurobiol. Aging 2013, 34, 1549–1554. [Google Scholar] [CrossRef]
- Chu, J.; Li, J.G.; Praticò, D. Zileuton improves memory deficits, amyloid and tau pathology in a mouse model of Alzheimer’s disease with plaques and tangles. PLoS ONE 2013, 8, e70991. [Google Scholar] [CrossRef]
- Tu, X.K.; Zhang, H.B.; Shi, S.S.; Liang, R.S.; Wang, C.H.; Chen, C.M.; Yang, W.Z. 5-LOX Inhibitor Zileuton Reduces Inflammatory Reaction and Ischemic Brain Damage Through the Activation of PI3K/Akt Signaling Pathway. Neurochem. Res. 2016, 41, 2779–2787. [Google Scholar] [CrossRef]
- Shekhar, S.; Yadav, S.K.; Rai, N.; Kumar, R.; Yadav, Y.; Tripathi, M.; Dey, A.B.; Dey, S. 5-LOX in Alzheimer’s Disease: Potential Serum Marker and In Vitro Evidences for Rescue of Neurotoxicity by Its Inhibitor YWCS. Mol. Neurobiol. 2018, 55, 2754–2762. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Singh, R.; Naz, F.; Chauhan, S.S.; Dinda, A.; Shukla, A.A.; Gill, K.; Kapoor, V.; Dey, S. Structure Based Design and Synthesis of Peptide Inhibitor of Human LOX-12: In Vitro and In Vivo Analysis of a Novel Therapeutic Agent for Breast Cancer. PLoS ONE 2012, 7, e32521. [Google Scholar] [CrossRef]
- Wei, G.; Shea, J.E. Effects of solvent on the structure of the Alzheimer amyloid-beta (25–35) peptide. Biophys. J. 2006, 91, 1638–1647. [Google Scholar] [CrossRef]
- Sato, K.; Wakamiya, A.; Maeda, T.; Noguchi, K.; Takashima, A.; Imahori, K. Correlation among secondary structure, amyloid precursor protein accumulation, and neurotoxicity of amyloid beta (25–35) peptide as analyzed by single alanine substitution. J. Biochem. 1995, 118, 1108–1111. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, G.; Jayakumar, R. Structural analysis of amyloid beta peptide fragment (25–35) in different microenvironments. Biopolymers 2004, 76, 421–434. [Google Scholar] [CrossRef]
- D’Ursi, A.M.; Armenante, M.R.; Guerrini, R.; Salvadori, S.; Sorrentino, G.; Picone, D. Solution structure of amyloid beta-peptide (25–35) in different media. J. Med. Chem. 2004, 47, 4231–4238. [Google Scholar] [CrossRef]
- Konno, T. Amyloid-induced aggregation and precipitation of soluble proteins: An electrostatic contribution of the Alzheimer’s beta(25–35) amyloid fibril. Biochemistry 2001, 40, 2148–2154. [Google Scholar] [CrossRef] [PubMed]
- Pike, C.J.; Walencewicz-Wasserman, A.J.; Kosmoski, J.; Cribbs, D.H.; Glabe, C.G.; Cotman, C.W. Structure-activity analyses of beta-amyloid peptides: Contributions of the beta 25-35 region to aggregation and neurotoxicity. J. Neurochem. 1995, 64, 253–265. [Google Scholar] [CrossRef]
- Maurice, T.; Mustafa, M.H.; Desrumaux, C.; Keller, E.; Naert, G.; de la CGarcía-Barceló, M.; Rodríguez Cruz, Y.; Garcia Rodríguez, J.C. Intranasal formulation of erythropoietin (EPO) showed potent protective activity against amyloid toxicity in the Aβ25–35 non-transgenic mouse model of Alzheimer’s disease. J. Psychopharmacol. 2013, 27, 1044–1057. [Google Scholar] [CrossRef] [PubMed]
- Stepanichev, M.Y.; Moiseeva, Y.V.; Lazareva, N.A.; Onufriev, M.V.; Gulyaeva, N.V. Single intracerebroventricular administration of amyloid-beta (25–35) peptide induces impairment in short-term rather than long-term memory in rats. Brain Res. Bull. 2003, 61, 197–205. [Google Scholar] [CrossRef]
- Cheng, L.; Yin, W.J.; Zhang, J.F.; Qi, J.S. Amyloid beta-protein fragments 25-35 and 31-35 potentiate long-term depression in hippocampal CA1 region of rats in vivo. Synapse 2009, 63, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Guo, H.; Li, C.; Wang, D.; Wu, J.; Wang, C.; Xu, J.; Qin, R.A. Cognitive improvement of compound danshen in an Aβ25-35 peptide-induced rat model of Alzheimer’s disease. BMC Complement. Altern. Med. 2015, 15, 382. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Chen, W.; Wang, P.; Chu, J. Asiatic acid protects differentiated PC12 cells from Aβ25-35-induced apoptosis and tau hyperphosphorylation via regulating PI3K/Akt/GSK-3β signaling. Life Sci. 2018, 208, 96–101. [Google Scholar] [CrossRef]
- Canet, G.; Zussy, C.; Hernandez, C.; Maurice, T.; Desrumaux, C.; Givalois, L. The pathomimetic oAβ25-35 model of Alzheimer’s disease: Potential for screening of new therapeutic agents. Pharmacol. Ther. 2023, 245, 108398. [Google Scholar] [CrossRef]
- Merrifield, R.B. Solid Phase Peptide Synthesis. I. The Synthesis of a Tetrapeptide. J. Am. Chem. Soc. 1963, 85, 2149–2154. [Google Scholar] [CrossRef]
- Millucci, L.; Ghezzi, L.; Bernardini, G.; Santucci, A. Conformations and biological activities of amyloid beta peptide 25-35. Curr. Protein Pept. Sci. 2010, 11, 54–67. [Google Scholar] [CrossRef]
- Kumar, R.; Singh, A.K.; Kumar, M.; Shekhar, S.; Rai, N.; Kaur, P.; Parshad, R.; Dey, S. Serum 5-LOX: A progressive protein marker for breast cancer and new approach for therapeutic target. Carcinogenesis 2016, 37, 912–917. [Google Scholar] [CrossRef]
- Manev, H.; Chen, H.; Dzitoyeva, S.; Manev, R. Cyclooxygenases and 5-lipoxygenase in Alzheimer’s disease. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 315–319. [Google Scholar] [CrossRef]
- Chu, J.; Praticò, D. The 5-Lipoxygenase as a Common Pathway for Pathological Brain and Vascular Aging. Cardiovasc. Psychiatry Neurol. 2009, 2009, 174657. [Google Scholar] [CrossRef]
- Lammers, C.H.; Schweitzer, P.; Facchinetti, P.; Arrang, J.M.; Madamba, S.G.; Siggins, G.R.; Piomelli, D. Arachidonate 5-Lipoxygenase and Its Activating Protein: Prominent Hippocampal Expression and Role in Somatostatin Signaling. J. Neurochem. 1996, 66, 147–152. [Google Scholar] [CrossRef]
- Chinnici, C.M.; Yao, Y.; Praticò, D. The 5-lipoxygenase enzymatic pathway in the mouse brain: Young versus old. Neurobiol. Aging 2007, 28, 1457–1462. [Google Scholar] [CrossRef]
- Kanaoka, Y.; Boyce, J.A. Cysteinyl Leukotrienes and Their Receptors; Emerging Concepts. Allergy Asthma Immunol. Res. 2014, 6, 288–295. [Google Scholar] [CrossRef]
- Kalra, J.; Kumar, P.; Majeed, A.B.; Prakash, A. Modulation of LOX and COX pathways via inhibition of amyloidogenesis contributes to mitoprotection against β-amyloid oligomer-induced toxicity in an animal model of Alzheimer’s disease in rats. Pharmacol. Biochem. Behav. 2016, 146–147, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Firuzi, O.; Zhuo, J.; Chinnici, C.M.; Wisniewski, T.; Praticò, D. 5-Lipoxygenase gene disruption reduces amyloid-β pathology in a mouse model of Alzheimer’s disease. FASEB J. 2008, 22, 1169–1178. [Google Scholar] [CrossRef]
- Ikonomovic, M.D.; Abrahamson, E.E.; Uz, T.; Manev, H.; DeKosky, S.T. Increased 5-Lipoxygenase Immunoreactivity in the Hippocampus of Patients With Alzheimer’s Disease. J. Histochem. Cytochem. 2008, 56, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Joshi, Y.B.; Giannopoulos, P.F.; Praticò, D. The 12/15Lipoxygenase as an emerging therapeutic target for Alzheimer’s disease. Trends Pharmacol. Sci. 2015, 36, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Joshi, Y.B.; Giannopoulos, P.F.; Chu, J.; Praticò, D. Modulation of LPS-induced memory insult, γ-secretase and neuroinflammation in 3xTg mice by 5-Lipoxygenase. Neurobiol. Aging 2014, 35, 1024–1031. [Google Scholar] [CrossRef]
- Gilbert, N.C.; Bartlett, S.G.; Waight, M.T.; Neau, D.B.; Boeglin, W.E.; Brash, A.R.; Newcomer, M.E. The structure of human 5-lipoxygenase. Science 2011, 331, 217–219. [Google Scholar] [CrossRef]
- Advani, D.; Gupta, R.; Tripathi, R.; Sharma, S.; Ambasta, R.K.; Kumar, P. Protective role of anticancer drugs in neurodegenerative disorders: A drug repurposing approach. Neurochem. Int. 2020, 140, 104841. [Google Scholar] [CrossRef]
- Habchi, J.; Arosio, P.; Perni, M.; Costa, A.R.; Yagi-Utsumi, M.; Joshi, P.; Chia, S.; Cohen, S.I.; Müller, M.B.; Linse, S.; et al. An anticancer drug suppresses the primary nucleation reaction that initiates the production of the toxic Aβ42 aggregates linked with Alzheimer’s disease. Sci. Adv. 2016, 2, e1501244. [Google Scholar] [CrossRef]
- Frozza, R.L.; Horn, A.P.; Hoppe, J.B.; Simão, F.; Gerhardt, D.; Comiran, R.A.; Salbego, C.G. A comparative study of beta-amyloid peptides Abeta1-42 and Abeta25-35 toxicity in organotypic hippocampal slice cultures. Neurochem. Res. 2009, 34, 295–303. [Google Scholar] [CrossRef]
- Filali, I.; Romdhane, A.; Znati, M.; Jannet, H.B.; Bouajila, J. Synthesis of New Harmine Isoxazoles and Evaluation of their Potential Anti-Alzheimer, Anti-inflammatory, and Anticancer Activities. Med. Chem. 2016, 12, 184–190. [Google Scholar] [CrossRef]
- Benedikz, E.; Kloskowska, E.; Winblad, B. The rat as an animal model of Alzheimer’s disease. J. Cell. Mol. Med. 2009, 13, 1034–1042. [Google Scholar] [CrossRef]
- Aquino, R.; de Concini, V.; Dhenain, M.; Lam, S.; Gosset, D.; Baquedano, L.; Forero, M.G.; Menuet, A.; Baril, P.; Pichon, C. Intrahippocampal Inoculation of Aβ1-42 Peptide in Rat as a Model of Alzheimer’s Disease Identified MicroRNA-146a-5p as Blood Marker with Anti-Inflammatory Function in Astrocyte Cells. Cells 2023, 12, 694. [Google Scholar] [CrossRef] [PubMed]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [PubMed]
- Rao, Y.L.; Ganaraja, B.; Murlimanju, B.V.; Joy, T.; Krishnamurthy, A.; Agrawal, A. Hippocampus and its involvement in Alzheimer’s disease: A review. 3 Biotech. 2022, 12, 55. [Google Scholar] [CrossRef] [PubMed]
- Tabatabaei-Jafari, H.; Shaw, M.E.; Cherbuin, N. Cerebral atrophy in mild cognitive impairment: A systematic review with meta-analysis. Alzheimer’s Dement. 2015, 1, 487–504. [Google Scholar] [CrossRef]
- Dudek, S.M.; Alexander, G.M.; Farris, S. Rediscovering area CA2: Unique properties and functions. Nat. Rev. Neurosci. 2016, 17, 89–102. [Google Scholar] [CrossRef]
- Upadhyay, R.K. Drug delivery systems, CNS protection, and the blood brain barrier. Biomed. Res. Int. 2014, 2014, 869269. [Google Scholar] [CrossRef]
- Bors, L.A.; Erdő, F. Overcoming the Blood–Brain Barrier. Challenges and Tricks for CNS Drug Delivery. Sci. Pharm. 2019, 87, 6. [Google Scholar] [CrossRef]
- Bahadur, S.; Pardhi, D.M.; Rautio, J.; Rosenholm, J.M.; Pathak, K. Intranasal Nanoemulsions for Direct Nose-to-Brain Delivery of Actives for CNS Disorders. Pharmaceutics 2020, 12, 1230. [Google Scholar] [CrossRef] [PubMed]
- Gänger, S.; Schindowski, K. Tailoring Formulations for Intranasal Nose-to-Brain Delivery: A Review on Architecture, Physico-Chemical Characteristics and Mucociliary Clearance of the Nasal Olfactory Mucosa. Pharmaceutics 2018, 10, 116. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yadav, Y.; Anwar, M.; Sharma, H.; Jain, S.; Sharma, U.; Haldar, P.; Dey, A.B.; Dey, S. Anti-Inflammatory Peptide Prevents Aβ25–35-Induced Inflammation in Rats via Lipoxygenase Inhibition. Cells 2025, 14, 957. https://doi.org/10.3390/cells14130957
Yadav Y, Anwar M, Sharma H, Jain S, Sharma U, Haldar P, Dey AB, Dey S. Anti-Inflammatory Peptide Prevents Aβ25–35-Induced Inflammation in Rats via Lipoxygenase Inhibition. Cells. 2025; 14(13):957. https://doi.org/10.3390/cells14130957
Chicago/Turabian StyleYadav, Yudhishthir, Masroor Anwar, Hanuman Sharma, Suman Jain, Uma Sharma, Partha Haldar, Aparajit B. Dey, and Sharmistha Dey. 2025. "Anti-Inflammatory Peptide Prevents Aβ25–35-Induced Inflammation in Rats via Lipoxygenase Inhibition" Cells 14, no. 13: 957. https://doi.org/10.3390/cells14130957
APA StyleYadav, Y., Anwar, M., Sharma, H., Jain, S., Sharma, U., Haldar, P., Dey, A. B., & Dey, S. (2025). Anti-Inflammatory Peptide Prevents Aβ25–35-Induced Inflammation in Rats via Lipoxygenase Inhibition. Cells, 14(13), 957. https://doi.org/10.3390/cells14130957