
Tyrosine 67 Phosphorylation Controls Respiration and Limits the Apoptotic Functions of Cytochrome c
 , , ,
, , ,  and
and 
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Methods
2.1. Isolation of Cytc from Bovine Heart Tissue
2.2. Mass Spectrometry of Purified Cytc to Detect Site-Specific Phosphorylation
2.3. Mutagenesis, Expression, and Purification of Recombinant Cytc
2.4. Concentration Determination of Cytc
2.5. Measurement of Cytochrome c Oxidase Activity
2.6. Caspase-3 Activity
2.7. Measurement of Cytc Redox Potential
2.8. Measurement of Cytc Rate of Oxidation
2.9. Measurement of Cytc Rate of Reduction by Superoxide Scavenging
2.10. Heme Degradation Assay
2.11. Cardiolipin Peroxidase Activity
2.12. Molecular Dynamics Simulations
2.13. Establishment of Stable Cell Lines Expressing Cytc Variants
2.14. Western Blotting for Cytc
2.15. Measurement of Oxygen Consumption Rate in Intact Cells
2.16. Measurement of Mitochondrial Membrane Potential
2.17. Measurement of Mitochondrial ROS
2.18. Cell Death Using Annexin V/Propidium Iodide Staining
2.19. Statistical Analyses
3. Results
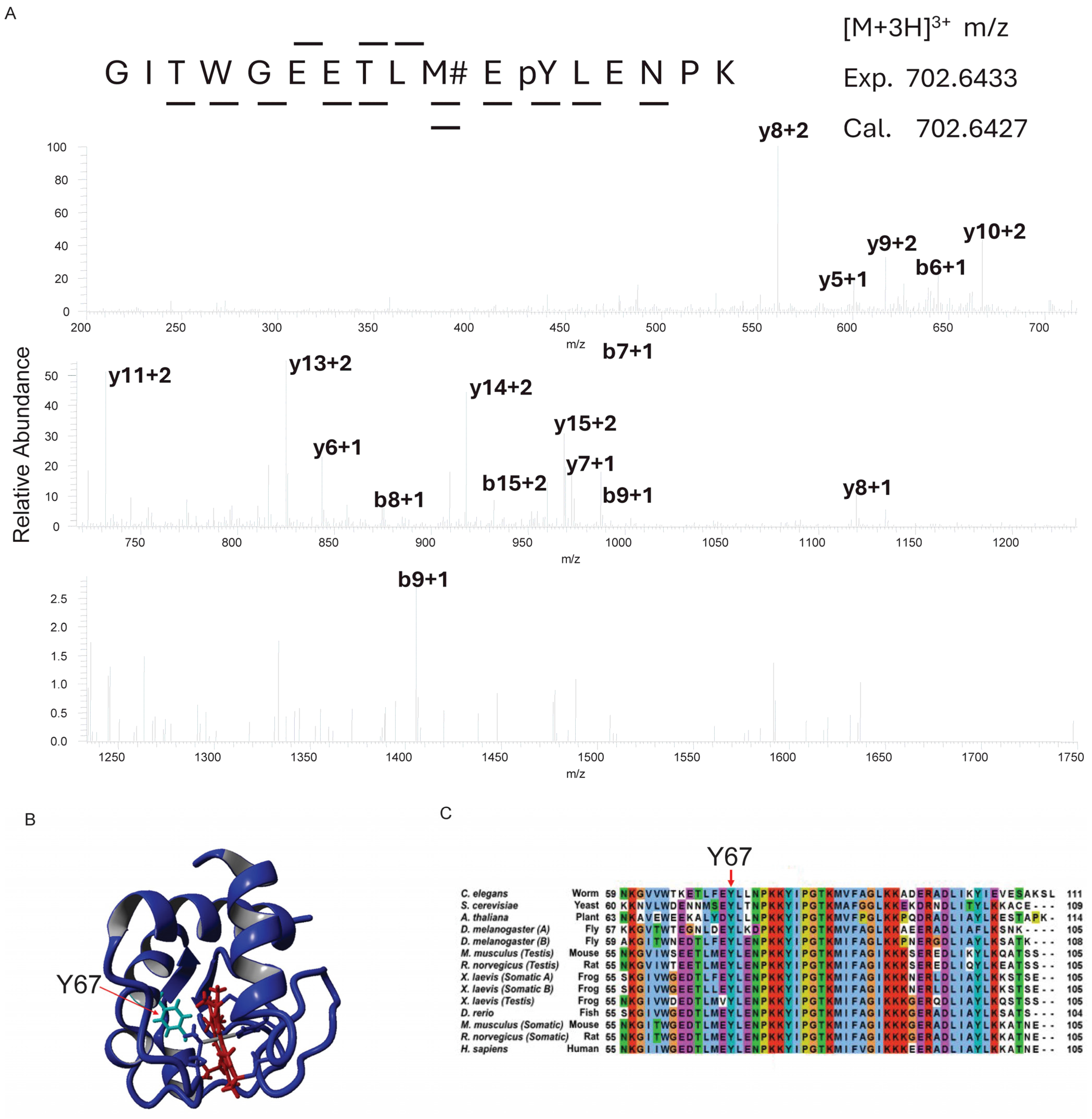
3.1. Cytc Y67 Is Conserved in Mammals and Phosphorylated in Bovine Heart
3.2. Overexpression and Purification of Functional Cytc Variants in E. coli Cells
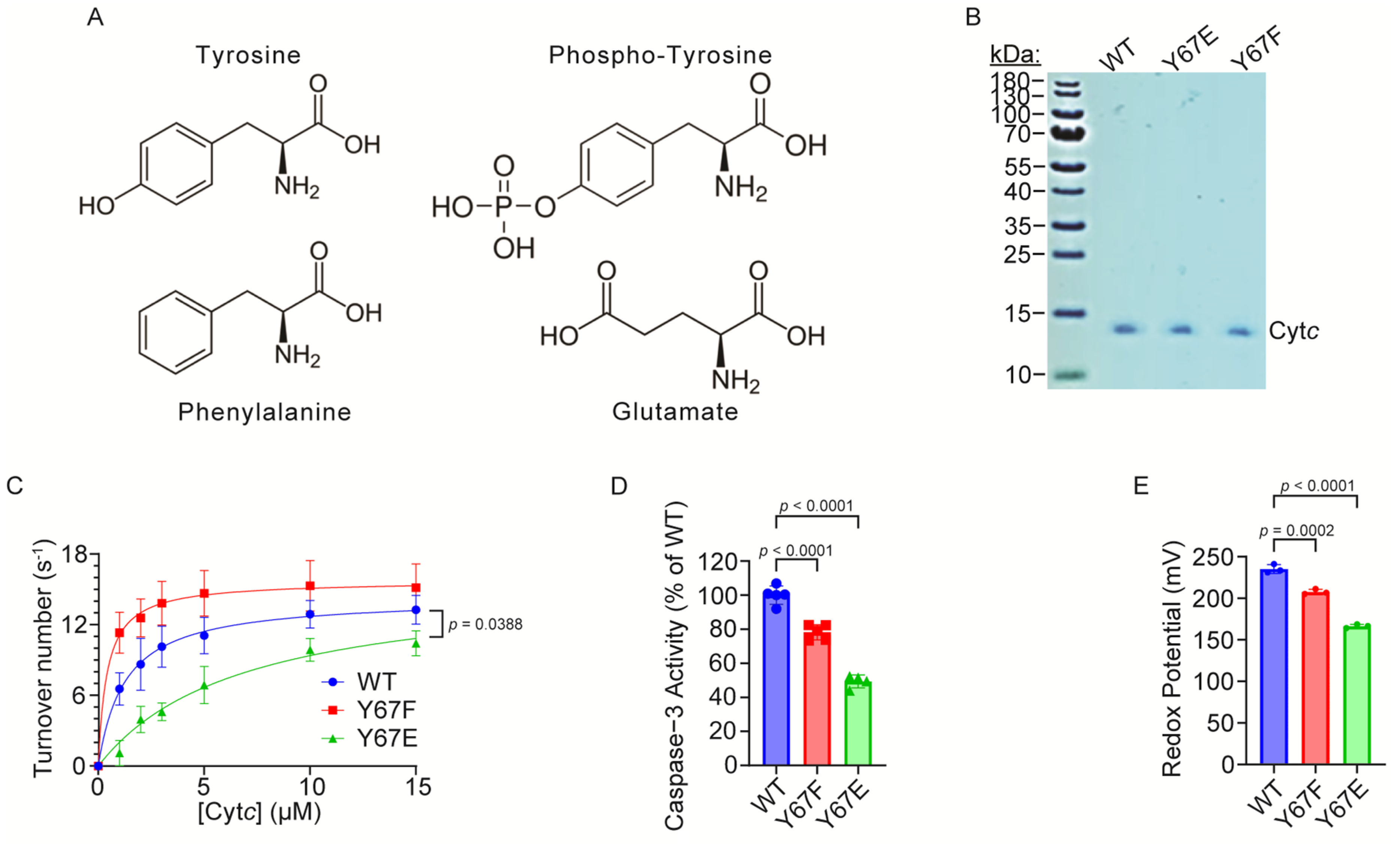
3.3. Cytochrome c Oxidase Activity Is Inhibited in the Reaction with Phosphomimetic Y67E Cytc
3.4. Caspase-3 Activity Is Inhibited by Phosphomimetic Y67E Cytc
3.5. Redox Potential, Susceptibility to Oxidation, and Superoxide Scavenging Activity Are Decreased in Phosphomimetic Cytc
3.6. Phosphomimetic Y67E Cytc Showed Increased Susceptibility to Degradation Under High Levels of Oxidative Stress
3.7. Phosphomimetic Y67E Cytc Impairs Cardiolipin Peroxidase Activity
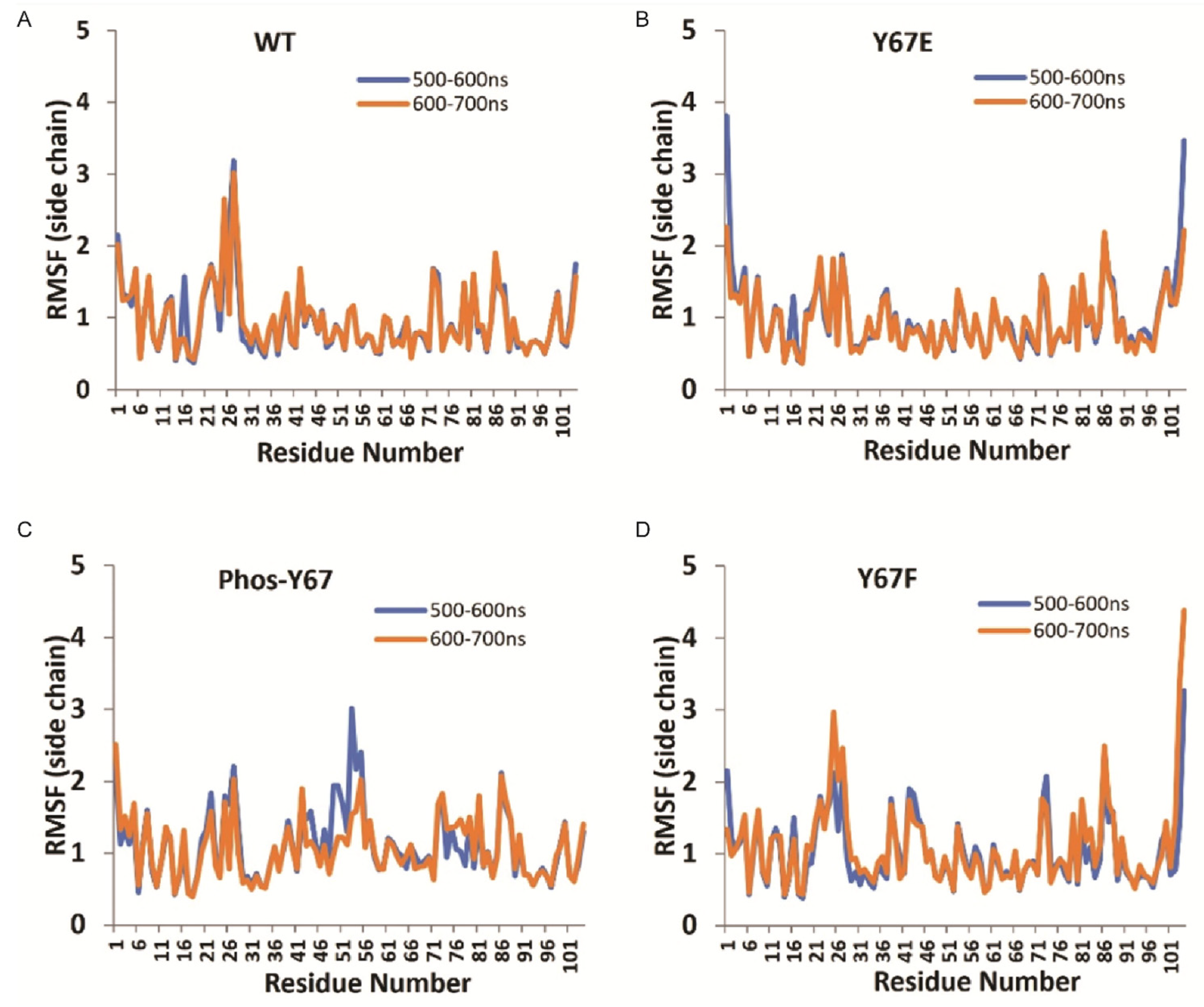
3.8. Molecular Dynamics Modeling of Cytc
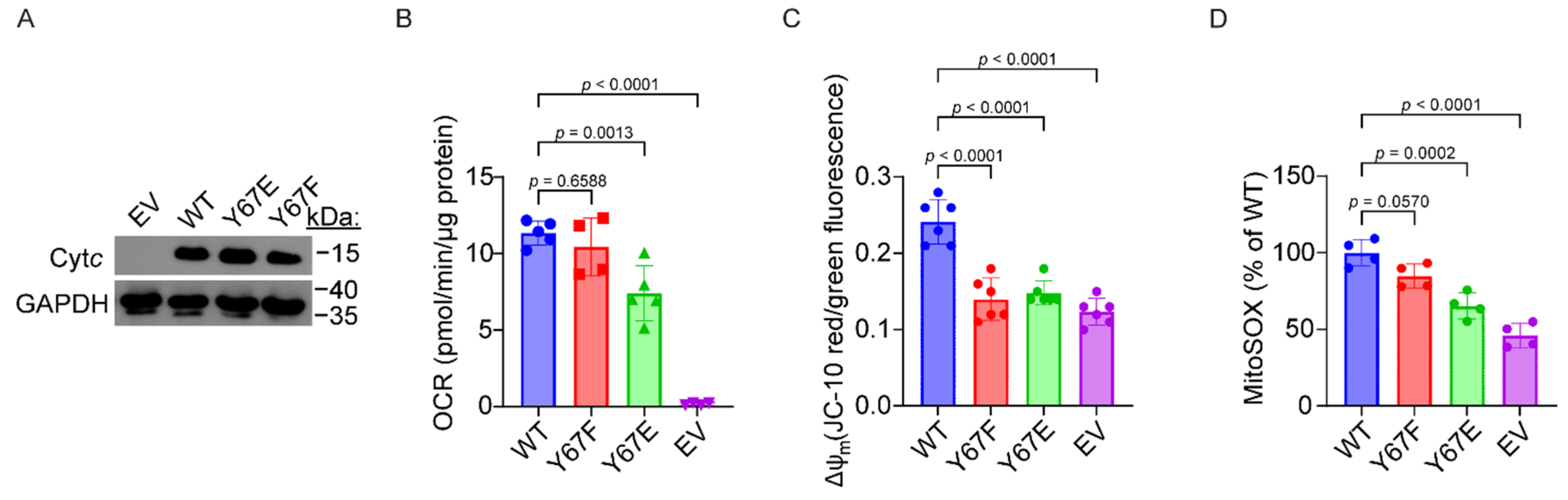
3.9. Cells Expressing Phosphomimetic Y67E Cytc Have Decreased Mitochondrial Respiration
3.10. Cells Expressing Phosphomimetic Y67E Cytc Have Decreased Mitochondrial Membrane Potential and ROS Production
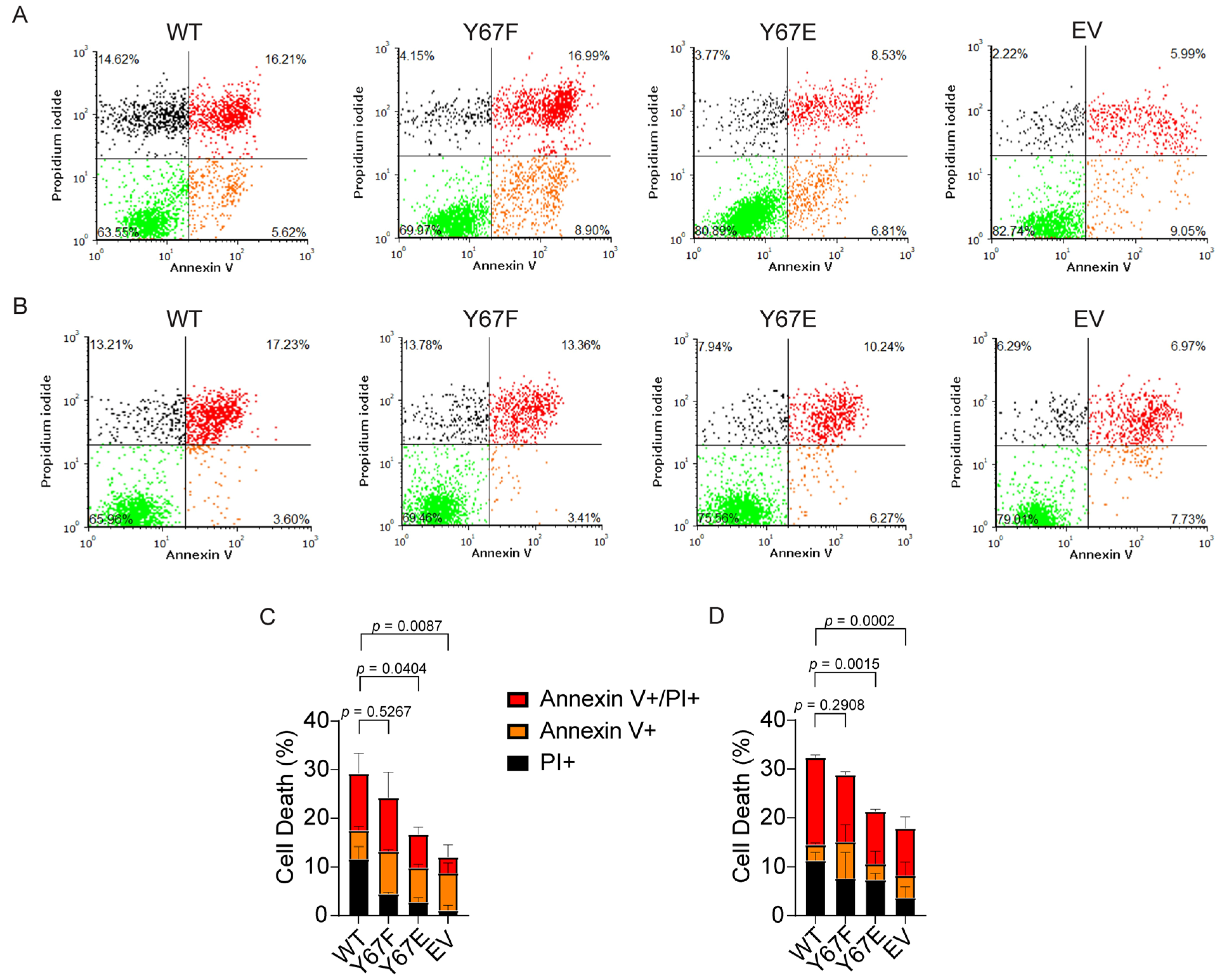
3.11. Cells Expressing Phosphomimetic Y67E Cytc Show Decreased Apoptosis After Cellular Injury
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Apaf-1 | protein apoptotic protease activating factor 1 |
| CL | cardiolipin |
| COX | cytochrome c oxidase (complex IV) |
| Cytc | cytochrome c |
| DCIP | 2,6-dichloroindophenol |
| ETC | electron transport chain |
| EV | empty vector (control cells that do not contain Cytc) |
| PI | propidium iodide |
| OCR | oxygen consumption rate |
| RMSF | root mean square fluctuations |
| ROS | reactive oxygen species |
| SOD | superoxide dismutase |
| WT | wild type |
| Y67E | replacement of tyrosine 67 (of Cytc) with glutamate |
| Y67F | replacement of tyrosine 67 (of Cytc) with phenylalanine |
| ∆Ѱm | mitochondrial membrane potential |
References
- Hüttemann, M.; Pecina, P.; Rainbolt, M.; Sanderson, T.H.; Kagan, V.E.; Samavati, L.; Doan, J.W.; Lee, I. The multiple functions of cytochrome c and their regulation in life and death decisions of the mammalian cell: From respiration to apoptosis. Mitochondrion 2011, 11, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Salemme, F.R. Structure and function of cytochromes c. Annu. Rev. Biochem. 1977, 46, 299–329. [Google Scholar] [CrossRef] [PubMed]
- Piccoli, C.; Scrima, R.; Boffoli, D.; Capitanio, N. Control by cytochrome c oxidase of the cellular oxidative phosphorylation system depends on the mitochondrial energy state. Biochem. J. 2006, 396, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Dalmonte, M.E.; Forte, E.; Genova, M.L.; Giuffre, A.; Sarti, P.; Lenaz, G. Control of respiration by cytochrome c oxidase in intact cells: Role of the membrane potential. J. Biol. Chem. 2009, 284, 32331–32335. [Google Scholar] [CrossRef]
- Kalpage, H.A.; Bazylianska, V.; Recanati, M.A.; Fite, A.; Liu, J.; Wan, J.; Mantena, N.; Malek, M.H.; Podgorski, I.; Heath, E.I.; et al. Tissue-specific regulation of cytochrome c by post-translational modifications: Respiration, the mitochondrial membrane potential, ROS, and apoptosis. FASEB J. 2019, 33, 1540–1553. [Google Scholar] [CrossRef]
- Nur, E.K.A.; Gross, S.R.; Pan, Z.; Balklava, Z.; Ma, J.; Liu, L.F. Nuclear translocation of cytochrome c during apoptosis. J. Biol. Chem. 2004, 279, 24911–24914. [Google Scholar] [CrossRef]
- González-Arzola, K.; Díaz-Moreno, I.; Cano-González, A.; Díaz-Quintana, A.; Velázquez-Campoy, A.; Moreno-Beltrán, B.; López-Rivas, A.; De la Rosa, M.A. Structural basis for inhibition of the histone chaperone activity of SET/TAF-Iβ by cytochrome c. Proc. Natl. Acad. Sci. USA 2015, 112, 9908–9913. [Google Scholar] [CrossRef]
- Korshunov, S.S.; Krasnikov, B.F.; Pereverzev, M.O.; Skulachev, V.P. The antioxidant functions of cytochrome c. FEBS Lett. 1999, 462, 192–198. [Google Scholar] [CrossRef]
- Pereverzev, M.O.; Vygodina, T.V.; Konstantinov, A.A.; Skulachev, V.P. Cytochrome c, an ideal antioxidant. Biochem. Soc. Trans. 2003, 31, 1312–1315. [Google Scholar] [CrossRef]
- Barros, M.H.; Netto, L.E.; Kowaltowski, A.J. H2O2 generation in Saccharomyces cerevisiae respiratory pet mutants: Effect of cytochrome c. Free Radic. Biol. Med. 2003, 35, 179–188. [Google Scholar] [CrossRef]
- Schlame, M.; Rua, D.; Greenberg, M.L. The biosynthesis and functional role of cardiolipin. Prog. Lipid Res. 2000, 39, 257–288. [Google Scholar] [CrossRef]
- Kagan, V.E.; Borisenko, G.G.; Tyurina, Y.Y.; Tyurin, V.A.; Jiang, J.; Potapovich, A.I.; Kini, V.; Amoscato, A.A.; Fujii, Y. Oxidative lipidomics of apoptosis: Redox catalytic interactions of cytochrome c with cardiolipin and phosphatidylserine. Free Radic. Biol. Med. 2004, 37, 1963–1985. [Google Scholar] [CrossRef]
- Bergstrom, C.L.; Beales, P.A.; Lv, Y.; Vanderlick, T.K.; Groves, J.T. Cytochrome c causes pore formation in cardiolipin-containing membranes. Proc. Natl. Acad. Sci. USA 2013, 110, 6269–6274. [Google Scholar] [CrossRef] [PubMed]
- Tuominen, E.K.; Wallace, C.J.; Kinnunen, P.K. Phospholipid-cytochrome c interaction: Evidence for the extended lipid anchorage. J. Biol. Chem. 2002, 277, 8822–8826. [Google Scholar] [CrossRef]
- Mandal, A.; Hoop, C.L.; DeLucia, M.; Kodali, R.; Kagan, V.E.; Ahn, J.; van der Wel, P.C. Structural Changes and Proapoptotic Peroxidase Activity of Cardiolipin-Bound Mitochondrial Cytochrome c. Biophys. J. 2015, 109, 1873–1884. [Google Scholar] [CrossRef] [PubMed]
- Kagan, V.E.; Tyurina, Y.Y.; Bayir, H.; Chu, C.T.; Kapralov, A.A.; Vlasova, I.I.; Belikova, N.A.; Tyurin, V.A.; Amoscato, A.; Epperly, M.; et al. The “pro-apoptotic genies” get out of mitochondria: Oxidative lipidomics and redox activity of cytochrome c/cardiolipin complexes. Chem. -Biol. Interact. 2006, 163, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Barayeu, U.; Lange, M.; Mendez, L.; Arnhold, J.; Shadyro, O.I.; Fedorova, M.; Flemmig, J. Cytochrome c autocatalyzed carbonylation in the presence of hydrogen peroxide and cardiolipins. J. Biol. Chem. 2019, 294, 1816–1830. [Google Scholar] [CrossRef]
- Gonzalez-Arzola, K.; Diaz-Quintana, A.; Bernardo-Garcia, N.; Martinez-Fabregas, J.; Rivero-Rodriguez, F.; Casado-Combreras, M.A.; Elena-Real, C.A.; Velazquez-Cruz, A.; Gil-Caballero, S.; Velazquez-Campoy, A.; et al. Nucleus-translocated mitochondrial cytochrome c liberates nucleophosmin-sequestered ARF tumor suppressor by changing nucleolar liquid-liquid phase separation. Nat. Struct. Mol. Biol. 2022, 29, 1024–1036. [Google Scholar] [CrossRef]
- Buzon, P.; Velazquez-Cruz, A.; Corrales-Guerrero, L.; Diaz-Quintana, A.; Diaz-Moreno, I.; Roos, W.H. The Histone Chaperones SET/TAF-1beta and NPM1 Exhibit Conserved Functionality in Nucleosome Remodeling and Histone Eviction in a Cytochrome c-Dependent Manner. Adv. Sci. 2023, 10, e2301859. [Google Scholar] [CrossRef]
- Casado-Combreras, M.A.; Velazquez-Campoy, A.; Martinho, M.; Belle, V.; De la Rosa, M.A.; Diaz-Moreno, I. Cytochrome c prompts the recruitment of its nuclear partners SET/TAF-Ibeta and NPM1 into biomolecular condensates. iScience 2024, 27, 110435. [Google Scholar] [CrossRef]
- Tamargo-Azpilicueta, J.; Casado-Combreras, M.A.; Giner-Arroyo, R.L.; Velazquez-Campoy, A.; Marquez, I.; Olloqui-Sariego, J.L.; De la Rosa, M.A.; Diaz-Moreno, I. Phosphorylation of cytochrome c at tyrosine 48 finely regulates its binding to the histone chaperone SET/TAF-Ibeta in the nucleus. Protein Sci. 2024, 33, e5213. [Google Scholar] [CrossRef] [PubMed]
- Guerra-Castellano, A.; Marquez, I.; Perez-Mejias, G.; Diaz-Quintana, A.; De la Rosa, M.A.; Diaz-Moreno, I. Post-Translational Modifications of Cytochrome c in Cell Life and Disease. Int. J. Mol. Sci. 2020, 21, 8483. [Google Scholar] [CrossRef]
- Morse, P.T.; Arroum, T.; Wan, J.; Pham, L.; Vaishnav, A.; Bell, J.; Pavelich, L.; Malek, M.H.; Sanderson, T.H.; Edwards, B.F.P.; et al. Phosphorylations and Acetylations of Cytochrome c Control Mitochondrial Respiration, Mitochondrial Membrane Potential, Energy, ROS, and Apoptosis. Cells 2024, 13, 493. [Google Scholar] [CrossRef]
- Yin, V.; Mian, S.H.; Konermann, L. Lysine carbonylation is a previously unrecognized contributor to peroxidase activation of cytochrome c by chloramine-T. Chem. Sci. 2019, 10, 2349–2359. [Google Scholar] [CrossRef] [PubMed]
- Yin, V.; Shaw, G.S.; Konermann, L. Cytochrome c as a Peroxidase: Activation of the Precatalytic Native State by H2O2-Induced Covalent Modifications. J. Am. Chem. Soc. 2017, 139, 15701–15709. [Google Scholar] [CrossRef]
- Flatmark, T. On the heterogeneity of beef heart cytochrome c. 3. A kinetic study of the non-enzymic deamidation of the main subfractions (Cy I-Cy 3). Acta Chem. Scand. 1966, 20, 1487–1496. [Google Scholar] [CrossRef]
- Sharma, G.S.; Warepam, M.; Bhattacharya, R.; Singh, L.R. Covalent Modification by Glyoxals Converts Cytochrome c Into its Apoptotically Competent State. Sci. Rep. 2019, 9, 4781. [Google Scholar] [CrossRef]
- Hildick-Smith, G.J.; Downey, M.C.; Gretebeck, L.M.; Gersten, R.A.; Sandwick, R.K. Ribose 5-phosphate glycation reduces cytochrome c respiratory activity and membrane affinity. Biochemistry 2011, 50, 11047–11057. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.M.; Gomes, R.A.; Yang, D.; Dennison, S.R.; Familia, C.; Lages, A.; Coelho, A.V.; Murphy, R.M.; Phoenix, D.A.; Quintas, A. Insights into the molecular mechanism of protein native-like aggregation upon glycation. Biochim. Biophys. Acta 2013, 1834, 1010–1022. [Google Scholar] [CrossRef]
- Mercado-Uribe, H.; Andrade-Medina, M.; Espinoza-Rodriguez, J.H.; Carrillo-Tripp, M.; Scheckhuber, C.Q. Analyzing structural alterations of mitochondrial intermembrane space superoxide scavengers cytochrome -c and SOD1 after methylglyoxal treatment. PLoS ONE 2020, 15, e0232408. [Google Scholar] [CrossRef]
- Delgado, Y.; Morales-Cruz, M.; Hernandez-Roman, J.; Martinez, Y.; Griebenow, K. Chemical glycosylation of cytochrome c improves physical and chemical protein stability. BMC Biochem. 2014, 15, 16. [Google Scholar] [CrossRef]
- Mendez, J.; Morales Cruz, M.; Delgado, Y.; Figueroa, C.M.; Orellano, E.A.; Morales, M.; Monteagudo, A.; Griebenow, K. Delivery of chemically glycosylated cytochrome c immobilized in mesoporous silica nanoparticles induces apoptosis in HeLa cancer cells. Mol. Pharm. 2014, 11, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.S.; Singh, L.R. Conformational status of cytochrome c upon N-homocysteinylation: Implications to cytochrome c release. Arch. Biochem. Biophys. 2017, 614, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Dopner, S.; Hildebrandt, P.; Rosell, F.I.; Mauk, A.G.; von Walter, M.; Buse, G.; Soulimane, T. The structural and functional role of lysine residues in the binding domain of cytochrome c in the electron transfer to cytochrome c oxidase. Eur. J. Biochem. 1999, 261, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Perla-Kajan, J.; Marczak, L.; Kajan, L.; Skowronek, P.; Twardowski, T.; Jakubowski, H. Modification by homocysteine thiolactone affects redox status of cytochrome C. Biochemistry 2007, 46, 6225–6231. [Google Scholar] [CrossRef]
- Gates, A.T.; Moore, L., Jr.; Sylvain, M.R.; Jones, C.M.; Lowry, M.; El-Zahab, B.; Robinson, J.W.; Strongin, R.M.; Warner, I.M. Mechanistic investigation of N-homocysteinylation-mediated protein-gold nanoconjugate assembly. Langmuir 2009, 25, 9346–9351. [Google Scholar] [CrossRef]
- Garcia-Heredia, J.M.; Diaz-Moreno, I.; Nieto, P.M.; Orzaez, M.; Kocanis, S.; Teixeira, M.; Perez-Paya, E.; Diaz-Quintana, A.; De la Rosa, M.A. Nitration of tyrosine 74 prevents human cytochrome c to play a key role in apoptosis signaling by blocking caspase-9 activation. Biochim. Biophys. Acta 2010, 1797, 981–993. [Google Scholar] [CrossRef]
- Garcia-Heredia, J.M.; Diaz-Moreno, I.; Diaz-Quintana, A.; Orzaez, M.; Navarro, J.A.; Hervas, M.; De la Rosa, M.A. Specific nitration of tyrosines 46 and 48 makes cytochrome c assemble a non-functional apoptosome. FEBS Lett. 2012, 586, 154–158. [Google Scholar] [CrossRef]
- Diaz-Moreno, I.; Nieto, P.M.; Del Conte, R.; Gairi, M.; Garcia-Heredia, J.M.; De la Rosa, M.A.; Diaz-Quintana, A. A non-damaging method to analyze the configuration and dynamics of nitrotyrosines in proteins. Chemistry 2012, 18, 3872–3878. [Google Scholar] [CrossRef]
- Nakagawa, D.; Ohshima, Y.; Takusagawa, M.; Ikota, N.; Takahashi, Y.; Shimizu, S.; Ozawa, T. Functional modification of cytochrome c by peroxynitrite in an electron transfer reaction. Chem. Pharm. Bull. 2001, 49, 1547–1554. [Google Scholar] [CrossRef]
- Diaz-Moreno, I.; Garcia-Heredia, J.M.; Diaz-Quintana, A.; Teixeira, M.; De la Rosa, M.A. Nitration of tyrosines 46 and 48 induces the specific degradation of cytochrome c upon change of the heme iron state to high-spin. Biochim. Biophys. Acta 2011, 1807, 1616–1623. [Google Scholar] [CrossRef]
- Vlasova, I.I.; Tyurin, V.A.; Kapralov, A.A.; Kurnikov, I.V.; Osipov, A.N.; Potapovich, M.V.; Stoyanovsky, D.A.; Kagan, V.E. Nitric oxide inhibits peroxidase activity of cytochrome c. cardiolipin complex and blocks cardiolipin oxidation. J. Biol. Chem. 2006, 281, 14554–14562. [Google Scholar] [CrossRef]
- Chen, Y.R.; Deterding, L.J.; Sturgeon, B.E.; Tomer, K.B.; Mason, R.P. Protein oxidation of cytochrome C by reactive halogen species enhances its peroxidase activity. J. Biol. Chem. 2002, 277, 29781–29791. [Google Scholar] [CrossRef]
- Ivanetich, K.M.; Bradshaw, J.J.; Kaminsky, L.S. Methionine sulfoxide cytochrome c. Biochemistry 1976, 15, 1144–1153. [Google Scholar] [CrossRef]
- Capdevila, D.A.; Oviedo Rouco, S.; Tomasina, F.; Tortora, V.; Demicheli, V.; Radi, R.; Murgida, D.H. Active Site Structure and Peroxidase Activity of Oxidatively Modified Cytochrome c Species in Complexes with Cardiolipin. Biochemistry 2015, 54, 7491–7504. [Google Scholar] [CrossRef] [PubMed]
- Capdevila, D.A.; Marmisolle, W.A.; Tomasina, F.; Demicheli, V.; Portela, M.; Radi, R.; Murgida, D.H. Specific methionine oxidation of cytochrome c in complexes with zwitterionic lipids by hydrogen peroxide: Potential implications for apoptosis. Chem. Sci. 2015, 6, 705–713. [Google Scholar] [CrossRef]
- Mahapatra, G.; Varughese, A.; Ji, Q.; Lee, I.; Liu, J.; Vaishnav, A.; Sinkler, C.; Kapralov, A.A.; Moraes, C.T.; Sanderson, T.H.; et al. Phosphorylation of Cytochrome c Threonine 28 Regulates Electron Transport Chain Activity in Kidney: IMPLICATIONS FOR AMP KINASE. J. Biol. Chem. 2017, 292, 64–79. [Google Scholar] [CrossRef] [PubMed]
- Guerra-Castellano, A.; Diaz-Moreno, I.; Velazquez-Campoy, A.; De la Rosa, M.A.; Diaz-Quintana, A. Structural and functional characterization of phosphomimetic mutants of cytochrome c at threonine 28 and serine 47. Biochim. Biophys. Acta 2016, 1857, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Kalpage, H.A.; Vaishnav, A.; Liu, J.; Lee, I.; Mahapatra, G.; Turner, A.A.; Zurek, M.P.; Ji, Q.; Moraes, C.T.; et al. Regulation of Respiration and Apoptosis by Cytochrome c Threonine 58 Phosphorylation. Sci. Rep. 2019, 9, 15815. [Google Scholar] [CrossRef]
- Kalpage, H.A.; Vaishnav, A.; Liu, J.; Varughese, A.; Wan, J.; Turner, A.A.; Ji, Q.; Zurek, M.P.; Kapralov, A.A.; Kagan, V.E.; et al. Serine-47 phosphorylation of cytochrome c in the mammalian brain regulates cytochrome c oxidase and caspase-3 activity. FASEB J. 2019, 33, 13503–13514. [Google Scholar] [CrossRef]
- Kalpage, H.A.; Wan, J.; Morse, P.T.; Lee, I.; Hüttemann, M. Brain-Specific Serine-47 Modification of Cytochrome c Regulates Cytochrome c Oxidase Activity Attenuating ROS Production and Cell Death: Implications for Ischemia/Reperfusion Injury and Akt Signaling. Cells 2020, 9, 1843. [Google Scholar] [CrossRef]
- Yu, H.; Lee, I.; Salomon, A.R.; Yu, K.; Hüttemann, M. Mammalian liver cytochrome c is tyrosine-48 phosphorylated in vivo, inhibiting mitochondrial respiration. Biochim. Biophys. Acta 2008, 1777, 1066–1071. [Google Scholar] [CrossRef]
- Pecina, P.; Borisenko, G.G.; Belikova, N.A.; Tyurina, Y.Y.; Pecinova, A.; Lee, I.; Samhan-Arias, A.K.; Przyklenk, K.; Kagan, V.E.; Hüttemann, M. Phosphomimetic substitution of cytochrome c tyrosine 48 decreases respiration and binding to cardiolipin and abolishes ability to trigger downstream caspase activation. Biochemistry 2010, 49, 6705–6714. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Heredia, J.M.; Diaz-Quintana, A.; Salzano, M.; Orzaez, M.; Perez-Paya, E.; Teixeira, M.; De la Rosa, M.A.; Diaz-Moreno, I. Tyrosine phosphorylation turns alkaline transition into a biologically relevant process and makes human cytochrome c behave as an anti-apoptotic switch. J. Biol. Inorg. Chem. 2011, 16, 1155–1168. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Beltran, B.; Guerra-Castellano, A.; Diaz-Quintana, A.; Del Conte, R.; Garcia-Maurino, S.M.; Diaz-Moreno, S.; Gonzalez-Arzola, K.; Santos-Ocana, C.; Velazquez-Campoy, A.; De la Rosa, M.A.; et al. Structural basis of mitochondrial dysfunction in response to cytochrome c phosphorylation at tyrosine 48. Proc. Natl. Acad. Sci. USA 2017, 114, E3041–E3050. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Sun, H.; Lin, X.; Li, Q.; Zhao, D.; Cheng, Z.; Liu, J.; Fan, Q. Increased cytochrome c threonine 50 phosphorylation in aging heart as a novel defensive signaling against hypoxia/reoxygenation induced apoptosis. Aging 2022, 14, 5699–5709. [Google Scholar] [CrossRef]
- Lee, I.; Salomon, A.R.; Yu, K.; Doan, J.W.; Grossman, L.I.; Hüttemann, M. New prospects for an old enzyme: Mammalian cytochrome c is tyrosine-phosphorylated in vivo. Biochemistry 2006, 45, 9121–9128. [Google Scholar] [CrossRef]
- Morse, P.T.; Pasupathi, V.; Vuljaj, S.; Yazdi, N.; Zurek, M.P.; Wan, J.; Lee, I.; Vaishnav, A.; Edwards, B.F.P.; Arroum, T.; et al. Cardiac Tyrosine 97 Phosphorylation of Cytochrome c Regulates Respiration and Apoptosis. Int. J. Mol. Sci. 2025, 26, 1314. [Google Scholar] [CrossRef]
- Guerra-Castellano, A.; Diaz-Quintana, A.; Perez-Mejias, G.; Elena-Real, C.A.; Gonzalez-Arzola, K.; Garcia-Maurino, S.M.; De la Rosa, M.A.; Diaz-Moreno, I. Oxidative stress is tightly regulated by cytochrome c phosphorylation and respirasome factors in mitochondria. Proc. Natl. Acad. Sci. USA 2018, 115, 7955–7960. [Google Scholar] [CrossRef]
- Morse, P.T.; Perez-Mejias, G.; Wan, J.; Turner, A.A.; Marquez, I.; Kalpage, H.A.; Vaishnav, A.; Zurek, M.P.; Huettemann, P.P.; Kim, K.; et al. Cytochrome c lysine acetylation regulates cellular respiration and cell death in ischemic skeletal muscle. Nat. Commun. 2023, 14, 4166. [Google Scholar] [CrossRef]
- Bazylianska, V.; Kalpage, H.A.; Wan, J.; Vaishnav, A.; Mahapatra, G.; Turner, A.A.; Chowdhury, D.D.; Kim, K.; Morse, P.T.; Lee, I.; et al. Lysine 53 Acetylation of Cytochrome c in Prostate Cancer: Warburg Metabolism and Evasion of Apoptosis. Cells 2021, 10, 802. [Google Scholar] [CrossRef] [PubMed]
- Marquez, I.; Perez-Mejias, G.; Guerra-Castellano, A.; Olloqui-Sariego, J.L.; Andreu, R.; Calvente, J.J.; De la Rosa, M.A.; Diaz-Moreno, I. Structural and functional insights into lysine acetylation of cytochrome c using mimetic point mutants. FEBS Open Bio 2021, 11, 3304–3323. [Google Scholar] [CrossRef] [PubMed]
- Morse, P.T.; Wan, J.; Arroum, T.; Herroon, M.K.; Kalpage, H.A.; Bazylianska, V.; Lee, I.; Heath, E.I.; Podgorski, I.; Hüttemann, M. Prostate Cancer-Specific Lysine 53 Acetylation of Cytochrome c Drives Metabolic Reprogramming and Protects from Apoptosis in Intact Cells. Biomolecules 2024, 14, 695. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Tiwari, S.; Singh, S. Pirh2 modulates the mitochondrial function and cytochrome c-mediated neuronal death during Alzheimer’s disease. Cell Death Dis. 2024, 15, 331. [Google Scholar] [CrossRef]
- Hüttemann, M.; Lee, I.; Samavati, L.; Yu, H.; Doan, J.W. Regulation of mitochondrial oxidative phosphorylation through cell signaling. Biochim. Biophys. Acta 2007, 1773, 1701–1720. [Google Scholar] [CrossRef]
- Hüttemann, M.; Lee, I.; Pecinova, A.; Pecina, P.; Przyklenk, K.; Doan, J.W. Regulation of oxidative phosphorylation, the mitochondrial membrane potential, and their role in human disease. J. Bioenerg. Biomembr. 2008, 40, 445–456. [Google Scholar] [CrossRef]
- Sanderson, T.H.; Mahapatra, G.; Pecina, P.; Ji, Q.; Yu, K.; Sinkler, C.; Varughese, A.; Kumar, R.; Bukowski, M.J.; Tousignant, R.N.; et al. Cytochrome c is tyrosine 97 phosphorylated by neuroprotective insulin treatment. PLoS ONE 2013, 8, e78627. [Google Scholar] [CrossRef]
- Deep, S.; Im, S.C.; Zuiderweg, E.R.; Waskell, L. Characterization and calculation of a cytochrome c-cytochrome b5 complex using NMR data. Biochemistry 2005, 44, 10654–10668. [Google Scholar] [CrossRef]
- Lee, I.; Salomon, A.R.; Yu, K.; Samavati, L.; Pecina, P.; Pecinova, A.; Hüttemann, M. Isolation of regulatory-competent, phosphorylated cytochrome c oxidase. Methods Enzymol. 2009, 457, 193–210. [Google Scholar] [CrossRef]
- Vempati, U.D.; Diaz, F.; Barrientos, A.; Narisawa, S.; Mian, A.M.; Millan, J.L.; Boise, L.H.; Moraes, C.T. Role of cytochrome c in apoptosis: Increased sensitivity to tumor necrosis factor alpha is associated with respiratory defects but not with lack of cytochrome c release. Mol. Cell Biol. 2007, 27, 1771–1783. [Google Scholar] [CrossRef]
- Cammack, R. Redox States and Potentials in Bioenergetics—A Practical Approach; Brown, G.C., Cooper, C.E., Eds.; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Liu, Z.; Lin, H.; Ye, S.; Liu, Q.Y.; Meng, Z.; Zhang, C.M.; Xia, Y.; Margoliash, E.; Rao, Z.; Liu, X.J. Remarkably high activities of testicular cytochrome c in destroying reactive oxygen species and in triggering apoptosis. Proc. Natl. Acad. Sci. USA 2006, 103, 8965–8970. [Google Scholar] [CrossRef]
- Fridovich, I. Quantitative aspects of the production of superoxide anion radical by milk xanthine oxidase. J. Biol. Chem. 1970, 245, 4053–4057. [Google Scholar] [CrossRef] [PubMed]
- Quick, K.L.; Hardt, J.I.; Dugan, L.L. Rapid microplate assay for superoxide scavenging efficiency. J. Neurosci. Methods 2000, 97, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Krieger, E.; Vriend, G. New ways to boost molecular dynamics simulations. J. Comput. Chem. 2015, 36, 996–1007. [Google Scholar] [CrossRef]
- Hornak, V.; Abel, R.; Okur, A.; Strockbine, B.; Roitberg, A.; Simmerling, C. Comparison of multiple Amber force fields and development of improved protein backbone parameters. Proteins 2006, 65, 712–725. [Google Scholar] [CrossRef]
- Zaidi, S.; Hassan, M.I.; Islam, A.; Ahmad, F. The role of key residues in structure, function, and stability of cytochrome -c. Cell Mol. Life Sci. 2014, 71, 229–255. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Cho, K.; Park, H.S. Chemical biology approaches for studying posttranslational modifications. RNA Biol. 2018, 15, 427–440. [Google Scholar] [CrossRef]
- Kalpage, H.A.; Wan, J.; Morse, P.T.; Zurek, M.P.; Turner, A.A.; Khobeir, A.; Yazdi, N.; Hakim, L.; Liu, J.; Vaishnav, A.; et al. Cytochrome c phosphorylation: Control of mitochondrial electron transport chain flux and apoptosis. Int. J. Biochem. Cell Biol. 2020, 121, 105704. [Google Scholar] [CrossRef]
- Nicholls, D.G.; Ferguson, S.J. Bioenergetics 2; Academic Press Limited: London, UK; San Diego, CA, USA, 1992. [Google Scholar]
- Kapralov, A.A.; Yanamala, N.; Tyurina, Y.Y.; Castro, L.; Samhan-Arias, A.; Vladimirov, Y.A.; Maeda, A.; Weitz, A.A.; Peterson, J.; Mylnikov, D.; et al. Topography of tyrosine residues and their involvement in peroxidation of polyunsaturated cardiolipin in cytochrome c/cardiolipin peroxidase complexes. Biochim. Biophys. Acta 2011, 1808, 2147–2155. [Google Scholar] [CrossRef]
- Battistuzzi, G.; Bortolotti, C.A.; Bellei, M.; Di Rocco, G.; Salewski, J.; Hildebrandt, P.; Sola, M. Role of Met80 and Tyr67 in the low-pH conformational equilibria of cytochrome c. Biochemistry 2012, 51, 5967–5978. [Google Scholar] [CrossRef]
- Hannibal, L.; Tomasina, F.; Capdevila, D.A.; Demicheli, V.; Tortora, V.; Alvarez-Paggi, D.; Jemmerson, R.; Murgida, D.H.; Radi, R. Alternative Conformations of Cytochrome c: Structure, Function, and Detection. Biochemistry 2016, 55, 407–428. [Google Scholar] [CrossRef] [PubMed]
- Ying, T.; Wang, Z.H.; Lin, Y.W.; Xie, J.; Tan, X.; Huang, Z.X. Tyrosine-67 in cytochrome c is a possible apoptotic trigger controlled by hydrogen bonds via a conformational transition. Chem. Commun. 2009, 4512–4514. [Google Scholar] [CrossRef]
- Lan, W.; Wang, Z.; Yang, Z.; Ying, T.; Zhang, X.; Tan, X.; Liu, M.; Cao, C.; Huang, Z.X. Structural basis for cytochrome c Y67H mutant to function as a peroxidase. PLoS ONE 2014, 9, e107305. [Google Scholar] [CrossRef]
- Berghuis, A.M.; Guillemette, J.G.; Smith, M.; Brayer, G.D. Mutation of tyrosine-67 to phenylalanine in cytochrome c significantly alters the local heme environment. J. Mol. Biol. 1994, 235, 1326–1341. [Google Scholar] [CrossRef]
- Tognaccini, L.; Ciaccio, C.; D’Oria, V.; Cervelli, M.; Howes, B.D.; Coletta, M.; Mariottini, P.; Smulevich, G.; Fiorucci, L. Structure-function relationships in human cytochrome c: The role of tyrosine 67. J. Inorg. Biochem. 2016, 155, 56–66. [Google Scholar] [CrossRef]
- Luntz, T.L.; Schejter, A.; Garber, E.A.; Margoliash, E. Structural significance of an internal water molecule studied by site-directed mutagenesis of tyrosine-67 in rat cytochrome c. Proc. Natl. Acad. Sci. USA 1989, 86, 3524–3528. [Google Scholar] [CrossRef] [PubMed]
- Cassina, A.M.; Hodara, R.; Souza, J.M.; Thomson, L.; Castro, L.; Ischiropoulos, H.; Freeman, B.A.; Radi, R. Cytochrome c nitration by peroxynitrite. J. Biol. Chem. 2000, 275, 21409–21415. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Roldán, V.; García-Heredia, J.M.; Navarro, J.A.; Rosa, M.A.D.l.; Hervás, M. Effect of Nitration on the Physicochemical and Kinetic Features of Wild-Type and Monotyrosine Mutants of Human Respiratory Cytochrome c. Biochemistry 2008, 47, 12371–12379. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.F.; Wang, Y.T.; Yen, H.Y.; Tsou, C.C.; Ku, W.C.; Lin, P.Y.; Chen, H.Y.; Nesvizhskii, A.I.; Ishihama, Y.; Chen, Y.J. Large-scale determination of absolute phosphorylation stoichiometries in human cells by motif-targeting quantitative proteomics. Nat. Commun. 2015, 6, 6622. [Google Scholar] [CrossRef]
- Zhou, M.; Li, Y.; Hu, Q.; Bai, X.C.; Huang, W.; Yan, C.; Scheres, S.H.; Shi, Y. Atomic structure of the apoptosome: Mechanism of cytochrome c- and dATP-mediated activation of Apaf-1. Genes Dev. 2015, 29, 2349–2361. [Google Scholar] [CrossRef]
- Cheng, T.C.; Hong, C.; Akey, I.V.; Yuan, S.; Akey, C.W. A near atomic structure of the active human apoptosome. eLife 2016, 5, e17755. [Google Scholar] [CrossRef] [PubMed]
- Tyurina, Y.Y.; Kini, V.; Tyurin, V.A.; Vlasova, I.I.; Jiang, J.; Kapralov, A.A.; Belikova, N.A.; Yalowich, J.C.; Kurnikov, I.V.; Kagan, V.E. Mechanisms of cardiolipin oxidation by cytochrome c: Relevance to pro- and antiapoptotic functions of etoposide. Mol. Pharmacol. 2006, 70, 706–717. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Sun, W.; Tyurin, V.A.; DeLucia, M.; Ahn, J.; Kagan, V.E.; van der Wel, P.C.A. Activation of Cytochrome c Peroxidase Function Through Coordinated Foldon Loop Dynamics upon Interaction with Anionic Lipids. J. Mol. Biol. 2021, 433, 167057. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, J.; Morse, P.T.; Zurek, M.P.; Turner, A.A.; Vaishnav, A.; Salomon, A.R.; Edwards, B.F.P.; Arroum, T.; Hüttemann, M. Tyrosine 67 Phosphorylation Controls Respiration and Limits the Apoptotic Functions of Cytochrome c. Cells 2025, 14, 951. https://doi.org/10.3390/cells14130951
Wan J, Morse PT, Zurek MP, Turner AA, Vaishnav A, Salomon AR, Edwards BFP, Arroum T, Hüttemann M. Tyrosine 67 Phosphorylation Controls Respiration and Limits the Apoptotic Functions of Cytochrome c. Cells. 2025; 14(13):951. https://doi.org/10.3390/cells14130951
Chicago/Turabian StyleWan, Junmei, Paul T. Morse, Matthew P. Zurek, Alice A. Turner, Asmita Vaishnav, Arthur R. Salomon, Brian F. P. Edwards, Tasnim Arroum, and Maik Hüttemann. 2025. "Tyrosine 67 Phosphorylation Controls Respiration and Limits the Apoptotic Functions of Cytochrome c" Cells 14, no. 13: 951. https://doi.org/10.3390/cells14130951
APA StyleWan, J., Morse, P. T., Zurek, M. P., Turner, A. A., Vaishnav, A., Salomon, A. R., Edwards, B. F. P., Arroum, T., & Hüttemann, M. (2025). Tyrosine 67 Phosphorylation Controls Respiration and Limits the Apoptotic Functions of Cytochrome c. Cells, 14(13), 951. https://doi.org/10.3390/cells14130951