
Hepatocellular Metabolic Profile: Understanding Post-Thawing Metabolic Shift in Primary Hepatocytes In Vitro

 , ,
, ,  , , , ,
, , , ,
Abstract

1. Introduction
2. Materials and Methods
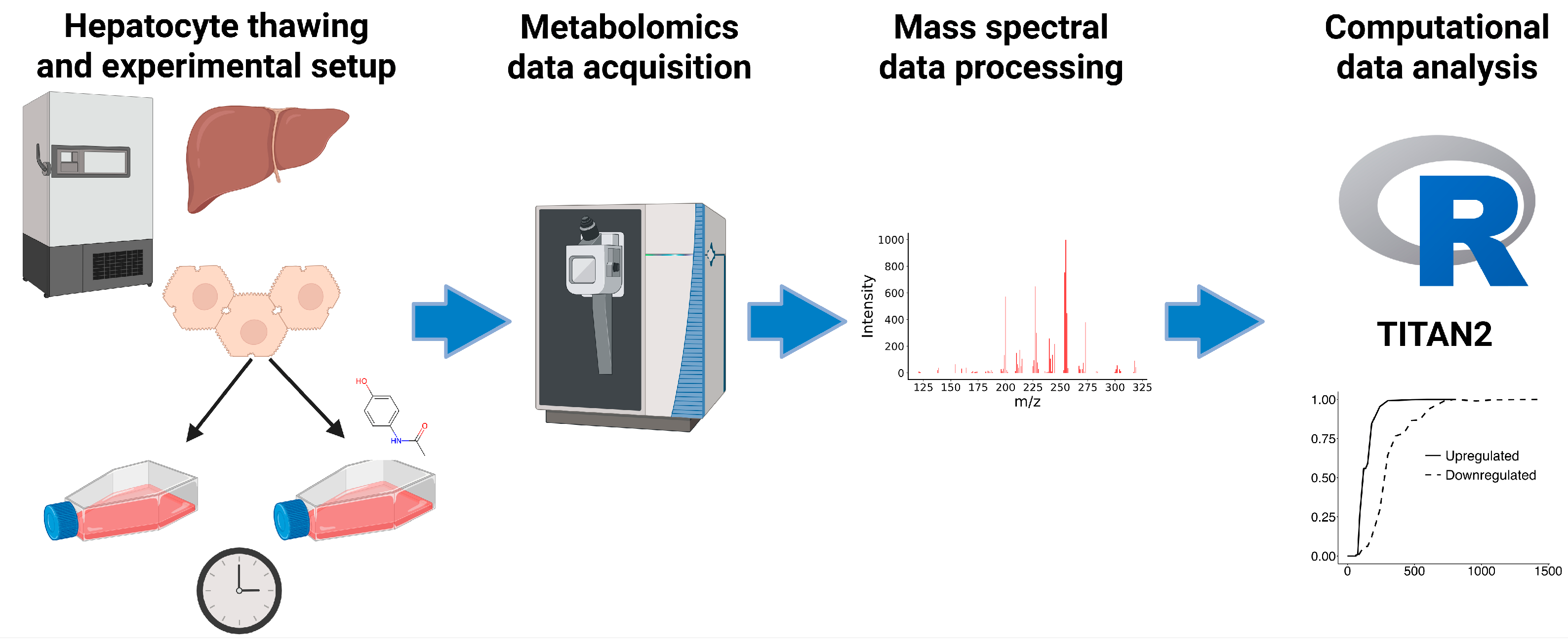
2.1. Culture of Human Hepatocytes
2.2. Preparation for LC-MS/MS Analysis
2.3. LC-MS/MS
2.4. Data Processing
2.5. Statistical Analysis
3. Results
3.1. Evidence of Hepatocyte Metabolic Restoration
3.2. Characterization of Metabolic Shift
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rhyu, J.; Yu, R. Newly discovered endocrine functions of the liver. World J. Hepatol. 2021, 13, 1611–1628. [Google Scholar] [CrossRef] [PubMed]
- Lubbers, R.; van Essen, M.F.; van Kooten, C.; Trouw, L.A. Production of complement components by cells of the immune system. Clin. Exp. Immunol. 2017, 188, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Tu, W.; Liu, J.; Tian, D. Hepatocytes: A key role in liver inflammation. Front. Immunol. 2022, 13, 1083780. [Google Scholar] [CrossRef]
- Houle, R.; Raoul, J.; Lévesque, J.F.; Pang, K.S.; Nicoll-Griffith, D.A.; Silva, J.M. Retention of transporter activities in cryopreserved, isolated rat hepatocytes. Drug Metab. Dispos. 2003, 31, 447–451. [Google Scholar] [CrossRef]
- Hu, C.; Li, L. In vitro culture of isolated primary hepatocytes and stem cell-derived hepatocyte-like cells for liver regeneration. Protein Cell 2015, 6, 562–574. [Google Scholar] [CrossRef]
- Jouin, D.; Blanchard, N.; Alexandre, E.; Delobel, F.; David-Pierson, P.; Lavé, T.; Jaeck, D.; Richert, L.; Coassolo, P. Cryopreserved human hepatocytes in suspension are a convenient high throughput tool for the prediction of metabolic clearance. Eur. J. Pharm. Biopharm. 2006, 63, 347–355. [Google Scholar] [CrossRef]
- Blanchard, N.; Richert, L.; Notter, B.; Delobel, F.; David, P.; Coassolo, P.; Lavé, T. Impact of serum on clearance predictions obtained from suspensions and primary cultures of rat hepatocytes. Eur. J. Pharm. Sci. 2004, 23, 189–199. [Google Scholar] [CrossRef]
- Klieber, S.; Torreilles, F.; Guillou, F.; Fabre, G. The Use of Human Hepatocytes to Investigate Drug Metabolism and CYP Enzyme Induction. In Hepatocytes: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2010; pp. 295–308. [Google Scholar]
- Vernetti, L.A.; Vogt, A.; Gough, A.; Taylor, D.L. Evolution of Experimental Models of the Liver to Predict Human Drug Hepatotoxicity and Efficacy. Clin. Liver Dis. 2017, 21, 197–214. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.D.; Yuen, G.; Tu, T.; Budzinska, M.A.; Spring, K.; Bryant, K.; Shackel, N.A. In Vitro Models of the Liver: Disease Modeling, Drug Discovery and Clinical Applications. In Hepatocellular Carcinoma [Internet]; Tirnitz-Parker, J.E.E., Ed.; Codon Publications: Brisbane, Australia, 2019. [Google Scholar]
- Kaur, I.; Vasudevan, A.; Rawal, P.; Tripathi, D.M.; Ramakrishna, S.; Kaur, S.; Sarin, S.K. Primary Hepatocyte Isolation and Cultures: Technical Aspects, Challenges and Advancements. Bioengineering 2023, 10, 131. [Google Scholar] [CrossRef]
- Hinson, J.A.; Roberts, D.W.; James, L.P. Mechanisms of acetaminophen-induced liver necrosis. Handb. Exp. Pharmacol. 2010, 196, 369–405. [Google Scholar]
- McGill, M.R.; Jaeschke, H. Metabolism and disposition of acetaminophen: Recent advances in relation to hepatotoxicity and diagnosis. Pharm. Res. 2013, 30, 2174–2187. [Google Scholar] [CrossRef] [PubMed]
- Rivera, E.S.; LeBrun, E.S.; Breidenbach, J.D.; Solomon, E.; Sanders, C.K.; Harvey, T.; Tseng, C.Y.; Thornhill, M.G.; Blackwell, B.R.; McBride, E.M.; et al. Feature-agnostic metabolomics for determining effective subcytotoxic doses of common pesticides in human cells. Toxicol. Sci. 2024, 202, 85–95. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. 2025. Available online: https://www.R-project.org/ (accessed on 1 March 2025).
- Shapiro, S.S.; Wilk, M.B. An Analysis of Variance Test for Normality (Complete Samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Kruskal, W.H.; Wallis, W.A. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Baker, M.E.; King, R.S.; Kahle, D. TITAN2: Threshold Indicator Taxa Analysis. 2023. Available online: https://CRAN.R-project.org/package=TITAN2 (accessed on 1 March 2025).
- Baker, M.E.; King, R.S. A new method for detecting and interpreting biodiversity and ecological community thresholds. Glob. Change Biol. 2019, 25, 25–37. [Google Scholar] [CrossRef]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted Metabolomics Strategies—Challenges and Emerging Directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef]
- Fisher, R.A. Statistical Methods for Research Workers. In Breakthroughs in Statistics: Methodology and Distribution; Kotz, S., Johnson, N.L., Eds.; Springer: New York, NY, USA, 1992; pp. 66–70. [Google Scholar]
- Yan, F.; Robert, M.; Li, Y. Statistical methods and common problems in medical or biomedical science research. Int. J. Physiol. Pathophysiol. Pharmacol. 2017, 9, 157–163. [Google Scholar]
- Prince, E.K.; Pohnert, G. Searching for signals in the noise: Metabolomics in chemical ecology. Anal. Bioanal. Chem. 2010, 396, 193–197. [Google Scholar] [CrossRef]
- Yuan, C.; Gao, J.; Guo, J.; Bai, L.; Marshall, C.; Cai, Z.; Wang, L.; Xiao, M. Dimethyl sulfoxide damages mitochondrial integrity and membrane potential in cultured astrocytes. PLoS ONE 2014, 9, e107447. [Google Scholar] [CrossRef]
- Hughes, R.D.; Mitry, R.R.; Lehec, S.C. Cryopreservation of Human Hepatocytes for Clinical Use. In Hepatocytes: Methods and Protocols; Maurel, P., Ed.; Humana Press: Totowa, NJ, USA, 2010; pp. 107–113. [Google Scholar]
- Święciło, A.; Januś, E.; Krzepiłko, A.; Skowrońska, M. The effect of DMSO on Saccharomyces cerevisiae yeast with different energy metabolism and antioxidant status. Sci. Rep. 2024, 14, 21974. [Google Scholar] [CrossRef]
- Len, J.S.; Koh, W.S.D.; Tan, S.X. The roles of reactive oxygen species and antioxidants in cryopreservation. Biosci. Rep. 2019, 39, BSR20191601. [Google Scholar] [CrossRef] [PubMed]
- Terry, C.; Mitry, R.R.; Lehec, S.C.; Muiesan, P.; Rela, M.; Heaton, N.D.; Hughes, R.D.; Dhawan, A. The Effects of Cryopreservation on Human Hepatocytes Obtained From Different Sources of Liver Tissue. Cell Transplant. 2005, 14, 585–594. [Google Scholar] [CrossRef]
- Baust, J.G.; Gao, D.; Baust, J.M. Cryopreservation: An emerging paradigm change. Organogenesis 2009, 5, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Pérez, L.C.; Padilla-Martínez, I.I.; Cruz, A.; Mendieta-Wejebe, J.E.; Tamay-Cach, F.; Rosales-Hernández, M.C. Evaluation of a new benzothiazole derivative with antioxidant activity in the initial phase of acetaminophen toxicity. Arab. J. Chem. 2019, 12, 3871–3882. [Google Scholar] [CrossRef]
- Satapati, S.; Sunny, N.E.; Kucejova, B.; Fu, X.; He, T.T.; Méndez-Lucas, A.; Shelton, J.M.; Perales, J.C.; Browning, J.D.; Burgess, S.C. Elevated TCA cycle function in the pathology of diet-induced hepatic insulin resistance and fatty liver. J. Lipid Res. 2012, 53, 1080–1092. [Google Scholar] [CrossRef]
- Lo, E.K.K.; Felicianna; Xu, J.H.; Zhan, Q.; Zeng, Z.; El-Nezami, H. The Emerging Role of Branched-Chain Amino Acids in Liver Diseases. Biomedicines 2022, 10, 1444. [Google Scholar] [CrossRef]
- Li, Z.; Wang, F.; Liang, B.; Su, Y.; Sun, S.; Xia, S.; Shao, J.; Zhang, Z.; Hong, M.; Zhang, F.; et al. Methionine metabolism in chronic liver diseases: An update on molecular mechanism and therapeutic implication. Signal Transduct. Target. Ther. 2020, 5, 280. [Google Scholar] [CrossRef] [PubMed]
- Shahin, R.; Sakai, Y.; Shirai, K.; Hashita, T.; Matsunaga, T.; Iwao, T. Converting suspension-type human primary hepatocytes to be plateable-type for use as an in vitro pharmacokinetics model. Drug Metab. Pharmacokinet. 2024, 62, 101044. [Google Scholar] [CrossRef]
- Schlegel, A.; Mergental, H.; Fondevila, C.; Porte, R.J.; Friend, P.J.; Dutkowski, P. Machine perfusion of the liver and bioengineering. J. Hepatol. 2023, 78, 1181–1198. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | # Input Features | # Filtered Features | # Upregulated | # Downregulated |
|---|---|---|---|---|
| Unexposed | 1675 | 504 | 398 | 106 |
| Exposed | 1679 | 227 | 196 | 31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palmisano, S.; Breidenbach, J.D.; Blackwell, B.R.; Harvey, T.; Luchini, K.A.; Thornhill, M.G.; LeBrun, E.S.; Mach, P.; Glaros, T.; Rivera, E.S. Hepatocellular Metabolic Profile: Understanding Post-Thawing Metabolic Shift in Primary Hepatocytes In Vitro. Cells 2025, 14, 803. https://doi.org/10.3390/cells14110803
Palmisano S, Breidenbach JD, Blackwell BR, Harvey T, Luchini KA, Thornhill MG, LeBrun ES, Mach P, Glaros T, Rivera ES. Hepatocellular Metabolic Profile: Understanding Post-Thawing Metabolic Shift in Primary Hepatocytes In Vitro. Cells. 2025; 14(11):803. https://doi.org/10.3390/cells14110803
Chicago/Turabian StylePalmisano, Salvator, Joshua D. Breidenbach, Brett R. Blackwell, Tara Harvey, Kes A. Luchini, M. Grace Thornhill, Erick S. LeBrun, Phillip Mach, Trevor Glaros, and Emilio S. Rivera. 2025. "Hepatocellular Metabolic Profile: Understanding Post-Thawing Metabolic Shift in Primary Hepatocytes In Vitro" Cells 14, no. 11: 803. https://doi.org/10.3390/cells14110803
APA StylePalmisano, S., Breidenbach, J. D., Blackwell, B. R., Harvey, T., Luchini, K. A., Thornhill, M. G., LeBrun, E. S., Mach, P., Glaros, T., & Rivera, E. S. (2025). Hepatocellular Metabolic Profile: Understanding Post-Thawing Metabolic Shift in Primary Hepatocytes In Vitro. Cells, 14(11), 803. https://doi.org/10.3390/cells14110803