
Oral Antiviral Defense: Saliva- and Beverage-like Hypotonicity Dynamically Regulate Formation of Membraneless Biomolecular Condensates of Antiviral Human MxA in Oral Epithelial Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Cell Culture
2.2. Plasmids and Transient Transfection
2.3. Live-Cell Fluorescence Imaging
2.4. Phase Transition Experiments
2.5. Immunofluorescence Imaging
2.6. Quantitation of Relative Amounts of GFP-MxA in Condensates vs. Dispersed State in a Cell
2.7. VSV Stock and Virus Infection
2.8. Antibody Reagents and Chemicals
2.9. Osmolarity Measurements
2.10. Statistical Testing
3. Results
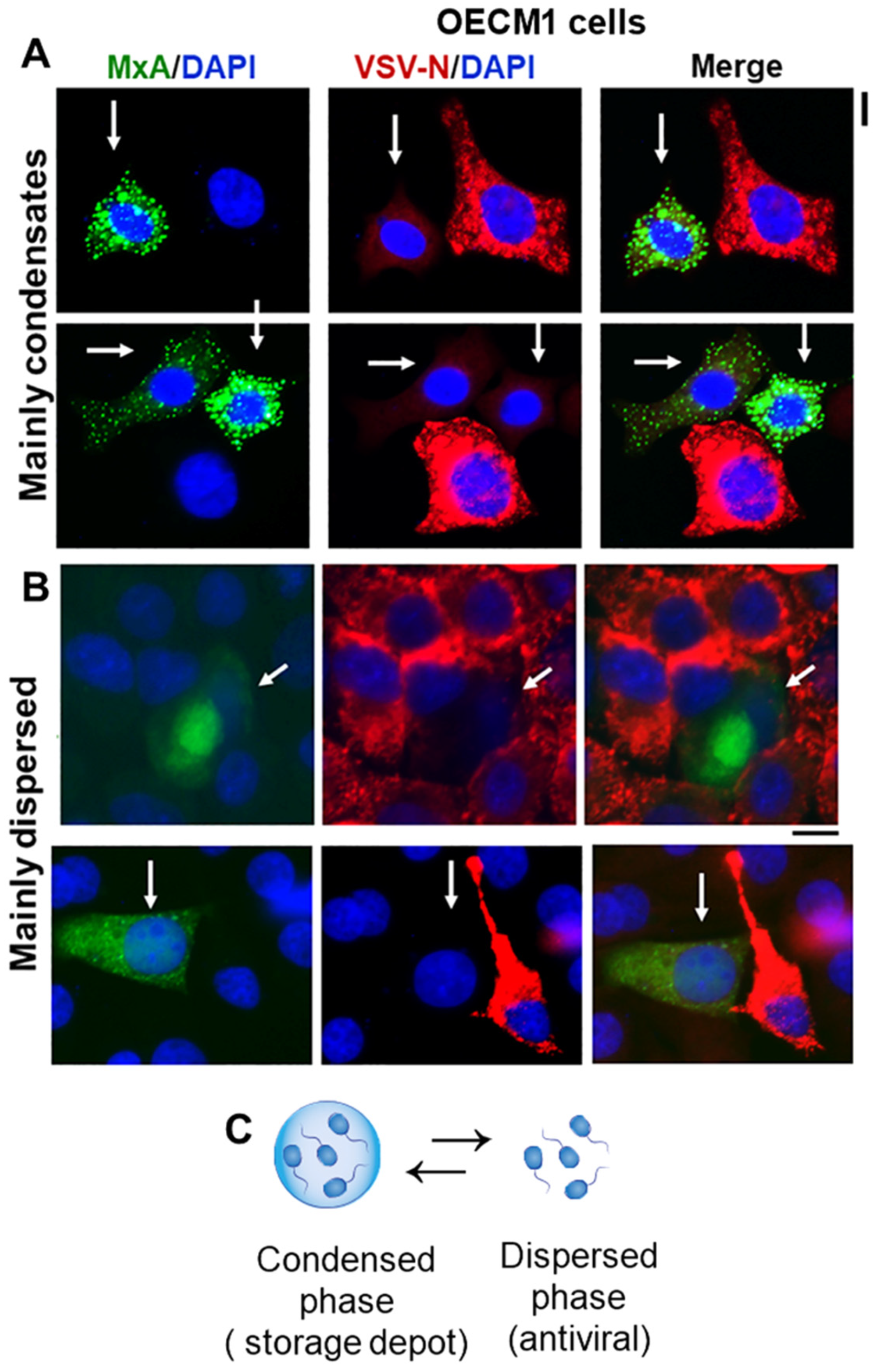
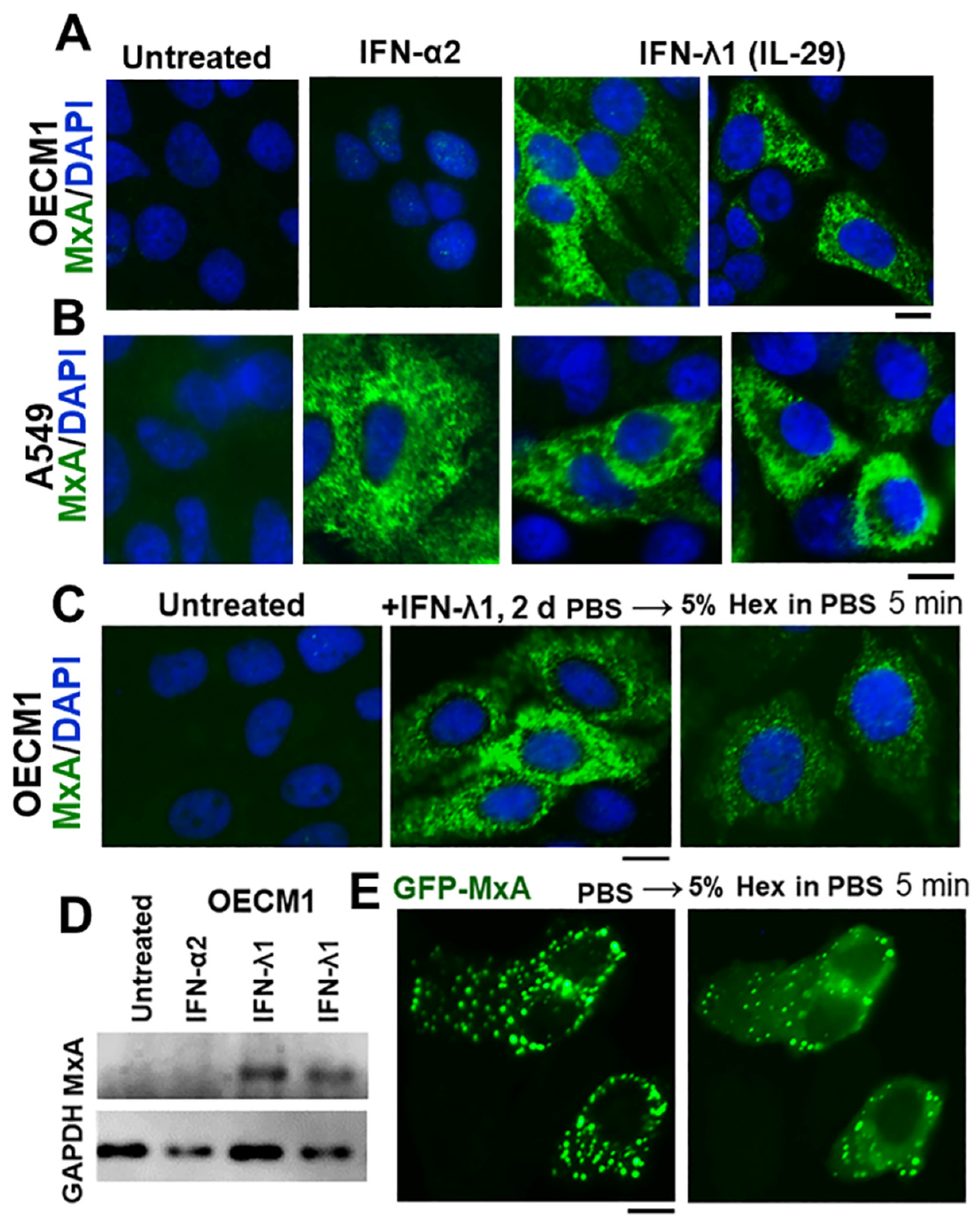
3.1. IFN-λ1 (IL-29), but Not IFN-α2, Induces Expression of MxA to Form Endogenous Cytoplasmic Condensates in Oral Epithelial Cells (OECM1 Cell Line)
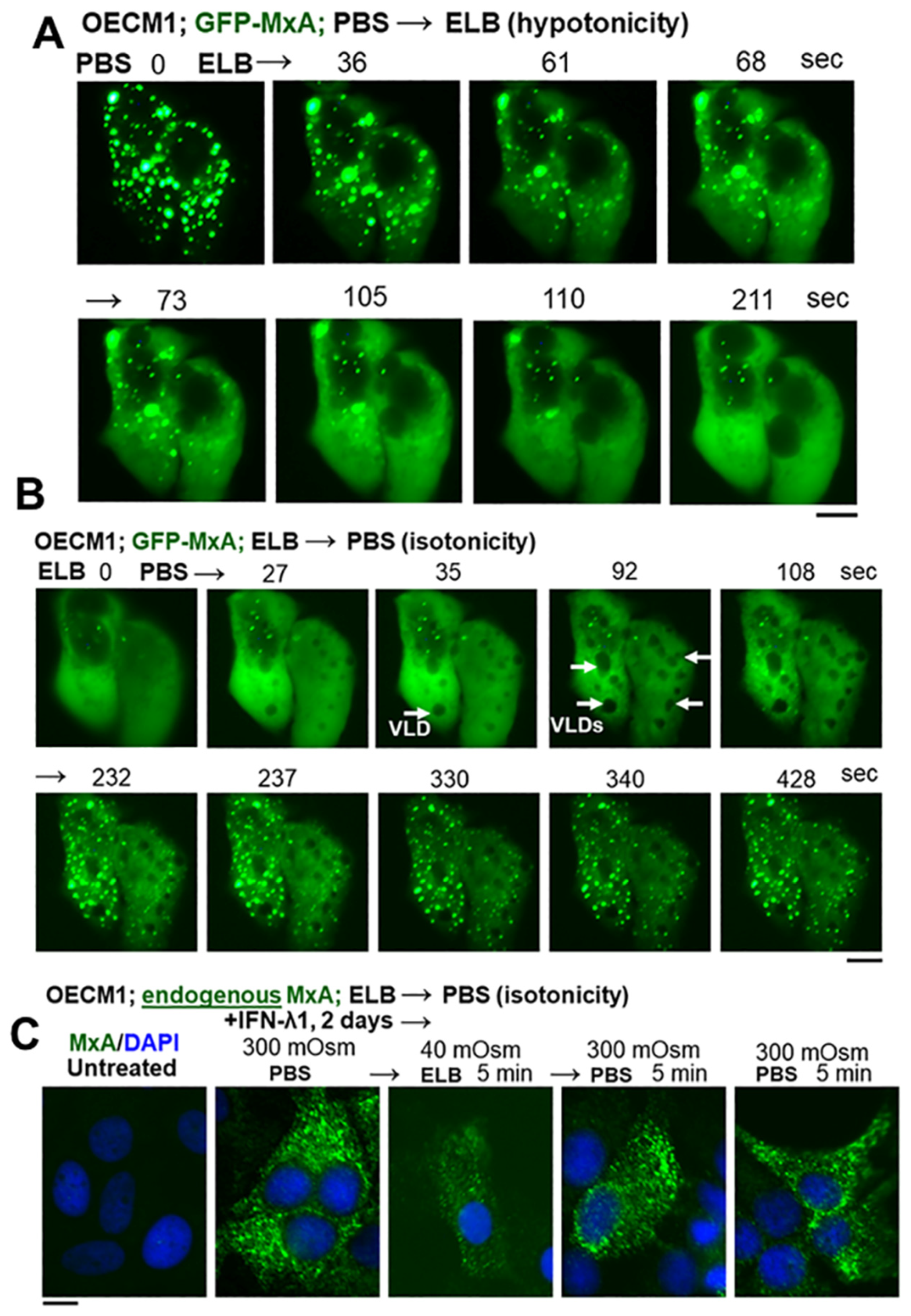
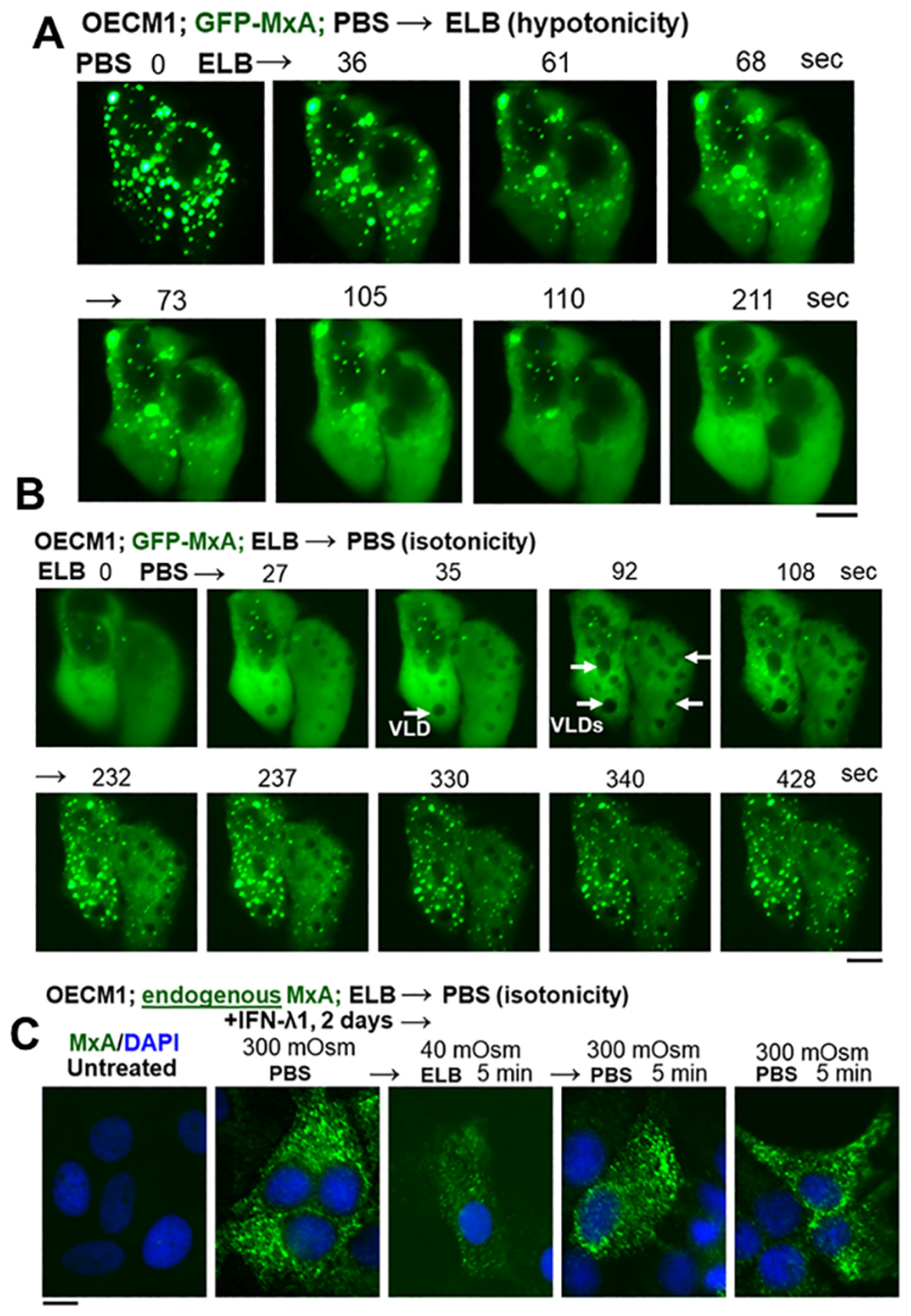
3.2. Hypotonicity Sensing by GFP-MxA and Endogenous MxA Condensates
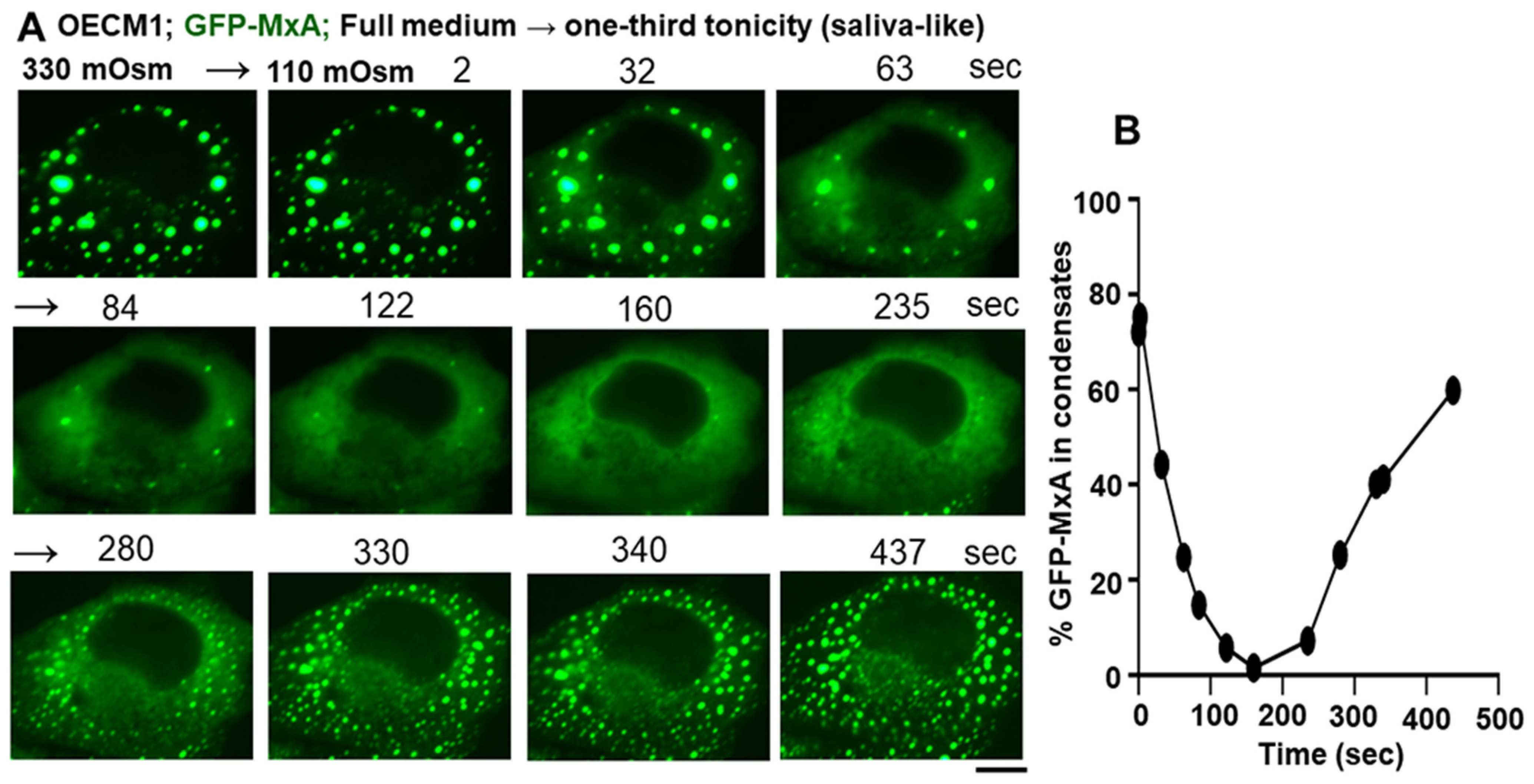
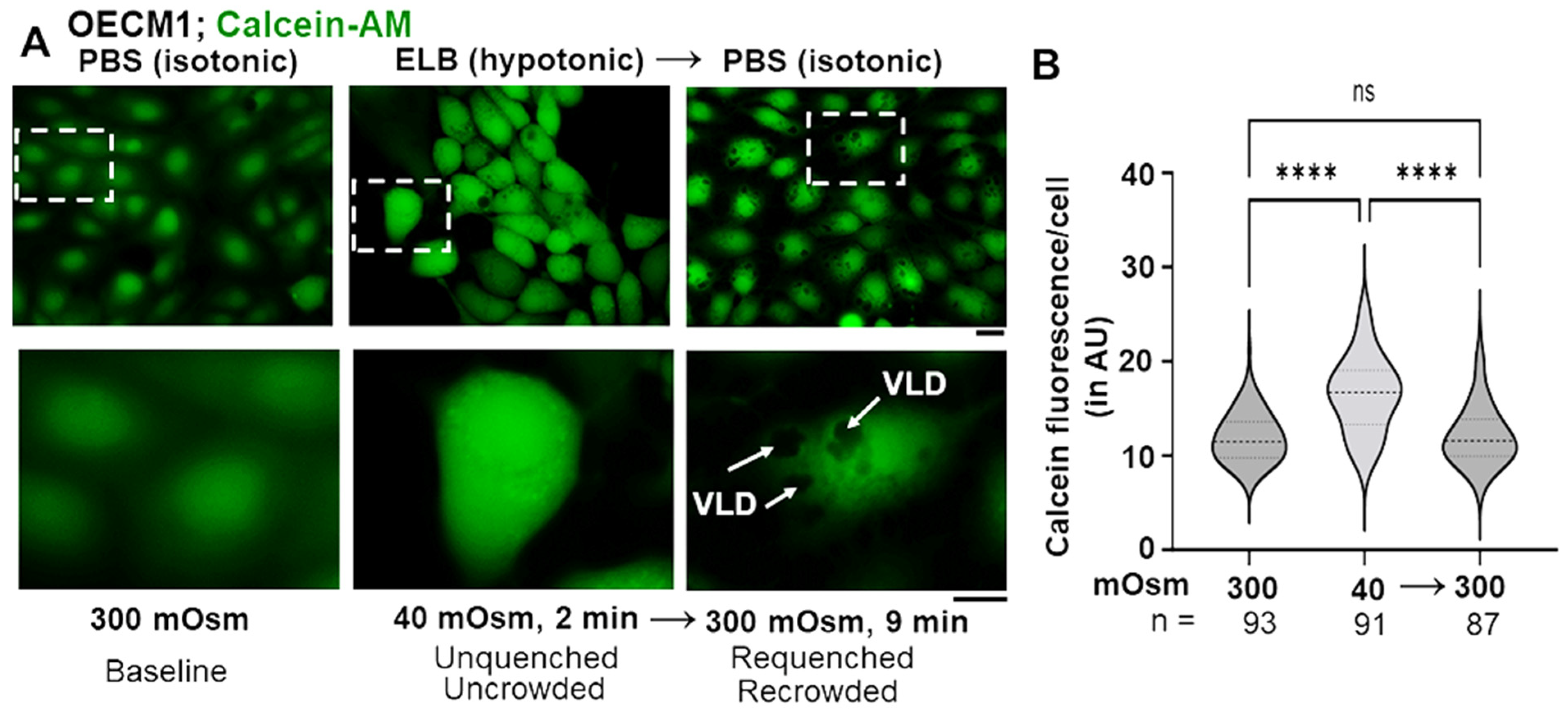
3.3. Biophysical Mechanism: Macromolecular Uncrowding and Recrowding during Regulated Volume Changes
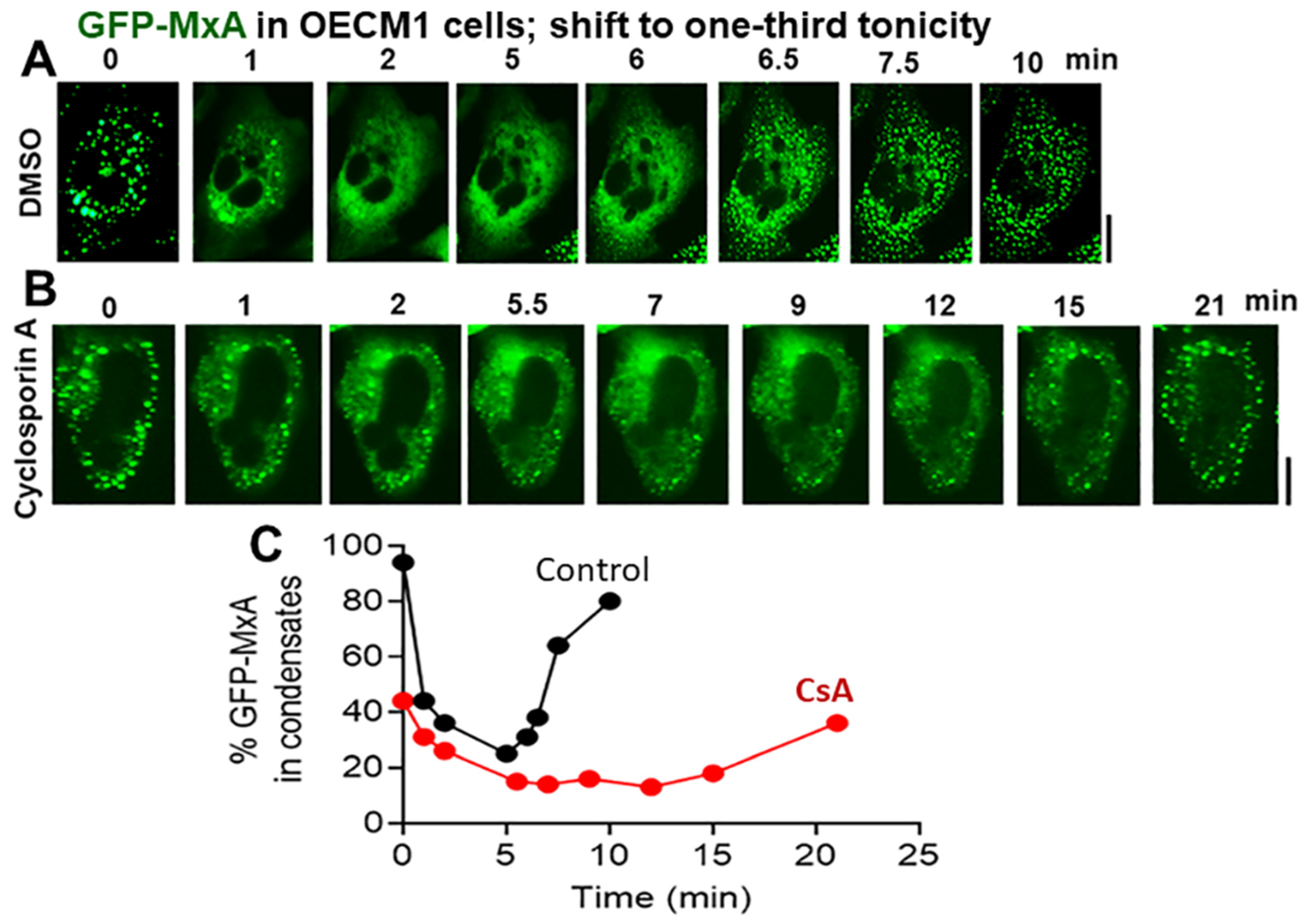
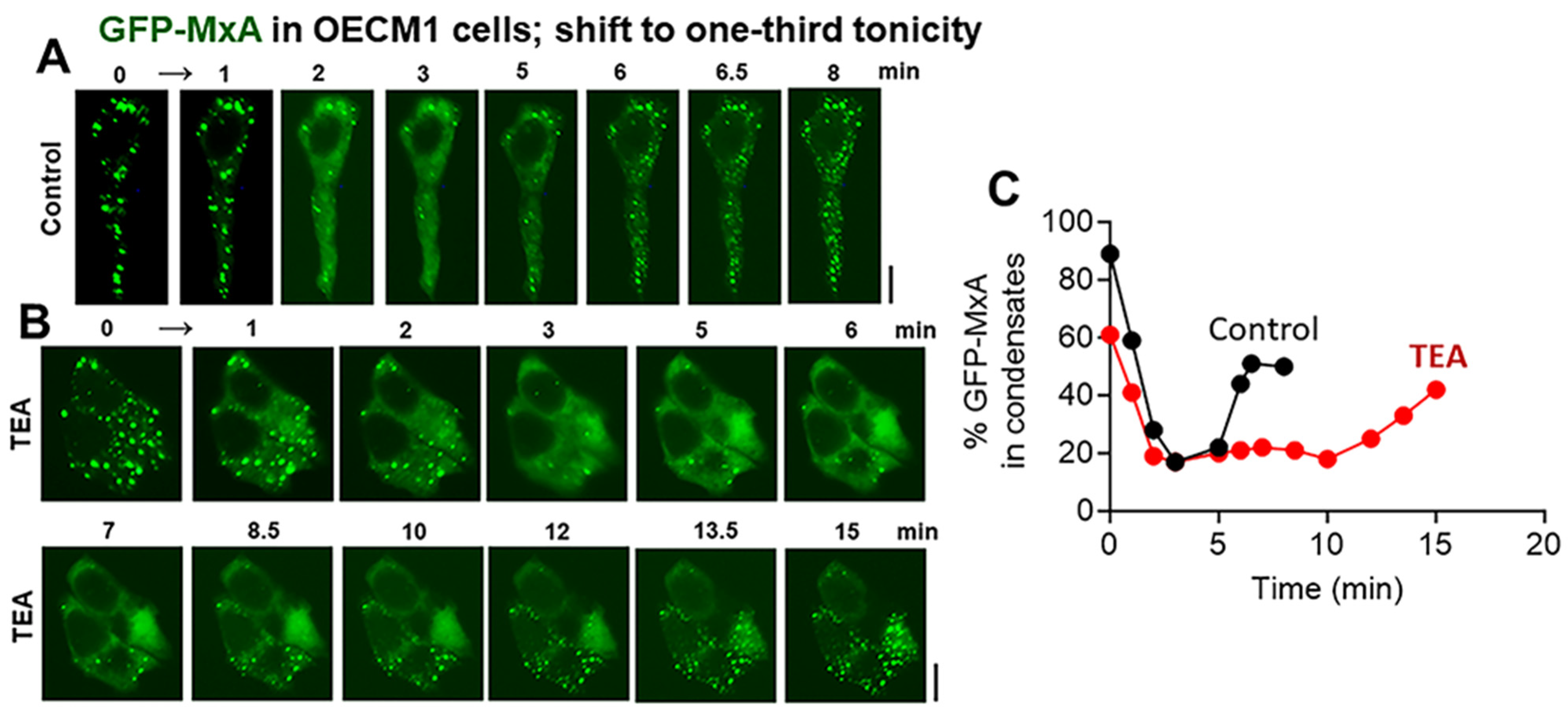
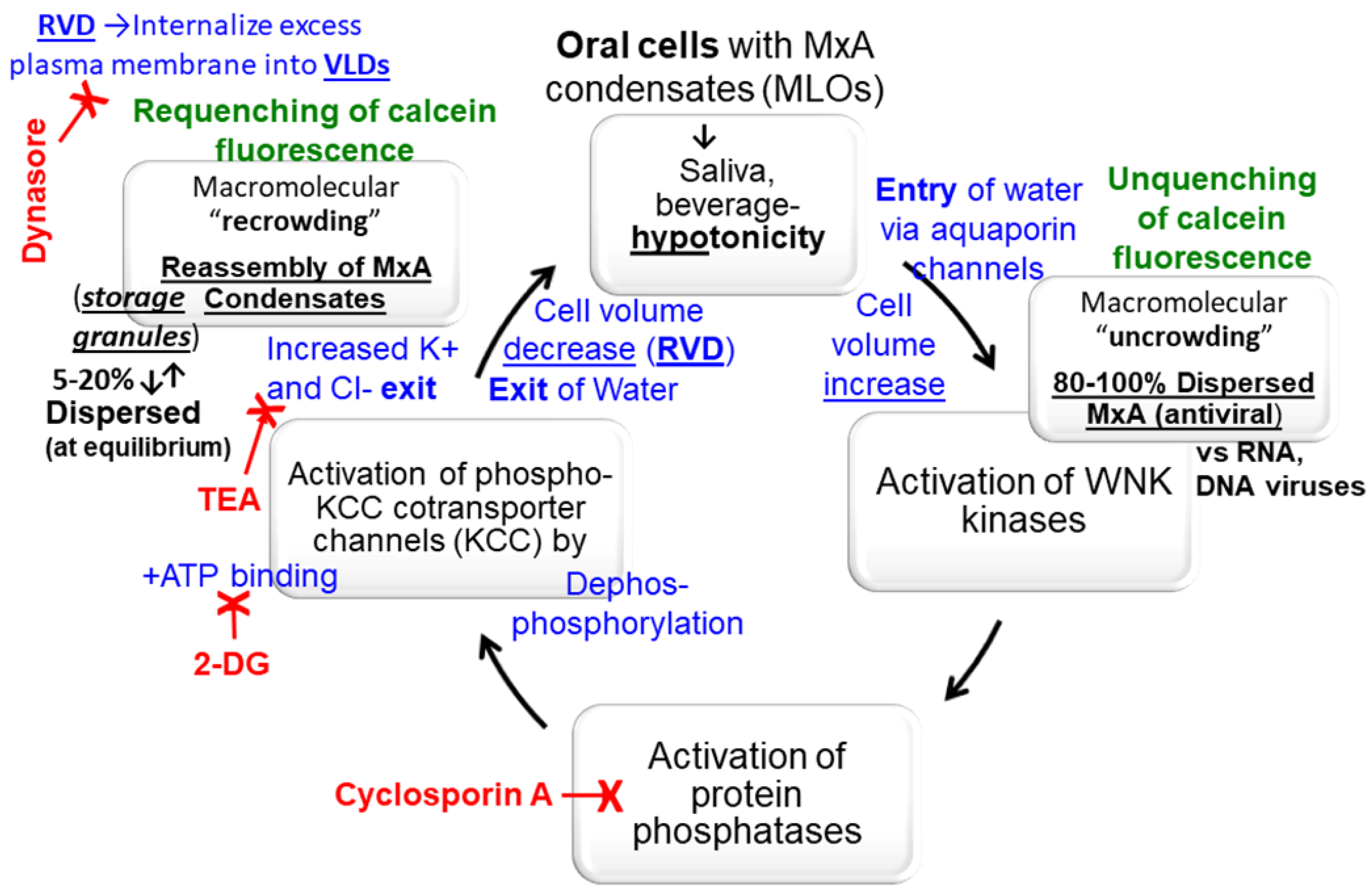
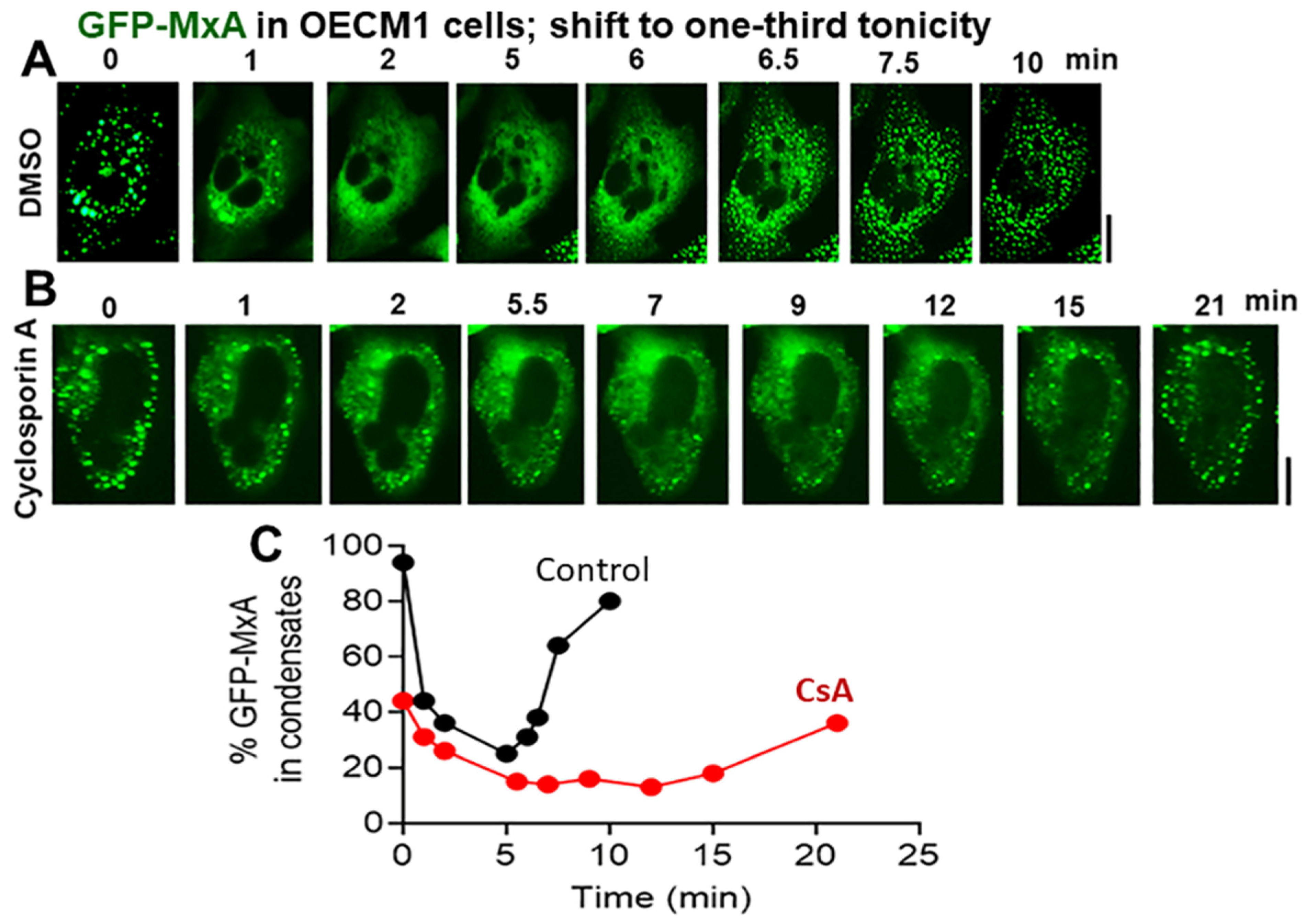
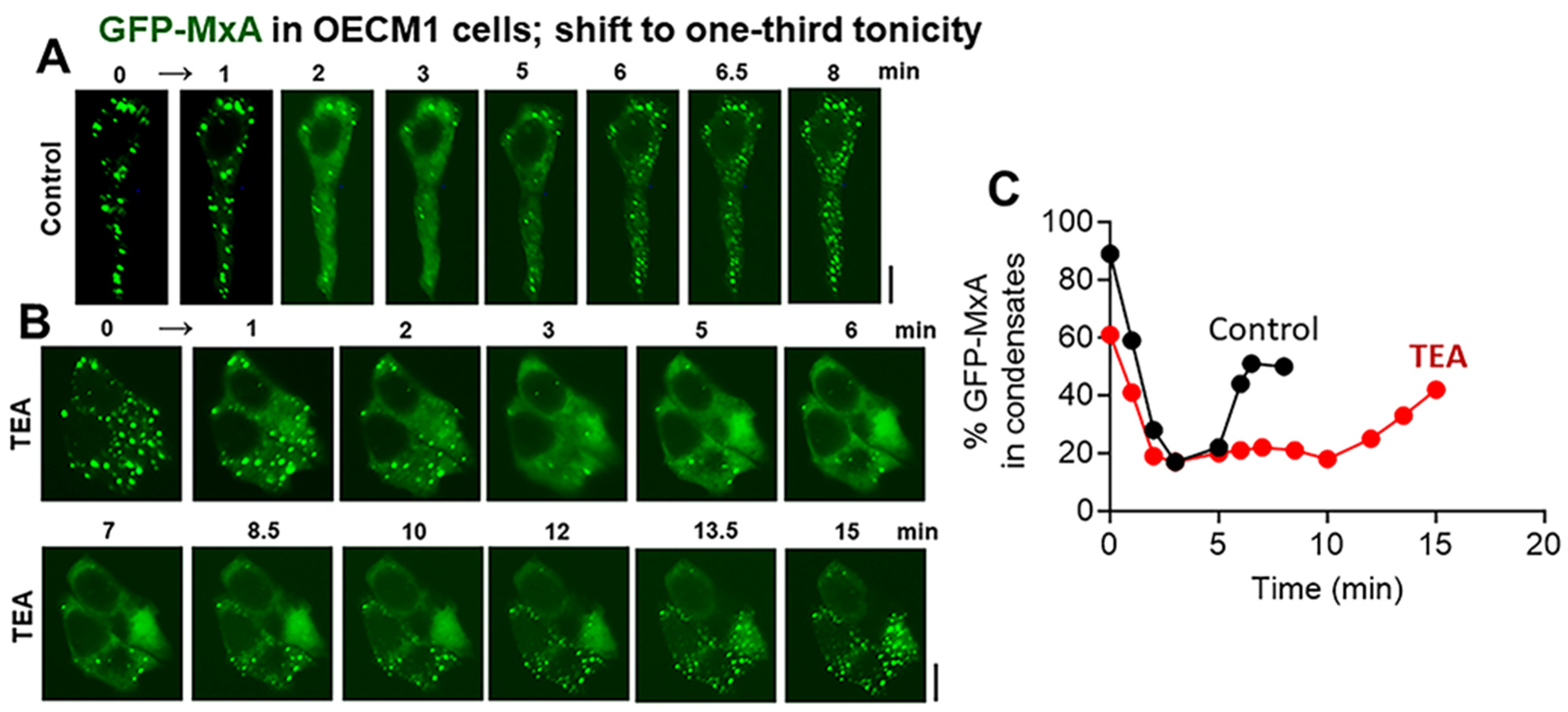
3.4. Biochemical Mechanisms: Volume-Sensitive WNK Kinase-Protein Phosphatase-K-Cl-Cotransporter Pathway
4. Discussion
5. Limitations of This Study
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Rodriguez-Hernandez, C.J.; Sokoloski, K.J.; Stocke, K.S.; Dukka, H.; Jin, S.; Metzler, M.A.; Zaitsev, K.; Shpak, B.; Shen, D.; Miller, D.P.; et al. Microbiome-mediated incapacitation of interferon lambda production in the oral mucosa. Proc. Natl. Acad. Sci. USA 2021, 118, e2105170118. [Google Scholar] [CrossRef] [PubMed]
- Atukorallya, D.S.; Ratyanake, R.K. Oral mucosa, saliva, and COVID-19 infection in oral health care. Front. Med. 2021, 8, 656926. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.T.; Ferreira, M.C.; Guaré, R.O.; Diniz, M.B.; Rösing, C.K.; Rodrigues, J.A.; Duarte, D.A. Gingivitis and salivary osmolality in children with cerebral palsy. Int. J. Paediatr. Dent. 2016, 26, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, L.A.; Diniz, M.B.; Loyola-Rodriguez, J.P.; Habibe, C.H.; Garrubbo, C.C. A controlled study comparing salivary osmolality, caries experience and caries risk in patients with cerebral palsy. Med. Oral. Patol. Oral. Cir. Bucal 2018, 23, e211–e215. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.; Barnett, C. Relationships between the acidity and osmolality of popular beverages and reported postprandial heartburn. Gasteroenterology 1995, 108, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Gresz, V.; Kwon, T.H.; Hurley, P.T.; Varga, G.; Zelles, T.; Nielsen, S.; Case, R.M.; Steward, M.C. Identification and localization of aquaporin water channels in human salivary glands. Am. J. Physiol. Gastroint Liver Physiol. 2001, 281, G247–G254. [Google Scholar] [CrossRef] [PubMed]
- Mahanonda, R.; Sa-Ard-Iam, N.; Rerkyen, P.; Thitithanyanont, A.; Subbalekha, K.; Pichyangkul, S. MxA expression induced by α-defensin in healthy human periodontal tissue. Eur. J. Immunol. 2012, 42, 946–956. [Google Scholar] [CrossRef] [PubMed]
- Imangulli, M.M.; Swaim, W.D.; League, S.C.; Gress, R.E.; Pavletic SZ Hakim, F.T. Increased T-bet+ cytotoxic effectors and type I interferon-mediated processes in chronic graft-versus-host disease of the oral mucosa. Blood 2009, 113, 3620–3630. [Google Scholar] [CrossRef] [PubMed]
- Holzinger, D.; Jorns, C.; Stertz, S.; Boisson-Dupuis, S.; Thimme, R.; Weidmann, M.; Casanova, J.L.; Haller, O.; Kochs, G. Induction of MxA gene expression by influenza A virus requires Type I or Type III interferon signaling. J. Virol. 2007, 81, 7776–7785. [Google Scholar] [CrossRef]
- Pervolaraki, K.; Rastgou Talemi, S.; Albrecht, D.; Bormann, F.; Bamford, C.; Mendoza, J.L.; Garcia, K.C.; McLauchlan, J.; Höfer, T.; Stanifer, M.L.; et al. Differential induction of interferon stimulated genes between type I and type III interferons is independent of interferon receptor abundance. PLoS Pathog. 2018, 14, e1007420. [Google Scholar] [CrossRef]
- Haller, O.; Kochs, G. Interferon-induced mx proteins: Dynamin-like GTPases with antiviral activity. Traffic 2002, 3, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Haller, O.; Staeheli, P.; Kochs, G. Interferon-induced Mx proteins in antiviral host defense. Biochimie 2007, 89, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Haller, O.; Staeheli, P.; Schwemmle, M.; Kochs, G. Mx GTPases: Dynamin-like antiviral machines of innate immunity. Trends Microbiol. 2015, 23, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Verhelst, J.; Hulpiau, P.; Saelens, X. Mx proteins: Antiviral gatekeepers that restrain the uninvited. Microbiol. Mol. Biol. Rev. 2013, 77, 551–566. [Google Scholar] [CrossRef]
- Staeheli, P.; Pavlovic, J. Inhibition of vesicular stomatitis virus mRNA synthesis by human MxA protein. J. Virol. 1991, 65, 4498–4501. [Google Scholar] [CrossRef] [PubMed]
- Schwemmle, M.; Weining, K.C.; Richter, M.F.; Shumacher, B.; Staeheli, P. Vesicular stomatitis virus transcription inhibited by purified MxA protein. Virology 1996, 206, 545–554. [Google Scholar] [CrossRef]
- Steiner, F.; Pavlovic, J. Subcellular localization of MxB determines its antiviral potential against influenza virus. J. Virol. 2020, 94, e00125-220. [Google Scholar] [CrossRef] [PubMed]
- DiPaolo, C.; Hefti, H.P.; Meli, M.; Landis, H.; Pavlovic, J. Intramolecular backfolding of the carboxyl-terminal end of MxA protein is a prerequisite for its oligomerization. J. Biol. Chem. 1999, 274, 32071–32078. [Google Scholar] [CrossRef]
- Davis, D.; Yuan, H.; Liang, F.X.; Yang, Y.M.; Westley, J.; Petzold, C.; Dancel-Manning, K.; Deng, Y.; Sall JSehgal, P.B. Human antiviral protein MxA forms novel metastable membraneless cytoplasmic condensates exhibiting rapid reversible tonicity-driven phase transitions. J. Virol. 2019, 93, e01014-19. [Google Scholar] [CrossRef]
- Sehgal, P.B.; Westley, J.; Lerea, K.M.; DiSenso-Browne, S.; Etlinger, J.D. Biomolecular condensates in cell biology and virology: Phase-separated membraneless organelles (MLOs). Anal. Biochem. 2020, 597, 113691. [Google Scholar] [CrossRef]
- Sehgal, P.B.; Lerea, K.L. Biomolecular condensates and membraneless organelles (MLOs). In Encyclopedia of Biological Chemistry, 3rd ed.; Joseph, J., Ed.; Elsevier: Oxford, UK, 2020; Volume 1, pp. 530–541. [Google Scholar] [CrossRef]
- Sehgal, P.B.; Yuan, H.; Scott, M.F.; Deng, Y.; Liang, F.-X.; Mackiewicz, A. Murine GFP-Mx1 forms phase-separated nuclear condensates and associates with cytoplasmic intermediate filaments: Novel antiviral activity against vesicular stomatitis virus. J. Biol. Chem. 2020, 294, 15218–15234. [Google Scholar]
- Sehgal, P.B. Metastable biomolecular condensates of interferon-inducible antiviral Mx-family GTPases: A paradigm shift in the last three years. J. Biosci. 2021, 46, 72. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, P.B.; Yuan, H.; Jin, Y. Rapid reversible osmoregulation of cytoplasmic biomolecular condensates of human interferon-α-induced antiviral MxA GTPase. Int. J. Mol. Sci. 2022, 23, 12739. [Google Scholar] [CrossRef] [PubMed]
- Sadowska, A.; Swiderski, F.; Laskowski, W. Osmolality of components and their application in the design of functional recovery drinks. Appl. Sci. 2020, 10, 7663. [Google Scholar] [CrossRef]
- Shen, M.R.; Chou, C.Y.; Ellory, J.C. Volume-sensitive KCl cotransport associated with human cervical carcinogenesis. Pflügers Arch. 2000, 440, 751–760. [Google Scholar] [CrossRef]
- Bize, I.; Guvenc, B.; Robb, A.; Buchbinder, G.; Brugnara, C. Serine/threonine protein phosphatases a2d regulation of K-Cl cotransport in human erythrocytes. Am. J. Physiol. 1999, 277, C926–C936. [Google Scholar] [CrossRef] [PubMed]
- Akella, R.; Humphreys, J.M.; Sekulski, K.; He, H.; Durbacz, M.; Chakravarthy, S.; Liwocha, J.; Mohammed, Z.J.; Brautigam, C.A.; Goldsmith, E.J. Osmosensing by WNK kinases. Mol. Biol. Cell 2021, 32, 1614–1623. [Google Scholar] [CrossRef] [PubMed]
- Boyd-Shiwarski, C.R.; Shiwarski, D.J.; Griffiths, S.E.; Beacham, R.T.; Norrell, L.; Morrison, D.E.; Wang, J.; Mann, J.; Tennant, W.; Anderson, E.N.; et al. WNK kinases sense molecular crowding and rescue cell volume via phase separation. Cell 2022, 185, 4488–4506. [Google Scholar] [CrossRef] [PubMed]
- Carey, B.L.; Ahmed, M.; Puckett, S.; Lyles, D.S. Early steps of the virus replication cycle are inhibited in prostate cancer cells resistant to oncolytic vesicular stomatitis virus. J. Virol. 2008, 82, 12104–12115. [Google Scholar] [CrossRef]
- Yool, A.J.; Brokl, O.H.; Pannabecker, T.L.; Dantzler, W.H.; Stamer, W.D. Tetraethylammonium block of water flux in aquaporin-1 channels expressed in kidney thin limbs of Henle’s loop and a kidney-deruived cell line. BMC Physiol. 2002, 2, 4. [Google Scholar] [CrossRef]
- Reuzeau, C.; Mills, L.R.; Harris, J.A.; Morris, C.E. Discrete and reversible vacuole-like dilations induced by osmomechanical perturbations of neurons. J. Membr. Biol. 1995, 145, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Mola, M.G.; Sparaneo, A.; Gargano, C.D.; Spray, D.C.; Svelto, M.; Frigeri, A.; Scemes, E.; Nicchia, G.P. The speed of swelling kinetics modulates cell volume regulation and calcium signaling in astrocytes: A different point of view on the role of aquaporins. Glia 2016, 64, 139–154. [Google Scholar] [CrossRef]
- de los Heros, P.; Kahle, K.T.; Rinehart, J.; Bobadilla, N.A.; Vazquez, N.; Cristobal, P.S.; Mount, D.B.; Lifton, R.P.; Hebert, S.C.; Gamba, G. WNK3 bypasses the tonicity requirement for K-Cl cotransporter activation via a phosphatase-dependent pathway. Proc. Natl. Acad. Sci. USA 2006, 103, 1976–1981. [Google Scholar] [CrossRef]
- Stertz, S.; Reichelt, M.; Krijnse-Locker, J.; Mackenzie, J.; Simpson, J.C.; Haller, O.; Kochs, G. Interferon-induced, antiviral human MxA protein localizes to a distinct subcompartment of the smooth endoplasmic reticulum. J. Interferon Cytokine Res. 2006, 26, 650–660. [Google Scholar] [CrossRef]
- Accola, M.A.; Huang, B.; Al Masri, A.; McNiven, M.A. The antiviral dynamin family member, MxA, tubulates lipids and localizes to the smooth endoplasmic reticulum. J. Biol. Chem. 2002, 277, 21829–21835. [Google Scholar] [CrossRef]
- Banani, S.F.; Lee, H.O.; Hyman, A.A.; Rosen, M.K. Biomolecular condensates: Organizers of cellular biochemistry. Nat. Rev. Mol. Cell Biol. 2017, 18, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.; Brangwynne, C.P. Liquid phase condensation in cell physiology and disease. Science 2017, 357, eaaf4382. [Google Scholar] [CrossRef]
- Alberti, S. The wisdom of crowds: Regulating cell function through condensed states of living matter. J. Cell Sci. 2017, 130, 2789–2796. [Google Scholar] [CrossRef] [PubMed]
- Alberti, S.; Gladfelter, A.; Mittag, T. Considerations and challenges in studying liquid-liquid phase separation and biomolecular condensates. Cell 2019, 176, 419–434. [Google Scholar] [CrossRef]
- Shin, Y.; Berry, J.; Pannucci, N.; Haataja NP Toettcher, J.E.; Brangwynne, C.P. Spatiotemporal control of intracellular phase transitions using light-activated optoDroplets. Cell 2017, 168, 159–171. [Google Scholar] [CrossRef]
- Heinrich, B.S.; Maliga, Z.; Stein, D.A.; Hyman, A.A.; Whelan, S.P.J. Phase transitions drive the formation of vesicular stomatitis virus replication compartments. mBio 2018, 9, e02290-17. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.R.; Asfaha, J.B.; Ghent, C.M.; Howard, C.J.; Hartooni, N.; Safari, M.; Frankel, A.D.; Morgan, D.O. Phosphoregulation of phase separation by the SARS-CoV-2 N protein suggests a biophysical basis for its dual functions. Mol. Cell 2020, 80, 1092–1103.e4. [Google Scholar] [CrossRef] [PubMed]
- Dick, A.; Graf, L.; Olal, D.; von der Malsburg, A.; Gao, S.; Kochs, G.; Daumke, O. Role of nucleotide binding and GTPase domain dimerization in dynamin-like myxovirus resistance protein A for GTPase activation and antiviral activity. J. Biol. Chem. 2015, 290, 12779–12792. [Google Scholar] [CrossRef] [PubMed]
- von der Malsburg, A.; Abutbul-Ionita, I.; Haller, O.; Kochs, G.; Danino, D. Stalk domain of the dynamin-like MxA GTPase protein mediates membrane binding and liposome tubulation via the unstructured L4 loop. J. Biol. Chem. 2011, 286, 37858–37865. [Google Scholar] [CrossRef]
- Moschonas, G.D.; Delhaye, L.; Cooreman, R.; Bhat, A.; Sutter, D.D.; Parthoens, E.; Desmer, A.; Maciejczak, A.; Grzek, H.; Lippens, S.; et al. MX2 restricts HIV-1 and herpes simplex virus-1 by forming cytoplasmic biomolecular conednsates that mimic nuclear pore complexes. bioRxiv 2023. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sehgal, P.B.; Yuan, H.; Centone, A.; DiSenso-Browne, S.V. Oral Antiviral Defense: Saliva- and Beverage-like Hypotonicity Dynamically Regulate Formation of Membraneless Biomolecular Condensates of Antiviral Human MxA in Oral Epithelial Cells. Cells 2024, 13, 590. https://doi.org/10.3390/cells13070590
Sehgal PB, Yuan H, Centone A, DiSenso-Browne SV. Oral Antiviral Defense: Saliva- and Beverage-like Hypotonicity Dynamically Regulate Formation of Membraneless Biomolecular Condensates of Antiviral Human MxA in Oral Epithelial Cells. Cells. 2024; 13(7):590. https://doi.org/10.3390/cells13070590
Chicago/Turabian StyleSehgal, Pravin B., Huijuan Yuan, Anthony Centone, and Susan V. DiSenso-Browne. 2024. "Oral Antiviral Defense: Saliva- and Beverage-like Hypotonicity Dynamically Regulate Formation of Membraneless Biomolecular Condensates of Antiviral Human MxA in Oral Epithelial Cells" Cells 13, no. 7: 590. https://doi.org/10.3390/cells13070590
APA StyleSehgal, P. B., Yuan, H., Centone, A., & DiSenso-Browne, S. V. (2024). Oral Antiviral Defense: Saliva- and Beverage-like Hypotonicity Dynamically Regulate Formation of Membraneless Biomolecular Condensates of Antiviral Human MxA in Oral Epithelial Cells. Cells, 13(7), 590. https://doi.org/10.3390/cells13070590