Abstract
The rate of spontaneous action potentials (APs) generated by sinoatrial node cells (SANC) is regulated by local Ca2+ release (LCR) from the sarcoplasmic reticulum via Ca2+ release channels (ryanodine receptors, RyRs). LCR events propagate and self-organize within the network of RyR clusters (Ca release units, CRUs) via Ca-induced-Ca-release (CICR) that depends on CRU sizes and locations: While larger CRUs generate stronger release signals, the network’s topology governs signal diffusion and propagation. This study used super-resolution structured illumination microscopy to image the 3D network of CRUs in rabbit SANC. The peripheral CRUs formed a spatial mesh, reflecting the cell surface geometry. Two distinct subpopulations of CRUs were identified within each cell, with size distributions conforming to a two-component Gamma mixture model. Furthermore, neighboring CRUs exhibited repulsive behavior. Functional properties of the CRU network were further examined in a novel numerical SANC model developed using our experimental data. Model simulations revealed that heterogeneities in both CRU sizes and locations facilitate CICR and increase the AP firing rate in a cooperative manner. However, these heterogeneities reduce the effect of β-adrenergic stimulation in terms of its relative change in AP firing rate. The presence of heterogeneities in both sizes and locations allows SANC to reach higher absolute AP firing rates during β-adrenergic stimulation. Thus, the CICR facilitation by heterogeneities in CRU sizes and locations regulates and optimizes cardiac pacemaker cell operation under various physiological conditions. Dysfunction of this optimization could be a key factor in heart rate reserve decline in aging and disease.
1. Introduction
Sinoatrial node cells (SANCs) located in the posterosuperior right atrium are specialized to generate rhythmic electrical impulses that drive cardiac contractions [1,2]. SANC dysfunction can lead to sick sinus syndrome, which is associated with a variety of life-threatening arrhythmias [3,4,5,6]. The generation of normal rhythmic impulses by SANC is executed via coupled signaling of both cell membrane ion channels and Ca2+ cycling, known as the coupled-clock system [7]. A vital component of this system is the sarcoplasmic reticulum (SR), the main intracellular Ca2+ store consisting of two parts: (i) the network SR that uptakes cytosolic Ca2+ via a Ca2+ pump and (ii) the junctional SR (JSR) that embeds clusters of 10–200 Ca2+ release channels (ryanodine receptors, RyRs [8]), forming functional couplons or Ca2+ release units (CRUs) linked to L-type channels [9]. While RyRs release Ca2+ from JSR, their openings are also activated by Ca, creating positive (i.e., explosive) feedback known as Ca-induced-Ca release (CICR) [10]. Thus, RyRs within a CRU tend to release Ca2+ in synchrony, creating an elementary Ca2+ signal known as a Ca2+ spark [11,12]. In SANCs under normal conditions, a Ca2+ spark, in turn, can interact with neighboring CRUs via CICR, creating a series of locally propagating sparks observed as Ca2+ wavelets, known as local Ca2+ release (LCR) (review [7]). The LCRs, in turn, activate an electrogenic Na/Ca exchanger embedded in the cell surface membrane that accelerates diastolic depolarization [7,13,14,15]. The diastolic depolarization is further accelerated by a positive feedback mechanism that includes voltage-activated L-type Ca2+ channels which not only depolarize the membrane, but also bring more Ca2+ into the cell. Thus, the Ca2+ influx brings more “fuel” to further explode CICR and diastolic depolarization, ensuring robust action potential (AP) ignition [16].
In this complex chain of signaling events, CICR plays a critical role, and its efficiency must strongly depend on the spatial organization of CRU network, i.e., the distributions of CRU sizes and locations. What is known thus far about the CRU network in SANC and what is still missing? RyRs are indeed clustered under the cell membrane, as observed in immunofluorescence images of SANCs of various species [17,18]. Early simple numerical models of CRU networks in SANCs considered identical CRUs in a perfect square grid [19,20]. These models demonstrated an important role of CICR in cardiac pacemaking, e.g., the emergence of a Ca2+ clock and robust AP firing (via stabilizing diastolic Na/Ca exchanger current [20]). Subsequentially, distributions of CRU sizes and nearest-neighbor distances (NNDs) were measured in 2D in tangential sections of confocal microscopy images and included in a 3D model of SANC [21]. That study raised important questions regarding CRU function, such as “when can a Ca2+ spark jump?”, i.e., when CICR propagates among neighboring CRUs. CICR and AP firing emerged in the model only when larger-sized CRUs were mixed with another population of smaller CRUs that provided bridges for CICR propagation, indicating the importance of CRU size heterogeneity. More recent theoretical studies have demonstrated that disorder in CRU locations confers robustness but reduces flexibility in heart pacemaking [22], showing the importance of heterogeneity in CRU distances. This was shown, however, in a SANC model with CRUs of identical sizes and hypothetical CRU repulsion, whereas precise organization of the CRU network in 3D and the functional role of heterogeneity of CRU sizes remain unknown. Thus, it is still not understood how the Ca2+ clock and the coupled-clock system emerge from the scale and organization of RyRs and CRUs towards the whole-cell function [23,24].
The present study approaches this problem by combining experimental methods and numerical modeling. We imaged the precise organization of the CRU network (both sizes and locations) in 3D in rabbit SANCs by super-resolution structured illumination microscopy (SIM), which provided a greater optical (about twice) resolution compared to confocal imaging [25]. Our results revealed that CRU sizes follow a mixture of two Gamma distributions, suggesting distinct subpopulations of smaller and larger CRUs. Furthermore, NNDs of RyR clusters exhibit a power law distribution at short distances, indicating repulsive interactions among CRUs that would restrain CICR and Ca2+ signal propagation. Based on these findings, we developed a new mathematical SANC model featuring CRUs of heterogeneous sizes and locations and performed model simulations with different CRU networks to obtain deeper insights. The simulations demonstrated that the complex and flexible structure of the RyR network, including partial disorder, represents a new pacemaker regulatory mechanism that optimizes cardiac pacemaker cell operation under various physiological conditions.
2. Materials and Methods
2.1. Enzymatic Isolation of Individual SANC
SANCs were isolated from male rabbits in accordance with NIH guidelines for the care and use of animals, protocol # 457-LCS-2024, as previously described [26]. New Zealand white rabbits (Charles River Laboratories, Wilmington, MA, USA) weighing 2.8–3.2 kg were anesthetized with sodium pentobarbital (50–90 mg/kg). The hearts were removed quickly and placed in solution containing (in mM): 130 NaCl, 24 NaHCO3, 1.2 NaH2PO4, 1.0 MgCl2, 1.8 CaCl2, 4.0 KCl, and 5.6 glucose equilibrated with 95% O2/5% CO2 (pH 7.4 at 35 °C). The SAN region was cut into small strips (~1.0 mm wide) perpendicular to the crista terminalis and excised. The final SA node preparation, which consisted of SA node strips attached to the small portion of crista terminalis, was washed twice in nominally Ca2+-free solution containing (in mM) 140 NaCl, 5.4 KCl, 0.5 MgCl2, 0.33 NaH2PO4, 5 HEPES, and 5.5 glucose (pH = 6.9) and incubated on a shaker at 35 °C for 30 min in the same solution with the addition of elastase type IV (0.6 mg/mL; Sigma, Chemical Co., St. Louis, MO, USA), collagenase type 2 (0.8 mg/mL; Worthington, NJ, USA), Protease XIV (0.12 mg/mL; Sigma, Chemical Co.), and 0.1% bovine serum albumin (Sigma, Chemical Co.). The SA node preparation was next placed in modified “Kraftbruhe” solution, containing (in mM) 70 potassium glutamate, 30 KCl, 10 KH2PO4, 1 MgCl2, 20 taurine, 10 glucose, 0.3 EGTA, and 10 HEPES (titrated to pH 7.4 with KOH), and kept at 4 °C for 1 h in KB solution containing 50 mg/mL polyvinylpyrrolidone (PVP, Sigma, Chemical Co.). Finally, cells were dispersed from the SA node preparation by gentle pipetting in the “Kraftbruhe” solution and stored at 4 °C.
2.2. Immunolabeling of RyRs
Isolated cells were fixed with 2% formaldehyde in phosphate-buffered saline (PBS, Sigma, St. Louis, MO) and then permeabilized with 0.5–1% Triton X-100/PBS. Nonspecific cross-reactivity was blocked by incubating the samples for 4 h in 1% BSA/PBS (Jackson ImmunoResearch, West Grove, PA, USA). The cells were then incubated with primary anti-RyR antibodies (IgG1, clone C3-33, Afinity BioReagents, Golden, CO, or AP20325PU-N (Origene, Rockville, MD, USA)); for details, see our database at https://doi.org/10.7910/DVN/N0OLGI, (accessed 13 November 2024). The antibodies were diluted in 1% BSA/PBS overnight, washed in PBS, and then incubated with appropriate fluorescence label-conjugated secondary antibodies (Jackson ImmunoResearch, West Grove, PA, USA).
2.3. Structured Illumination Microscopy (SIM)
SIM uses a sharply patterned light source to improve spatial resolution in fluorescence microscopy [27]. SIM can be used to study the structure and function of cells in great detail, as it can provide images with higher resolutions than those of traditional microscopy techniques. It is particularly useful for studying cells and tissues that have fine structural features, such as the SANC, as we report here. Experiments with RyR detection in fixed cells were performed in the experimental instant SIM setup in Hari Shroff’s laboratory at the National Institute of Biomedical Imaging and Bioengineering, as described elsewhere [27,28,29]. The instant SIM is an implementation of SIM well-suited for high-speed imaging, as the images are processed optically rather than computationally to improve the resolution ~1.4×. SANCs were imaged via an oil ×100, 1.49 NA objective and 488 nm excitation; in-house software controlled the optimal laser power and positioning (sectioning) of cell preparation. The system allowed for imaging at resolutions as low as 145 nm laterally and 320 nm axially. Three-dimensional sectioning was performed with a pixel size of 55.5 × 55.5 nm in xy and z stacking of 150 nm.
2.4. Segmentation of Peripheral RyR Clusters in 3D
Our dataset consisted of 31 rabbit SANCs, which included an aggregated total of 70,982 RyR clusters. We present a novel algorithm for high-throughput segmentation of peripheral ryanodine receptor (RyR) clusters in 3D SANCs (Figure 1). The preprocessing procedure included the following steps in order: a 3D median filter to remove salt-and-pepper noise, signal normalization to standardize the intensity range, and finally, 3D contrast-limited adaptive histogram equalization (CLAHE) [30] to enhance local contrast while preserving overall image structure and global contrast. The segmentation was then performed using the 3D StarDist neural network [31], which directly predicts star-convex polyhedra representations for each RyR cluster without relying on the watershed algorithm. The 3D StarDist network learns to separate partially overlapping or contacting RyR clusters by predicting appropriate polyhedra shapes based on the trained model. The ground truth data used for training 3D StarDist were initially generated using Squassh 1.0.24 [32], an open-source software algorithm in ImageJ Fiji Release Version 1.54f, which is part of the MOSAIC suite that utilizes globally optimal detection and segmentation methodologies [33] and incorporates corrections for the microscope’s point spread function [34,35]. The Squassh segmentation results were then fine-tuned using the Adaptive Watershed tool in ORS Dragonfly (2024.1) [36] and further refined by manual editing in ORS Dragonfly to ensure accurate separation of RyR clusters (Figure 2). The final trained model for 3D StarDist achieved an Intersection over Union (IoU) score of 0.68.
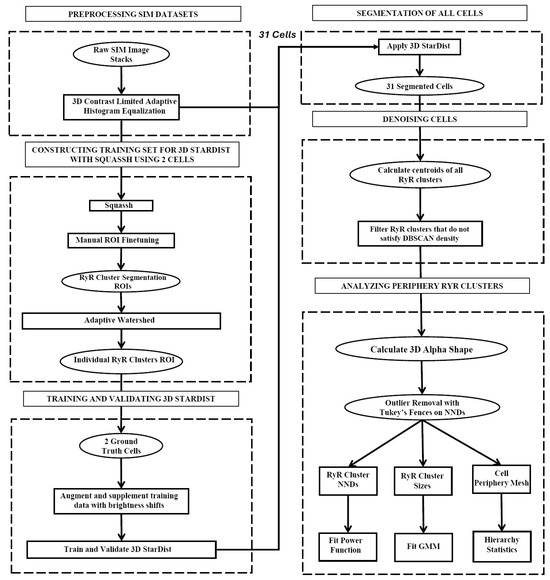
Figure 1.
Flowchart of the peripheral RyR cluster analysis algorithm. Presented is our image-processing algorithm for the precise segmentation and analysis of peripheral RyR clusters in 3D SIM imaging data in SANCs. Initially, the data undergo cropping and contrast enhancement through 3D CLAHE. The main segmentation process is conducted using the 3D StarDist neural network, which is trained using ground truth data generated through the Squassh software and refined via adaptive watershed. Following segmentation, RyR clusters that are part of the cell are identified and extracted using the density-based spatial clustering of applications with noise (DBSCAN) algorithm, which detects high-density clusters within the dataset. The culmination of this process is the generation of a 3D alpha shape encapsulating the spatial distribution of the RyR clusters on the periphery of SANCs. The peripheral RyR cluster sizes, the distances, and their alpha shape mesh are exported for further statistical processing.

Figure 2.
Visual representation of transformations and segmentation of flowchart. (A): The initial, unprocessed SIM image data (in 3 dimensions) of RyR immunofluorescence in SANC, serving as the primary source for image analysis. (B): The enhanced and denoised raw SIM data. (C): The preliminary segmentation from Squassh with a purple ROI. (D): The stage where distinct RyR clusters are isolated, establishing boundaries between adjoining regions using a 3D 26-connectivity strategy. (E): The Watershed algorithm’s role in further refining the segmentation, capable of isolating individual RyR clusters even in complex spatial arrangements.
To further refine the segmentation of RyR clusters and remove false positives after 3D StarDist segmentation, DBSCAN (Density-Based Spatial Clustering of Applications with Noise) clustering was used on the centroid data of the segmented RyR clusters. DBSCAN is a density-based clustering algorithm that groups together points that are closely packed, marking points that are in low-density regions as outliers. The key parameters for DBSCAN were ϵ and min_samples. ϵ determines the maximum distance between two points for them to be considered as part of the same cluster, or in our case, the imaged SANCs. ϵ was set using the median absolute deviation (MAD) method with a scaling factor c.
By using the scaled MAD and the median, the remained robust against the influence of extreme values:
where A represents the set of nearest-neighbor distances. Furthermore, the scaling factor c was defined as:
where erfcinv is the inverse complementary error function. This adaptation ensures that the scaled MAD approximates the standard deviation when the data follow a normal distribution. The second parameter, min_samples, is the minimum number of points required to form a dense region, and it was set to 3. If a point has at least min_samples within its ϵ-neighborhood (including itself), it is considered a core point. Points that are not core points but are within the ϵ-neighborhood of a core point are called border points. Points that are neither core points nor border points are classified as noise or outliers.
The largest cluster deduced by DBSCAN (which is the cell itself) was further refined by computing its alpha shape, which captured the peripheral RyR clusters. The optimal alpha value was determined by iterating through a range of values and selecting the one that maximized the density of points on the surface of the alpha shape. Finally, the algorithm generated various data and visualizations, including the alpha shape, surface area, volume, centroid data, volume data, voxel lists, cluster labels, adjacency matrix, and nearest-neighbor distances, for further analysis. An additional outlier test was performed by taking Tukey’s Fences where extreme nearest-neighbor values are filtered out by keeping only the interval:
The visualizations for all 31 cells are presented in Figure 3, and mouse-interactive HTML files constructed with plotly Python library are published in GitHub https://github.com/alexmaltsev/SANC/tree/main/3D%20Visualizations (accessed 13 November 2024). Each cell can be individually opened and rotated (with zoom-in or -out) by the reader. We also published our code to use 3D StarDist and all code for data application and processing in GitHub. See the Code Availability section for more details.
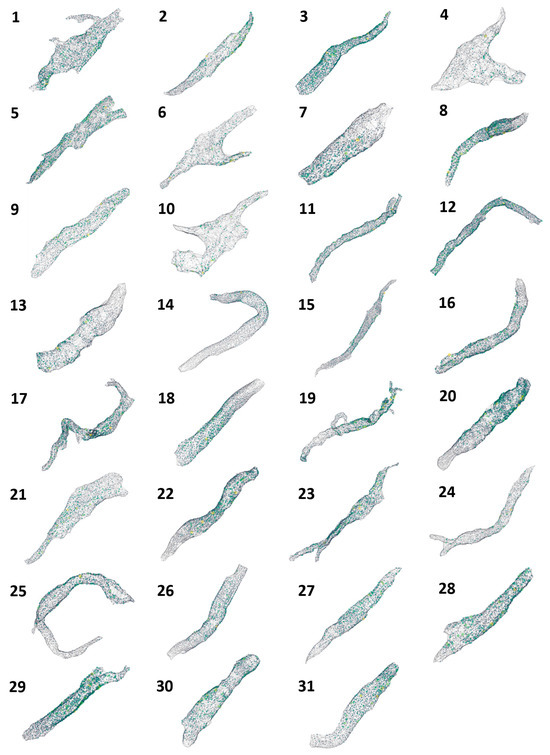
Figure 3.
Three-dimensional visualization of RyR cluster distribution in all 31 rabbit SANCs analyzed in the study. Each cell is represented by a unique number (1–31) corresponding to the cell identifiers used in Table 1 and Table 2. The visualizations depict the spatial distribution of RyR clusters within each cell, with clusters represented as colored points. The color intensity indicates the relative sizes of the clusters, with brighter colors (yellow to green) representing larger clusters and dimmer colors (blue to gray) representing smaller clusters. These visualizations provide a comprehensive overview of the RyR cluster organization across the entire sample set, allowing for visual comparison of cluster distributions and densities among cells of different morphologies. Cells are shown at various zoom levels to closely fit the panel size. Three-dimensional representations of each cell with its scale bar are available in html format in GitHub https://github.com/alexmaltsev/SANC/tree/main/3D%20Visualizations (accessed 13 November 2024). Each html file can be opened in a web browser for a computer mouse-interactive view (rotation and zoom-in and -out).
3. Results
3.1. Two Subpopulations of RyR Cluster Sizes
The distributions of RyR cluster sizes in individual SANC were analyzed, with the results shown in Table 1.

Table 1.
Morphological and RyR cluster characteristics of rabbit sinoatrial node cells. Detailed statistics for 31 rabbit sinoatrial node cells, including their surface area, volume, number of RyR clusters, and metrics related to RyR cluster distances and sizes. The data encompass a wide range of cellular morphologies and RyR cluster distributions. Notably, cells 3, 8, 14, 24, and 27 (in Figure 3) are included in this comprehensive dataset, allowing for direct comparison of their characteristics with the broader cell population.
On average, we detected 2285 clusters per cell, and the overall mean of RyR cluster sizes was 0.18 μm3 with a standard deviation of 0.13 μm3. The distributions were plotted as histograms with 50 bins, and the existence of two right-side-skewed subpopulations was visually apparent, with four representative cells shown in Figure 4. Thus, a Gamma Mixture Model (GMM) was used to model this distribution. A Gamma distribution is often used to model distributions that are asymmetric, with a longer tail on the right side. A GMM is a probabilistic model that assumes the data are generated from a mixture of several Gamma distributions, in our case two distributions. Each component in the mixture is a Gamma distribution with its own parameters, and the mixture is weighted by coefficients that sum to 1. The PDF for a GMM with two components in our case is:
where is the Gamma function, π is the mixture weight, and are the shape and scale parameters for their respective Gamma distribution components. The GMM was fit to the data using the scikit-learn library in Python 3.14, which implements the expectation maximization algorithm to estimate the parameters of the mixture model. The number of Gamma components in the mixture was determined by evaluating models with different numbers of components and selecting the one with the lowest Bayesian Information Criterion to balance goodness of fit with model complexity.
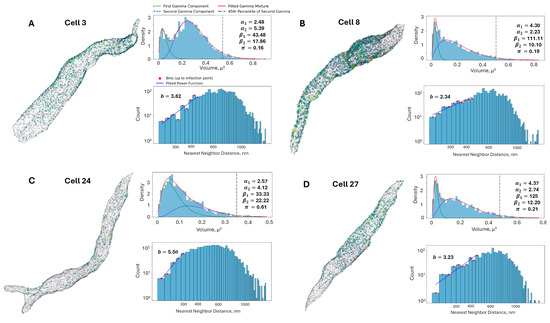
Figure 4.
Analysis of RyR cluster distribution and nearest-neighbor distances in representative rabbit SANC. (A–D): Four representative cells (3, 8, 24, and 27 in Figure 3), each presenting three key analyses. First, a 3D visualization depicts RyR clusters as colored spheres scaled to their volumes (yellow for largest, purple for smallest) within a gray mesh representing the cell surface. Second, a fitted Gamma Mixture Model for RyR cluster size distributions is shown, with blue histograms representing observed data, green and blue dashed lines indicating individual gamma components, and a red line showing the overall fitted mixture. Model parameters (α1, α2, β1, β2, π) and the 95th percentile of the second gamma component (vertical black dash-dot line, marking large CRUs) are displayed. Third, a log-log histogram of RyR cluster nearest neighbor distances is presented, where blue bars show observed distances, red dots mark bins up to the inflection point used for power law fitting (blue line), and the fitted power law exponent (B) is provided. This comprehensive analysis reveals cell-to-cell variability in RyR cluster organization and spacing but overall consistency, offering insights into the spatial arrangement of Ca2+ release sites in SANC.
To initialize the GMM, an estimate of the peak locations in the data was obtained using the method of moments for initial parameter estimation, which splits the sorted data at the median and uses the statistics of each half to approximate the parameters of two gamma distributions. The two most prominent peaks were identified and used as initial estimates for the means of the Gamma components. The optimization of the GMM parameters was performed using the minimize function from the scipy.optimize module, with bounds set on the parameters to ensure they remain positive and the mixing coefficient remaining between 0 and 1. The GMM analysis revealed that the distribution of RyR cluster sizes in the rabbit sinoatrial node cells is best described by a mixture of two Gamma components, with the results for individual cells shown in Table 2.
Table 2.
RyR cluster distribution analysis in rabbit SANCs. Detailed analytical results for the 31 rabbit SANCs, focusing on key aspects of RyR cluster distribution: repulsion (b) and Gamma Mixture Model (GMM) parameters (α1, β1, α2, β2, π). The repulsion parameter b indicates the strength of spatial repulsion between RyR clusters. The GMM parameters describe the distribution of RyR cluster sizes. Notably, cells 3, 8, 14, 24, and 27 in previous figures are included in this dataset, allowing for a more comprehensive understanding of their RyR cluster characteristics in the context of the entire cell population. The aggregated results refer to the results that are calculated from all RyR clusters in all 31 cells in a single plot.
After application of Tukey’s Fences on the sizes, the RyR clusters were aggregated into a set of 70,849 clusters. Next, a histogram with 100 bins was created and a GMM was fit on top of this aggregated set. The fitted parameters of the GMM were as follows in Figure 5: = 4.35, = 125, = 2.22, = 10.64, π = 0.16. The first gamma component had an average of 0.0333 μm3 and a standard deviation of 0.0160 μm3, while the second gamma component had an average of 0.2083 μm3 and a standard deviation of 0.1398 μm3. These results provide a comprehensive characterization of the RyR cluster size distribution in the rabbit SANC, highlighting the presence of two distinct subpopulations with different average sizes and variability. The kurtosis values of the first and second Gamma components were calculated to be 1.38 and 2.70, respectively. The higher kurtosis of the second component suggests that it represents a subpopulation of larger RyR clusters with a heavier tail in the distribution. The 95th percentile of the second Gamma component was found to be 0.478 μm3, which was used as a threshold to define the tail of the distribution representing the largest RyR clusters used in further analysis.
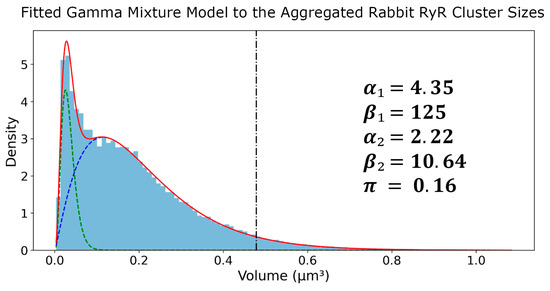
Figure 5.
Gamma mixture model analysis of RyR cluster sizes in rabbit SANCs. Probability density function (PDF) of RyR cluster sizes derived from 31 rabbit SANCs, encompassing 70,982 clusters. The distribution is fitted with a two-component Gamma Mixture Model, with parameters (α1, β1, α2, β2, π) displayed. The blue histogram represents experimental data, while green and blue dashed lines show individual gamma components. The red line depicts the overall fitted mixture, and the vertical black dash–dot line marks the 95th percentile of the second gamma component (marking large CRUs).
3.2. Repulsive Behavior of RyR Clusters in Spatial Distribution
To investigate the spatial arrangement of RyR clusters, the aggregated distances between nearest-neighbor clusters were analyzed. The distribution of nearest-neighbor distances provides insights into potential spatial interactions, such as repulsion or attraction, among the clusters. The overall mean distance between nearest-neighbor RyR clusters was found to be 638.62 nm, with a standard deviation of 208.14 nm (Table 1). Histograms of the nearest-neighbor distances for each individual SANC were constructed using 50 bins and a power law function which was fit to the ascending part of the distribution up to the inflection point, determined as the bin where the histogram values first fell below half of the peak value.
The power law function used to model the nearest-neighbor distances is given by , where is the probability of observing a nearest-neighbor distance, is a normalization constant, and b is the power law exponent. Transforming the dataset into a log-log plot, a linear relationship is obtained between ) and This transformation allows for a more straightforward analysis of the power law behavior. The resulting equation in log-log form is: . The variable b characterizes the spatial distribution of the clusters, with b ≈ 1 indicating a random Poisson process, b > 1 suggesting repulsion, and b < 1 indicating clustering. The power law function is fit to the ascending part of the histogram until the inflection. Four example cells and their associated analysis are shown in Figure 4. The results in Figure 6B and Table 2 show that each cell exhibited repulsion with b > 1. Figure 7 is a visual representation of the spatial distribution of RyR clusters, showing varying levels of cluster density on the surface mesh on sample cell 14. Higher-density areas are colored in blue → green → yellow, representing RyR cluster “hotspot zones”, while low-density areas are in purple, exhibiting strong repulsive forces.
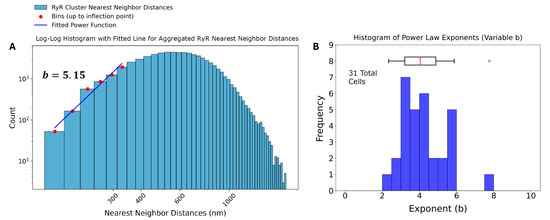
Figure 6.
Analysis of RyR cluster nearest-neighbor distances in rabbit sinoatrial node cells. (A) Log-log histogram of nearest-neighbor distances (NND) for ryanodine receptor (RyR) clusters. The blue bars represent the observed NND distribution. Red dots indicate bins up to the inflection point, which were used to fit a power function (blue line). The fitted power law exponent b = 3.81 is displayed, suggesting strong repulsion among RyR clusters at short distances. (B) Histogram of power law exponents (b) derived from individual analysis of 31 cells, with an accompanying box plot.
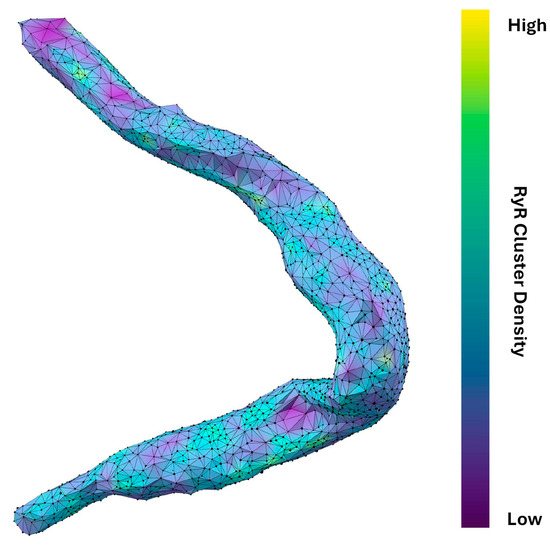
Figure 7.
Spatial distribution and connectivity analysis of RyR clusters in a rabbit SANC. Detailed visualization of RyR cluster distribution in cell #14 (in Figure 3). The cell’s shape is outlined by a network of interconnected black vertices, each representing an RyR cluster. The surface coloring, transitioning from purple to yellow (via blue and green), indicates the density and connectivity of these clusters. Purple areas signify regions where clusters strongly repel each other, resulting in lower cluster density and fewer neighboring connections. Conversely, blue → green → yellow areas represent regions where this repulsion is weaker, leading to higher-density regions with more interconnected clusters. The varying degrees of repulsion between RyR clusters across different areas of the cell provide insights into the spatial organization of Ca2+ release sites, which influence CICR and AP firing rate. Specifically, this visualization technique effectively highlights potential hotspots of Ca2+ signaling activity within the cell that is likely associated with higher density of RyR clusters.
After application of Tukey’s Fences on the distances, the aggregated set of 70,887 nearest neighbor distances was also analyzed, and the results of the analysis revealed a power law exponent of b = 5.15, also indicating a strong repulsion among the RyR clusters at short distances (Figure 6A).
3.3. Structure–Function Relationship of CRU Network in Numerical Simulations of SANC Function
We simulated AP firing in numerical SANC models (Figure A1) with different distributions of sizes and locations. With respect to sizes, we tested and compared two distributions: one was the realistic (i.e., heterogeneous) distribution obtained in SIM data adopted to the SANC model (re-scaled and re-binned) and the other was the distribution featuring identical CRU sizes, with each CRU size fixed to 48 RyRs representing the average CRU size in the realistic distribution. It is also important to note that each cell model had the same total number of RyRs. With respect to CRU locations, we tested three CRU distributions of NNDs: (i) uniformly random, (ii) realistic, and (iii) crystal-like grid (Figure A2). The distributions were generated by our CRU repulsion algorithm that dynamically changed positions of CRUs from uniformly random locations with no repulsion towards a crystal-like structure with the strongest repulsion. The change was performed in 100 repulsion steps (see details in Appendix A and Videos S1 and S2). The realistic NND distribution was found at an intermediate repulsion step with the repulsion, yielding an average NND value closely matching that measured experimentally.
Finally, each model was tested in basal-state AP firing and in the presence of β-adrenergic receptor (βAR) stimulation. Thus, we tested the operation of six different SANC models under two conditions, resulting in 12 simulations altogether. The results of these simulations are summarized in Figure 8. The simulations (see examples in Videos S3–S8) showed that, for each CRU location distribution type (random, realistic, crystal), the models with realistic CRU sizes had faster AP firing rates vs. those with identical CRU sizes. At the same time, the absolute values of AP firing rates were the largest (reflecting shortest cycle length) in cells with random CRU locations (Figure 8A), but the smallest (reflecting longest cycle length) in crystal-like grids with the strongest CRU repulsion (Figure 8C). The scenario with realistic locations (Figure 8B) had an intermediate AP firing rate of 157 bpm, close to the experimentally reported rates of isolated rabbit SANCs.
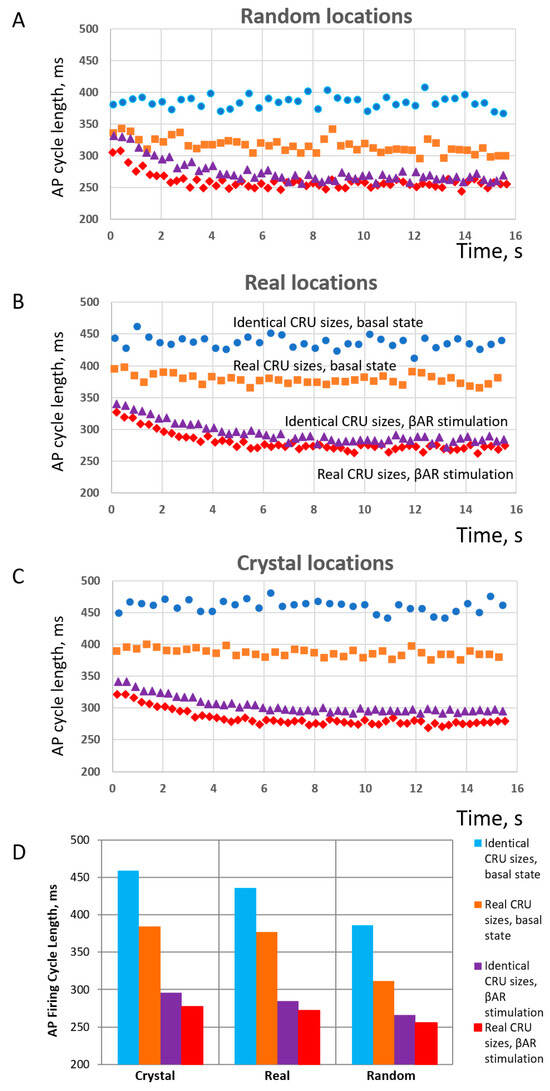
Figure 8.
Results of numerical model simulations of SANC function: heterogeneities in both CRU sizes and locations facilitate LCR propagation and increase AP firing rate in a cooperative manner but decrease the effect of βAR stimulation in terms of relative change in AP firing rate; at the same time, the presence of heterogeneities in both sizes and locations allows higher absolute AP firing rates to be reached during βAR stimulation. (A–C): Intervalograms for AP cycle length (CL) in 12 numerical model simulations during 16 s for 6 models featuring different distributions of CRU sizes and distances in basal state and during βAR stimulation (shown by text labels in the panel (B)). (D): Graph showing average CL calculated for steady-state AP firing during 6 to 16 s of the simulations (see also Videos S3–S8). Explanation for symbols: blue circles-identical CRU sizes in basal state; orange squares–real CRU sizes in basal state; purple triangles–identical CRU sizes in βAR stimulation; red diamonds–real CRU sizes in βAR stimulation.
With respect to βAR stimulation, its effect in terms of relative change of AP firing CL (or rate) increased as the order of CRU sizes and position increased, i.e., heterogeneity or disorder decreased and reached its maximum in the model with identical CRU located in a crystal-like grid with the least heterogeneity and maximum order. See panel D, with blue and magenta bars in the “Crystal section”, 459 vs. 296 ms, yielding a 55% increase in AP firing rate from 131 vs. 202 bpm. And vice versa, the βAR stimulation effect decreased as the disorder of CRU sizes and position increased (i.e., order decreased) and reached its minimum in the model with realistic CRU located in (uniformly) random locations. See Figure 8D, with orange and red bars in the “Random” section, 311 vs. 255 ms, yielding only a 22% increase in AP firing rate from 192 vs. 225 bpm. On the other hand, the disorder (heterogeneity) of CRU sizes and locations allowed the models to reach higher absolute values of AP firing rates (shorter cycle lengths) both in the basal state and during βAR stimulation. As a result, the highest beating rate of 225 bpm among all 12 tested scenarios was achieved in the SANC model with the highest degree of disorder, representing a cooperative effect of the most heterogeneous (realistic) sizes and uniformly random locations of their CRUs (red diamonds in panel C). The model with realistic CRU sizes and locations demonstrated a 39% βAR increase in AP firing rate (377 vs. 271 ms of CL, or 159 vs. 220 bpm).
Next, we analyzed our simulations to obtain insights into the biophysical mechanism of the contribution of the heterogenous CRU network to regulating pacemaker function. According to our previous studies, the critical part of the diastolic depolarization is the AP ignition [16] at the threshold of L-type Ca2+ current (ICaL) activation. In our model, each CRU size is given by the number of RyRs embedded in that CRU. Therefore, in all 12 simulation scenarios, we determined the total number of RyRs in all firing CRUs at −45 mV, i.e., at the threshold of ICaL activation (Cav1.3 current component in our model of ICaL). We found that the AP cycle length in the basal state and during βAR stimulation was linked to the number of firing RyRs (Figure 9), reflecting the degree of order and disorder (heterogeneity) of CRUs and their ability to self-organize to fire earlier and synchronously within the AP cycle.
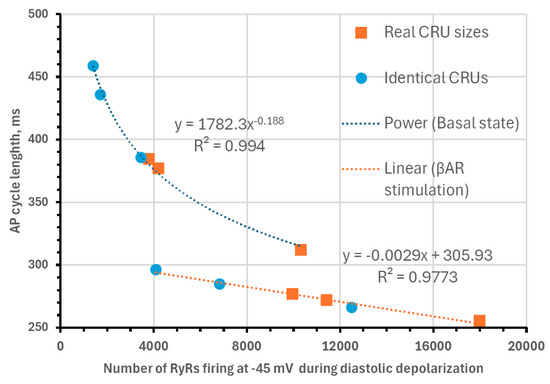
Figure 9.
Biophysical mechanism of how CRU distribution affects AP firing rate. Shown is the relationship between of AP cycle length (CL) and the total number of RyRs in all firing CRUs at −45 mV, i.e., at the threshold of ICaL activation. Both in basal-state firing (fitted by power function, R2 = 0.994) and during βAR stimulation (fitted by linear function, R2 = 0.9773), CL was closely linked to the RyR firing at this critical timing within diastolic depolarization.
4. Discussion
4.1. Result Summary: Major Findings
The present study, for the first time, examined the detailed structure of the RyR network under the cell membrane in SANCs (Figure 3) at the nanoscale level via SIM super-resolution microscopy. Previous lower-resolution confocal microscopy studies have shown that RyR appears in clusters which are mainly located under the cell membrane in central (primary) SANCs [17]. It was also shown that RyR-mediated LCR events play a crucial role in the coupled-clock system that ensures robust and flexible pacemaking [7]. Many important properties of the network, however, remained unknown. The major findings of the present study include:
- (1)
- Two distinct RyR cluster subpopulations with sizes adhering to a Gamma mixture distribution.
- (2)
- Notable repulsion between neighboring RyR clusters.
- (3)
- Model simulations showed that heterogeneities in both CRU sizes and locations facilitate CICR and increase AP firing rate in a cooperative manner, but decrease the effects of βAR stimulation. However, the highest possible absolute rate during βAR stimulation is reached with heterogeneities in both CRU sizes and locations.
4.2. New Image Analysis Methods and Numerical Models
We developed a novel, high-throughput algorithm for the segmentation of peripheral RyR clusters in 3D in SANCs, enabling the analysis of over 70,000 RyR clusters across 31 rabbit SANCs (Figure 1). Our advanced image preprocessing techniques included 3D median filtering, signal normalization, and CLAHE [30]. It enhanced the local contrast while preserving the global image structure, facilitating more accurate segmentation. For the first time in SANCs, the 3D StarDist neural network [31] was trained for the segmentation of RyR clusters and accurately predicted star-convex polyhedra representations for each cluster, improving upon traditional methods that rely on watershed algorithms and often struggle with overlap. The ground truth data for training the neural network were prepared using Squassh [32] and fine-tuned with the Adaptive Watershed tool in ORS Dragonfly [36]. Further refinement through DBSCAN clustering removed false positives. To accurately capture the peripheral RyR clusters directly interacting with cell membrane ion channels and transporters critical for heart rate regulation, the cell periphery was isolated using an alpha shape algorithm, constructing a minimal bounding surface that closely conforms to the shape of the data points, effectively representing the cell’s geometry.
We also modified our previous 3D numerical model of SANC [22] by including new formulations for CRU function (see Appendix A for details). Recent theoretical studies [37,38,39] showed that Ca2+ release activation in a CRU (i.e., Ca2+ spark activation) depends substantially on the CRU size, i.e., the number of RyRs in a CRU. Thus, our new formulations were created to reflect actual distributions obtained by segmentation of SIM images: they generate repulsion between the CRUs (Figure A2, Videos S1 and S2) and CRUs of various sizes (Figure A3). Using the new formulations, we created and tested a series of models with real and extreme settings that revealed importance of heterogeneities in CRU sizes and locations and their joint regulation of CICR to ensure robust and flexible pacemaker function.
4.3. Structure–Function Relations of RyR Network in SANC
While it is known that LCRs play an important role in pacemaker function [7], the detailed mechanisms regarding how LCRs emerge from the RyR network remain mainly unknown. Because the RyR network operates via CICR, its properties must critically depend on both CRU sizes (generating stronger releases) and distances, which define whether the release of each CRU can reach and activate its neighbors (i.e., “when can a Ca2+ spark jump?” [21]). Our previous theoretical studies of CRU function in SANCs first showed that synchronization of stochastic CRUs creates a rhythmic Ca2+ clock in SANCs via repetitive phase-like transitions within a perfect rectangular grid of CRU networks [19]. Further linking this model with the membrane clock revealed that RyR-Na/Ca exchanger (NCX)-SERCA local crosstalk ensures pacemaker cell function at rest and during the fight-or-flight reflex [20].
While previous studies have tested the effects of CRU sizes and distances in separation, here, for the first time, we found that they act together to regulate pacemaking (Figure 8). The CRUs form a functional network in which Ca2+ release events, which self-organize (via CICR) into local Ca2+ oscillators that create a rhythmic diastolic Ca2+ signal known as the “Ca clock”. This, together with the “membrane clock”, forms a coupled-clock system that ensures robust and flexible pacemaking [7]. The diastolic Ca2+ signal acts not alone, but within a strong positive feedback mechanism (known as AP ignition [16]) with NCX, membrane potential, and ICaL. The self-organized Ca2+ signal formed by CRUs at the threshold of ICaL (at the ignition point) must play a critical role in the ignition and, ultimately, AP firing rate regulation.
Our simulations showed that this critical signal at the threshold of ICaL is tightly linked to the structure of the RyR network. The heterogeneity in both CRU sizes and distances strongly increased the CICR and the AP firing rate both in the basal state and during βAR stimulation (Figure 9). Thus, to reach the maximal relative increase in firing rate and, at the same time, the maximum absolute rate, the CRU network can transform to increase its heterogeneity, i.e., to increase the contribution of larger RyR clusters. Recent dSTORM studies in ventricular myocytes showed that the RyR clusters can expand and coalesce after application of isoproterenol [40]. Future dSTORM studies will show whether this also happens in SANCs, allowing the RyR network to transform itself to reach maximum performance in fight-or-flight reflex.
We have previously shown [22] that disorder in CRU locations increases CICR due to Poisson clumping characterized by empty spaces and clusters. The CRU clustering makes it easier for released Ca2+ to reach a neighboring CRU. But why does a CRU network with heterogeneous CRU sizes also enhance CICR and AP firing rate? Larger CRUs are naturally present in such network (Figure 4 and Figure 5, beyond the vertical line). These larger CRUs have a greater probability and capacity for Ca2+ release [39], acting as central hubs that initiate and amplify CICR to the surrounding smaller CRUs. This multi-scale spatial organization enhances the likelihood of CICR propagation from large clusters to smaller ones, promoting synchronization necessary for effective Ca2+ signaling.
Pilot dSTORM studies of RyR clusters in rat SANCs (abstracts [41,42]) fully support the results of the present study. It was found that the cluster sizes vary from 10 to 150 RyRs, and they are organized in couplons, i.e., in apposition to L-type Ca2+ channels found largely at the outer membrane in SAN cells. Heterogenous RyR2 clusters were also found, often with small inter-cluster distances, which is indicative of inter-cluster RyR2 activation.
4.4. Limitations and Future Studies
Our cell database was limited to 31 cells, and our model simulations were limited to 12 basic scenarios. Future studies in a larger number of cells and with even stronger imaging resolution, like dSTORM (up to 20 nm resolution [43]), will clarify further details of RyR network structure and its relationship with other networks formed, e.g., by mitochondria [44] or Ca2+ channels of different types and isoforms [45,46,47] directly or indirectly interacting with the RyR network.
Future numerical modeling will include and test the functional importance of these relationships. The presence of spatial repulsion between CRUs suggests that there exists a molecular mechanism that actively maintains minimum spacing. Further biophysical studies will also clarify the nature of CRU repulsion and, possibly, coalescence [40]. Thus, the CRU sizes are likely determined, in part, by the repulsion/coalescence balance controlled by autonomic modulation to reach the maximal (or optimal) effect.
The antibody mAb C33-3 used to detect RyRs in SANCs was generated against RyR2, but also cross-reacted with RyR1. However, using RT-PCR, it was reported that the sinoatrial node contained both RyR2 and RyR3, but not RyR1 [48]. The relative abundance of RyR2 and RyR3 was distinct between cells located at the periphery vs. those at the center of the sinoatrial node: RyR2 was higher at the periphery and RyR3 was higher in cells at the center of the node [49]. Thus, the present study did not investigate all RyRs, but was focused on cardiac type RyR2. Further studies are needed to clarify CRU composition and function with respect to both RyR2 and RyR3 isoforms in SANCs residing in different regions of the node.
4.5. Implications for Clinical and Aging Research
Disorder within biological systems tends to increase with aging across all scales. Thus, heterogeneity (i.e., disorder) in CRU sizes and spatial distribution is also expected to increase in older people, but as a result, CICR will actually be enhanced, thereby elevating the AP rate. This enhancement could partially compensate for the reduced intrinsic heart rate commonly associated with aging. However, the same increase in structural disorder of the RyR network would diminish β-adrenergic responsiveness, as demonstrated in this study, contributing to (and explaining in part) the decline in heart rate reserve observed in older people.
As we mentioned in the Introduction, dysfunction of SANCs can lead to sick sinus syndrome, which is associated with a variety of life-threatening arrhythmias [3,4,5,6]. Electronic pacemakers are used to treat this condition, but impose lifestyle restriction and can cause severe side effects. To improve electronic pacemakers and create new approaches, for example, biological pacemakers [50], it is important to understand the natural origin of the heartbeat, which is “still mysterious after all these years” (cited from [23]). SANC operates via a coupled-clock mechanism [7], and its “Ca2+ clock” operates via synchronous and rhythmic release of Ca2+ via the RyR cluster network. Our new findings regarding the structure–function relation of the network can help direct the development of new prophylactic and therapeutic strategies. Specifically, our results suggest that healthy operation of the network requires a delicate balance of order and disorder in RyR cluster positions and sizes that determines and ensures the respective healthy balance of robustness and flexibility of cardiac pacemaking. Dysfunction of this balance could be a new factor in heart rate reserve decline in aging and disease. Our numerical model provides a new platform to investigate how molecular defects in CRUs and CRU network structure may disrupt pacemaking in diseases like sick sinus syndrome. Incorporating measured CRU network properties will help to provide mechanistic insights into pacemaker pathologies and develop new biological pacemakers.
4.6. Conclusions
CICR interplay of CRUs of various sizes and locations regulates and optimizes cardiac pacemaker cell operation under various physiological conditions and could be a key factor in heart rate reserve decline in aging and disease. Overall, our study elucidates how the specialized architecture of the CRU network in SANC regulates ignition and propagation of Ca2+ sparks to generate rhythmic AP firing. The combination of super-resolution imaging and computational modeling provides a powerful approach to link subcellular Ca2+ release patterns to whole-cell pacemaker activity.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cells13221885/s1, Videos S1 and S2: The sequence of CRU distributions (for real CRU sizes and identical CRUs, respectively) generated by our CRU repulsion algorithm that dynamically changed positions of CRUs from uniformly random locations with no repulsion towards a crystal-like structure with the strongest repulsion (cylinder cell surfaces were unwrapped to squares). The change was performed in 100 repulsion steps. The CRUs (green objects) on the top panel are shown together with respective histograms of NNDs and plots of average NND and standard deviation at each step. For a better representation of nonlinear changes, the first 13 steps are shown at a slower rate of 0.5 s/frame, with the rest of the steps at 0.1 s/frame. Videos S3–S8: Ca2+ dynamics under the cell membrane in numerical simulations in the basal state of our 6 models with different CRU distributions as follows: Video S3, random positions and realistic sizes; Video S4, realistic positions and sizes; Video S5, crystal-like positions and realistic sizes; Video S6, random positions and identical sizes (48 RyRs); Video S7, realistic positions and identical sizes, Video S8, crystal-like positions and identical sizes. Ca2+ concentration is coded by red shades from black (0.15 μM) to pure red (>10 μM). CRU functional states are coded by colors as follows: refractory CRUs are in blue; CRUs ready to release are in green; and CRUs releasing Ca2+ are in grey shades, reflecting JSR Ca2+ changes, with a saturation level set at 0.3 mM. Simulation time and membrane potential are shown in the left top corner of each video.
Author Contributions
Conceptualization, A.V.M. (Alexander V. Maltsev), E.G.L., and V.A.M.; methodology, A.V.M. (Alexander V. Maltsev), V.V.S., O.M., M.J., S.T. A.V.M. (Anna V. Maltsev), and V.A.M.; software, A.V.M. (Alexander V. Maltsev), V.V.S., S.R., and V.A.M.; validation, A.V.M. (Anna V. Maltsev), S.T., M.D.S., and V.A.M.; investigation, A.V.M. (Alexander V. Maltsev), V.V.S., O.M., M.J., P.A.W., S.R., A.V.M. (Anna V. Maltsev), and V.A.M.; resources, E.G.L. and M.D.S.; data curation, A.V.M. (Alexander V. Maltsev), V.V.S., and V.A.M.; formal analysis, A.V.M. (Alexander V. Maltsev), V.V.S., O.M., P.A.W., S.R., and V.A.M.; writing—original draft preparation, A.V.M. (Alexander V. Maltsev) and V.A.M.; writing—review and editing, A.V.M. (Alexander V. Maltsev), V.V.S., A.V.M. (Anna V. Maltsev), E.G.L., and V.A.M.; visualization, A.V.M. (Alexander V. Maltsev), V.V.S., and V.A.M.; supervision, M.D.S., E.G.L., and V.A.M.; project administration, E.G.L. and M.D.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Intramural Research Program of the National Institutes of Health, National Institute on Aging. Anna V. Maltsev acknowledges the support of the Royal Society University Research Fellowship (Grant number RF/ERE/21005). Valeria Ventura Subirachs acknowledges the support of the U-RISE Program at the University of Maryland, Baltimore County (UMBC), which is supported by the National Institute of General Medical Sciences of the National Institutes of Health under Award Number T34GM136497.
Institutional Review Board Statement
The animal study was reviewed and approved by the Animal Care and Use Committee of the National Institutes of Health (protocol #457-LCS-2024).
Informed Consent Statement
Not applicable.
Data Availability Statement
All original data files were deposited to Harvard Dataverse at https://doi.org/10.7910/DVN/N0OLGI (accessed 13 November 2024). Code Availability: Code for image data analysis was uploaded to GitHub: https://github.com/alexmaltsev/SANC (accessed 13 November 2024), https://github.com/valventura/SANC (accessed 13 November 2024). Our new model of rabbit SANC with CRUs of various sizes written in Delphi programming language was uploaded to GitHub: https://github.com/victoramaltsev/CRU-based-SANC-model (accessed 13 November 2024).
Acknowledgments
The authors thank Hari Shroff (currently at Janelia Research Campus) for the opportunity to perform SIM imaging in his research laboratory at the National Institute of Biomedical Imaging and Bioengineering, NIH. The authors wish to acknowledge the assistance of Bruce D. Ziman for skillful isolation of sinoatrial node cells. The authors also thank Daniel Riordon and Larissa Maltseva for their assistance in performing SIM imaging.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
| SANC = sinoatrial node cell; |
| AP = action potential; |
| LCR = local Ca2+ release; |
| βAR = β-adrenergic receptor; |
| RyR = ryanodine receptor; |
| SERCA= sarco/endoplasmic reticulum Ca-ATPase; |
| Cycle length = action potential cycle length; |
| CRU = Ca2+ release unit; |
| SR = sarcoplasmic reticulum; |
| JSR = the junctional SR; |
| CICR = Ca-induced Ca release; |
| SIM = structured illumination microscopy; |
| NND = nearest-neighbor distances; |
| PBS = phosphate-buffered saline; |
| CLAHE = contrast-limited adaptive histogram equalization; |
| IoU = Intersection over Union; |
| DBSCAN = Density-Based Spatial Clustering of Applications with Noise; |
| MAD = median absolute deviation; |
| GMM = Gamma Mixture Model; |
| CDF = cumulative distribution function. |
Appendix A. Details of a Novel Numerical Model of SANC
In the present study, we investigate functional importance of both heterogeneous CRU sizes and locations (Figure A1). To gain further insights into the operation of the CRU network featuring realistic locations and sizes (as observed in SIM images), we developed a new numerical cell model that has a novel Ca2+ release mechanism that depends on the CRU size. The new model is based on our previous model of rabbit SANCs [22], which included a full set of electrophysiological equations and local Ca2+ dynamics generating LCRs via CRUs, but it was limited to CRUs of identical sizes. With respect to CRU location, the new model simulates repulsion between CRUs (see Videos S1 and S2), which was revealed in SIM images. The LCR emergence is approximated in the model at the scale of an individual CRU, i.e., LCRs appear as Ca2+ sparks and abrupted Ca2+ waves (wavelets, as observed in prior experimental studies) via saltatory propagation of CICR among neighboring CRUs. Each CRU, in turn, is represented by a cluster of RyRs embedded in the JSR, located in close proximity (20 nm) to the cell surface membrane. We did not simulate operation of each individual RyR, but (in the spirit of multi-scale modeling [51]) we tuned the CRU formulations, i.e., clusters of RyRs, by performing simulations of the Stern model of CRU [52] featuring individual RyR firing. All equations of this new numerical model (and model tuning), including novel formulation of CRU activation and generation of realistic heterogeneous CRU distributions (both size and location), are described below.
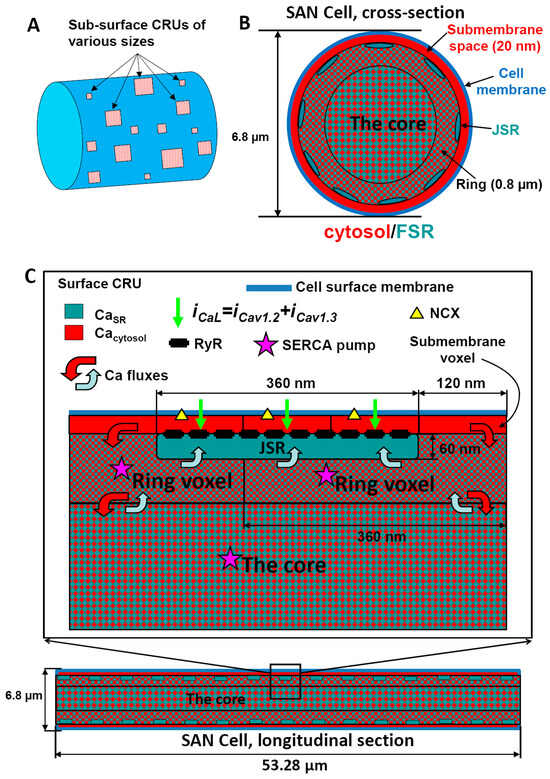
Figure A1.
Schematic illustration of approximation of local Ca2+ dynamics in our CRU-based SANC model. Panel (A) shows the spatial positioning of Ca2+ release units (CRUs) beneath the cell membrane. CRUs are shown in different sizes (a key feature of our new model), reflecting different numbers of ryanodine receptors (RyRs) included in each CRU. In Panels (B,C), a cross-sectional and a longitudinal section of our simulation are presented, respectively, revealing a three-layer voxel-based approximation of intracellular Ca2+ dynamics. These voxel layers offer a discretized 3D view of the cellular space, enabling the detailed spatio-temporal representation of Ca2+ signals within the cell. Specifically, the first layer (red) approximates the submembrane space where the CRUs are located and interact with cell membrane proteins. The second layer (ring) approximates cell space associated with the junctional sarcoplasmic reticulum (JSR), and the third layer (the core) is representative of the bulk cytosolic space. Both the ring and the core include the network sarcoplasmic reticulum, the primary intracellular Ca2+ storage site that is equipped with Ca2+ pump (SERCA, labeled by star). Collectively, this updated 3D SANC model, with its refined representation of CRU sizes and locations, provides a more accurate and realistic approximation of Ca2+ signaling in sinoatrial node cells. Adopted from [22].
Figure A1.
Schematic illustration of approximation of local Ca2+ dynamics in our CRU-based SANC model. Panel (A) shows the spatial positioning of Ca2+ release units (CRUs) beneath the cell membrane. CRUs are shown in different sizes (a key feature of our new model), reflecting different numbers of ryanodine receptors (RyRs) included in each CRU. In Panels (B,C), a cross-sectional and a longitudinal section of our simulation are presented, respectively, revealing a three-layer voxel-based approximation of intracellular Ca2+ dynamics. These voxel layers offer a discretized 3D view of the cellular space, enabling the detailed spatio-temporal representation of Ca2+ signals within the cell. Specifically, the first layer (red) approximates the submembrane space where the CRUs are located and interact with cell membrane proteins. The second layer (ring) approximates cell space associated with the junctional sarcoplasmic reticulum (JSR), and the third layer (the core) is representative of the bulk cytosolic space. Both the ring and the core include the network sarcoplasmic reticulum, the primary intracellular Ca2+ storage site that is equipped with Ca2+ pump (SERCA, labeled by star). Collectively, this updated 3D SANC model, with its refined representation of CRU sizes and locations, provides a more accurate and realistic approximation of Ca2+ signaling in sinoatrial node cells. Adopted from [22].
Appendix A.1. Cell Geometry, Compartments, Voxels, and Membrane Patches
We model a small SANC shaped as a cylinder 53.28 μm in length and 6.876 μm in diameter. The cell membrane electrical capacitance of 19.8 pF is similar to that of 20 pF in Zhang et al.’s model [53] of a central SANC. Details of local Ca2+ dynamics under the cell membrane are simulated on a sub-micron-resolution square grid (120 nm, i.e., similar to SIM resolution used in this study). The grid divides the cell membrane and the submembrane space (dubbed subspace) into respective membrane patches and subspace voxels. Locations within the grid are defined by coordinates in the respective plane of the cell surface cylinder: along the cylinder (x axis) and around the cell cross-section (y axis). Our cell partitioning and respective voxel sizes are schematically illustrated in Figure A1. To avoid special considerations at the cell borders, the ends of the cylinder are connected (yielding a torus). Our cell compartments and voxel structure are essentially similar to those in Stern et al.’s model [21]. which approximates Ca2+ dynamics in three dimensions at the single RyR scale. However, we limited the cell partition to only three nested layers of voxels of substantially different scales, reflecting the respective essential Ca2+ cycling components and processes happening at these scales (described below).
Appendix A.1.1. Submembrane Voxels
The first, most detailed level of approximation of local Ca2+ dynamics is achieved via 79,920 voxels, 444 in x and 180 in y. Each voxel of 120 nm × 120 nm × 20 nm (ΔxΔyΔr) is located under the entire cell membrane, including the 20 nm dyadic space (or cleft space) separating CRUs and the cell membrane. This thin layer of voxels describes local Ca2+ release from individual CRUs, CRU-to-CRU interactions via Ca2+ diffusion, and CRU interactions with the cell membrane (including ICaL, INCX, ICaT, and IbCa). Ca2+ currents were computed for each membrane patch (120 nm × 120 nm) to generate local Ca2+ fluxes, contributing to local Ca2+ dynamics.
Appendix A.1.2. JSR Level Voxels (“Ring” Voxels)
The next approximation level of Ca2+ dynamics is a deeper layer of voxels with a larger size of 360 nm × 360 nm × 800 nm (ΔxΔyΔr) and that includes the scale of JSR depth (60 nm). Each ring voxel has its cytoplasmic part and FSR part, and some ring voxels are diffusively connected to JSRs (Figure A1). While it appears that the geometric scale of the ring voxel is of an order of magnitude larger than the JSR size, the actual FSR volume within each ring voxel is comparable with the JSR volume (described below in details). Thus, this level of voxels describes the local dynamics of Ca2+ transfer from FSR to JSR, as well as enhanced local Ca2+ pumping and diffusion fluxes due to close proximity to Ca2+ release and Ca2+ influx in its neighboring submembrane voxels.
Appendix A.1.3. The Core
The rest of the cell does not have further geometric partitioning, and it is lumped into one compartment, “the core”, where local Ca2+ dynamics are less important. The core also has its cytosolic and FSR parts (as in ring voxels). Thus, it describes the bulk Ca2+ uptake from cytosol to FSR and further accumulation, diffusion, and redistribution of the pumped and released Ca2+ within the cell interior.
Appendix A.1.4. JSR
While individual release channels are not modeled directly, each CRU is characterized by its number of RyR channels, determining its JSR size. Because the thickness of pancake-like JSR is fixed to 60 nm, the volume of each JSR is directly proportional to the number of RyRs (NRyR) residing in the JSR and facing the dyadic space. Because RyRs form a 30 nm × 30 nm crystal grid and because the dyadic space (as a part of the submembrane space) is split into voxels of 120 nm × 120 nm in the xy plane, each xy voxel size can accommodate 120 × 120/(30 × 30) = 16 RyRs. This dictates the possible NRyR and respective geometries that JSRs can have: NRyR varies in multiples of 16, i.e., NRyR = 16*n, whereas n is positive integer, i.e., 16, 32, 48, 64, etc. Thus, each CRU is characterized by a set of adjacent submembrane voxels fully populated by RyRs. Our computer algorithm configured CRU geometries as ideal squares of the voxels when possible (e.g., 2 × 2 for 4 voxels populated by 64 RyRs or 3 × 3 for 9 voxels populated by 144 RyRs), or formed incomplete squares for intermediate cases with random rotation.
Based on available experimental data, our previous SANC model [21] had 196,000 RyRs. That cell model had a length of 112 μm and a diameter of 8 μm, yielding a cell cylinder surface area of π × 8 × 112 = 2814.86701762 μm2, with an RyR density of 196,000/2814.86701762 = 69.6302876026 ~70 RyRs per μm2. A similar RyR density was also used in our original CRU-based model of SANC [22] and in the present model. We examine two types of JSR size models here: (1) all JSRs have identical number of RyRs, i.e., homogeneous JSR models; (2) JSR sizes are distributed as in the experimental data. For a correct comparison of structure–function performance, we generate all models with approximately the same number (83,009) of RyRs per cell, as determined by the aforementioned density of 70 RyRs per μm2 and total cell surface area in our cell model of 2 × π × 3.495043 × 54 = 1185.84015258 μm2. The exact number of RyRs per cell slightly varies in different RyR distributions because (i) RyRs populate JSRs in multiples of 16, as described above, and (ii) there is a small remainder when CRUs are populated following a specific RyR distribution. When we created CRUs and populated them with RyRs in the cell model using experimental data, we first calculated the number of RyRs for CRU size (16, 32, 48, etc.) by simple scaling: ModelCRUSizeDistr(i) = 83,009 × ExpCRUSizeDistr(i)/Σ ExpCRUSizeDistr(i).
In our original model [22] with identical CRUs, we simulated various degrees of order and disorder in JSR positions by generating respective various degrees of deviations from their crystalline positions. In our new model, we developed a different approach that simulates CRU repulsion found in SIM images of RyR clusters. In short, the new algorithm considers CRUs as charged particles of same polarity with the effective charge and mass proportional to their sizes (i.e., NRyR). Initially, CRUs are distributed randomly (at time 0). JSR overlaps are excluded, i.e., any two JSRs cannot occupy the same cell volume. Then, CRUs move freely (but with friction), driven by respective Coulomb forces. The friction is important for the system to ultimately (in ~100 steps) evolve into a steady state exhibiting a crystal-like structure due to strong repulsion (see Videos S1 and S2, for CRUs of heterogeneous and identical sizes, respectively). Thus, iteration steps create a series of distributions of CRU positions reflecting various degrees of repulsion (or order and disorder) (Figure A2). It is important to note, however, that we do not know the nature of repulsion in real CRU distributions, and we found all parameters in the repulsion algorithm empirically. We use Coulomb forces here just to create a computationally effective repulsion process that generates respective CRU distributions with various degrees of repulsion. Those distributions are further tested to evaluate the functional effect of the repulsion per se, regardless the nature of the repulsion.
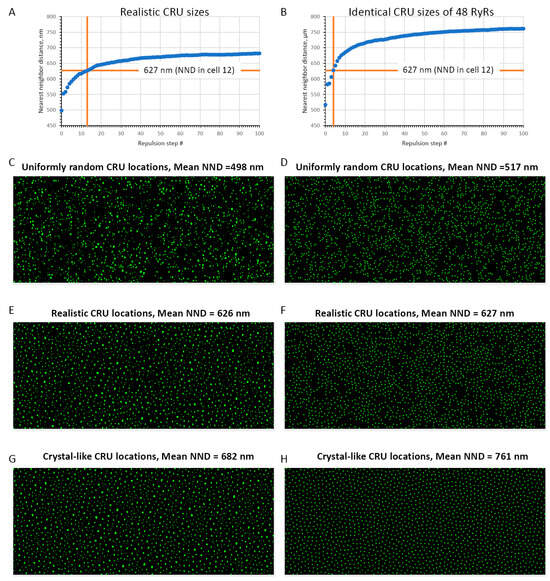
Figure A2.
Various degrees of CRU repulsion (i.e., order and disorder) generated by our repulsion algorithm in a cell with realistic CRU sizes (as in cell #8, Table 1) ((left) panels) and in a virtual cell with CRUs of identical sizes accommodating 48 RyRs ((right) panels). Top panels (A,B) show in blue circles how average NND changed in the respective CRU networks under cell membrane as repulsion transformed uniformly randomly distributed CRUs (step 0) towards a crystal-like structure (step 100). An intermediate CRU network with mean nearest neighbor distance (NND) close to that measured experimentally was used in our model simulations (steps 13 and 4 for networks of realistic and identical CRU sizes, respectively, vertical orange lines in (A,B)). Panels (C–H) show CRU distributions under the cell membrane (cylinder surfaces unwrapping to squares) that were used in our numerical model simulations of SANC function. See also Videos S1 and S2 for more details, including NND distributions, mean NND, and NND standard deviation at each repulsion step.
Figure A2.
Various degrees of CRU repulsion (i.e., order and disorder) generated by our repulsion algorithm in a cell with realistic CRU sizes (as in cell #8, Table 1) ((left) panels) and in a virtual cell with CRUs of identical sizes accommodating 48 RyRs ((right) panels). Top panels (A,B) show in blue circles how average NND changed in the respective CRU networks under cell membrane as repulsion transformed uniformly randomly distributed CRUs (step 0) towards a crystal-like structure (step 100). An intermediate CRU network with mean nearest neighbor distance (NND) close to that measured experimentally was used in our model simulations (steps 13 and 4 for networks of realistic and identical CRU sizes, respectively, vertical orange lines in (A,B)). Panels (C–H) show CRU distributions under the cell membrane (cylinder surfaces unwrapping to squares) that were used in our numerical model simulations of SANC function. See also Videos S1 and S2 for more details, including NND distributions, mean NND, and NND standard deviation at each repulsion step.
Each JSR is diffusely linked to the network of FSR that uptakes Ca2+ from cytosol via sarco/endoplasmic reticulum Ca-ATPase (SERCA) pumping. We placed JSRs inside the respective ring voxels just below their outer side facing the cell membrane (Figure A1). Volumes of the ring voxels were kept the same by their extending into the core by the exact volume that the JSR occupied at their outer side. Thus, the actual core volume was calculated as the volume of the cylinder core decreased by the volume of all JSR volumes.
Appendix A.2. Ca2+ Cycling
Appendix A.2.1. Free SR (FSR)
As mentioned above, each ring voxel and the core were further partitioned into cytosol and FSR fractions. We model FSR as a homogeneously distributed network within the cytosol. The cytosol fraction is set to 0.46 and the FSR fraction to 0.035 [21]. The remainder presumably contains the myofilaments, mitochondria, nucleus, and other organelles. In turn, each FSR portion has capability to pump Ca2+ locally from the respective cytosol portion of the same voxel, simulating local SERCA function. Because the submembrane voxels are extremely thin, only 20 nm in depth, their contributions to Ca2+ pumping and intra-SR diffusion are negligible and not modeled.
Appendix A.2.2. SR Ca2+ Pump
The SERCA pump is present uniformly throughout the cell, transferring Ca2+ from the cytosolic to the FSR compartment of each voxel (of ring and core), with the Ca2+ uptake rate given by the reversible Ca2+ pump formulation adopted from Shannon et al. [54]
where Pup = 0.012 mM/ms, Kmf = 0.000246 mM, Kmr = 1.7 mM, and H = 1.787.
Appendix A.2.3. Ca2+ Diffusion Within and Among Cell Compartments
Ca diffusion fluxes between voxels within and among cell compartments are approximated by the first Fick’s law:
where D is a diffusion coefficient, and Δ[Ca]/Δx is the Ca2+ concentration gradient, i.e., Δ[Ca] is the concentration difference and Δx is the distance between the voxel centers.
J = −D·Δ[Ca]/Δx
The respective rate of change of [Ca] is defined as:
where s is the diffusion area sharing by voxels and v is the receiving volume. Thus,
dCa/dt = J·s/v
dCa/dt = −D·(Δ[Ca]/Δx)·s/v
For any two diffusively interacting voxels with volumes v1 and v2, Ca2+ dynamics are described by a set of differential equations:
where Ca1 and Ca2 are Ca2+ concentrations in the respective voxels, and
dCa1/dt = (Ca2 − Ca1)/τ1
dCa2/dt = (Ca1 − Ca2)/τ2
τ1 = v1·Δx/(D·s)
τ2 = v2·Δx/(D·s)
τ1 (or τ2, symmetrically) is the respective time constant of Ca1 change in time in a special case if the other compartment with volume v2 is substantially larger than v1 (i.e., v2 >> v1), and therefore, Ca2 remains approximately constant. In a general case, the set is analytically solved to the respective exponential decays:
where C∞= (Ca1(0)·v1 + Ca2(0)·v2)/(v1 + v2) is the equilibrium concentration (t = ∞) in both voxels defined by the matter conservation principle and τ = τ1·τ2/(τ1 + τ2) is the common time constant of the exponential decay of the system to reach the equilibrium. The respective Ca2+ change from its initial value in voxel v1 over time is given as follows:
Ca1(t) = (Ca1(0) − C∞)·exp(−t/τ)+ C∞
Ca2(t) = (Ca2(0) − C∞)·exp(−t/τ)+ C∞
ΔCa1(t) = (C∞ −Ca1(0)) ·(1 − exp(−t/τ))
Then, by substituting C∞, we obtain:
ΔCa1(Δt) = (v2/(v1 + v2))·(Ca2(0) −Ca1(0)) ·(1 − exp(−Δt/τ))
ΔCa2(Δt) = −(v1/(v1+v2))·(Ca2(0) −Ca1(0))·(1 − exp(−Δt/τ))= −ΔCa1(t)·v1/v2
These formulations are used in all our computations of [Ca] changes for all neighboring voxels within and among cell compartments for the model integration for each time update Δt (during time tick or several time ticks for slower processes). In our computer algorithm, we calculate the fractional Ca2+ change (FCC) before the model run and use it simply as a scaling factor to determine the actual [Ca] change from the difference in [Ca] between any two diffusely interacting voxels at the beginning of each integration step (from t = 0 to t = Δt). Thus,
FCC = (v2/(v1+v2))·(1 − exp(−Δt/τ)) (calculated before model run)
ΔCa1(Δt) = FCC·(Ca2(0) − Ca1(0)) (calculated during model run)
ΔCa2(Δt) = −ΔCa1(t)·v1/v2 (calculated during model run)
In the case of identical voxels, i.e., within the subspace and within the ring (i.e., when v1 = v2 and τ1= τ2), the formulations are simplified to:
FCC = 0.5·(1−exp(−2·Δt/τ1))
Note 1: If Δt/τ1 << 1, then FCC can be approximated (e.g., via respective Taylor series) as:
FCC = Δt/τ1 = Δt·D·s/(v1·Δx)
Further, if v1 can be described as v1 = s·Δx, e.g., for diffusion along the cell length (axis x in our model), FCC can be further simplified to
FCC = Δt·D/Δx2
However, because FCC is calculated only once, before the model run, and does not carry any additional computational burden during actual simulations of Ca2+ dynamics, here, we always use the full approximation for the diffusion, i.e., more precise exponential decay, rather than a linear change over Δt. An advantage of this approach is that the model features more stable behavior in case we want to vary cell geometry, cell compartments, voxel sizes, or integration time (Δt).
Note 2: We have only a fraction of cell volume occupied with cytosol (or FSR). However, the same fraction will be for v and s in τ formulations, and it cancels. The volume ratios v2/(v1+v2) and v1/v2 also remain unchanged because the fraction factor also cancels. Thus, all above formulations with formal geometric volumes are also valid for fractional volumes, assuming that the fraction of cytosol (or FSR) is evenly distributed within the volume.
Appendix A.2.4. Junctional SR (JSR) and Ca2+ Diffusion Between JSR and FSR
Each JSR is refilled with Ca2+ locally from the FSR network via a fixed diffusional resistance, yielding a mono-exponential process with a fixed time constant tau. In our original model [22] with fixed JSR sizes (144 RyRs), this tau was set to 40 ms, similarly to previous common pool models that approximated diffusion between JSR and FSR pools [55]. Here, we want to do the same for JSRs of various sizes. We do not consider Ca2+ diffusion within the JSR (i.e., each JSR Ca2+ is described by one variable), but each JSR can touch several FSR voxels (Figure A1). As described above, complex JSR geometries represent a collection of N-adjacent 120 × 120 × 60 nm parts/blocks, with each part accommodating 16 RyRs (N= NRyR/16). Hence, we set a diffusional link for each such JSR part to its adjacent FSR voxel (360 × 360 nm in xy). The net Ca2+ flux to a given JSR from FSR is calculated as a sum of Ca2+ fluxes via those individual links, with each link transferring Ca2+ flux with τ = N·40 ms·vFSR/(vFSR + vJSR), where vJSR is the volume of a given JSR and vFSR is the FSR volume within a ring voxel.
Appendix A.2.5. Spark Activation and Inactivation
Each CRU can be in either an open or closed state. The capability of a given CRU to open, i.e., to generate a Ca2+ spark, is controlled by its JSR Ca2+ loading. Experimental and theoretical studies have shown that sparks are generated when SR Ca2+ loading reaches a certain critical level [39,56]. This critical CajSR level is implemented in our mechanism of spark activation by prohibiting CRU firing while SR Ca2+ loading remains below 300 μM (CaSRfire). When JSR is refilled with Ca2+ above this level, it can open. The switch from a closed state to an open state is probabilistic. The probability density for a given closed CRU to open is described by a power function of Ca2+ concentration (Ca) in the dyadic space. In our original model [22], CRU size was fixed to 144 RyRs. The probability p144 for a CRU of this size to open during a short time interval, TimeTick, is given by
where Casens = 0.15 μM is the sensitivity of CRU to Ca2+ in dyadic space, ProbConst144 = 0.00027 ms−1 is the open probability rate at Ca2+ = Casens, and ProbPower = 3 defines the cooperativity of CRU activation by Ca2+ in dyadic space. In our new model, CRU sizes vary, and therefore, we adopted the original formulation as follows:
p144 = ProbConst144·(Ca/Casens)ProbPower·TimeTick
p(NRyR) = NRyR·ProbConst1·[b(NRyR)·Ca/Casens]ProbPower·TimeTick
In the new formulation, we scaled the probability rate with respect to NRyR in the CRU, so that ProbConst1 = ProbConst144/144 = 1.875 × 10−6 ms−1. We also introduced a boundary factor b(NRyR) that reflects a lower probability to ignite a spark by an RyR opening in smaller CRUs due to stronger escape of the released Ca2+ to bulk cytosol from the tight dyadic space via a closer CRU boundary to the open channel. This Ca2+ escape results in a lower (smaller) spatial Ca2+ profile from one open RyR, resulting in a weaker CICR with respect to neighboring RyRs and a lower probability of spark ignition. To find the boundary factor b(NRyR), we performed a series of numerical model simulations using Stern at al.’s spark model [52], which describes individual RyRs within a CRU. The results of these simulations are presented in Figure A3.
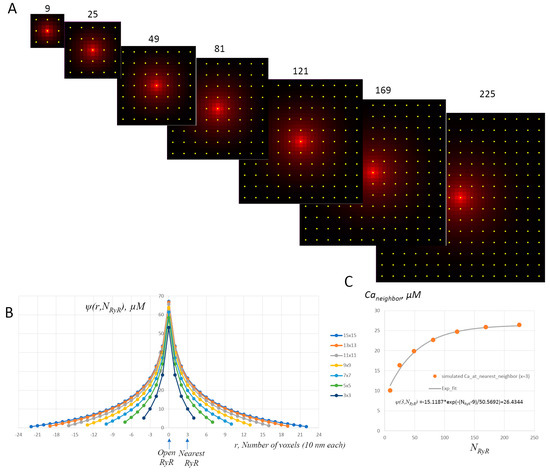
Figure A3.
Effect of Ca2+ escape from dyadic space on RyR interaction profile in CRUs of different sizes (boundary effect). (A): A cascade of distributions of Ca2+ in dyadic space for CRUs with different numbers of RyRs (shown by yellow circles): 3 × 3, 5 × 5, 7 × 7 etc. The distributions were simulated by running Stern at al.’s spark model [52], which describes the function of each individual RyR within an individual CRU. The distributions were assessed 10 ms after one RyR channel opened in the middle of each CRU. Other RyRs were not allowed to open. The initial [Ca] in JSR was set to 1 mM. [Ca] is coded by red shades, saturating (pure red) at 60 μM. The voxel size in the model is 10 × 10 nm in the xy plane, parallel to the cell surface membrane. (B): Ca2+ profiles (interaction profiles) via the CRU center of the respective distributions in the panel. The distance r is given by the model voxel size (10 nm). Arrows show positions of open RyRs in the CRU center (r = 0) and its nearest RyR neighbor (r = 3 voxels). (C): Plot of Ca2+ at the nearest neighbor for CRUs of different sizes Caneighbor = ψ(3,NRyR) together with its exponential fit (R2 = 0.976) is shown by the grey line with the equation at the bottom of the plot.
Figure A3.
Effect of Ca2+ escape from dyadic space on RyR interaction profile in CRUs of different sizes (boundary effect). (A): A cascade of distributions of Ca2+ in dyadic space for CRUs with different numbers of RyRs (shown by yellow circles): 3 × 3, 5 × 5, 7 × 7 etc. The distributions were simulated by running Stern at al.’s spark model [52], which describes the function of each individual RyR within an individual CRU. The distributions were assessed 10 ms after one RyR channel opened in the middle of each CRU. Other RyRs were not allowed to open. The initial [Ca] in JSR was set to 1 mM. [Ca] is coded by red shades, saturating (pure red) at 60 μM. The voxel size in the model is 10 × 10 nm in the xy plane, parallel to the cell surface membrane. (B): Ca2+ profiles (interaction profiles) via the CRU center of the respective distributions in the panel. The distance r is given by the model voxel size (10 nm). Arrows show positions of open RyRs in the CRU center (r = 0) and its nearest RyR neighbor (r = 3 voxels). (C): Plot of Ca2+ at the nearest neighbor for CRUs of different sizes Caneighbor = ψ(3,NRyR) together with its exponential fit (R2 = 0.976) is shown by the grey line with the equation at the bottom of the plot.
Our previous studies showed that the critical interaction of RyRs within a CRU with respect to CICR and spark activation is described by the interaction profile, ψ(r), i.e., the profile of Ca2+ drop in dyadic space when one RyR stays open, and especially Ca2+ at the nearest RyR neighbor [39,57]. Thus, we found a series of interaction profiles ψ(r) as a function of NRyR for a large variety of CRU sizes from 3 × 3 to 15 × 15 RyRs. Then, we plotted Ca2+ at the nearest RyR neighbor of the open channel and fitted it to a phenomenological exponential curve Caneighbor(N) that closely described waning RyR interactions for smaller CRUs (grey line in Figure A3C). To obtain the boundary factor in Equation (A2b), we normalized the exponential curve to its value at 144 (i.e., b(144) = 1) to be consistent with our previous formulation for the CRU with 144 RyRs [22]. Because Caneighbor(144) = 25.387 μM, we finally obtained:
b(NRyR) = (−15.1187 × exp(−(NRyR− 9)/50.5692) + 26.4344)/25.387
At each time tick, our computer algorithm tried to activate a closed CRU by generating a random number within (0, 1). If this number was less than p (in Equation (A2b)), then the CRU opened. The Ca2+ current amplitude, Ispark, is defined by the spark activation kinetics a(t), RyR unitary current (IRyR,1mM, the current via a single RyR at 1 mM of Ca2+ gradient), NRyR, and concentration difference between inside and outside JSR:
Ispark = a(t)· i(t)·IRyR,1mM·NRyR· (Ca− CaJSR)
IRyR,1mM is set to 0.35 pA [52]. Spark activation a(t) kinetics is described as a single exponential time-dependent process with τa = 10 ms:
a(t) = 1− exp(−t/τa)
Because each open RyR undergoes time-dependent inactivation, this is also reflected in our CRU model by adding simple inactivation kinetic variable i(t) with a time constant τi = 1/kom = 8.547 ms (inactivation rate constant kom = 0.117 ms−1 in Stern et al.’s spark model [52]):
i(t) = exp(−t/τi)
Appendix A.2.6. Spark Termination Mechanism
Our spark termination mechanism is based on the current knowledge in this research area [52,57,58], i.e., a Ca2+ spark is generated via CICR among individual RyRs within a CRU, and it sharply terminates due to induction decay [58], CICR interruption [52], or a phase transition (similar to that known in Ising model) [57] when the RyR current IRyR(t) becomes too small (due to JSR Ca2+ depletion) to further support the CICR among RyRs. The specific value of the critical current Ispark_termination is defined by interactions of individual RyRs and is beyond the capability of our CRU-based model. Thus, we terminated the spark when the spark current Ispark(t) became very small, equal to RyR current IRyR(t), i.e., the current via one RyR channel at a given JSR Ca2+ load.
Appendix A.2.7. Ca2+ Buffering
Cytosolic Ca2+ is buffered by calmodulin (0.045 mM) throughout the cell: submembrane voxels, ring voxels, and the core. Each JSR features Ca2+ buffering with calsequestrin (30 mM).
Appendix A.2.8. Summary of Equations of Local Ca2+ Dynamics
In a submembrane voxel:
Ispark in voxels outside the dyadic space is absent. Each CRU releases Ca2+ (given by Ispark) into its dyadic space. Ispark is evenly distributed among submembrane voxels of the dyadic space (Nvoxels_in_dyad). Ispark is given in Equation (A3), and it is positive, i.e., increasing [Ca] in the submembrane voxel. iCa is the sum of local Ca2+ transmembrane currents (described in detail below in the Electrophysiology section) via the membrane patch facing this submembrane voxel:
iCa = iCaL + iCaT + ibCa− 2∙iNCX
ICaL is included in this equation only for submembrane voxels facing a CRU (ICaL is injected into the subspace voxels of the respective dyadic space). The local Ca2+ currents iCaL, iCaT, ibCa have inward direction, and (by convention) are defined as negative. Therefore, the minus sign before iCa in Equation (A5) ensures a positive change in [Ca] in the submembrane voxel due to respective Ca2+ influx. During diastole, local iNCX also flows inwardly, but NCX exchanges 1 Ca2+ ion to 3 Na ions. Hence, the inward iNCX generates a Ca2+ efflux. That is why it has a different sign.
In a voxel of the ring layer:
is the sum of diffusion fluxes from neighboring smaller submembrane voxels.
The SERCA uptake flux jup is given by Equation (A1).
In a given JSR:
where Ispark is given by Equation (A3).
is the sum of diffusion fluxes between JSR and the respective FSR parts of the neighboring ring voxels. jCQ is Ca2+ flux of Ca2+ buffering by calsequestrin:
dfCQ/dt = kfCQ ∙CajSR∙(1- fCQ) − kbCQ ∙ fCQ(t)
In the core:
where and are the respective sums of diffusion fluxes in cytosol and FSR of all ring voxels. In any cytoplasmic voxel (subspace, ring, and the core), jCM is Ca2+ flux of Ca2+ buffering by calmodulin:
Appendix A.3. Electrophysiology
Major changes in the model include introduction of local Ca2+ currents and the modulation of local currents by local Ca. To introduce local currents and local modulation by Ca, the cell membrane is partitioned into small patches, with each patch facing its respective subspace voxel. We also omitted the sustained inward current Ist and the background Na current IbNa because, thus far, the molecular identities for these currents have not been found, and these currents are likely produced by NCX or other currents [7].
Appendix A.3.1. Fixed Ion Concentrations, mM
Cao = 2: Extracellular Ca2+ concentration.
Ko = 5.4: Extracellular K concentration.
Ki = 140: Intracellular K concentration.
Nao = 140: Extracellular Na concentration.
Nai = 10: Intracellular Na concentration.
Appendix A.3.2. The Nernst Equation and Electric Potentials, mV
ENa = ET∙ln(Nao/Nai): Equilibrium potential for Na
EK = ET∙ln(Ko/Ki): Equilibrium potential for K
EKs = ET∙ln[(Ko + 0.12∙ Nao)/(Ki + 0.12 ∙ Nai)]: Reversal potential of IKs
Where ET is “RT/F” factor = 26.72655 mV at 37 °C,
ECaL = 45: Apparent reversal potential of ICaL
ECaT = 45: Apparent reversal potential of ICaT
Appendix A.3.3. Membrane Potential, Vm
Net membrane current determines the time derivative of the membrane potential.
dVm/dt = −(ICaL + ICaT + IKr + IKs + Ito + Isus + If + INaK + IbCa + INCX)/Cm
Appendix A.3.4. Formulation of Cell Membrane Ion Currents
The kinetics of ion currents are described by gating variables (described below) in respective differential equations:
dyi/dt = (yi,∞ − y)/τyi
(yi = dL, fL, fCa, dT, fT, paF, paS, pi_, n, q, r, y)
τyi: time constant for a gating variable yi.
αyi and βyi: opening and closing rates for channel gating.
yi,∞: steady-state curve for a gating variable yi.
L-type Ca2+ current (ICaL)
The whole cell ICaL is calculated as a sum of local currents iCaL,i in each dyadic space.
This reflects reports that L-type Ca2+ cannels (LCCs) are colocalized with RyRs [59]. Thus, the whole-cell maximum ICaL conductance (gCaL) is distributed locally among dyadic spaces proportionally to their areas:
where si is the submembrane area of the i-th CRU and Σsi is the area occupied by all CRUs. The respective local iCaL,i is calculated in each CRU as a sum of two currents generated by two major LCC isoforms, Cav1.2 and Cav1.3, present in SANCs [2,60]
where η1.2 and η1.3 are the respective fractions of the Cav1.2 and Cav1.3 isoforms (η1.2 + η1.3 = 1). Cav1.3 has a lower (more hyperpolarized) voltage activation threshold.
gCaL,i = gCaL·si/Σsi
iCaL,i = iCaL1.2,i + iCaL1.3,i
iCaL1.2,i = Cm·gCaL,i ·η1.2· (Vm- ECaL)·dL1.2·fL·fCa,i
iCaL1.3,i = Cm·gCaL,i·η1.3· (Vm- ECaL)·dL1.3·fL·fCa,i
dL1.2,∞ = 1/{1+ exp[−(Vm − Vd1.2)/6]}
dL1.3,∞ = 1/{1+ exp[−(Vm − Vd1.3)/6]}
We set Vd1.2 the midpoint of ICaL1.2 activation to −6.6 mV, as in the earlier SANC models of Wilders et al. [61] and Dokos et al. [62]. This value of Vd1.2 is relatively high with respect to other SANC models, and therefore was considered in this study (and our previous study [22]) to simulate ICaL generated by Cav1.2. Vd1.3, the midpoint of ICaL1.3 activation, is set to −13.5 mV. The resultant shift in activation increased dL1.3,∞vs. dL1.2,∞within the range of diastolic depolarization by a factor of ~3 [22], generating a much larger current and stronger recruitment of CRUs to fire during diastolic depolarization, which is consistent with the current concept of the important role of the Cav1.3 isoform in cardiac pacemaker function [46,47]. Other parameters of L-type Ca2+ currents are the same for both isoforms:
fL,∞ = 1/{1 + exp[(Vm + 35)/7.3]}
αdL = −0.02839·(Vm+ 35)/{exp[−(Vm +35)/2.5] − 1} − 0.0849 ·Vm/[exp(−Vm/4.8) − 1]
βdL = 0.01143·(Vm − 5)/{exp[(Vm − 5)/2.5] 1}
τdL = 1/(αdL + βdL)
τfL = 128.55· exp{−[(Vm+ 32.5)/13.9]2}+ 44.3)
fCa,∞ = KmfCa/(KmfCa + Casub,i)
τfCa = fCa,∞/αfCa
Ca-dependent ICaL inactivation is described by the parameters KmfCa and αfCa. The T-type Ca2+ current (ICaT) is based on formulations of Demir et al. [63] and modified by Kurata et al. [55].
ICaT = Cm∙gCaT,max ∙ (Vm − ECaT)∙dT∙fT
dT,∞ = 1/{1 + exp[-(Vm + 26.3)/6.0]}
fT,∞ = 1/{1+ exp[(Vm+ 61.7)/5.6]}
τdT = 1/{1.068∙exp[(Vm+ 26.3)/30] + 1.068∙exp[−(Vm + 26.3)/30]}
τfT = 1/{0.0153∙exp[−(Vm + 61.7)/83.3] + 0.015∙exp[(Vm + 61.7)/15.38]}
The whole-cell ICaT is evenly distributed over the cell membrane patches to generate the respective homogeneous Ca2+ influx. In each submembrane, the i-th voxel ICaT,i = ICaT/Nvoxels. The rapidly activating delayed rectifier K+ current (IKr) is based on the formulations of Zhang et al. [53] and further modified by Kurata et al. [55].
IKr = Cm∙gKr,max ∙(Vm − EK)∙(0.6∙ paF + 0.4∙ paS) ∙ pi
pa,∞ = 1/{1 + exp[−(Vm +23.2)/10.6]}
pi,∞ = 1/{1 + exp[(Vm + 28.6)/17.1]}
τpaF = k_tau_IKr ∙ 0.84655354/[0.0372 ∙ exp(Vm/15.9) + 0.00096 ∙ exp(−Vm/22.5)]
τpaS = k_ tau_IKr ∙ 0.84655354/[0.0042 ∙ exp(Vm/17.0) + 0.00015 ∙ exp(−Vm/21.6)]
τpi = 1/[0.1 ∙ exp(−Vm/54.645) + 0.656 ∙ exp(Vm/106.157)]
The slowly activating delayed rectifier K+ current (IKs) is based on the formulations of Zhang et al. [53].
IKs = Cm∙gKs,max ∙(Vm − EKs)∙ n2
αn = 0.014/{1 + exp[-(Vm − 40)/9]}
βn = 0.001 ∙ exp(−Vm/45)
n∞ = αn/(αn + βn)
τn = 1/(αn + βn)
The 4-aminopyridine-sensitive currents (I4AP = Ito + Isus) are based on the formulations of Zhang et al. [53].
Ito = Cm ∙ gto,max ∙ (Vm − EK) ∙ q r
Isus = Cm ∙ gsus,max ∙ (Vm − EK) ∙ r
q∞ = 1/{1 + exp[(Vm + 49)/13]}
r∞ = 1/{1 + exp[-(Vm − 19.3)/15]}
τq = 39.102/{0.57∙exp[−0.08∙(Vm +44)] + 0.065∙exp [0.1∙(Vm +45.93)]}+ 6.06
τr = 14.40516/{1.037∙exp [0.09∙(Vm +30.61)] + 0.369∙exp[−0.12∙(Vm +23.84)]}+ 2.75352
The hyperpolarization-activated, “funny” current (If) is based on formulations of Wilders et al. [61] and Kurata et al. [55].
If = IfNa+ IfK
y∞ = 1/{1 + exp[(Vm −VIf,1/2)/13.5]}
τy = 0.7166529/{exp[-(Vm+ 386.9)/45.302] + exp[(Vm − 73.08)/19.231]}
IfNa = Cm∙0.3833 ∙gIf,max ∙ (Vm − ENa)∙y2
IfK = Cm∙0.6167 ∙ gIf,max ∙ (Vm − EK)∙y2
The Na+-K+ pump current (INaK) is based on the formulations of Kurata et al. [55], which were in turn based on the experimental work of Sakai et al. [64] for rabbit SANCs.
INaK = Cm ∙INaK,max ∙{1 + (KmKp/Ko)1.2}−1 ∙ {1 + (KmNap/Nai)1.3} −1 ∙ {1 + exp[-(Vm- ENa + 120)/30]}−
Ca- background current (IbCa)
IbCa = Cm∙ gbCa ∙(Vm − ECaL)
The whole-cell IbCa is evenly distributed over the cell membrane to generate a respective homogeneous Ca2+ influx. In each submembrane, the i-th voxel IbCa,i = IbCa/Nvoxels. The Na-Ca exchanger current (INCX) is based on original formulations from Dokos et al. [62]. INCX is modulated by local Ca2+, and therefore, the whole-cell INCX is calculated as a sum of local currents INCX,i in respective membrane patches facing each submembrane voxel. Thus,
For membrane voltage Vm in each i-th membrane patch with subspace Casub,i, the respective local NCX current (INCX,i) is calculated as follows:
INCX,i = Cm ∙kNCX ∙(k21 ∙ x2 − k12 ∙ x1)/(x1 + x2 + x3 + x4)
do = 1 + (Cao/Kco)∙{1 + exp(Qco∙Vm/ET)} + (Nao/K1no)∙{1 + (Nao/K2no)∙(1 + Nao/K3no)}
k43 = Nai/(K3ni + Nai)
k41 = exp[−Qn∙Vm/(2ET)]
k34 = Nao/(K3no + Nao)
k21 = (Cao/Kco)∙exp(Qco∙Vm/ET)/do
k23 = (Nao/K1no)∙(Nao/K2no)∙(1 + Nao/K3no)∙ exp[−Qn∙Vm/(2ET)]/do
k32 = exp[Qn∙Vm/(2ET)]
x1 = k34 ∙ k41 ∙(k23 + k21) + k21 ∙ k32 ∙(k43 + k41)
di = 1 + (Casub,i/Kci)∙{1 + exp(−Qci∙Vm/ET) + Nai/Kcni} + (Nai/K1ni)∙{1 + (Nai/K2ni)∙(1 + Nai/K3ni)}
k12 = (Casub,i/Kci)∙exp(−Qci∙Vm/ET)/di
k14 = (Nai/K1ni)∙(Nai/K2ni)∙(1 + Nai/K3ni)∙ exp[Qn∙Vm/(2ET)]/di
x2 = k43∙ k32 (k14 + k12) + k41 k12∙k34 + k32)
x3 = k43 ∙k14 ∙(k23 + k21) + k12∙k23∙(k43 + k41)
x4 = k34 ∙k23 ∙(k14 + k12) + k21 ∙k14∙(k34+ k32)
Appendix A.4. Initial Values
Appendix A.4.1. Electrophysiology
| −57.96393469 | Vm | Membrane potential, mV |
| 0.000584546 | dL | ICaL voltage-dependent activation |
| 0.86238125 | fL | ICaL voltage-dependent inactivation |
| 0.71139592 | fCa | ICaL Ca-dependent inactivation (local) |
| 0.144755091 | paF | IKr fast activation |
| 0.453100577 | paS | IKr slow activation |
| 0.849409822 | pi_ | IKr inactivation |
| 0.026460041 | n | IKs activation |
| 0.113643187 | y | If activation |
| 0.005043934 | dT | ICaT activation |
| 0.420757825 | fT | ICaT inactivation |
| 0.694241314 | q | Ito inactivation |
| 0.005581317 | r | Ito and Isus activation |
Appendix A.4.2. Ca2+ Dynamics
| 200 nM | [Ca] in cytosol |
| 1 mM | [Ca] in FSR |
| 0.8 mM | Ca concentration in JSR |
| 0.0787 | Fractional occupancy of calmodulin by Ca2+ in cytoplasm |
| 0.6 | Fractional occupancy of calsequestrin by Ca2+ in JSR |
All CRUs are set in the closed state.
Appendix A.5. Simulations of βAR Stimulation Effect
The effect of βAR stimulation is modeled essentially as we previously reported [22], by increasing the ICaL, IKr, If, and Ca2+ uptake rates by FSR via SERCA pumping. Specifically, the whole-cell maximum ICaL conductance gCaL was increased by a factor of 1.75 from 0.464 to 0.812 nS/pF; the whole-cell maximum IKr conductance gKr was increased by a factor of 1.5 from 0.05679781 to 0.085196715 nS/pF; the midpoint of the If activation curve was shifted to more depolarized potential by 7.8 mV from −64 mV to −56.2 mV; and the maximum Ca2+ uptake rate Pup was increased by a factor of 2 from 0.012 mM/ms to 0.024 mM/ms.
Appendix A.6. Model Integration
The model code was written in Delphi Language (Delphi 10.4) and was computed with a fixed time tick of 0.0075 ms on a workstation running Windows 10 with an Intel® Xeon® W-2145 CPU @3.7GHz processor.
Appendix A.7. Appendix Glossary and Parameters’ Values
Fixed ion concentrations:
Cao = 2 mM: extracellular [Ca]
Ko = 5.4 mM: extracellular [K]
Nao = 140 mM: extracellular [Na]
Ki = 140 mM: intracellular [K]
Nai = 10 mM: intracellular [Na]
Membrane currents
ECaL = 45 mV: apparent reversal potential of ICaL
gCaL = 0.464 nS/pF: conductance of ICaL
η1.2= η1.3 = 0.5: contributions of Cav1.2 and Cav1.3 isoforms to total gCaL
KmfCa = 0.03 mM: dissociation constant of Ca2+ -dependent ICaL inactivation
betafCa = 60 mM−1·ms−1: Ca2+ association rate constant for ICaL.
alfafCa = 0.021 ms−1: Ca2+ dissociation rate constant for ICaL
ECaT = 45: Apparent reversal potential of ICaT, mV
gCaT = 0.1832 nS/pF: conductance of ICaT
gIf = 0.105 nS/pF: conductance of If
VIf,1/2 = −64: half activation voltage of If, mV
gKr = 0.05679781 nS/pF: conductance of delayed rectifier K current rapid component
k_ tau_IKr = 0.3: scaling factor for tau_paF and tau_paS used to tune the model
gKs = 0.0259 nS/pF: conductance of delayed rectifier K current slow component
gto = 0.252 nS/pF: conductance of 4-aminopyridine sensitive transient K+ current
gsus = 0.02 nS/pF: conductance of 4-aminopyridine sensitive sustained K+ current
INaKmax = 1.44 pA/pF: maximum Na+/K+ pump current
KmKp = 1.4 mM: half-maximal Ko for INaK.
KmNap = 14 mM: half-maximal Nai for INaK.
gbCa = 0.003 nS/pF: conductance of background Ca2+ current,
kNCX = 48.75 pA/pF: maximumal amplitude of INCX
Dissociation constants for NCX:
K1ni = 395.3 mM: intracellular Na binding to first site on NCX
K2ni = 2.289 mM: intracellular Na binding to second site on NCX
K3ni = 26.44 mM: intracellular Na binding to third site on NCX
K1no = 1628 mM: extracellular Na binding to first site on NCX
K2no = 561.4 mM: extracellular Na binding to second site on NCX
K3no = 4.663 mM: extracellular Na binding to third site on NCX
Kci = 0.0207 mM: intracellular Ca2+ binding to NCX transporter
Kco = 3.663 mM: extracellular Ca2+ binding to NCX transporter
Kcni = 26.44 mM: intracellular Na and Ca2+ simultaneous binding to NCX
NCX fractional charge movement
Qci= 0.1369: intracellular Ca2+ occlusion reaction of NCX
Qco = 0: extracellular Ca2+ occlusion reaction of NCX
Qn= 0.4315: Na occlusion reactions of NCX
Ca buffering
kbCM = 0.542 ms−1: Ca2+ dissociation constant for calmodulin
kfCM = 227.7 mM−1 ms−1: Ca2+ association constant for calmodulin
kbCQ = 0.445 ms−1: Ca2+ dissociation constant for calsequestrin
kfCQ = 0.534 mM−1 ms−1: Ca2+ association constant for calsequestrin
CQtot = 30 mM: total calsequestrin concentration
CMtot = 0.045 mM: total calmodulin concentration
SR Ca2+ ATPase function
Kmf = 0.000246 mM: the cytosolic side Kd of SR Ca2+ pump
Kmr = 1.7 mM: the lumenal side Kd of SR Ca2+ pump
H = 1.787: cooperativity of SR Ca2+ pump
Pup = 0.012 mM/ms: maximal rate of Ca2+ uptake by SR Ca2+ pump
CRU (Ca release and JSR)
Casens = 0.00015 mM: sensitivity of Ca2+ release to Casub
ProbConst1 = 1.875 × 10−6 ms−1: CRU probability constant per each RyR in the CRU
ProbPower = 3: cooperativity of CRU activation by Casub
CaJSR_spark_activation = 0.3 mM: critical JSR Ca2+ loading to generate a spark (CRU can open)
τCRUactivation = 10 ms: time constant of spark activation (a)
τCRUinactivation = 8.547 ms: time constant of spark inactivation (i)
Iryr_at_1mM_CaJSR = 0.35 pA: unitary RyR current at 1 mM delta Ca
RyR_to_RyR_distance_um = 0.03 μm: RyR crystal grid size in JSR
JSR_depth_um = 0.06 μm: JSR depth
JSR_Xsize_um = 0.36 μm: JSR size in x
JSR_Ysize_um = 0.36 μm: JSR size in y
JSR_to_JSR_X_um = 1.44 μm: JSR crystal grid size in x
JSR_to_JSR_Y_um = 1.44 μm: JSR crystal grid size in y
Cell geometry, compartments, and voxels:
Lcell = 53.28 μm: cell length
rcell = 3.437747 μm: cell radius
Cm = 19.80142 pF: membrane electrical capacitance of our cell model with 0.0172059397937 pF/μm2 specific membrane capacitance calculated from 2002 Kurata et al. model [55] for its 32 pF cylinder cell of 70 μm length and 4 μm radius.
0.12 μm: the grid size
0.12 μm: submembrane voxel size in x
0.12 μm: submembrane voxel size in y
0.02 μm: submembrane voxel size depth
0.36 μm: ring voxel size in x
0.36 μm: ring voxel size in y
0.8 μm: ring voxel depth
0.46: fractional volume of cytosol
0.035: fractional volume of FSR
Ca diffusion
Dcyt = 0.35 μm2/ms: diffusion coefficient of free Ca2+ in cytosol
DFSR = 0.06 μm2/ms: diffusion coefficient of free Ca2+ in FSR.
References
- Boyett, M.R.; Honjo, H.; Kodama, I. The sinoatrial node, a heterogeneous pacemaker structure. Cardiovasc. Res. 2000, 47, 658–687. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.E.; Nargeot, J. Genesis and regulation of the heart automaticity. Physiol. Rev. 2008, 88, 919–982. [Google Scholar] [CrossRef] [PubMed]
- Dobrzynski, H.; Boyett, M.R.; Anderson, R.H. New insights into pacemaker activity: Promoting understanding of sick sinus syndrome. Circulation 2007, 115, 1921–1932. [Google Scholar] [CrossRef]
- Liu, Y.; Zheng, Y.; Tse, G.; Bazoukis, G.; Letsas, K.; Goudis, C.; Korantzopoulos, P.; Li, G.; Liu, T. Association between sick sinus syndrome and atrial fibrillation: A systematic review and meta-analysis. Int. J. Cardiol. 2023, 381, 20–36. [Google Scholar] [CrossRef]
- Lang, D.; Glukhov, A.V. Cellular and Molecular Mechanisms of Functional Hierarchy of Pacemaker Clusters in the Sinoatrial Node: New Insights into Sick Sinus Syndrome. J. Cardiovasc. Dev. Dis. 2021, 8, 43. [Google Scholar] [CrossRef]
- Jensen, P.N.; Gronroos, N.N.; Chen, L.Y.; Folsom, A.R.; deFilippi, C.; Heckbert, S.R.; Alonso, A. Incidence of and risk factors for sick sinus syndrome in the general population. J. Am. Coll. Cardiol. 2014, 64, 531–538. [Google Scholar] [CrossRef]
- Lakatta, E.G.; Maltsev, V.A.; Vinogradova, T.M. A coupled SYSTEM of intracellular Ca2+ clocks and surface membrane voltage clocks controls the timekeeping mechanism of the heart’s pacemaker. Circ. Res. 2010, 106, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Inui, M.; Saito, A.; Fleischer, S. Isolation of the ryanodine receptor from cardiac sarcoplasmic reticulum and identity with the feet structures. J. Biol. Chem. 1987, 262, 15637–15642. [Google Scholar] [CrossRef]
- Franzini-Armstrong, C.; Protasi, F.; Ramesh, V. Shape, size, and distribution of Ca2+ release units and couplons in skeletal and cardiac muscles. Biophys. J. 1999, 77, 1528–1539. [Google Scholar] [CrossRef]
- Fabiato, A. Calcium-induced release of calcium from the cardiac sarcoplasmic reticulum. Am. J. Physiol. 1983, 245, C1–C14. [Google Scholar] [CrossRef]
- Cheng, H.; Lederer, W.J.; Cannell, M.B. Calcium sparks: Elementary events underlying excitation-contraction coupling in heart muscle. Science 1993, 262, 740–744. [Google Scholar] [CrossRef]
- Cheng, H.; Lederer, W.J. Calcium sparks. Physiol. Rev. 2008, 88, 1491–1545. [Google Scholar] [CrossRef] [PubMed]
- Rubenstein, D.S.; Lipsius, S.L. Mechanisms of automaticity in subsidiary pacemakers from cat right atrium. Circ. Res. 1989, 64, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Huser, J.; Blatter, L.A.; Lipsius, S.L. Intracellular Ca2+ release contributes to automaticity in cat atrial pacemaker cells. J. Physiol. 2000, 524 Pt 2, 415–422. [Google Scholar] [CrossRef]
- Groenke, S.; Larson, E.D.; Alber, S.; Zhang, R.; Lamp, S.T.; Ren, X.; Nakano, H.; Jordan, M.C.; Karagueuzian, H.S.; Roos, K.P.; et al. Complete atrial-specific knockout of sodium-calcium exchange eliminates sinoatrial node pacemaker activity. PLoS ONE 2013, 8, e81633. [Google Scholar] [CrossRef] [PubMed]
- Lyashkov, A.E.; Behar, J.; Lakatta, E.G.; Yaniv, Y.; Maltsev, V.A. Positive Feedback Mechanisms among Local Ca Releases, NCX, and ICaL Ignite Pacemaker Action Potentials. Biophys. J. 2018, 114, 1176–1189. [Google Scholar] [CrossRef]
- Rigg, L.; Heath, B.M.; Cui, Y.; Terrar, D.A. Localisation and functional significance of ryanodine receptors during beta-adrenoceptor stimulation in the guinea-pig sino-atrial node. Cardiovasc. Res. 2000, 48, 254–264. [Google Scholar] [CrossRef]
- Musa, H.; Lei, M.; Honjo, H.; Jones, S.A.; Dobrzynski, H.; Lancaster, M.K.; Takagishi, Y.; Henderson, Z.; Kodama, I.; Boyett, M.R. Heterogeneous expression of Ca2+ handling proteins in rabbit sinoatrial node. J. Histochem. Cytochem. 2002, 50, 311–324. [Google Scholar] [CrossRef]
- Maltsev, A.V.; Maltsev, V.A.; Mikheev, M.; Maltseva, L.A.; Sirenko, S.G.; Lakatta, E.G.; Stern, M.D. Synchronization of stochastic Ca2+ release units creates a rhythmic Ca2+ clock in cardiac pacemaker cells. Biophys. J. 2011, 100, 271–283. [Google Scholar] [CrossRef]
- Maltsev, A.V.; Yaniv, Y.; Stern, M.D.; Lakatta, E.G.; Maltsev, V.A. RyR-NCX-SERCA local crosstalk ensures pacemaker cell function at rest and during the fight-or-flight reflex. Circ. Res. 2013, 113, e94–e100. [Google Scholar] [CrossRef]
- Stern, M.D.; Maltseva, L.A.; Juhaszova, M.; Sollott, S.J.; Lakatta, E.G.; Maltsev, V.A. Hierarchical clustering of ryanodine receptors enables emergence of a calcium clock in sinoatrial node cells. J. Gen. Physiol. 2014, 143, 577–604. [Google Scholar] [CrossRef] [PubMed]
- Maltsev, A.V.; Stern, M.D.; Maltsev, V.A. Disorder in Ca2+ release unit locations confers robustness but cuts flexibility of heart pacemaking. J. Gen. Physiol. 2022, 154, e202113061. [Google Scholar] [CrossRef]
- Weiss, J.N.; Qu, Z. The Sinus Node: Still Mysterious After All These Years. JACC Clin. Electrophysiol. 2020, 6, 1841–1843. [Google Scholar] [CrossRef]
- Clancy, C.E.; Santana, L.F. Evolving Discovery of the Origin of the Heartbeat: A New Perspective on Sinus Rhythm. JACC Clin. Electrophysiol. 2020, 6, 932–934. [Google Scholar] [CrossRef]
- Gustafsson, M.G. Surpassing the lateral resolution limit by a factor of two using structured illumination microscopy. J. Microsc. 2000, 198, 82–87. [Google Scholar] [CrossRef]
- Vinogradova, T.M.; Zhou, Y.Y.; Bogdanov, K.Y.; Yang, D.; Kuschel, M.; Cheng, H.; Xiao, R.P. Sinoatrial node pacemaker activity requires Ca2+/calmodulin-dependent protein kinase II activation. Circ. Res. 2000, 87, 760–767. [Google Scholar] [CrossRef]
- Wu, Y.; Shroff, H. Faster, sharper, and deeper: Structured illumination microscopy for biological imaging. Nat. Methods 2018, 15, 1011–1019. [Google Scholar] [CrossRef]
- York, A.G.; Chandris, P.; Nogare, D.D.; Head, J.; Wawrzusin, P.; Fischer, R.S.; Chitnis, A.; Shroff, H. Instant super-resolution imaging in live cells and embryos via analog image processing. Nat. Methods 2013, 10, 1122–1126. [Google Scholar] [CrossRef]
- Curd, A.; Cleasby, A.; Makowska, K.; York, A.; Shroff, H.; Peckham, M. Construction of an instant structured illumination microscope. Methods 2015, 88, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Stimper, V.; Bauer, S.; Ernstorfer, R.; Schölkopf, B.; Xian, R.P. Multidimensional Contrast Limited Adaptive Histogram Equalization. IEEE Access 2019, 7, 165437–165447. [Google Scholar] [CrossRef]
- Schmidt, U.; Weigert, M.; Broaddus, C.; Myers, G. Cell Detection with Star-Convex Polygons. In Lecture Notes in Computer Science, Proceedings of the Medical Image Computing and Computer Assisted Intervention—MICCAI 2018, Granada, Spain, 16–20 September 2018; Frangi, A., Schnabel, J., Davatzikos, C., Alberola-López, C., Fichtinger, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; Volume 11071, pp. 265–273. [Google Scholar]
- Rizk, A.; Paul, G.; Incardona, P.; Bugarski, M.; Mansouri, M.; Niemann, A.; Ziegler, U.; Berger, P.; Sbalzarini, I.F. Segmentation and quantification of subcellular structures in fluorescence microscopy images using Squassh. Nat. Protoc. 2014, 9, 586–596. [Google Scholar] [CrossRef] [PubMed]
- Paul, G.; Cardinale, J.; Sbalzarini, I.F. Coupling Image Restoration and Segmentation: A Generalized Linear Model/Bregman Perspective. Int. J. Comput. Vis. 2013, 104, 69–93. [Google Scholar] [CrossRef]
- Helmuth, J.A.; Sbalzarini, I.F. Deconvolving active contours for fluorescence microscopy images. In Advances in Visual Computing, Proceedings of the International Conference on Medical Image Computing and Computer-Assisted Intervention, Las Vegas, NV, USA, 30 November–2 December 2009; Springer: Berlin/Heidelberg, Germany, 2009; pp. 544–551. [Google Scholar]
- Helmuth, J.A.; Paul, G.; Sbalzarini, I.F. Beyond co-localization: Inferring spatial interactions between sub-cellular structures from microscopy images. BMC Bioinform. 2010, 11, 372. [Google Scholar] [CrossRef] [PubMed]
- Roerdink, J.B.; Meijster, A. The watershed transform: Definitions, algorithms and parallelization strategies. Fundam. Informaticae 2000, 41, 187–228. [Google Scholar] [CrossRef]
- Fill, M.; Gillespie, D. Simulating cardiac Ca2+ release units: Effects of RyR cluster size and Ca2+ buffers on diastolic Ca2+ leak. Pflug. Arch. 2021, 473, 435–446. [Google Scholar] [CrossRef]
- Xie, Y.; Yang, Y.; Galice, S.; Bers, D.M.; Sato, D. Size Matters: Ryanodine Receptor Cluster Size Heterogeneity Potentiates Calcium Waves. Biophys. J. 2019, 116, 530–539. [Google Scholar] [CrossRef]
- Veron, G.; Maltsev, V.A.; Stern, M.D.; Maltsev, A.V. Elementary intracellular Ca signals approximated as a transition of release channel system from a metastable state. J. Appl. Phys. 2023, 134, 124701. [Google Scholar] [CrossRef]
- Scriven, D.R.L.; Johnsen, A.B.; Asghari, P.; Chou, K.C.; Moore, E.D.W. Cardiomyocyte ryanodine receptor clusters expand and coalesce after application of isoproterenol. J. Gen. Physiol. 2023, 155, e202213109. [Google Scholar] [CrossRef]
- Greiser, M.; Joca, H.C.; Lederer, W.J. Pacemaker organization at the nanoscale: Imaging of ryanodine receptors as clusters in single sinoatrial nodal cells. Biophys. J. 2019, 116, 380A. [Google Scholar] [CrossRef]
- Yasin, S.; Kaplan, A.D.; Joca, H.C.; Lederer, W.J.; Greiser, M. ‘Calcium Clock’ at the Nanoscale in the RAT SA Node: 3D Ryanodine Receptor Cluster Organization and Intracellular Ca2+ Signaling. Biophys. J. 2020, 118, 173A. [Google Scholar] [CrossRef]
- Jensen, E.; Crossman, D.J. Technical review: Types of imaging-direct STORM. Anat. Rec. 2014, 297, 2227–2231. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Gopireddy, R.R.; Perkins, G.; Zhang, H.; Timofeyev, V.; Lyu, Y.; Diloretto, D.A.; Trinh, P.; Sirish, P.; Overton, J.L.; et al. Disruption of mitochondria-sarcoplasmic reticulum microdomain connectomics contributes to sinus node dysfunction in heart failure. Proc. Natl. Acad. Sci. USA 2022, 119, e2206708119. [Google Scholar] [CrossRef] [PubMed]
- Louradour, J.; Bortolotti, O.; Torre, E.; Bidaud, I.; Lamb, N.; Fernandez, A.; Le Guennec, J.Y.; Mangoni, M.E.; Mesirca, P. L-Type Cav1.3 Calcium Channels Are Required for Beta-Adrenergic Triggered Automaticity in Dormant Mouse Sinoatrial Pacemaker Cells. Cells 2022, 11, 1114. [Google Scholar] [CrossRef] [PubMed]
- Torrente, A.G.; Mesirca, P.; Neco, P.; Rizzetto, R.; Dubel, S.; Barrere, C.; Sinegger-Brauns, M.; Striessnig, J.; Richard, S.; Nargeot, J.; et al. L-type Cav1.3 channels regulate ryanodine receptor-dependent Ca2+ release during sino-atrial node pacemaker activity. Cardiovasc. Res. 2016, 109, 451–461. [Google Scholar] [CrossRef]
- Mesirca, P.; Torrente, A.G.; Mangoni, M.E. Functional role of voltage gated Ca2+ channels in heart automaticity. Front. Physiol. 2015, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Masumiya, H.; Yamamoto, H.; Hemberger, M.; Tanaka, H.; Shigenobu, K.; Chen, S.R.; Furukawa, T. The mouse sino-atrial node expresses both the type 2 and type 3 Ca2+ release channels/ryanodine receptors. FEBS Lett. 2003, 553, 141–144. [Google Scholar] [CrossRef]
- Tellez, J.O.; Dobrzynski, H.; Greener, I.D.; Graham, G.M.; Laing, E.; Honjo, H.; Hubbard, S.J.; Boyett, M.R.; Billeter, R. Differential expression of ion channel transcripts in atrial muscle and sinoatrial node in rabbit. Circ. Res. 2006, 99, 1384–1393. [Google Scholar] [CrossRef]
- Cingolani, E.; Goldhaber, J.I.; Marban, E. Next-generation pacemakers: From small devices to biological pacemakers. Nat. Rev. Cardiol. 2018, 15, 139–150. [Google Scholar] [CrossRef]
- Qu, Z.; Garfinkel, A.; Weiss, J.N.; Nivala, M. Multi-scale modeling in biology: How to bridge the gaps between scales? Prog. Biophys. Mol. Biol. 2011, 107, 21–31. [Google Scholar] [CrossRef]
- Stern, M.D.; Rios, E.; Maltsev, V.A. Life and death of a cardiac calcium spark. J. Gen. Physiol. 2013, 142, 257–274. [Google Scholar] [CrossRef]
- Zhang, H.; Holden, A.V.; Kodama, I.; Honjo, H.; Lei, M.; Varghese, T.; Boyett, M.R. Mathematical models of action potentials in the periphery and center of the rabbit sinoatrial node. Am. J. Physiol. 2000, 279, H397–H421. [Google Scholar] [CrossRef] [PubMed]
- Shannon, T.R.; Wang, F.; Puglisi, J.; Weber, C.; Bers, D.M. A mathematical treatment of integrated Ca dynamics within the ventricular myocyte. Biophys. J. 2004, 87, 3351–3371. [Google Scholar] [CrossRef]
- Kurata, Y.; Hisatome, I.; Imanishi, S.; Shibamoto, T. Dynamical description of sinoatrial node pacemaking: Improved mathematical model for primary pacemaker cell. Am. J. Physiol. 2002, 283, H2074–H2101. [Google Scholar] [CrossRef]
- Zima, A.V.; Bovo, E.; Bers, D.M.; Blatter, L.A. Ca2+ spark-dependent and -independent sarcoplasmic reticulum Ca2+ leak in normal and failing rabbit ventricular myocytes. J. Physiol. 2010, 588, 4743–4757. [Google Scholar] [CrossRef]
- Maltsev, A.V.; Maltsev, V.A.; Stern, M.D. Clusters of calcium release channels harness the Ising phase transition to confine their elementary intracellular signals. Proc. Natl. Acad. Sci. USA 2017, 114, 7525–7530. [Google Scholar] [CrossRef] [PubMed]
- Laver, D.R.; Kong, C.H.; Imtiaz, M.S.; Cannell, M.B. Termination of calcium-induced calcium release by induction decay: An emergent property of stochastic channel gating and molecular scale architecture. J. Mol. Cell. Cardiol. 2013, 54, 98–100. [Google Scholar] [CrossRef]
- Christel, C.J.; Cardona, N.; Mesirca, P.; Herrmann, S.; Hofmann, F.; Striessnig, J.; Ludwig, A.; Mangoni, M.E.; Lee, A. Distinct localization and modulation of Cav1.2 and Cav1.3 L-type Ca2+ channels in mouse sinoatrial node. J. Physiol. 2012, 590, 6327–6342. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.E.; Couette, B.; Bourinet, E.; Platzer, J.; Reimer, D.; Striessnig, J.; Nargeot, J. Functional role of L-type Cav1.3 Ca2+ channels in cardiac pacemaker activity. Proc. Natl. Acad. Sci. USA 2003, 100, 5543–5548. [Google Scholar] [CrossRef] [PubMed]
- Wilders, R.; Jongsma, H.J.; van Ginneken, A.C. Pacemaker activity of the rabbit sinoatrial node. A comparison of mathematical models. Biophys. J. 1991, 60, 1202–1216. [Google Scholar] [CrossRef]
- Dokos, S.; Celler, B.; Lovell, N. Ion currents underlying sinoatrial node pacemaker activity: A new single cell mathematical model. J. Theor. Biol. 1996, 181, 245–272. [Google Scholar] [CrossRef]
- Demir, S.S.; Clark, J.W.; Murphey, C.R.; Giles, W.R. A mathematical model of a rabbit sinoatrial node cell. Am. J. Physiol. 1994, 266, C832–C852. [Google Scholar] [CrossRef] [PubMed]
- Sakai, R.; Hagiwara, N.; Matsuda, N.; Kassanuki, H.; Hosoda, S. Sodium--potassium pump current in rabbit sino-atrial node cells. J. Physiol. 1996, 490 Pt 1, 51–62. [Google Scholar] [CrossRef] [PubMed]
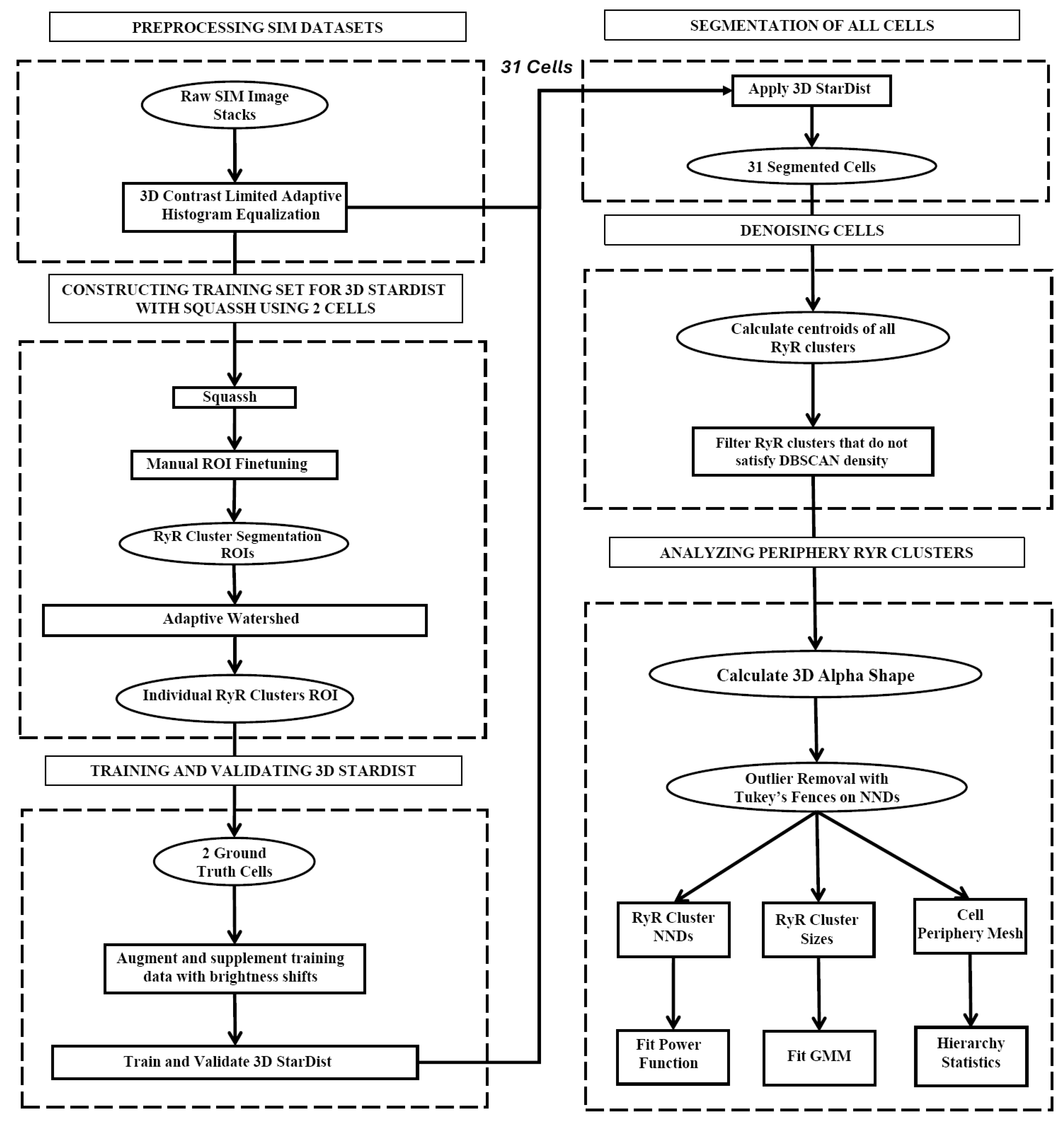

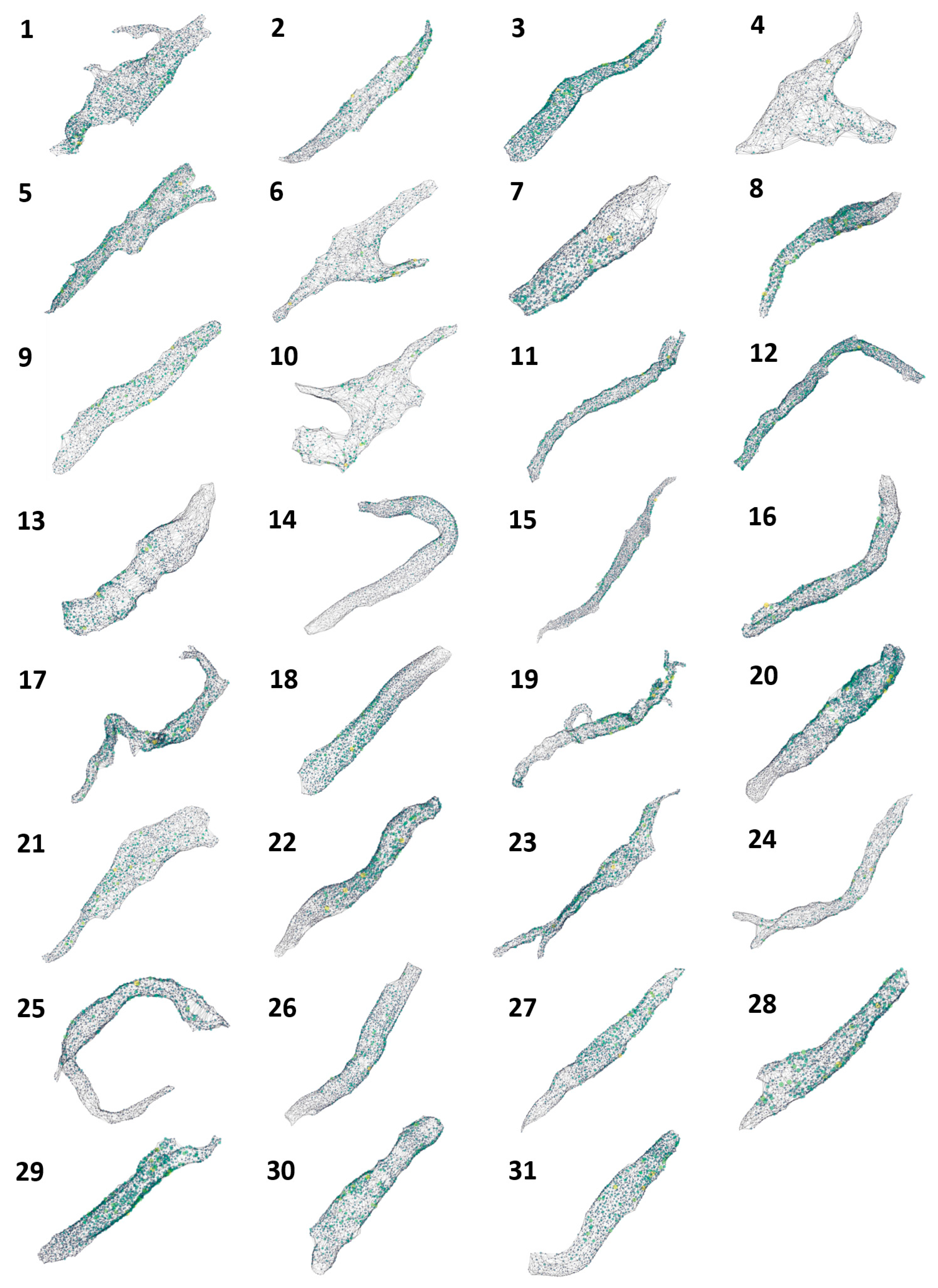
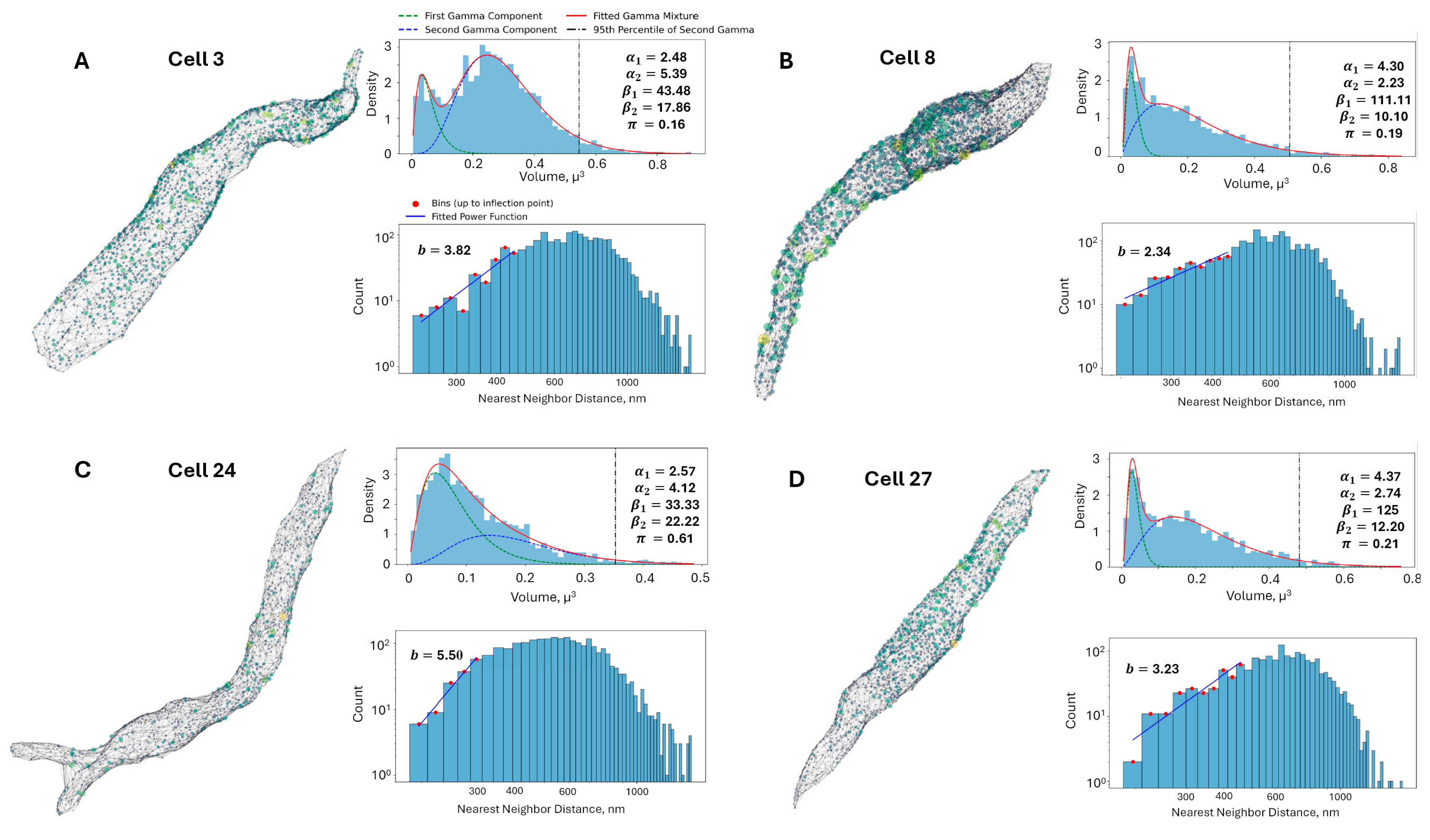
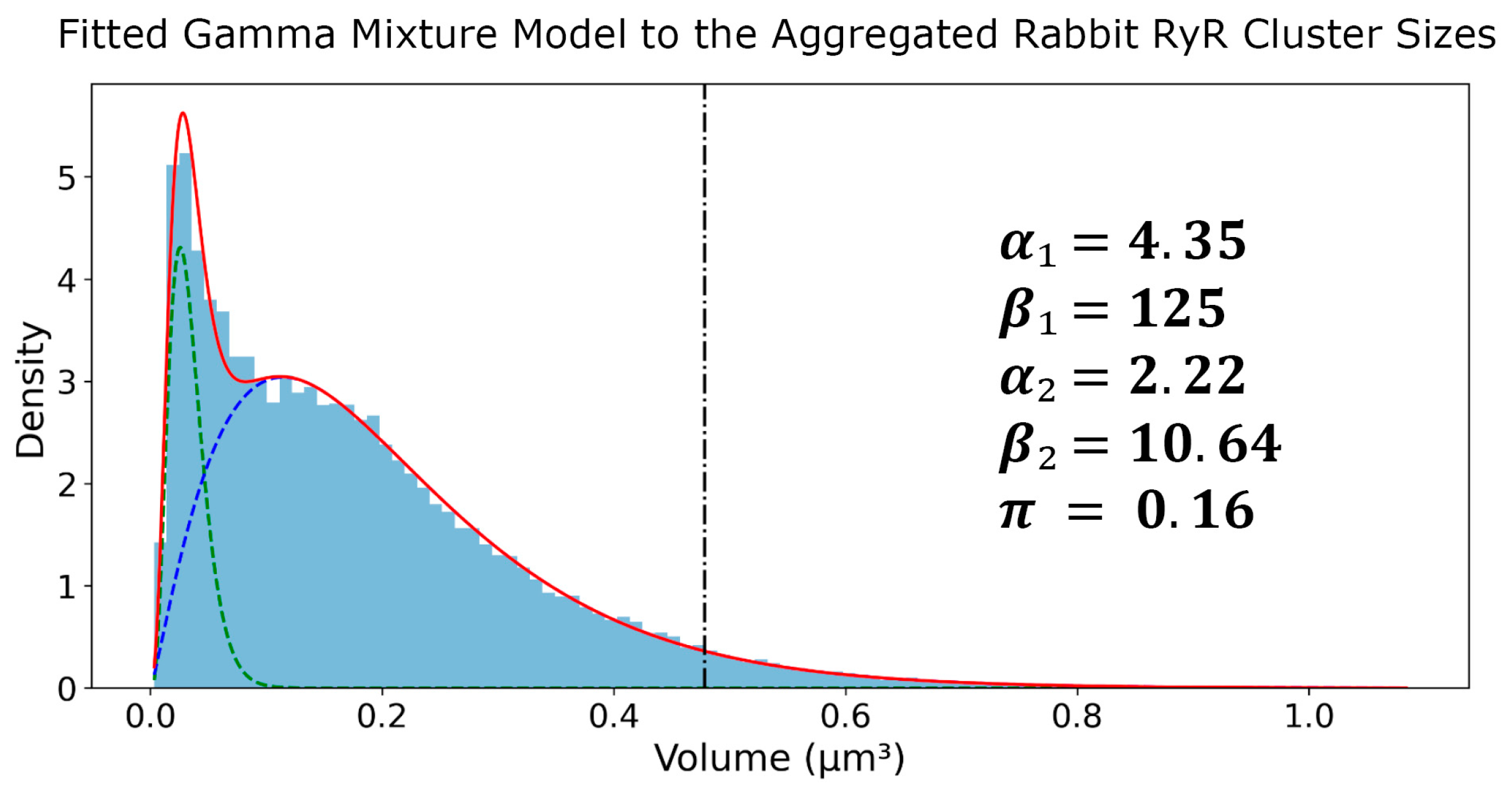
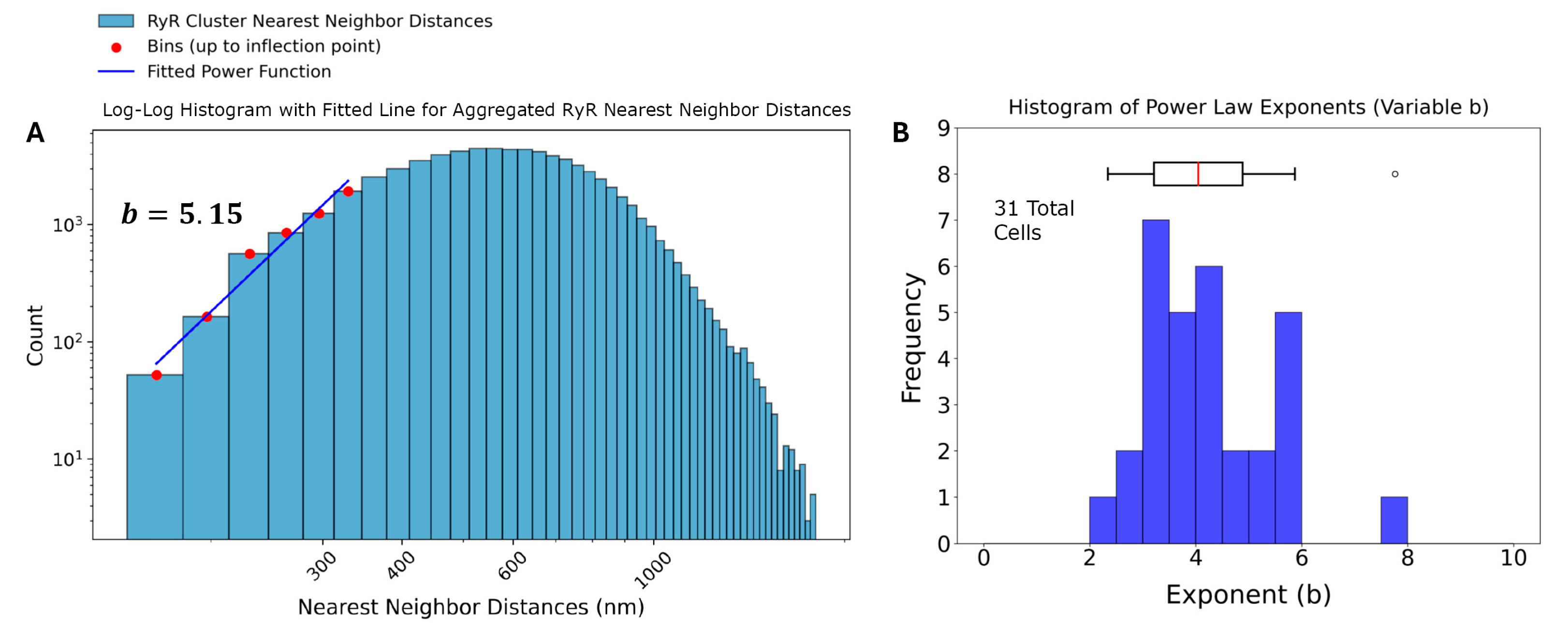
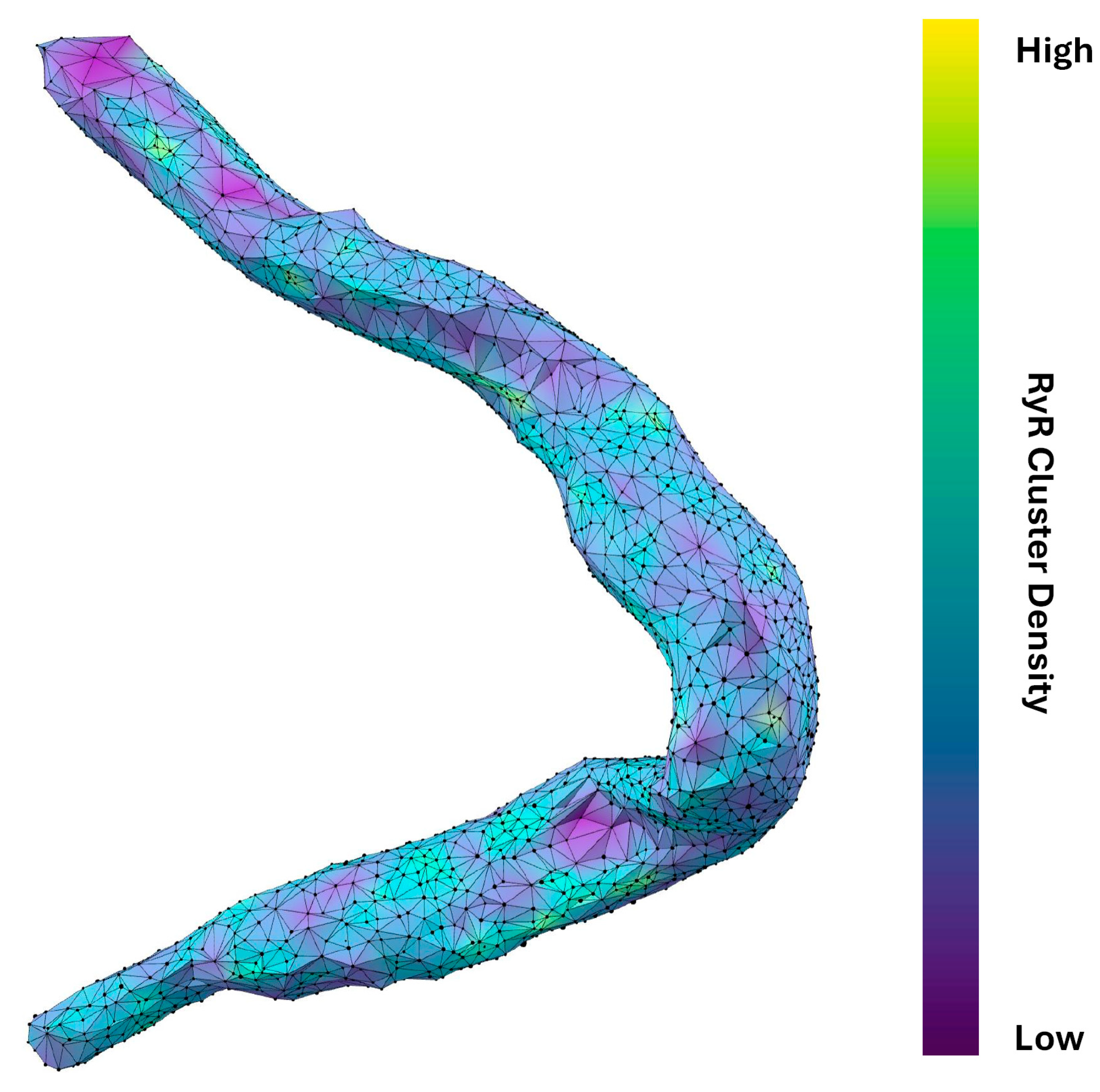
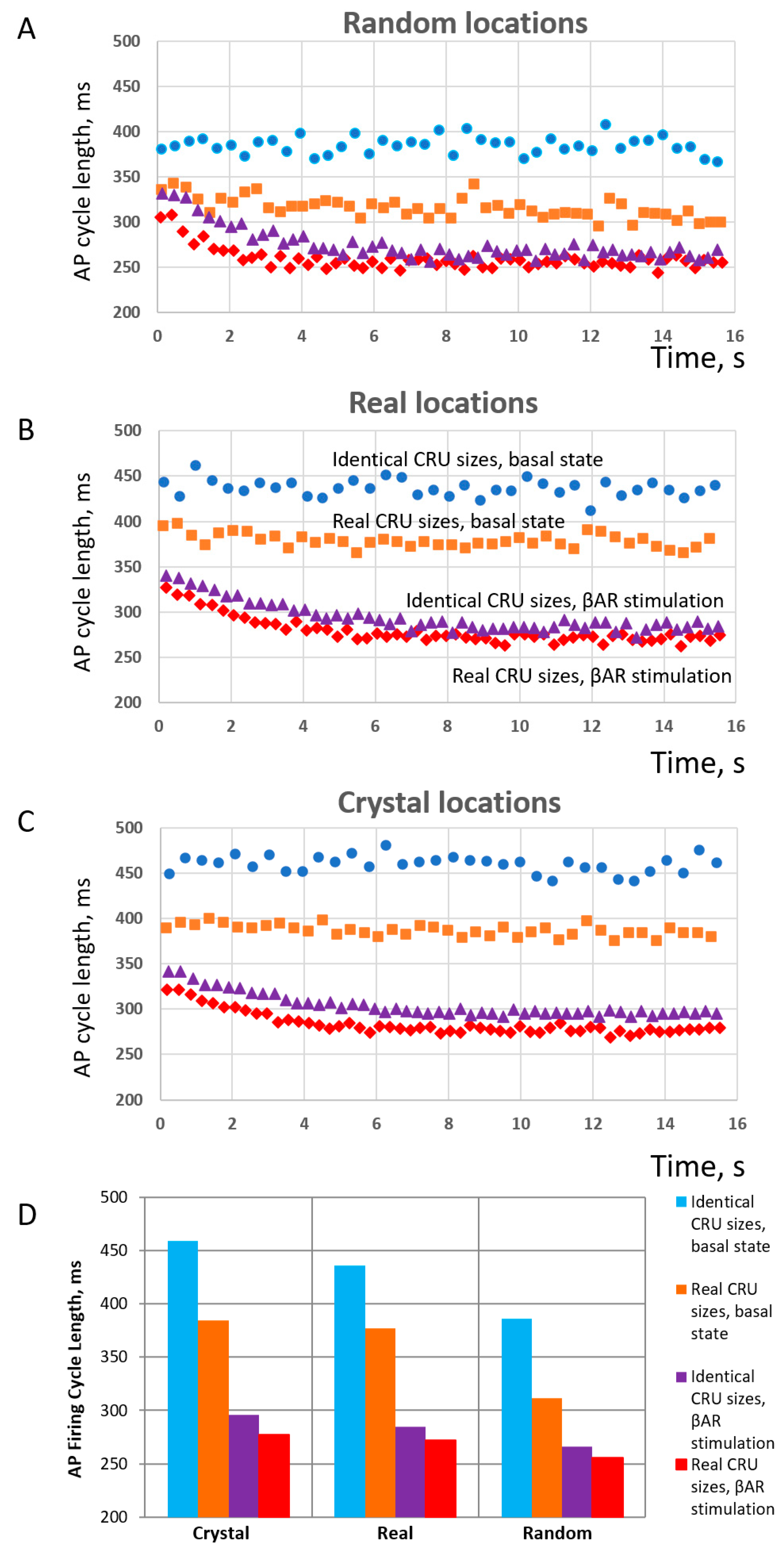
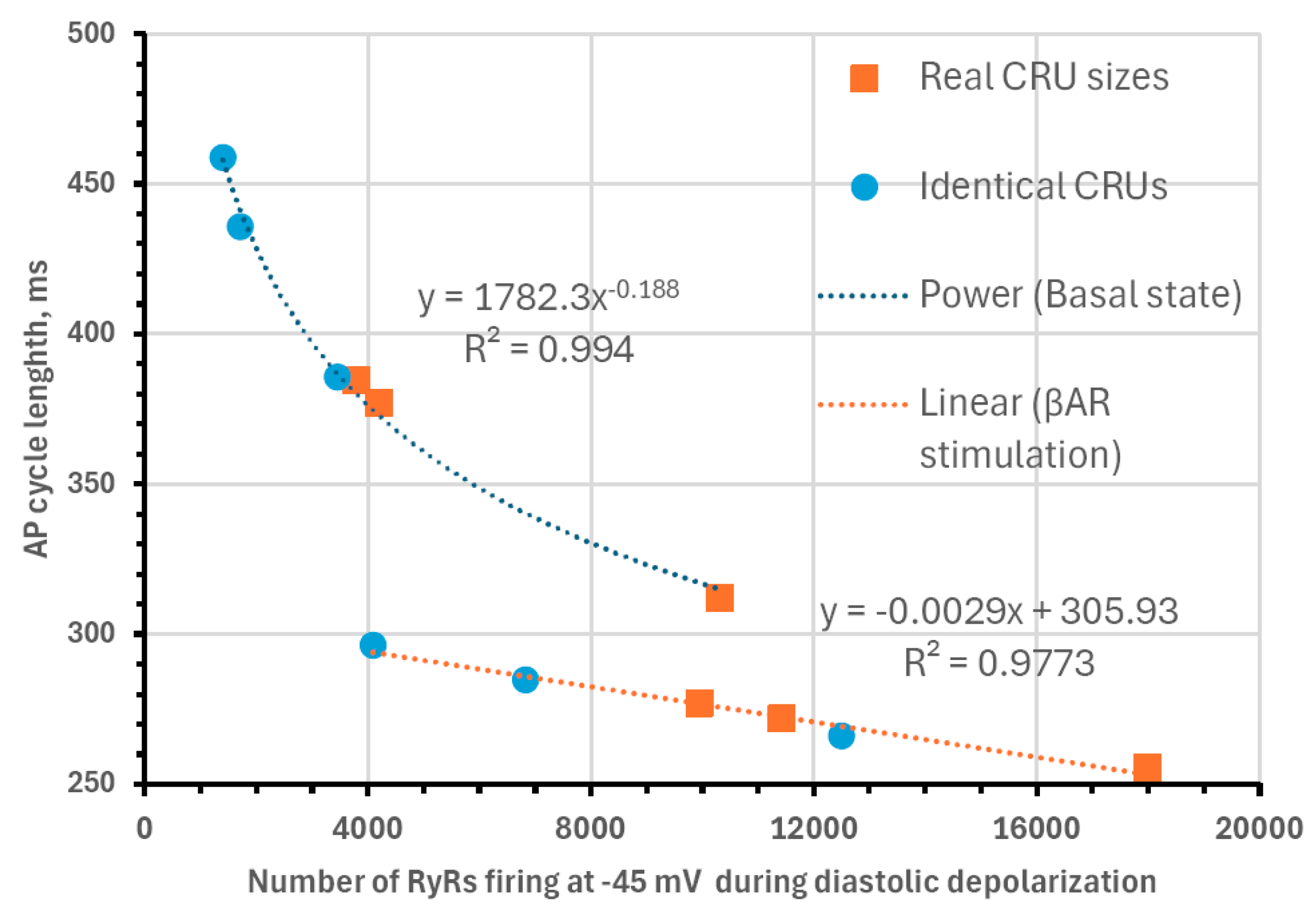
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).