
FAAH Inhibition Reverses Depressive-like Behavior and Sex-Specific Neuroinflammatory Alterations Induced by Early Life Stress


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Early Life Stress (ELS) Model
2.3. Pharmacological Treatment
2.4. Behavioral Tests
2.4.1. Activity in an Open-Field Test (OFT)
2.4.2. Social Preference (SP) and Social Recognition (SR)
2.4.3. Forced Swim Test (FST)
2.4.4. Real-Time (RT) PCR
2.4.5. Statistical Analysis
3. Results
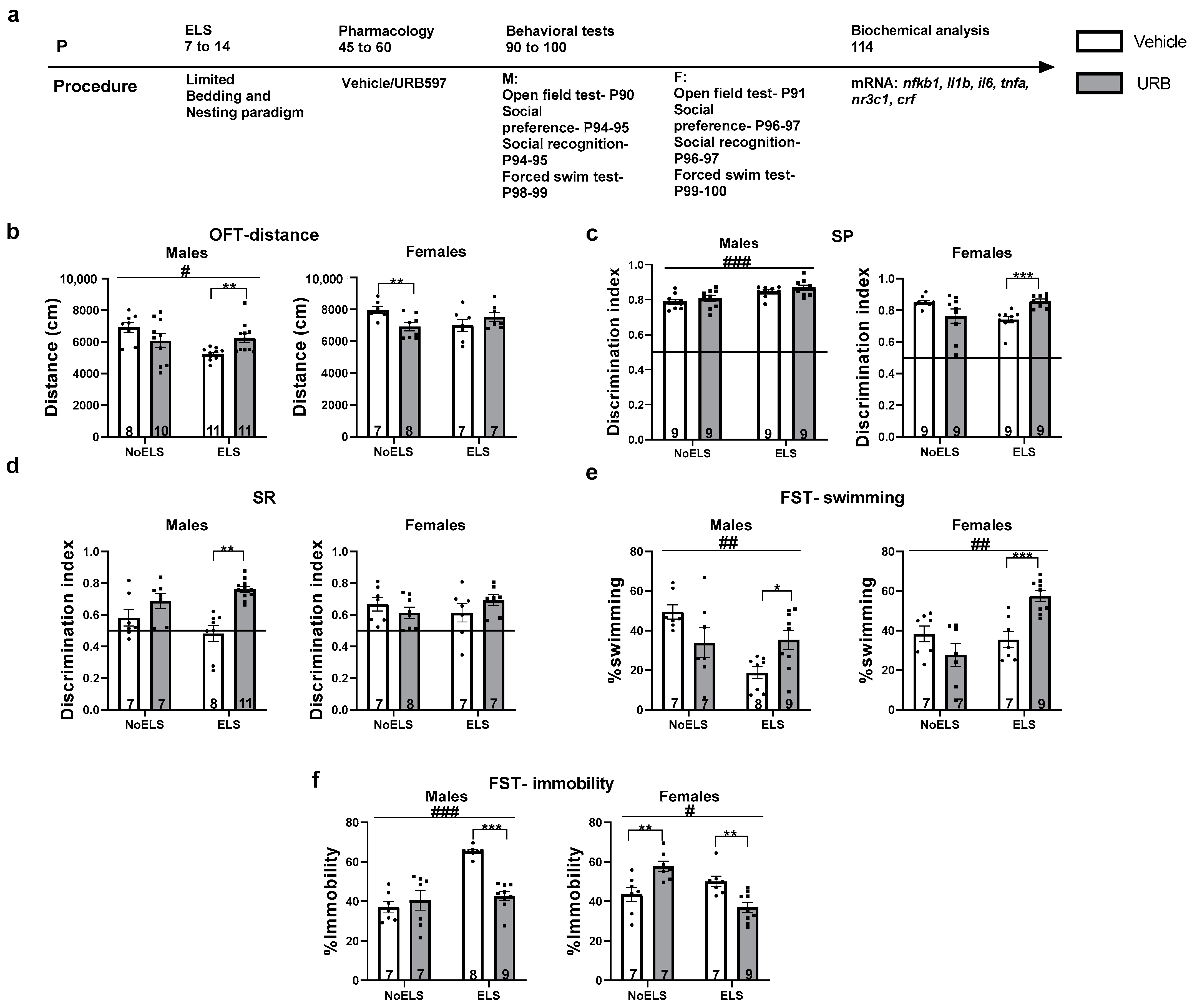
3.1. Experiment 1: Chronic Late Adolescence URB597 Treatment Ameliorates ELS-Induced Depressive-like Phenotype and Modulates Changes in Gene Expression in Adult Male and Female Rats
3.1.1. Chronic Late Adolescence URB597 Treatment Ameliorates ELS-Induced Depressive-like Phenotype in Adult Males and Females
Distance Traveled in the Open Field (OFT)
Social Preference Test (SP)
Social Recognition Test (SR)
Swimming in the Forced Swim Test (FST)
Immobility in the FST
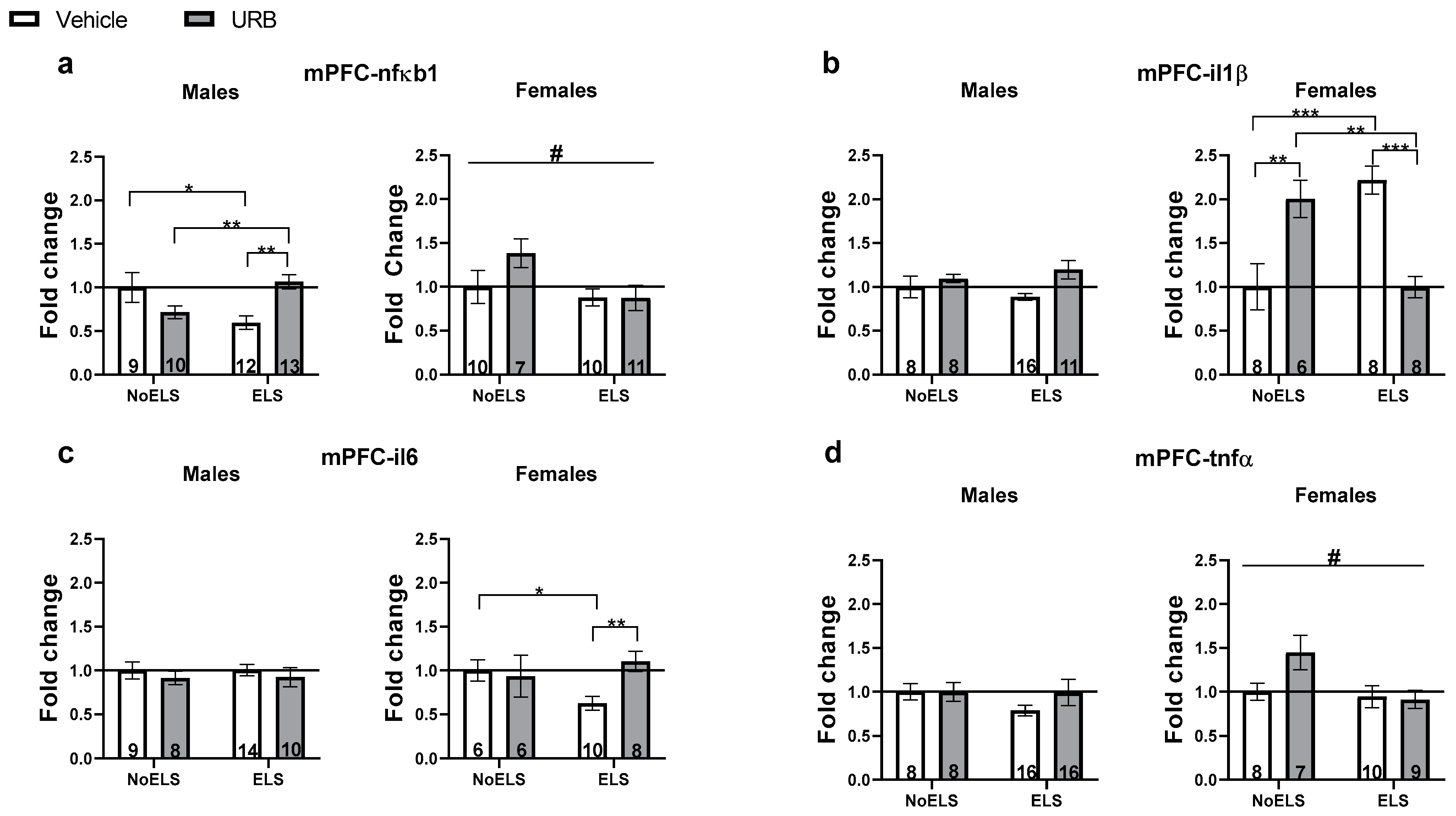
3.1.2. Chronic Late Adolescence URB597 Treatment Ameliorates ELS-Induced Changes in Inflammatory Gene Expression in the mPFC
nfκb1
il1β
il6
tnfα
Correlations Between the Expression of Inflammatory Genes in the mPFC and Behavioral Responses in Adult Males and Females
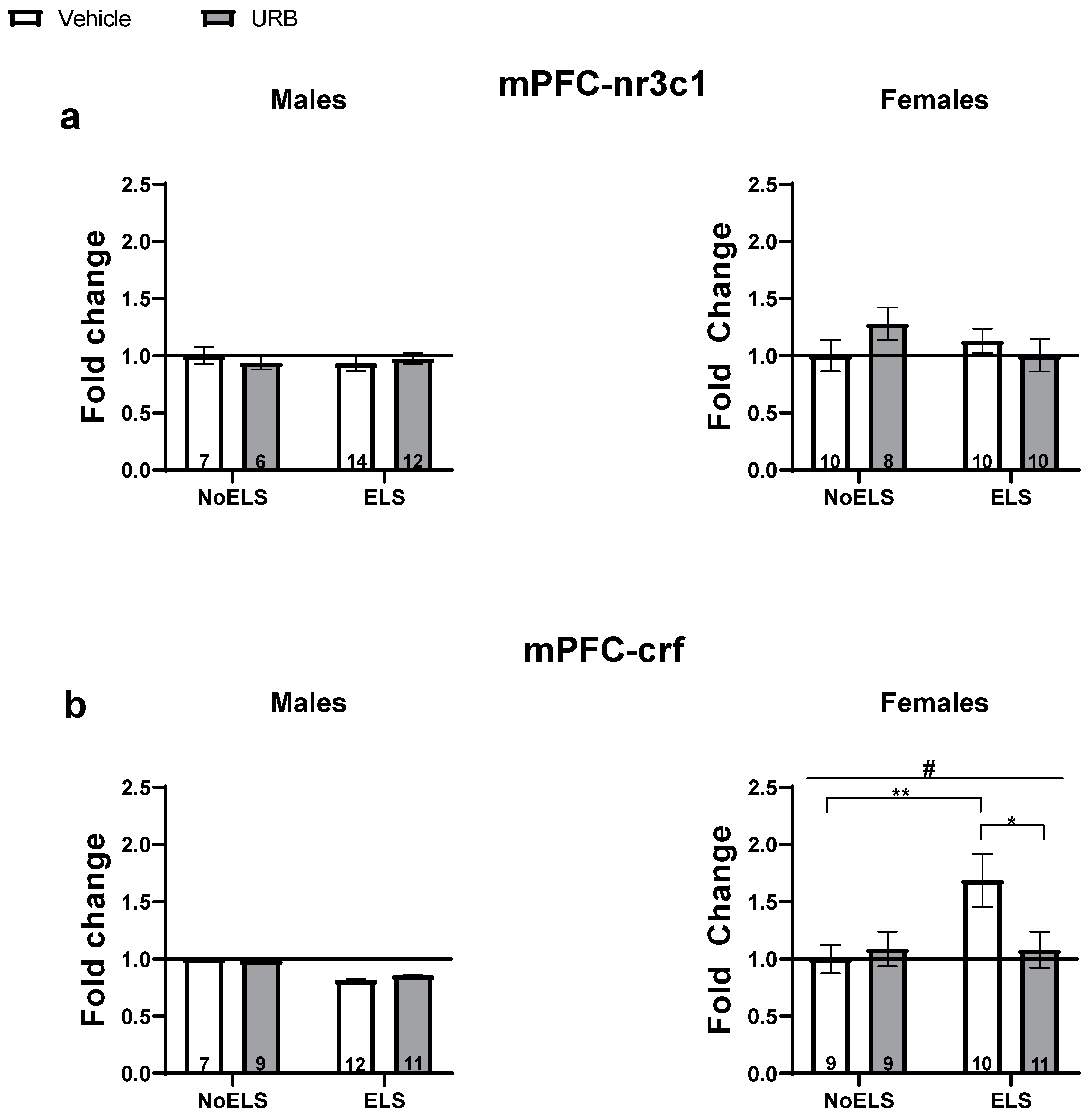
3.1.3. Chronic Late Adolescence URB597 Treatment Ameliorates ELS-Induced Changes in Stress-Related Gene Expression in the mPFC
nr3c1
crf
Correlations Between the Expression of Stress-Related Genes in the mPFC and Behavioral Responses in Adult Males and Females
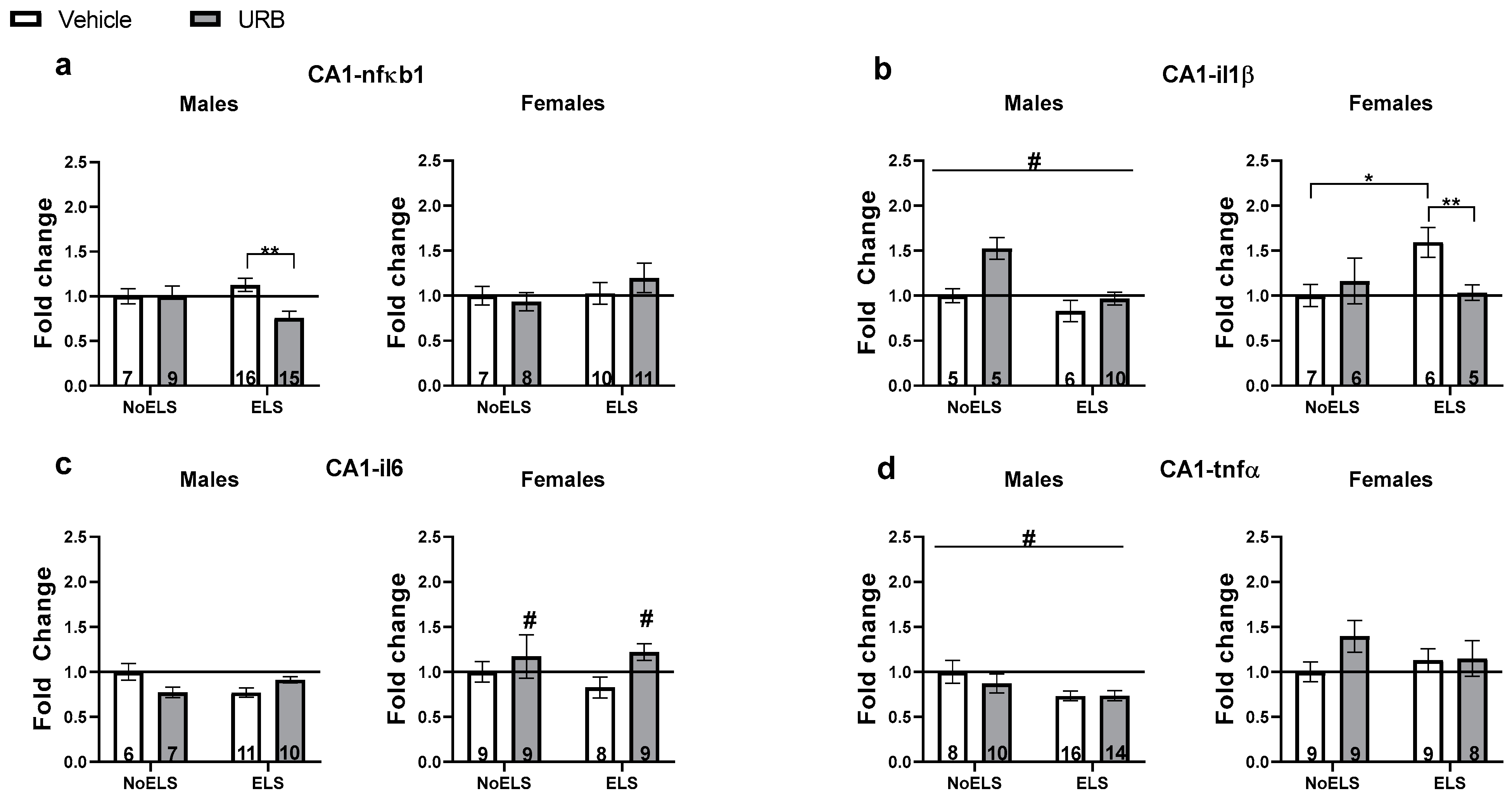
3.1.4. Chronic Late Adolescence URB597 Treatment Ameliorates ELS-Induced Changes in Inflammatory Genes’ Expression in the Hippocampal CA1
nfκb1
il1β
il6
tnfα
Correlations Between Inflammatory Gene Expression in the Hippocampal CA1 and Behavior in Adult Males and Females
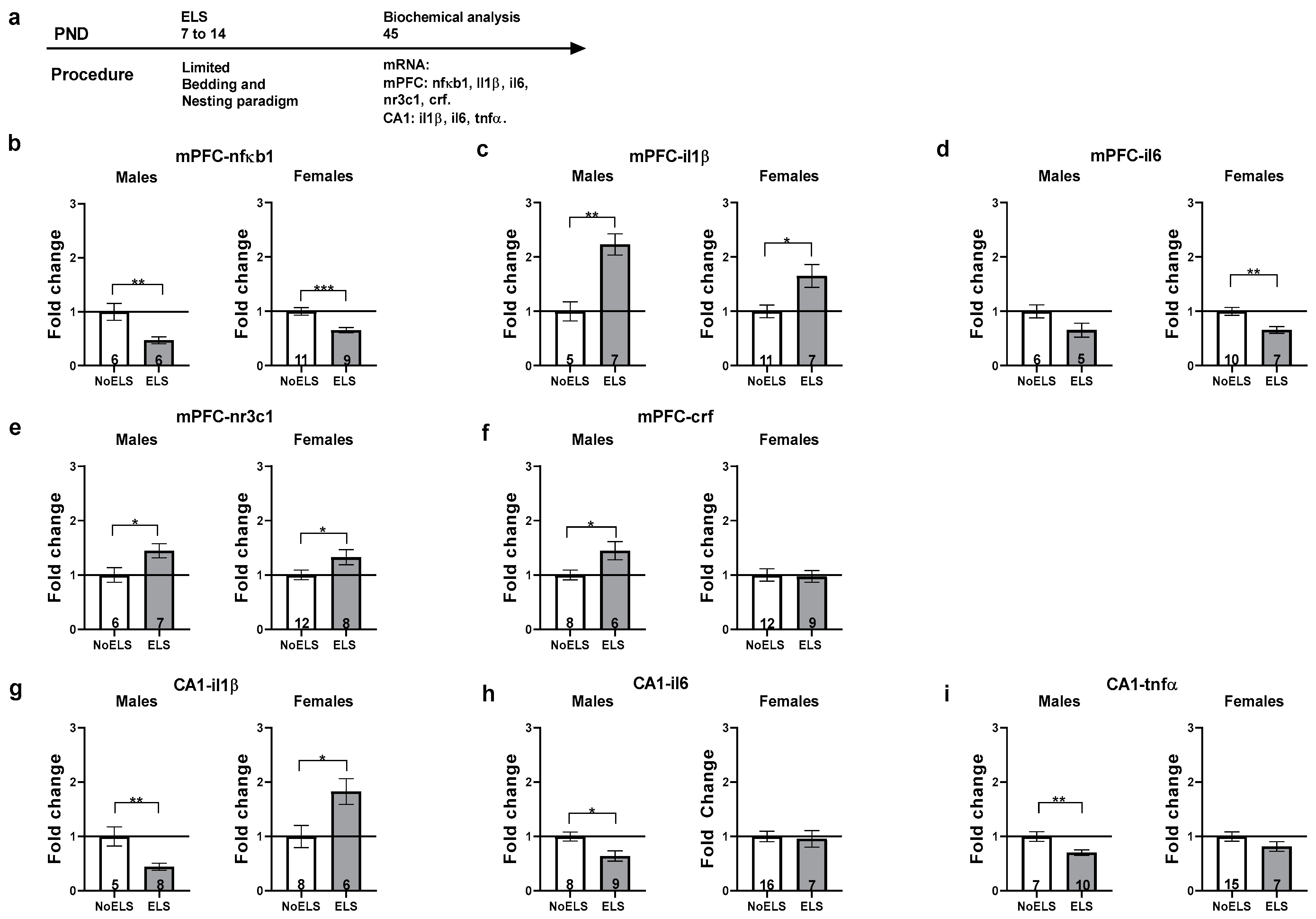
3.2. Experiment 2: ELS-Induced Alterations in Inflammatory and Stress-Related Genes in the mPFC and Hippocampal CA1 of Late Adolescent Rats, Prior to URB Treatment
3.2.1. mPFC-nfκb1
3.2.2. mPFC-il1β
3.2.3. mPFC-il6
3.2.4. mPFC-nr3c1
3.2.5. mPFC-crf
3.2.6. CA1-il1β
3.2.7. CA1-il6
3.2.8. CA1-tnfα
4. Discussion
4.1. Impact of ELS and URB597 Treatment on Behavioral Outcome in Adult Male and Female Rats
4.2. Impact of ELS and URB597 on Inflammatory and Stress-Related Gene Expression in the mPFC
4.3. Impact of ELS and URB597 on Inflammatory and Stress-Related Gene Expression in the CA1 Region
4.4. Sex Differences and ECS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Z.; Ruan, M.; Chen, J.; Fang, Y. Major Depressive Disorder: Advances in Neuroscience Research and Translational Applications. Neurosci. Bull. 2021, 37, 863–880. [Google Scholar] [CrossRef] [PubMed]
- Waters, R.C.; Gould, E. Early Life Adversity and Neuropsychiatric Disease: Differential Outcomes and Translational Relevance of Rodent Models. Front. Syst. Neurosci. 2022, 16, 860847. [Google Scholar] [CrossRef]
- Williams, L.M.; Debattista, C.; Duchemin, A.-M.; Schatzberg, A.F.; Nemeroff, C.B. Childhood Trauma Predicts Antidepressant Response in Adults with Major Depression: Data from the Randomized International Study to Predict Optimized Treatment for Depression. Transl. Psychiatry 2016, 6, e799. [Google Scholar] [CrossRef]
- Bertollo, A.G.; Galvan, A.C.L.; Dallagnol, C.; Cortez, A.D.; Ignácio, Z.M. Early Life Stress and Major Depressive Disorder—An Update on Molecular Mechanisms and Synaptic Impairments. Mol. Neurobiol. 2024, 61, 6469–6483. [Google Scholar] [CrossRef] [PubMed]
- Blaisdell, K.N.; Imhof, A.M.; Fisher, P.A. Early Adversity, Child Neglect, and Stress Neurobiology: From Observations of Impact to Empirical Evaluations of Mechanisms. Int. J. Dev. Neurosci. 2019, 78, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Réus, G.Z.; Fernandes, G.C.; de Moura, A.B.; Silva, R.H.; Darabas, A.C.; de Souza, T.G.; Abelaira, H.M.; Carneiro, C.; Wendhausen, D.; Michels, M. Early Life Experience Contributes to the Developmental Programming of Depressive-like Behaviour, Neuroinflammation and Oxidative Stress. J. Psychiatr. Res. 2017, 95, 196–207. [Google Scholar] [CrossRef]
- Smith, K.E.; Pollak, S.D. Early Life Stress and Development: Potential Mechanisms for Adverse Outcomes. J. Neurodev. Disord. 2020, 12, 34. [Google Scholar] [CrossRef]
- Speranza, L.; Filiz, K.D.; Lippiello, P.; Ferraro, M.G.; Pascarella, S.; Miniaci, M.C.; Volpicelli, F. Enduring Neurobiological Consequences of Early-Life Stress: Insights from Rodent Behavioral Paradigms. Biomedicines 2024, 12, 1978. [Google Scholar] [CrossRef]
- Gunn, B.G.; Baram, T.Z. Stress and Seizures: Space, Time and Hippocampal Circuits. Trends Neurosci. 2017, 40, 667–679. [Google Scholar] [CrossRef]
- Rahman, M.F.; McGowan, P.O. Cell-Type-Specific Epigenetic Effects of Early Life Stress on the Brain. Transl. Psychiatry 2022, 12, 326. [Google Scholar] [CrossRef]
- Danielewicz, J.; Hess, G. Early Life Stress Alters Synaptic Modification Range in the Rat Lateral Amygdala. Behav. Brain Res. 2014, 265, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Callaghan, B.L.; Richardson, R. Maternal Separation Results in Early Emergence of Adult-like Fear and Extinction Learning in Infant Rats. Behav. Neurosci. 2011, 125, 20. [Google Scholar] [CrossRef] [PubMed]
- Ivy, A.S.; Rex, C.S.; Chen, Y.; Dubé, C.; Maras, P.M.; Grigoriadis, D.E.; Gall, C.M.; Lynch, G.; Baram, T.Z. Hippocampal Dysfunction and Cognitive Impairments Provoked by Chronic Early-Life Stress Involve Excessive Activation of CRH Receptors. J. Neurosci. 2010, 30, 13005–13015. [Google Scholar] [CrossRef]
- Anilkumar, S.; Wright-Jin, E. NF-ΚB as an Inducible Regulator of Inflammation in the Central Nervous System. Cells 2024, 13, 485. [Google Scholar] [CrossRef]
- Kuhlman, K.R.; Robles, T.F.; Haydon, M.D.; Dooley, L.; Boyle, C.C.; Bower, J.E. Early Life Stress Sensitizes Individuals to the Psychological Correlates of Mild Fluctuations in Inflammation. Dev. Psychobiol. 2020, 62, 400–408. [Google Scholar] [CrossRef]
- Silva, R.C.; Maffioletti, E.; Gennarelli, M.; Baune, B.T.; Minelli, A. Biological Correlates of Early Life Stressful Events in Major Depressive Disorder. Psychoneuroendocrinology 2021, 125, 105103. [Google Scholar] [CrossRef]
- Lumertz, F.S.; Kestering-Ferreira, E.; Orso, R.; Creutzberg, K.C.; Tractenberg, S.G.; Stocchero, B.A.; Viola, T.W.; Grassi-Oliveira, R. Effects of Early Life Stress on Brain Cytokines: A Systematic Review and Meta-Analysis of Rodent Studies. Neurosci. Biobehav. Rev. 2022, 139, 104746. [Google Scholar] [CrossRef] [PubMed]
- Dutcher, E.G.; Pama, E.A.C.; Lynall, M.-E.; Khan, S.; Clatworthy, M.R.; Robbins, T.W.; Bullmore, E.T.; Dalley, J.W. Early-Life Stress and Inflammation: A Systematic Review of a Key Experimental Approach in Rodents. Brain Neurosci. Adv. 2020, 4, 2398212820978049. [Google Scholar] [CrossRef] [PubMed]
- Troubat, R.; Barone, P.; Leman, S.; Desmidt, T.; Cressant, A.; Atanasova, B.; Brizard, B.; El Hage, W.; Surget, A.; Belzung, C. Neuroinflammation and Depression: A Review. Eur. J. Neurosci. 2021, 53, 151–171. [Google Scholar] [CrossRef]
- Kasatkina, L.A.; Rittchen, S.; Sturm, E.M. Neuroprotective and Immunomodulatory Action of the Endocannabinoid System under Neuroinflammation. Int. J. Mol. Sci. 2021, 22, 5431. [Google Scholar] [CrossRef]
- Rathod, S.S.; Agrawal, Y.O.; Nakhate, K.T.; Meeran, M.F.N.; Ojha, S.; Goyal, S.N. Neuroinflammation in the Central Nervous System: Exploring the Evolving Influence of Endocannabinoid System. Biomedicines 2023, 11, 2642. [Google Scholar] [CrossRef] [PubMed]
- Su, S.-H.; Wu, Y.-F.; Lin, Q.; Hai, J. Cannabinoid Receptor Agonist WIN55, 212–2 and Fatty Acid Amide Hydrolase Inhibitor URB597 Ameliorate Neuroinflammatory Responses in Chronic Cerebral Hypoperfusion Model by Blocking NF-ΚB Pathways. Naunyn. Schmiedebergs. Arch. Pharmacol. 2017, 390, 1189–1200. [Google Scholar] [CrossRef]
- Bellozi, P.M.Q.; Pelição, R.; Santos, M.C.; Lima, I.V.A.; Saliba, S.W.; Vieira, É.L.M.; Campos, A.C.; Teixeira, A.L.; de Oliveira, A.C.P.; Nakamura-Palacios, E.M.; et al. URB597 Ameliorates the Deleterious Effects Induced by Binge Alcohol Consumption in Adolescent Rats. Neurosci. Lett. 2019, 711, 134408. [Google Scholar] [CrossRef] [PubMed]
- Rivera, P.; del Mar Fernández-Arjona, M.; Silva-Peña, D.; Blanco, E.; Vargas, A.; López-Ávalos, M.D.; Grondona, J.M.; Serrano, A.; Pavón, F.J.; de Fonseca, F.R. Pharmacological Blockade of Fatty Acid Amide Hydrolase (FAAH) by URB597 Improves Memory and Changes the Phenotype of Hippocampal Microglia despite Ethanol Exposure. Biochem. Pharmacol. 2018, 157, 244–257. [Google Scholar] [CrossRef]
- Piomelli, D.; Tarzia, G.; Duranti, A.; Tontini, A.; Mor, M.; Compton, T.R.; Dasse, O.; Monaghan, E.P.; Parrott, J.A.; Putman, D. Pharmacological Profile of the Selective FAAH Inhibitor KDS-4103 (URB597). CNS Drug Rev. 2006, 12, 21–38. [Google Scholar] [CrossRef]
- McKinney, M.K.; Cravatt, B.F. Structure and Function of Fatty Acid Amide Hydrolase. Annu. Rev. Biochem. 2005, 74, 411–432. [Google Scholar] [CrossRef]
- Portugalov, A.; Zaidan, H.; Gaisler-Salomon, I.; Hillard, C.J.; Akirav, I. FAAH Inhibition Restores Early Life Stress-Induced Alterations in PFC MicroRNAs Associated with Depressive-like Behavior in Male and Female Rats. Int. J. Mol. Sci. 2022, 23, 16101. [Google Scholar] [CrossRef] [PubMed]
- Portugalov, A.; Akirav, I. Do Adolescent Exposure to Cannabinoids and Early Adverse Experience Interact to Increase the Risk of Psychiatric Disorders: Evidence from Rodent Models. Int. J. Mol. Sci. 2021, 22, 730. [Google Scholar] [CrossRef] [PubMed]
- De Bosscher, K.; Vanden Berghe, W.; Haegeman, G. The Interplay between the Glucocorticoid Receptor and Nuclear Factor-ΚB or Activator Protein-1: Molecular Mechanisms for Gene Repression. Endocr. Rev. 2003, 24, 488–522. [Google Scholar]
- Gilles, E.E.; Schultz, L.; Baram, T.Z. Abnormal Corticosterone Regulation in an Immature Rat Model of Continuous Chronic Stress. Pediatr. Neurol. 1996, 15, 114–119. [Google Scholar] [CrossRef]
- Brunson, K.L.; Kramár, E.; Lin, B.; Chen, Y.; Colgin, L.L.; Yanagihara, T.K.; Lynch, G.; Baram, T.Z. Mechanisms of Late-Onset Cognitive Decline after Early-Life Stress. J. Neurosci. 2005, 25, 9328–9338. [Google Scholar] [CrossRef] [PubMed]
- Ivy, A.S.; Brunson, K.L.; Sandman, C.; Baram, T.Z. Dysfunctional Nurturing Behavior in Rat Dams with Limited Access to Nesting Material: A Clinically Relevant Model for Early-Life Stress. Neuroscience 2008, 154, 1132–1142. [Google Scholar] [CrossRef] [PubMed]
- Alteba, S.; Korem, N.; Akirav, I. Cannabinoids Reverse the Effects of Early Stress on Neurocognitive Performance in Adulthood. Learn. Mem. 2016, 23, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Alteba, S.; Portugalov, A.; Hillard, C.J.; Akirav, I. Inhibition of Fatty Acid Amide Hydrolase (FAAH) during Adolescence and Exposure to Early Life Stress May Exacerbate Depression-like Behaviors in Male and Female Rats. Neuroscience 2021, 455, 89–106. [Google Scholar] [CrossRef] [PubMed]
- Segev, A.; Rubin, A.S.; Abush, H.; Richter-Levin, G.; Akirav, I. Cannabinoid Receptor Activation Prevents the Effects of Chronic Mild Stress on Emotional Learning and LTP in a Rat Model of Depression. Neuropsychopharmacology 2014, 39, 919–933. [Google Scholar] [CrossRef]
- Bauminger, H.; Zaidan, H.; Akirav, I.; Gaisler-Salomon, I. Anandamide Hydrolysis Inhibition Reverses the Long-Term Behavioral and Gene Expression Alterations Induced by MK-801 in Male Rats: Differential CB1 and CB2 Receptor-Mediated Effects. Schizophr. Bull. 2022, 48, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of Stable Housekeeping Genes, Differentially Regulated Target Genes and Sample Integrity: BestKeeper–Excel-Based Tool Using Pair-Wise Correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Alteba, S.; Zer-Aviv, T.M.; Tenenhaus, A.; David, G.B.; Adelman, J.; Hillard, C.J.; Doron, R.; Akirav, I. Antidepressant-like Effects of URB597 and JZL184 in Male and Female Rats Exposed to Early Life Stress. Eur. Neuropsychopharmacol. 2020, 39, 70–86. [Google Scholar] [CrossRef]
- Eid, R.S.; Gobinath, A.R.; Galea, L.A.M. Sex Differences in Depression: Insights from Clinical and Preclinical Studies. Prog. Neurobiol. 2019, 176, 86–102. [Google Scholar] [CrossRef]
- Viola, T.W.; Creutzberg, K.C.; Zaparte, A.; Kestering-Ferreira, É.; Tractenberg, S.G.; Centeno-Silva, A.; Orso, R.; Lumertz, F.S.; Brietzke, E.; Wearick-Silva, L.E. Acute Neuroinflammation Elicited by TLR-3 Systemic Activation Combined with Early Life Stress Induces Working Memory Impairments in Male Adolescent Mice. Behav. Brain Res. 2019, 376, 112221. [Google Scholar] [CrossRef]
- Tseng, A.H.; Craft, R.M. Sex Differences in Antinociceptive and Motoric Effects of Cannabinoids. Eur. J. Pharmacol. 2001, 430, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Craft, R.M.; Wakley, A.A.; Tsutsui, K.T.; Laggart, J.D. Sex Differences in Cannabinoid 1 vs. Cannabinoid 2 Receptor-Selective Antagonism of Antinociception Produced by Δ9-Tetrahydrocannabinol and CP55, 940 in the Rat. J. Pharmacol. Exp. Ther. 2012, 340, 787–800. [Google Scholar] [CrossRef] [PubMed]
- Burston, J.J.; Wiley, J.L.; Craig, A.A.; Selley, D.E.; Sim-Selley, L.J. Regional Enhancement of Cannabinoid CB1 Receptor Desensitization in Female Adolescent Rats Following Repeated Δ9-tetrahydrocannabinol Exposure. Br. J. Pharmacol. 2010, 161, 103–112. [Google Scholar] [CrossRef]
- Mateos, B.; Borcel, E.; Loriga, R.; Luesu, W.; Bini, V.; Llorente, R.; Castelli, M.P.; Viveros, M.P. Adolescent Exposure to Nicotine and/or the Cannabinoid Agonist CP 55,940 Induces Gender-Dependent Long-Lasting Memory Impairments and Changes in Brain Nicotinic and CB1 Cannabinoid Receptors. J. Psychopharmacol. 2011, 25, 1676–1690. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Signaling to NF-kappaB. Genes Dev. 2004, 18, 2195–2224. [Google Scholar] [CrossRef] [PubMed]
- Koo, J.W.; Russo, S.J.; Ferguson, D.; Nestler, E.J.; Duman, R.S. Nuclear Factor-ΚB Is a Critical Mediator of Stress-Impaired Neurogenesis and Depressive Behavior. Proc. Natl. Acad. Sci. USA 2010, 107, 2669–2674. [Google Scholar] [CrossRef]
- Hermoso, M.A.; Cidlowski, J.A. Putting the Brake on Inflammatory Responses: The Role of Glucocorticoids. IUBMB Life 2003, 55, 497–504. [Google Scholar] [CrossRef]
- Burke, N.N.; Llorente, R.; Marco, E.M.; Tong, K.; Finn, D.P.; Viveros, M.-P.; Roche, M. Maternal Deprivation Is Associated with Sex-Dependent Alterations in Nociceptive Behavior and Neuroinflammatory Mediators in the Rat Following Peripheral Nerve Injury. J. Pain 2013, 14, 1173–1184. [Google Scholar] [CrossRef]
- Wang, R.; Wang, W.; Xu, J.; Liu, D.; Wu, H.; Qin, X.; Jiang, H.; Pan, F. Jmjd3 Is Involved in the Susceptibility to Depression Induced by Maternal Separation via Enhancing the Neuroinflammation in the Prefrontal Cortex and Hippocampus of Male Rats. Exp. Neurol. 2020, 328, 113254. [Google Scholar] [CrossRef]
- Pryce, C.R.; Ruedi-Bettschen, D.; Dettling, A.C.; Feldon, J. Early Life Stress: Long-Term Physiological Impact in Rodents and Primates. Physiology 2002, 17, 150–155. [Google Scholar] [CrossRef]
- Jankovic, M.; Spasojevic, N.; Ferizovic, H.; Stefanovic, B.; Dronjak, S. Inhibition of the Fatty Acid Amide Hydrolase Changes Behaviors and Brain Catecholamines in a Sex-Specific Manner in Rats Exposed to Chronic Unpredictable Stress. Physiol. Behav. 2020, 227, 113174. [Google Scholar] [CrossRef]
- Kerr, D.M.; Burke, N.N.; Ford, G.K.; Connor, T.J.; Harhen, B.; Egan, L.J.; Finn, D.P.; Roche, M. Pharmacological Inhibition of Endocannabinoid Degradation Modulates the Expression of Inflammatory Mediators in the Hypothalamus Following an Immunological Stressor. Neuroscience 2012, 204, 53–63. [Google Scholar] [CrossRef]
- Hill, M.N.; McLaughlin, R.J.; Pan, B.; Fitzgerald, M.L.; Roberts, C.J.; Lee, T.T.-Y.; Karatsoreos, I.N.; Mackie, K.; Viau, V.; Pickel, V.M. Recruitment of Prefrontal Cortical Endocannabinoid Signaling by Glucocorticoids Contributes to Termination of the Stress Response. J. Neurosci. 2011, 31, 10506–10515. [Google Scholar] [CrossRef] [PubMed]
- Cota, D.; Steiner, M.-A.; Marsicano, G.; Cervino, C.; Herman, J.P.; Grubler, Y.; Stalla, J.; Pasquali, R.; Lutz, B.; Stalla, G.K. Requirement of Cannabinoid Receptor Type 1 for the Basal Modulation of Hypothalamic-Pituitary-Adrenal Axis Function. Endocrinology 2007, 148, 1574–1581. [Google Scholar] [CrossRef] [PubMed]
- Gorzalka, B.B.; Dang, S.S. Minireview: Endocannabinoids and Gonadal Hormones: Bidirectional Interactions in Physiology and Behavior. Endocrinology 2012, 153, 1016–1024. [Google Scholar] [CrossRef] [PubMed]
- Santoro, A.; Mele, E.; Marino, M.; Viggiano, A.; Nori, S.L.; Meccariello, R. The Complex Interplay between Endocannabinoid System and the Estrogen System in Central Nervous System and Periphery. Int. J. Mol. Sci. 2021, 22, 972. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Portugalov, A.; Akirav, I. FAAH Inhibition Reverses Depressive-like Behavior and Sex-Specific Neuroinflammatory Alterations Induced by Early Life Stress. Cells 2024, 13, 1881. https://doi.org/10.3390/cells13221881
Portugalov A, Akirav I. FAAH Inhibition Reverses Depressive-like Behavior and Sex-Specific Neuroinflammatory Alterations Induced by Early Life Stress. Cells. 2024; 13(22):1881. https://doi.org/10.3390/cells13221881
Chicago/Turabian StylePortugalov, Anna, and Irit Akirav. 2024. "FAAH Inhibition Reverses Depressive-like Behavior and Sex-Specific Neuroinflammatory Alterations Induced by Early Life Stress" Cells 13, no. 22: 1881. https://doi.org/10.3390/cells13221881
APA StylePortugalov, A., & Akirav, I. (2024). FAAH Inhibition Reverses Depressive-like Behavior and Sex-Specific Neuroinflammatory Alterations Induced by Early Life Stress. Cells, 13(22), 1881. https://doi.org/10.3390/cells13221881