
An Investigation of RNA Methylations with Biophysical Approaches in a Cervical Cancer Cell Model

 , , and
, , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Cell Culture, Flow Cytometry and Western Blotting
2.2. Cell Transfection
2.3. Total RNA Isolation and Global m6A Detection
2.4. SELECT (Single-Base Elongation- and Ligation-Based qPCR Amplification Method)
2.5. FT-IR Spectroscopy Measurements
2.6. CD Spectroscopy Measurements
3. Results
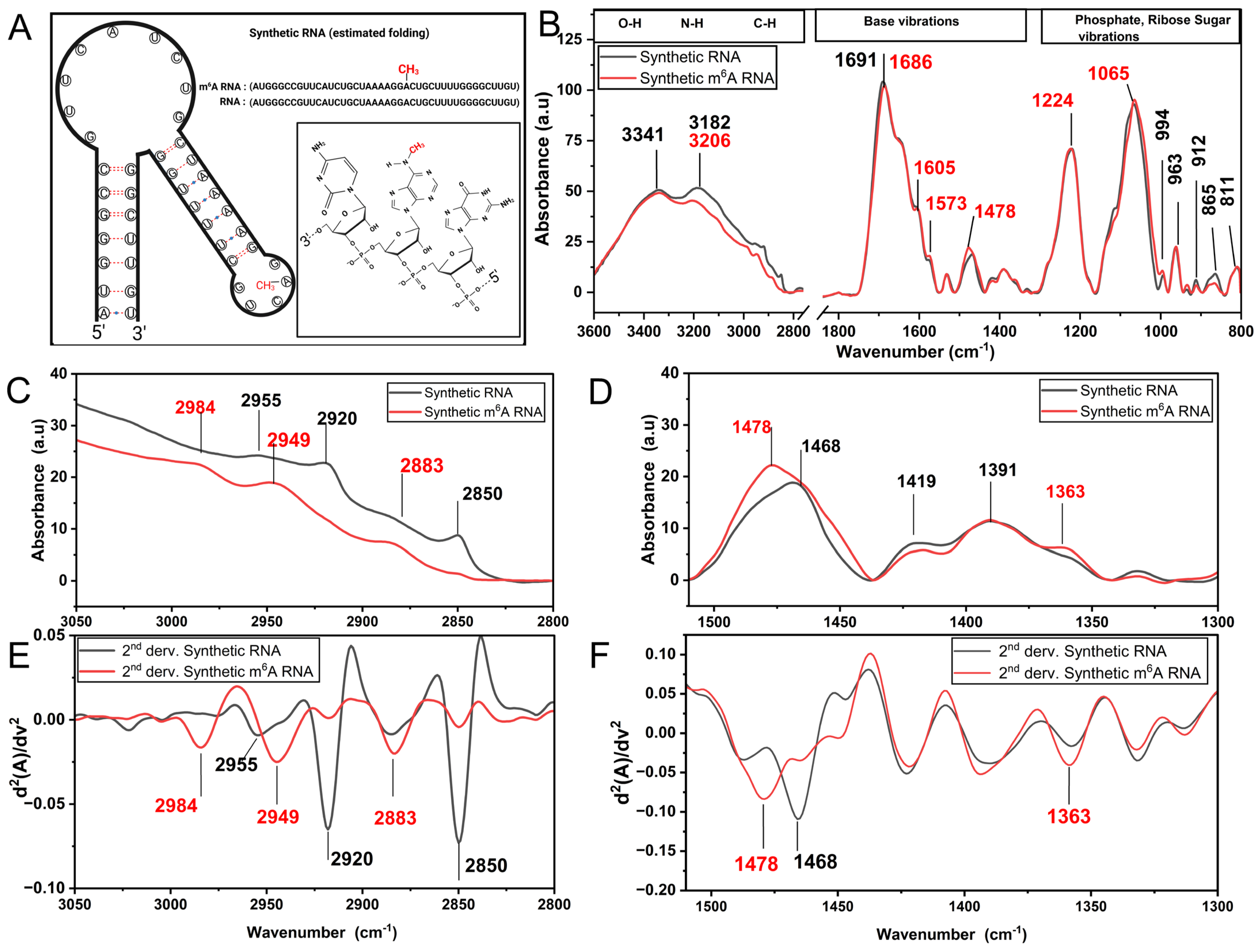
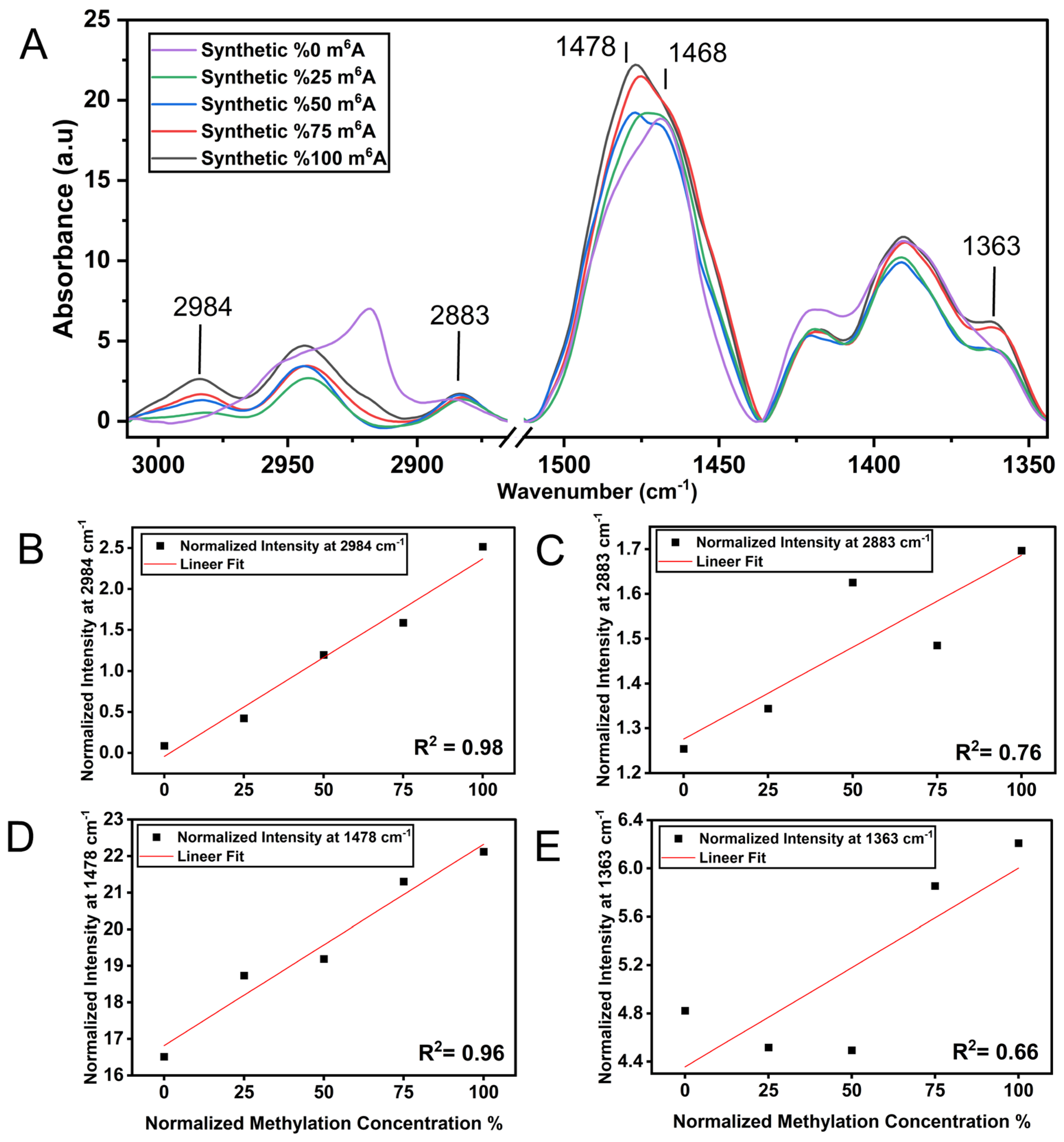
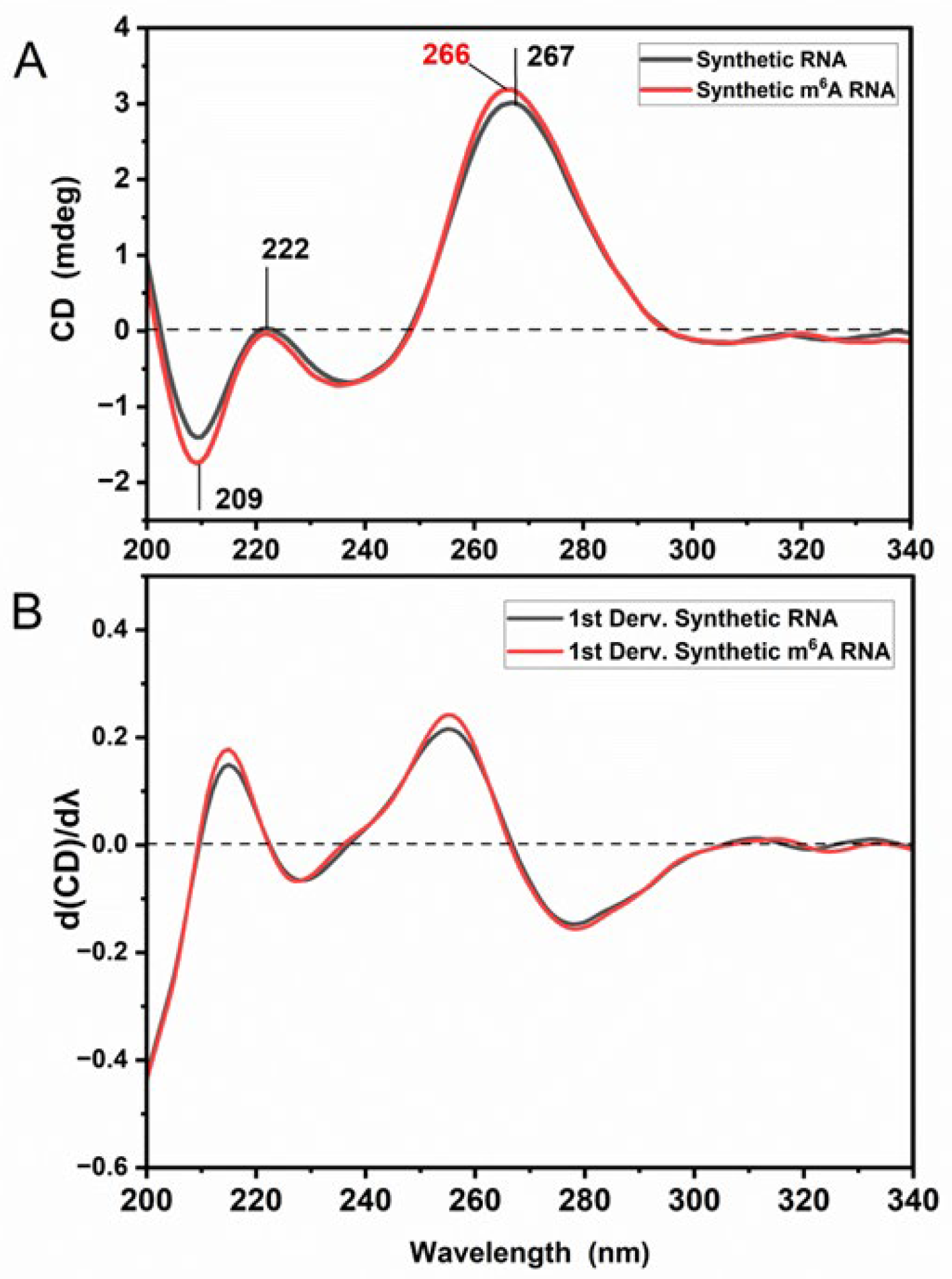
3.1. m6A Methylation Induces a Unique Spectroscopy Profile in Synthetic RNA Oligonucleotides
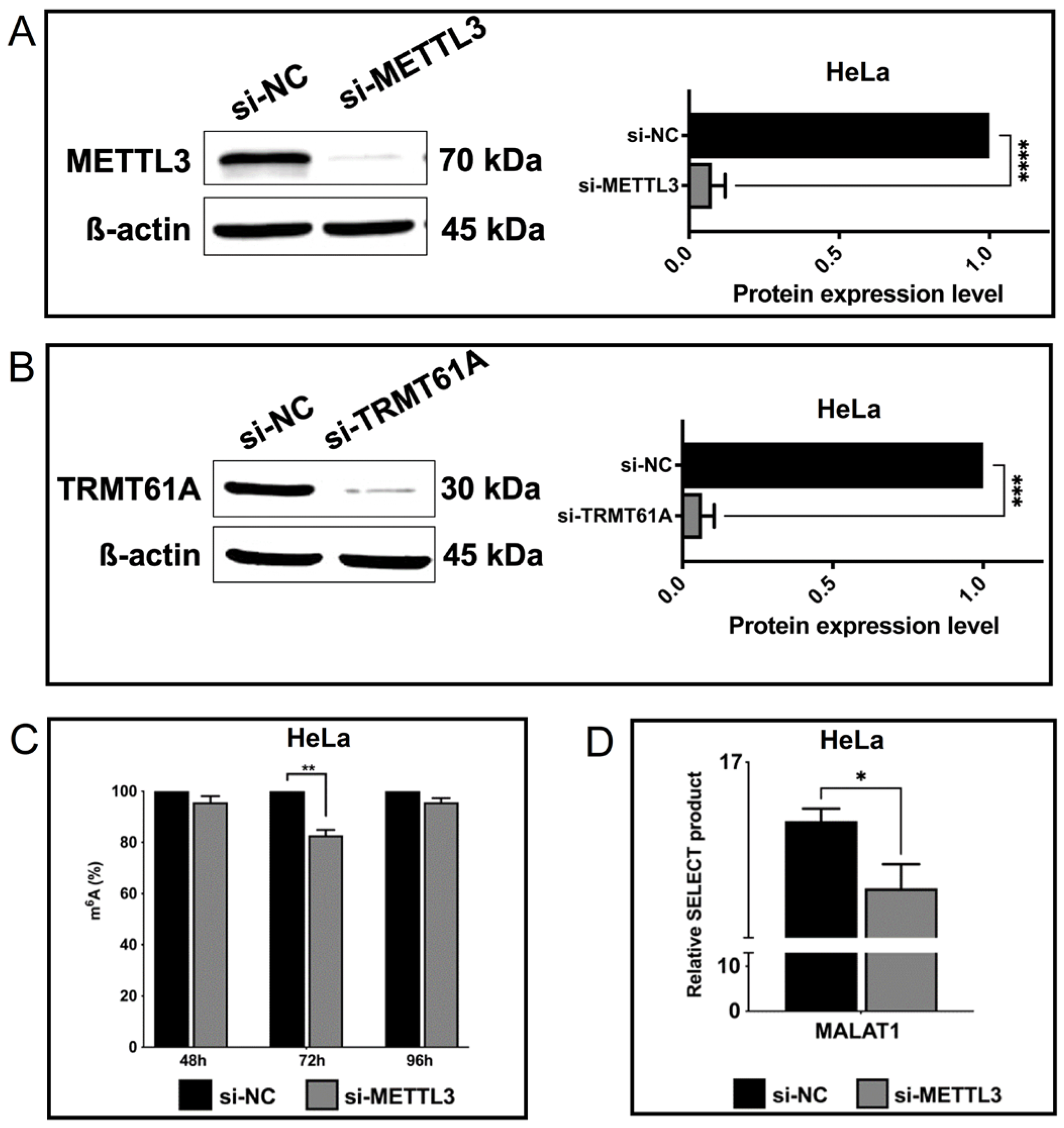
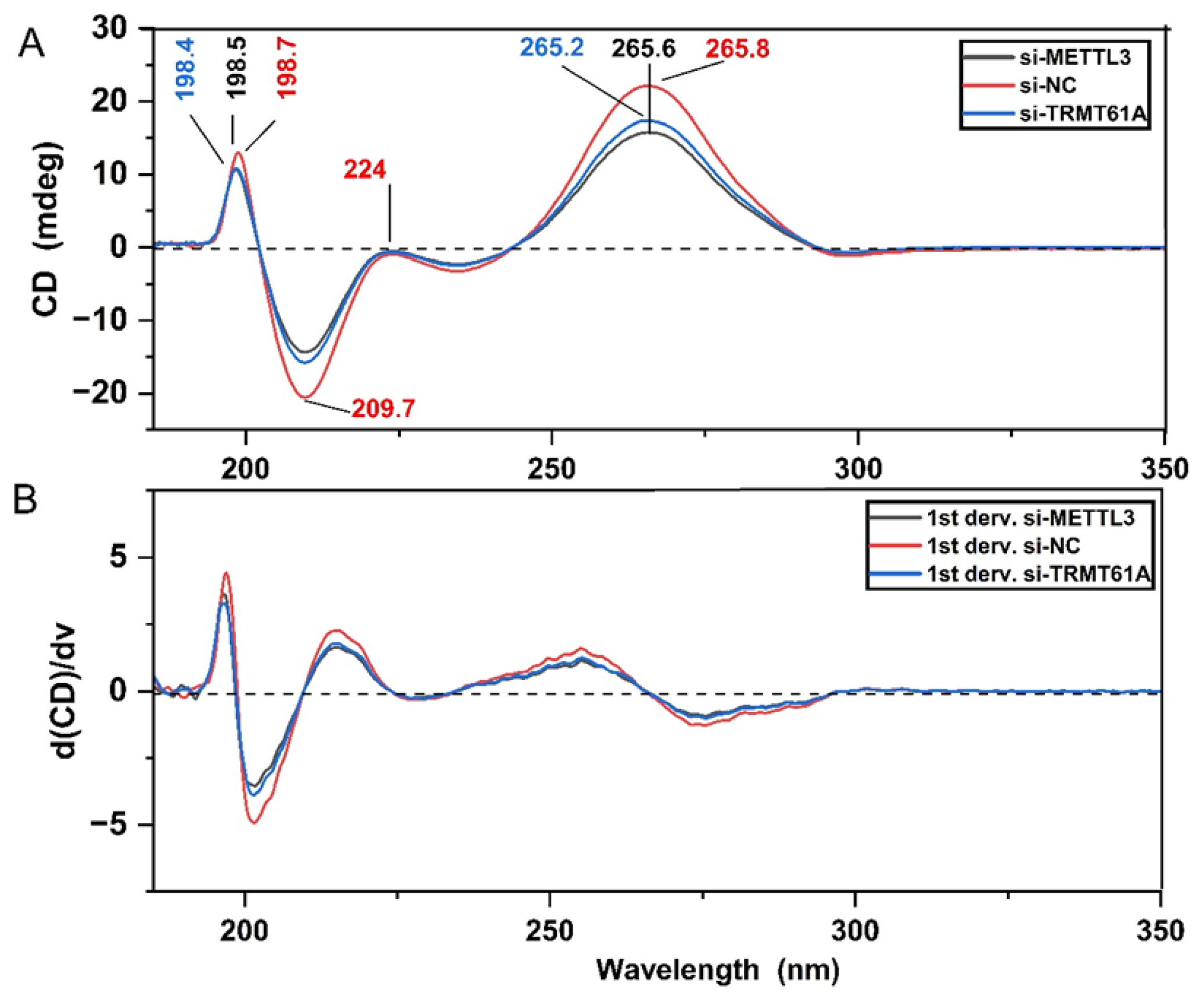
3.2. Spectroscopic Analysis of Perturbations in Cellular m6A Marks
3.3. TNF-α-Mediated Changes in the Biophysical Properties of RNAs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilbert, W.V.; Bell, T.A.; Schaening, C. Messenger RNA Modifications: Form, Distribution, and Function. Science 2016, 352, 1408–1412. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Wei, J.; He, C. Where, When, and How: Context-Dependent Functions of RNA Methylation Writers, Readers, and Erasers. Mol. Cell 2019, 74, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Alasar, A.A.; Tüncel, Ö.; Gelmez, A.B.; Sağlam, B.; Vatansever, İ.E.; Akgül, B. Genomewide M6A Mapping Uncovers Dynamic Changes in the M6A Epitranscriptome of Cisplatin-Treated Apoptotic HeLa Cells. Cells 2022, 11, 3905. [Google Scholar] [CrossRef] [PubMed]
- Akçaöz-Alasar, A.; Tüncel, Ö.; Sağlam, B.; Gazaloğlu, Y.; Atbinek, M.; Cagiral, U.; Iscan, E.; Ozhan, G.; Akgül, B. Epitranscriptomics M6A Analyses Reveal Distinct M6A Marks under Tumor Necrosis Factor α (TNF-α)-Induced Apoptotic Conditions in HeLa Cells. J. Cell. Physiol. 2024, 239, e31176. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Xu, G.; Zhang, X.; Ye, Y.; Cai, W.; Shao, Q. RNA M6A Modification Orchestrates the Rhythm of Immune Cell Development from Hematopoietic Stem Cells to T and B Cells. Front. Immunol. 2022, 13, 839291. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Xu, Y.; Gao, S.; Qin, L.; Austria, Q.; Siedlak, S.L.; Pajdzik, K.; Dai, Q.; He, C.; Wang, W.; et al. METTL3-Dependent RNA M6A Dysregulation Contributes to Neurodegeneration in Alzheimer’s Disease through Aberrant Cell Cycle Events. Mol. Neurodegener. 2021, 16, 70. [Google Scholar] [CrossRef]
- Barbieri, I.; Kouzarides, T. Role of RNA Modifications in Cancer. Nat. Rev. Cancer 2020, 20, 303–322. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Toth, J.I.; Petroski, M.D.; Zhang, Z.; Zhao, J.C. N6 -Methyladenosine Modification Destabilizes Developmental Regulators in Embryonic Stem Cells. Nat. Cell Biol. 2014, 16, 191–198. [Google Scholar] [CrossRef]
- Akgül, B.; Akçaöz-Alasar, A.; Sağlam, B. RNA M6A Methylation at the Juxtaposition of Apoptosis and RNA Therapeutics. Trends Cell Biol. 2024, 34, 801–804. [Google Scholar] [CrossRef] [PubMed]
- Sağlam, B.; Akgül, B. An Overview of Current Detection Methods for RNA Methylation. Int. J. Mol. Sci. 2024, 25, 3098. [Google Scholar] [CrossRef] [PubMed]
- Helm, M.; Motorin, Y. Detecting RNA Modifications in the Epitranscriptome: Predict and Validate. Nat. Rev. Genet. 2017, 18, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Ensinck, I.; Sideri, T.; Modic, M.; Capitanchik, C.; Vivori, C.; Toolan-Kerr, P.; Van Werven, F.J. M6A-ELISA, a Simple Method for Quantifying N6-Methyladenosine from MRNA Populations. RNA 2023, 29, 705–712. [Google Scholar] [CrossRef]
- Güler, G.; Acikgoz, E.; Mukhtarova, G.; Oktem, G. Biomolecular Fingerprints of the Effect of Zoledronic Acid on Prostate Cancer Stem Cells: Comparison of 2D and 3D Cell Culture Models. Arch. Biochem. Biophys. 2024, 753, 109920. [Google Scholar] [CrossRef]
- Güler, G.; Gärtner, R.M.; Ziegler, C.; Mäntele, W. Lipid-Protein Interactions in the Regulated Betaine Symporter BetP Probed by Infrared Spectroscopy. J. Biol. Chem. 2016, 291, 4295–4307. [Google Scholar] [CrossRef] [PubMed]
- Vorob’ev, M.M.; Açıkgöz, B.D.; Güler, G.; Golovanov, A.V.; Sinitsyna, O.V. Proteolysis of Micellar β-Casein by Trypsin: Secondary Structure Characterization and Kinetic Modeling at Different Enzyme Concentrations. Int. J. Mol. Sci. 2023, 24, 3874. [Google Scholar] [CrossRef] [PubMed]
- Zucchiatti, P.; Mitri, E.; Kenig, S.; Bille, F.; Kourousias, G.; Bedolla, D.E.; Vaccari, L. Contribution of Ribonucleic Acid (RNA) to the Fourier Transform Infrared (FTIR) Spectrum of Eukaryotic Cells. Anal. Chem. 2016, 88, 12090–12098. [Google Scholar] [CrossRef] [PubMed]
- Bourassa, P.; Tajmir-Riahi, H.A. Folic Acid Binds DNA and RNA at Different Locations. Int. J. Biol. Macromol. 2015, 74, 337–342. [Google Scholar] [CrossRef]
- Simsek Ozek, N.; Tuna, S.; Erson-Bensan, A.E.; Severcan, F. Characterization of MicroRNA-125b Expression in MCF7 Breast Cancer Cells by ATR-FTIR Spectroscopy. Analyst 2010, 135, 3094–3102. [Google Scholar] [CrossRef] [PubMed]
- Sarić, A.; Rajić, J.; Tolić, A.; Dučić, T.; Vidaković, M. Synchrotron-Based FTIR Microspectroscopy Reveals DNA Methylation Profile in DNA-HALO Structure. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2023, 302, 123090. [Google Scholar] [CrossRef]
- Kelly, J.G.; Najand, G.M.; Martin, F.L. Characterisation of DNA Methylation Status Using Spectroscopy (Mid-IR versus Raman) with Multivariate Analysis. J. Biophotonics 2011, 4, 345–354. [Google Scholar] [CrossRef]
- Duan, M.; Li, Y.; Zhang, F.; Huang, Q. Assessing B-Z DNA Transitions in Solutions via Infrared Spectroscopy. Biomolecules 2023, 13, 964. [Google Scholar] [CrossRef] [PubMed]
- Banyay, M.; Gräslund, A. Structural Effects of Cytosine Methylation on DNA Sugar Pucker Studied by FTIR. J. Mol. Biol. 2002, 324, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Sousa, V.K.; Pedro, J.A.F.; Kumagai, P.S.; Lopes, J.L.S. Effect of Setting Data Collection Parameters on the Reliability of a Circular Dichroism Spectrum. Eur. Biophys. J. 2021, 50, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Le Brun, E.; Arluison, V.; Wien, F. Application of Synchrotron Radiation Circular Dichroism for RNA Structural Analysis. Methods Mol. Biol. 2020, 2113, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Nichols, P.J.; Bevers, S.; Henen, M.; Kieft, J.S.; Vicens, Q.; Vögeli, B. Recognition of Non-CpG Repeats in Alu and Ribosomal RNAs by the Z-RNA Binding Domain of ADAR1 Induces A-Z Junctions. Nat. Commun. 2021, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Güler, G.; Džafić, E.; Vorob’Ev, M.M.; Vogel, V.; Mäntele, W. Real Time Observation of Proteolysis with Fourier Transform Infrared (FT-IR) and UV-Circular Dichroism Spectroscopy: Watching a Protease Eat a Protein. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2011, 79, 104–111. [Google Scholar] [CrossRef]
- Meiser, N.; Mench, N.; Hengesbach, M. RNA Secondary Structure Dependence in METTL3–METTL14 MRNA Methylation Is Modulated by the N-Terminal Domain of METTL. Biol. Chem. 2020, 402, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Szabat, M.; Gudanis, D.; Kotkowiak, W.; Gdaniec, Z.; Kierzek, R.; Pasternak, A. Thermodynamic Features of Structural Motifs Formed by β-L-RNA. PLoS ONE 2016, 11, e0149478. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Toh, J.D.W.; Wong, K.H.Q.; Gao, Y.G.; Hong, W.; Woon, E.C.Y. N 6 -Methyladenosine: A Conformational Marker That Regulates the Substrate Specificity of Human Demethylases FTO and ALKBH5. Sci. Rep. 2016, 6, 25677. [Google Scholar] [CrossRef] [PubMed]
- Arluison, V.; Wien, F. RNA Spectroscopy Methods and Protocols Methods in Molecular Biology; Humana: Louisville, KY, USA, 2020. [Google Scholar]
- Ranjbar, B.; Gill, P. Circular Dichroism Techniques: Biomolecular and Nanostructural Analyses- A Review. Chem. Biol. Drug Des. 2009, 74, 101–120. [Google Scholar] [CrossRef] [PubMed]
- Cheong, A.; Low, J.J.A.; Lim, A.; Yen, P.M.; Woon, E.C.Y. A Fluorescent Methylation-Switchable Probe for Highly Sensitive Analysis of FTO N6-Methyladenosine Demethylase Activity in Cells. Chem. Sci. 2018, 9, 7174–7185. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Yin, P. Structural Insights into N6-Methyladenosine (m6A) Modification in the Transcriptome. Genom. Proteom. Bioinform. 2018, 16, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Wang, Y.; Tang, Q.; Wei, L.; Zhang, X.; Jia, G. An Elongation- and Ligation-Based QPCR Amplification Method for the Radiolabeling-Free Detection of Locus-Specific N 6 -Methyladenosine Modification. Angew. Chem. 2018, 130, 16227–16232. [Google Scholar] [CrossRef]
- Senguen, F.T.; Doran, T.M.; Anderson, E.A.; Nilsson, B.L. Clarifying the Influence of Core Amino Acid Hydrophobicity, Secondary Structure Propensity, and Molecular Volume on Amyloid-β 16-22 Self-Assembly. Mol. Biosyst. 2011, 7, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Andrade, J.; Pereira, C.G.; Ranquine, T.; Azarias, C.A.; Bell, M.J.V.; De Carvalho Dos Anjos, V. Long-Term Ripening Evaluation of Ewes’ Cheeses by Fourier-Transformed Infrared Spectroscopy under Real Industrial Conditions. J. Spectrosc. 2018, 2018, 1381864. [Google Scholar] [CrossRef]
- Banyay, M.; Sarkar, M.; Graslund, A. A Library of IR Bands of Nucleic Acids in Solution. Biophys. Chem. 2003, 104, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Micura, R.; Pils, W.; Höbartner, C.; Grubmayr, K.; Ebert, M.-O.; Jaun, B. Methylation of the Nucleobases in RNA Oligonucleotides Mediates Duplex–Hairpin Conversion. Nucleic Acids Res. 2001, 29, 3997–4005. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C.J.T.; Pan, T.; Kalsotra, A. RNA Modifications and Structures Cooperate to Guide RNA-Protein Interactions. Nat. Rev. Mol. Cell Biol. 2017, 18, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Kierzek, E.; Zhang, X.; Watson, R.M.; Kennedy, S.D.; Szabat, M.; Kierzek, R.; Mathews, D.H. Secondary Structure Prediction for RNA Sequences Including N6-Methyladenosine. Nat. Commun. 2022, 13, 1271. [Google Scholar] [CrossRef] [PubMed]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the Human and Mouse M6A RNA Methylomes Revealed by M6A-Seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Dovbeshko, G.I.; Gridina, N.Y.; Kruglova, E.B.; Pashchuk, O.P. FTIR Spectroscopy Studies of Nucleic Acid Damage. Talanta 2000, 53, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Geinguenaud, F.; Militello, V.; Arluison, V. Application of FTIR Spectroscopy to Analyze RNA Structure. RNA Spectrosc. Methods Protoc. 2020, 2113, 119–133. [Google Scholar] [CrossRef]
- Wood, B.R. The Importance of Hydration and DNA Conformation in Interpreting Infrared Spectra of Cells and Tissues. Chem. Soc. Rev. 2016, 45, 1980–1998. [Google Scholar] [CrossRef]
- Hwang, W.; Arluison, V.; Hohng, S. Dynamic Competition of DsrA and RpoS Fragments for the Proximal Binding Site of Hfq as a Means for Efficient Annealing. Nucleic Acids Res. 2011, 39, 5131–5139. [Google Scholar] [CrossRef] [PubMed]
- Bokar, J.A.; Shambaugh, M.E.; Polayes, D.; Matera, A.G.; Rottman, F.M. Purification and CDNA Cloning of the AdoMet-Binding Subunit of the Human MRNA (N6-Adenosine)-Methyltransferase. RNA 1997, 3, 1233–1247. [Google Scholar] [PubMed]
- Anderson, J.; Phan, L.; Hinnebusch, A.G. The Gcd10pGcd14p Complex Is the Essential Two-Subunit TRNA(1-Methyladenosine) Methyltransferase of Saccharomyces Cerevisiae. Proc. Natl. Acad. Sci. USA 2000, 97, 5173–5178. [Google Scholar] [CrossRef]
- Lindqvist, M.; Sarkar, M.; Winqvist, A.; Rozners, E.; Strömberg, R.; Gräslund, A. Optical Spectroscopic Study of the Effects of a Single Deoxyribose Substitution in a Ribose Backbone: Implications in RNA-RNA Interaction. Biochemistry 2000, 39, 1693–1701. [Google Scholar] [CrossRef]
- Güler, G.; Vorob’Ev, M.M.; Vogel, V.; Mäntele, W. Proteolytically-Induced Changes of Secondary Structural Protein Conformation of Bovine Serum Albumin Monitored by Fourier Transform Infrared (FT-IR) and UV-Circular Dichroism Spectroscopy. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2016, 161, 8–18. [Google Scholar] [CrossRef]
- Nandiyanto, A.B.D.; Ragadhita, R.; Fiandini, M. Interpretation of Fourier Transform Infrared Spectra (FTIR): A Practical Approach in the Polymer/Plastic Thermal Decomposition. Indones. J. Sci. Technol. 2023, 8, 113–126. [Google Scholar] [CrossRef]
- Banyay, M.; Sandbrink, J.; Strömberg, R.; Gräslund, A. Characterization of an RNA Bulge Structure by Fourier Transform Infrared Spectroscopy. Biochem. Biophys. Res. Commun. 2004, 324, 634–639. [Google Scholar] [CrossRef]
- Liu, Z.-G.; Hsu, H.; Goeddel, D.V. Dissection of TNF Receptor 1 Effector Functions: JNK Activation Is Not Linked to Apoptosis While NF-B Activation Prevents Cell Death. Cell 1996, 87, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Riazance, J.H.; Baase, W.A.; Johnson, W.C.; Hall, K.; Cruz, P.; Tinoco, I. Evidence for Z-form RNA by vacuum UV circular dichroism. Nucleic Acids Res. 1985, 13, 4983–4989. [Google Scholar] [CrossRef] [PubMed]
- Vanloon, J.; Bennett, H.A.; Martin, A.; Wien, F.; Harroun, T.; Yan, H. Synchrotron Radiation Circular Dichroism Spectroscopy of Oligonucleotides at Millimolar Concentrations. Bioorg. Med. Chem. Lett. 2023, 92, 129376. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′ → 3′) |
|---|---|
| MALAT1 m6A2515 UP | tagccagtaccgtagtgcgtgAATTACTTCCGTTACGAAAG |
| MALAT1 m6A2515 DOWN | 5phos/CCTTCACATTTTTCAAACTAAGCTACTcagaggctgagtcgctgcat |
| MALAT1 A2511 UP | tagccagtaccgtagtgcgtgAATTACTTCCGTTACGAAAGTCCT |
| MALAT1 A2511 DOWN | 5phos/CACATTTTTCAAACTAAGCTACTcagaggctgagtcgctgcat |
| SELECT qRT-PCR Forward | ATGCAGCGACTCAGCCTCTG |
| SELECT qRT-PCR Reverse | TAGCCAGTACCGTAGTGCGTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sağlam, B.; Akkuş, O.; Akçaöz-Alasar, A.; Ceylan, Ç.; Güler, G.; Akgül, B. An Investigation of RNA Methylations with Biophysical Approaches in a Cervical Cancer Cell Model. Cells 2024, 13, 1832. https://doi.org/10.3390/cells13221832
Sağlam B, Akkuş O, Akçaöz-Alasar A, Ceylan Ç, Güler G, Akgül B. An Investigation of RNA Methylations with Biophysical Approaches in a Cervical Cancer Cell Model. Cells. 2024; 13(22):1832. https://doi.org/10.3390/cells13221832
Chicago/Turabian StyleSağlam, Buket, Onur Akkuş, Azime Akçaöz-Alasar, Çağatay Ceylan, Günnur Güler, and Bünyamin Akgül. 2024. "An Investigation of RNA Methylations with Biophysical Approaches in a Cervical Cancer Cell Model" Cells 13, no. 22: 1832. https://doi.org/10.3390/cells13221832
APA StyleSağlam, B., Akkuş, O., Akçaöz-Alasar, A., Ceylan, Ç., Güler, G., & Akgül, B. (2024). An Investigation of RNA Methylations with Biophysical Approaches in a Cervical Cancer Cell Model. Cells, 13(22), 1832. https://doi.org/10.3390/cells13221832