

The Importance of the Position of the Nucleus in Drosophila Oocyte Development

 and
and 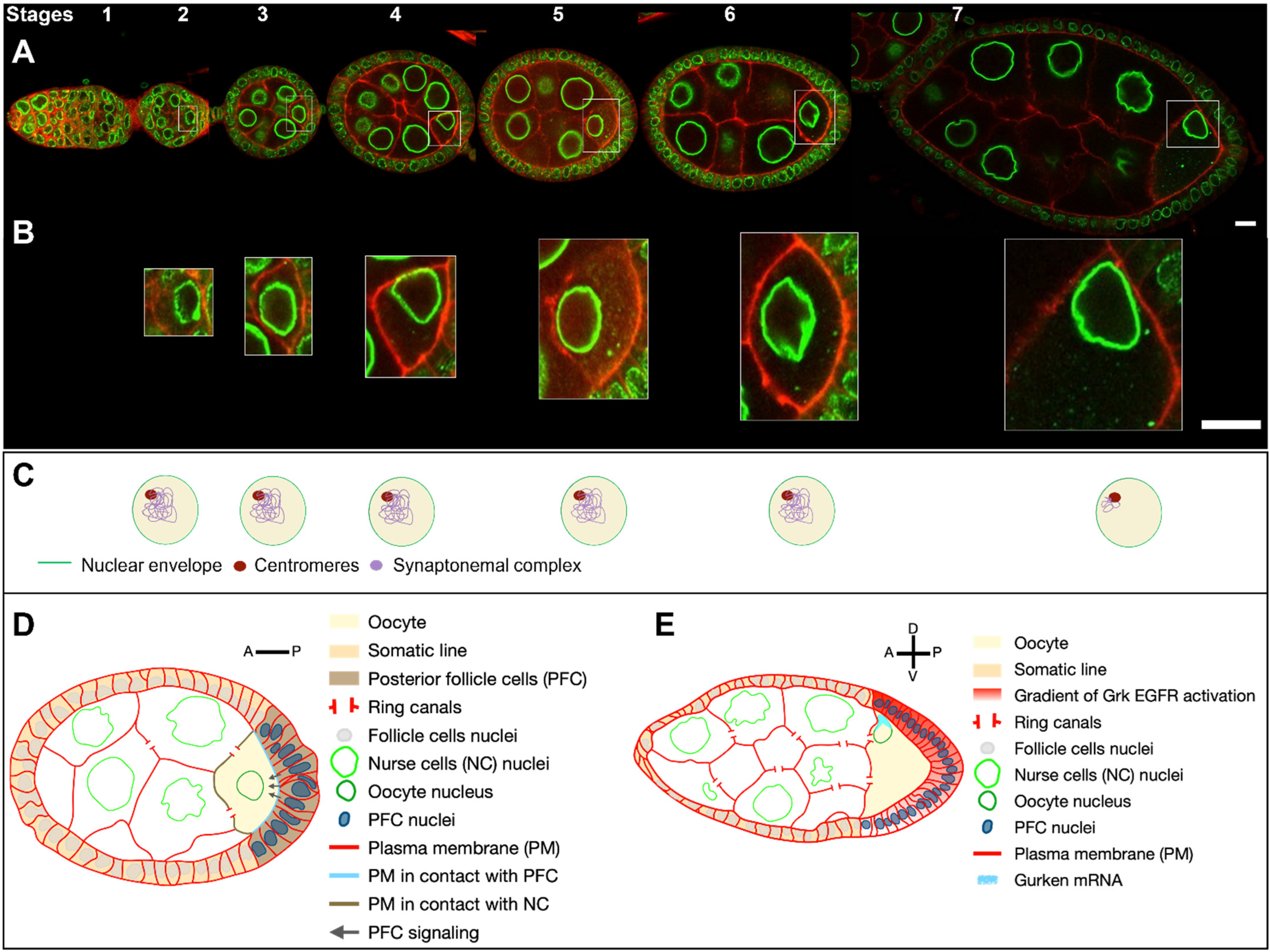{kind=link}
{kind=link}
{kind=link}
Abstract
1. Anatomy and Development of the Drosophila Egg Chamber
- During early oogenesis, before stage 6, the grk mRNA is localized close to the nucleus hemisphere facing the posterior of the oocyte, and its translation leads to the activation of the EGFr in about 20 follicle cells adjacent to the oocyte. This induces a differentiation program necessary for building the posterior structures of the eggshell [35,36,37,38] and for the emission, in conjunction with the JAK-STAT signalling pathway, of a signal from those cells, that later triggers a repolarization of the MT network of the oocyte and the start of nucleus migration [37,38,39] (Figure 1C).
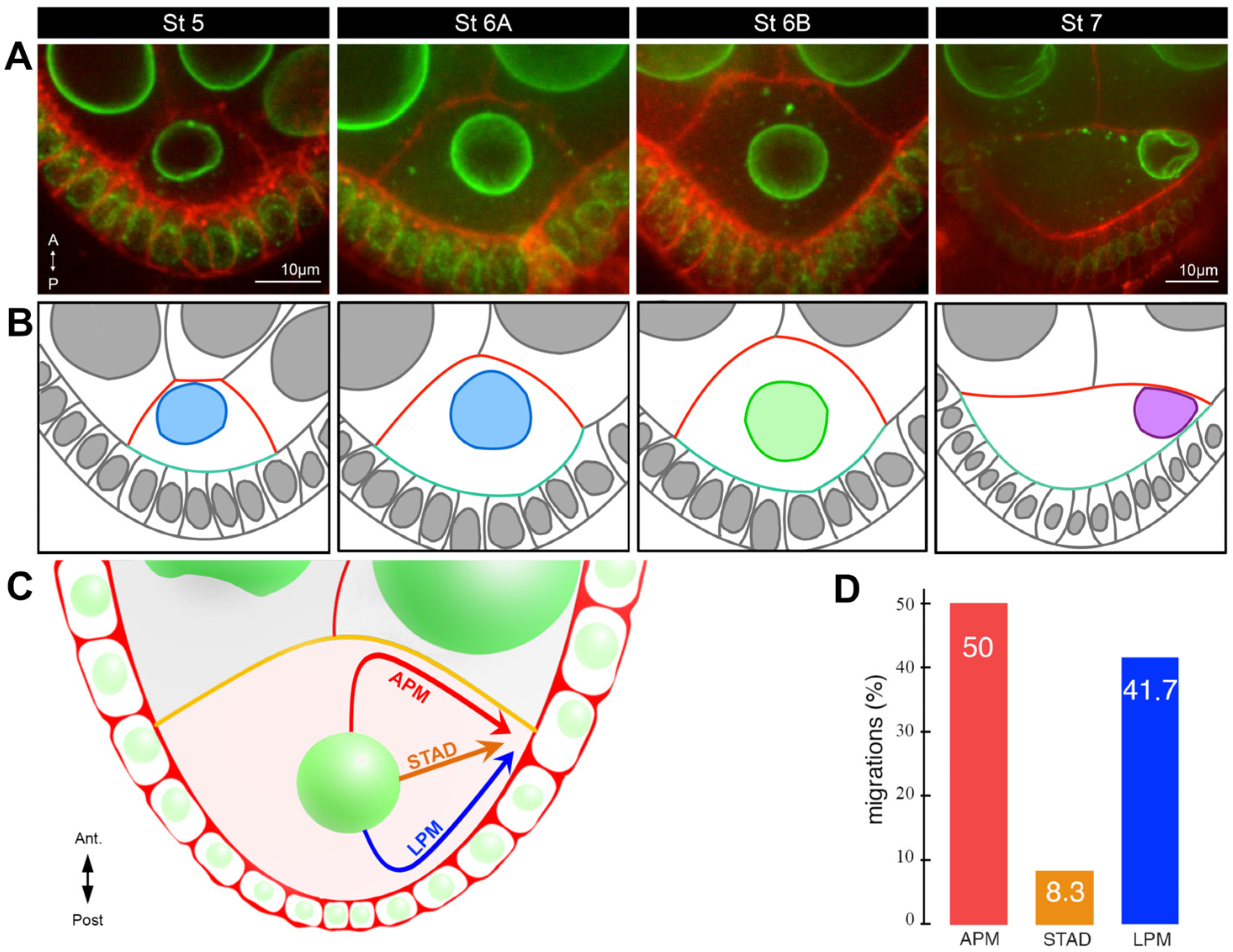
- With the asymmetrical positioning of the nucleus to an antero-dorsal position, the grk mRNA, still associated with the nucleus, relocalises in the vicinity of the antero-dorsal follicle cells where its translation triggers a second wave of EGFr activation in a gradient pattern (Figure 1E). This enables the formation of two groups of follicle cells, which forms the dorsal appendages. Through a series of steps, the cells that lack EGFr activation because they do not receive the Grk signal, secrete the ligand triggering the formation of most of the different germ layers of the embryo via the activation of the Toll receptor [7,8]. A crucial step in this process is the asymmetrical positioning of the nucleus, which functions as a symmetry-breaking event for the formation of the dorsoventral axis of the eggshell and of the future embryo. Because the lateral follicle cells are all equivalent before this second wave of Grk signaling, and because there are no markers that predict where the nucleus would move within the oocyte, it is thought the nucleus can migrate towards any position representing the intersection between the anterior and lateral parts of the oocyte. Further support for this view comes from an elegant experiment showing in oocytes with two nuclei, the nuclei migrate to random positions with respect to each other [40].
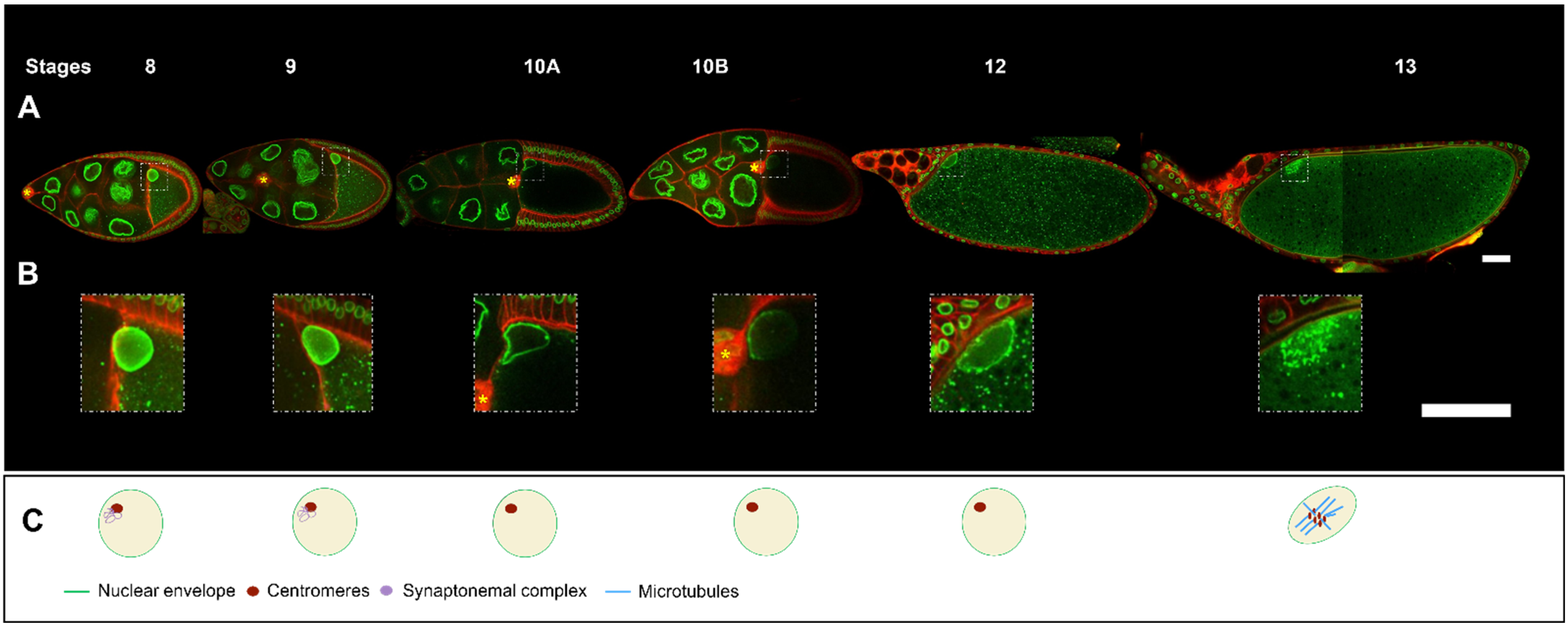
2. Steps in Nucleus Positioning during Oocyte Development
Oocyte Nucleus Positioning and Cytoskeleton
3. Several MT Networks Participate in Nuclear Positioning
3.1. Centrosome Involvement
3.2. A Second MT Network Operates at the Level of the Nuclear Envelope
4. The Relationship between the Mechanical Forces Exerted on the Nucleus and Its Intranuclear Organisation and Positioning
5. What Mechanism Triggers Nucleus Migration?
6. Oocyte Nucleus and Meiotic Divisions
7. Oocyte Nucleus Anchoring
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- King, R.C. Ovarian Development in Drosophila Melanogaster; Academic Press: New York, NY, USA, 1970. [Google Scholar]
- McLaughlin, J.M.; Bratu, D.P. Drosophila melanogaster Oogenesis: An Overview. In Drosophila Oogenesis: Methods and Protocols; Bratu, D.P., McNeil, G.P., Eds.; Springer New York: New York, NY, USA, 2015; pp. 1–20. [Google Scholar] [CrossRef]
- Almonacid, M.; Terret, M.-E.; Verlhac, M.-H. Control of nucleus positioning in mouse oocytes. Semin. Cell Dev. Biol. 2017, 82, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.E.; Miller, D.E.; Miller, A.L.; Hawley, R.S. Female Meiosis: Synapsis, Recombination, and Segregation in Drosophila melanogaster. Genetics 2018, 208, 875–908. [Google Scholar] [CrossRef] [PubMed]
- Mahowald, A.P.; Tiefert, M. Fine structural changes in the Drosophila oocyte nucleus during a short period of RNA synthesis—An autoradiographic and ultrastructural study of RNA synthesis in the oocyte nucleus of Drosophila. Wilhelm Roux. Arch. Entwickl. Mech. Org. 1970, 165, 8–25. [Google Scholar] [CrossRef]
- Liu, J.L.; Buszczak, M.; Gall, J.G. Nuclear bodies in the Drosophila germinal vesicle. Chromosom. Res. 2006, 14, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Roth, S.; Lynch, J.A. Symmetry breaking during Drosophila oogenesis. Cold Spring Harb. Perspect. Biol. 2009, 1, a001891. [Google Scholar] [CrossRef]
- Moussian, B.; Roth, S. Dorsoventral axis formation in the Drosophila embryo—Shaping and transducing a morphogen gradient. Curr. Biol. 2005, 15, 887–899. [Google Scholar] [CrossRef]
- Duchek, P.; Rørth, P. Guidance of Cell Migration by EGF Receptor Signaling During Drosophila Oogenesis. Science 2001, 291, 131–133. [Google Scholar] [CrossRef]
- Huynh, J.R.; St Johnston, D. The origin of asymmetry: Early polarisation of the Drosophila germline cyst and oocyte. Curr. Biol. 2004, 14, 438–449. [Google Scholar] [CrossRef]
- Adelaide, C. Electron microscopy of meiosis in Drosophila melanogaster females. I. Structure, arrangement, and temporal change of the synaptonemal complex in wild-type. Chromosoma 1975, 51, 157–182. [Google Scholar]
- Theurkauf, W.E.; Alberts, B.M.; Nung Jan, Y.; Jongens, T.A.; Jan, Y.N.; Jongens, T.A. A central role for microtubules in the differentiation of Drosophila oocytes. Development 1993, 1169–1180. [Google Scholar] [CrossRef]
- Nicolas, E.; Chenouard, N.; Olivo-Marin, J.-C.J.-C.; Guichet, A. A dual role for actin and microtubule cytoskeleton in the transport of golgi units from the nurse cells to the oocyte across ring canals. Mol. Biol. Cell 2009, 20, 556–568. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Lakonishok, M.; Gelfand, V.I. Gatekeeper function for Short stop at the ring canals of the Drosophila ovary. Curr. Biol. 2021, 31, 3207–3219.e5. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Lakonishok, M.; Serpinskaya, A.S.; Gelfand, V.I. A novel mechanism of bulk cytoplasmic transport by cortical dynein in Drosophila ovary. eLife 2022, 11, e75538. [Google Scholar] [CrossRef] [PubMed]
- Januschke, J.; Gervais, L.; Gillet, L.; Keryer, G.; Bornens, M.; Guichet, A. The centrosome-nucleus complex and microtubule organization in the Drosophila oocyte. Development 2006, 133, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Nashchekin, D.; Fernandes, A.R.; St. Johnston, D. Patronin/Shot Cortical Foci Assemble the Noncentrosomal Microtubule Array that Specifies the Drosophila Anterior-Posterior Axis. Dev. Cell 2016, 38, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Tissot, N.; Lepesant, J.-A.A.; Bernard, F.; Legent, K.; Bosveld, F.; Martin, C.; Faklaris, O.; Bellaïche, Y.; Coppey, M.; Guichet, A. Distinct molecular cues ensure a robust microtubule-dependent nuclear positioning in the Drosophila oocyte. Nat. Commun. 2017, 8, 15168. [Google Scholar] [CrossRef] [PubMed]
- Nashchekin, D.; Busby, L.; Jakobs, M.; Squires, I.; St. Johnston, D. Symmetry breaking in the female germline cyst. Science 2021, 374, 874–879. [Google Scholar] [CrossRef]
- Roper, K.; Brown, N. A Spectraplakin Is Enriched on the Fusome and Organizes Microtubules during Oocyte Specification in Drosophila. Curr. Biol. 2004, 14, 99–110. [Google Scholar] [CrossRef]
- Bolívar, J.; Huynh, J.-R.; López-Schier, H.; González, C.; St. Johnston, D.; González-Reyes, A. Centrosome migration into the Drosophila oocyte is independent of BicD and egl, and of the organisation of the microtubule cytoskeleton. Development 2001, 1909, 1889–1909. [Google Scholar] [CrossRef]
- Zhao, T.; Graham, O.S.; Raposo, A.; St. Johnston, D. Growing Microtubules Push the Oocyte Nucleus to Polarize the Drosophila Dorsal-Ventral Axis. Science 2012, 336, 999–1003. [Google Scholar] [CrossRef]
- Loh, M.; Bernard, F.; Guichet, A. Kinesin-1 promotes centrosome clustering and nuclear migration in the Drosophila oocyte. Development 2023, 150, dev201728. [Google Scholar] [CrossRef] [PubMed]
- Pimenta-Marques, A.; Bento, I.; Lopes, C.A.M.; Duarte, P.; Jana, S.C.; Bettencourt-Dias, M. A mechanism for the elimination of the female gamete centrosome in Drosophila melanogaster. Science 2016, 353, aaf4866. [Google Scholar] [CrossRef]
- St Johnston, D. Polarity and axis formation in the Drosophila female germ line. Curr. Top. Dev. Biol. 2023, 154, 73–97. [Google Scholar] [PubMed]
- Robinson, D.N.; Cant, K.; Cooley, L. Morphogenesis of Drosophila ovarian ring canals. Development 2015, 2025, 2015–2025. [Google Scholar] [CrossRef] [PubMed]
- Driever, W.; Nüsslein-Volhard, C. A gradient of bicoid protein in Drosophila embryos. Cell 1988, 54, 83–93. [Google Scholar] [CrossRef] [PubMed]
- St. Johnston, D.; Driever, W.; Berleth, T.; Richstein, S.; Nusslein-Volhard, C. Multiple steps in the localization of bicoid RNA to the anterior pole of the Drosophila oocyte. Development 1989, 107, 13–19. [Google Scholar] [CrossRef]
- Ephrussi, A.; Lehmann, R. Induction of germ cell fate by oskar. Nature 1992, 358, 387–392. [Google Scholar] [CrossRef]
- Ephrussi, A.; Dickinson, L.K.; Lehmann, R. Oskar Organizes the Germ Plasm and Directs Localization of the Posterior Determinant Nanos. Cell 1991, 66, 37–50. [Google Scholar] [CrossRef]
- Gavis, E.R.; Lehmann, R. Localization of nanos RNA controls embryonic polarity. Cell 1992, 71, 301–313. [Google Scholar] [CrossRef]
- Kim-Ha, J.; Smith, J.L.; Macdonald, P.M. oskar mRNA is localized to the posterior pole of the Drosophila oocyte. Cell 1991, 66, 23–35. [Google Scholar] [CrossRef]
- Jeske, M.; Müller, C.W.; Ephrussi, A. The LOTUS domain is a conserved DEAD-box RNA helicase regulator essential for the recruitment of Vasa to the germ plasm and nuage. Genes Dev. 2017, 31, 939–952. [Google Scholar] [CrossRef] [PubMed]
- Neuman-Silberberg, F.S.; Schüpbach, T. The drosophila dorsoventral patterning gene gurken produces a dorsally localized RNA and encodes a TGFα-like protein. Cell 1993, 75, 165–174. [Google Scholar] [CrossRef]
- Acaimo, G.-R.; St. Johnston, D. Role of Oocyte Position in Establishment of Anterior-Posterior Polarity in Drosophila. Science 1994, 266, 639–642. [Google Scholar]
- González-Reyes, A.; St. Johnston, D. Patterning of the follicle cell epithelium along the anterior-posterior axis during Drosophila oogenesis. Development 1998, 125, 2837–2846. [Google Scholar] [CrossRef] [PubMed]
- Roth, S.; Neuman-Silberberg, F.S.; Barcelo, G.; Schüpbach, T. cornichon and the EGF receptor signaling process are necessary for both anterior-posterior and dorsal-ventral pattern formation in Drosophila. Cell 1995, 81, 967–978. [Google Scholar] [CrossRef] [PubMed]
- González-Reyes, A.; Elliott, H.; St. Johnston, D. Polarization of both major body axes in Drosophila by gurken-torpedo signalling. Nature 1995, 375, 654–658. [Google Scholar] [CrossRef] [PubMed]
- Xi, R.; McGregor, J.R.; Harrison, D.A. A gradient of JAK pathway activity patterns the anterior-posterior axis of the follicular epithelium. Dev. Cell 2003, 4, 167–177. [Google Scholar] [CrossRef]
- Roth, S.; Jordan, P.; Karess, R. Binuclear Drosophila oocytes: Consequences and implications for dorsal-ventral patterning in oogenesis and embryogenesis. Development 1999, 126, 927–934. [Google Scholar] [CrossRef]
- Paré, C.; Suter, B. Subcellular localization of Bic-D::GFP is linked to an asymmetric oocyte nucleus. J. Cell Sci. 2000, 113, 2119–2127. [Google Scholar] [CrossRef]
- Guichet, A.; Peri, F.; Roth, S. Stable anterior anchoring of the oocyte nucleus is required to establish dorsoventral polarity of the drosophila egg. Dev. Biol. 2001, 237, 93–106. [Google Scholar] [CrossRef]
- Horne-Badovinac, S. The Drosophila micropyle as a system to study how epithelia build complex extracellular structures. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190561. [Google Scholar] [CrossRef] [PubMed]
- Koch, E.A.; Spitzer, R.H. Multiple effects of colchicine on oogenesis in Drosophila: Induced sterility and switch of potential oocyte to nurse-cell developmental pathway. Cell Tissue Res. 1983, 228, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Baffet, A.D.D.; Benoit, B.; Januschke, J.; Audo, J.; Gourhand, V.; Roth, S.; Guichet, A. Drosophila tubulin-binding cofactor B is required for microtubule network formation and for cell polarity. Mol. Biol. Cell 2012, 23, 3591–3601. [Google Scholar] [CrossRef] [PubMed]
- Chemla, L.; Guichet, A. Université Paris-Cité, CNRS, Institut Jacques Monod, Paris, France. 2024; manuscript in preparation. [Google Scholar]
- Manseau, L.; Galley, J.; Phan, H. Profilin is required for posterior patterning of the Drosophila oocyte. Development 1996, 122, 2109–2116. [Google Scholar] [CrossRef] [PubMed]
- Verheyen, E.M.; Cooley, L. Profilin mutations disrupt multiple actin-dependent processes during Drosophila development. Development 1994, 120, 717–728. [Google Scholar] [CrossRef]
- Mahowald, A.P.; Strassheim, J.M. Intercellular migration of centrioles in the germarium of Drosophila melanogaster. An electron microscopic study. J. Cell Biol. 1970, 45, 306–320. [Google Scholar] [CrossRef] [PubMed]
- Matthies, H.J.G.; McDonald, H.B.; Goldstein, L.S.B.; Theurkauf, W.E. Anastral meiotic spindle morphogenesis: Role of the non-claret disjunctional kinesin-like protein. J. Cell Biol. 1996, 134, 455–464. [Google Scholar] [CrossRef]
- Riparbelli, M.G.; Persico, V.; Callaini, G. Cell-to-Cell Interactions during Early Drosophila Oogenesis: An Ultrastructural Analysis. Cells 2022, 11, 2658. [Google Scholar] [CrossRef]
- Stevens, N.R.; Raposo, A.A.S.F.; Basto, R.; St. Johnston, D.; Raff, J.W.W. From Stem Cell to Embryo without Centrioles. Curr. Biol. 2007, 17, 1498–1503. [Google Scholar] [CrossRef]
- Hannaford, M.R.; Liu, R.; Billington, N.; Swider, Z.T.; Galletta, B.J.; Fagerstrom, C.J.; Combs, C.; Sellers, J.R.; Rusan, N.M. Pericentrin interacts with Kinesin-1 to drive centriole motility. J. Cell Biol. 2022, 221, e202112097. [Google Scholar] [CrossRef]
- Roque, H.; Saurya, S.; Pratt, M.B.; Johnson, E.; Raff, J.W. Drosophila PLP assembles pericentriolar clouds that promote centriole stability, cohesion and MT nucleation. PLoS Genet. 2018, 14, e1007198. [Google Scholar] [CrossRef] [PubMed]
- Lele, T.P.; Dickinson, R.B.; Gundersen, G.G. Mechanical principles of nuclear shaping and positioning. J. Cell Biol. 2018, 217, 3330–3342. [Google Scholar] [CrossRef]
- Sun, T.; Song, Y.; Dai, J.; Mao, D.; Ma, M.; Ni, J.-Q.Q.; Liang, X.; Pastor-Pareja, J.C. Spectraplakin Shot Maintains Perinuclear Microtubule Organization in Drosophila Polyploid Cells. Dev. Cell 2019, 49, 731–747.e7. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Buchwalter, R.A.; Zheng, C.; Wight, E.M.; Chen, J.V.; Megraw, T.L. A perinuclear microtubule-organizing centre controls nuclear positioning and basement membrane secretion. Nat. Cell Biol. 2020, 22, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Roman, W.; Gomes, E.R. Nuclear positioning in skeletal muscle. Semin. Cell Dev. Biol. 2017, 82, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.X.; Guan, Z.; Nash, H.A. The mushroom body defect gene product is an essential component of the meiosis II spindle apparatus in Drosophila oocytes. Genetics 2006, 173, 243–253. [Google Scholar] [CrossRef]
- Januschke, J.; Gervais, L.; Dass, S.; Kaltschmidt, J.A.; Lopez-Schier, H.; St. Johnston, D.; Brand, A.H.; Roth, S.; Guichet, A. Polar transport in the Drosophila oocyte requires Dynein and Kinesin I cooperation. Curr. Biol. 2002, 12, 1971–1981. [Google Scholar] [CrossRef] [PubMed]
- van der Voet, M.; Berends, C.W.H.; Perreault, A.; Nguyen-Ngoc, T.; Gönczy, P.; Vidal, M.; Boxem, M.; van den Heuvel, S. NuMA-related LIN-5, ASPM-1, calmodulin and dynein promote meiotic spindle rotation independently of cortical LIN-5/GPR/Gα. Nat. Cell Biol. 2009, 11, 269–277. [Google Scholar] [CrossRef]
- Roland-Gosselin, F.; Guichet, A.; Bernard, F. Université Paris-Cité, CNRS, Institut Jacques Monod, Paris, France. 2024; manuscript in preparation. [Google Scholar]
- Jahed, Z.; Soheilypour, M.; Peyro, M.; Mofrad, M.R.K. The LINC and NPC relationship—It’s complicated! J. Cell Sci. 2016, 129, 3219–3229. [Google Scholar] [CrossRef]
- Kracklauer, M.P.; Wiora, H.M.; Deery, W.J.; Chen, X.; Bolival, B.; Romanowicz, D.; Simonette, R.A.; Fuller, M.T.; Fischer, J.A.; Beckingham, K.M. The Drosophila SUN protein Spag4 cooperates with the coiled-coil protein Yuri Gagarin to maintain association of the basal body and spermatid nucleus. J. Cell Sci. 2010, 123, 2763–2772. [Google Scholar] [CrossRef]
- Technau, M.; Roth, S. The Drosophila KASH domain proteins Msp-300 and Klarsicht and the SUN domain protein klaroid have no essential function during oogenesis. Fly 2008, 2, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Christophorou, N.; Rubin, T.; Bonnet, I.; Piolot, T.; Arnaud, M.; Huynh, J.-R. Microtubule-driven nuclear rotations promote meiotic chromosome dynamics. Nat. Cell Biol. 2015, 17, 1388–1400. [Google Scholar] [CrossRef] [PubMed]
- Lepesant, J.A.; Guichet, A. Université Paris-Cité, CNRS, Institut Jacques Monod, Paris, France. 2024; manuscript in preparation. [Google Scholar]
- Al Jord, A.; Letort, G.; Chanet, S.; Tsai, F.C.; Antoniewski, C.; Eichmuller, A.; Da Silva, C.; Huynh, J.R.; Gov, N.S.; Voituriez, R.; et al. Cytoplasmic forces functionally reorganize nuclear condensates in oocytes. Nat. Commun. 2022, 13, 5070. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Costa, P.; McCarthy, A.; Prudêncio, P.; Greer, C.; Guilgur, L.G.; Becker, J.D.; Secombe, J.; Rangan, P.; Martinho, R.G. Early programming of the oocyte epigenome temporally controls late prophase I transcription and chromatin remodelling. Nat. Commun. 2016, 7, 12331. [Google Scholar] [CrossRef] [PubMed]
- Milas, A.; De-Carvalho, J.; Telley, I.A. Follicle cell contact maintains main body axis polarity in the Drosophila melanogaster oocyte. J. Cell Biol. 2023, 222, e202209052. [Google Scholar] [CrossRef]
- Doerflinger, H.; Zimyanin, V.; St. Johnston, D. The Drosophila anterior-posterior axis is polarized by asymmetric myosin activation. Curr. Biol. 2022, 32, 374–385.e4. [Google Scholar] [CrossRef]
- Skold, H.; Komma, D.; Endow, S. Assembly pathway of the anastral Drosophila oocyte meiosis I spindle. J. Cell Sci. 2005, 118, 1745. [Google Scholar] [CrossRef]
- Riparbelli, M.G.; Callaini, G. The meiotic spindle of the Drosophila oocyte: The role of centrosomin and the central aster. J. Cell Sci. 2005, 118, 2827–2836. [Google Scholar] [CrossRef]
- Quinlan, M.E. Cytoplasmic Streaming in the Drosophila Oocyte. Annu. Rev. Cell Dev. Biol. 2016, 32, 173–195. [Google Scholar] [CrossRef]
- Theurkauf, W.E.; Smiley, S.; Wong, M.L.; Alberts, B.M. Reorganization of the cytoskeleton during Drosophila oogenesis: Implications for axis specification and intercellular transport. Development 1992, 115, 923–936. [Google Scholar] [CrossRef]
- Duncan, J.E.; Warrior, R. The cytoplasmic dynein and kinesin motors have interdependent roles in patterning the Drosophila oocyte. Curr. Biol. 2002, 12, 1982–1991. [Google Scholar] [CrossRef]
- Swan, A.; Nguyen, T.; Suter, B. Drosophila Lissencephaly-1 functions with Bic-D and dynein in oocyte determination and nuclear positioning. Nat. Cell Biol. 1999, 1, 444–449. [Google Scholar] [CrossRef]
- Swan, A.; Suter, B. Role of Bicaudal-D in patterning the Drosophila egg chamber in mid-oogenesis. Development 1996, 122, 3577–3586. [Google Scholar] [CrossRef]
- Gervais, L.; Claret, S.; Januschke, J.; Roth, S.; Guichet, A. PIP5K-dependent production of PIP2 sustains microtubule organization to establish polarized transport in the Drosophila oocyte. Development 2008, 135, 3829–3838. [Google Scholar] [CrossRef] [PubMed]
- Kotak, S.; Busso, C.; Gönczy, P. NuMA interacts with phosphoinositides and links the mitotic spindle with the plasma membrane. EMBO J. 2014, 33, 1815–1830. [Google Scholar] [CrossRef] [PubMed]
- Claret, S.; Guichet, A. Université Paris-Cité, CNRS, Institut Jacques Monod, Paris, France. 2024; manuscript in preparation. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lepesant, J.-A.; Roland-Gosselin, F.; Guillemet, C.; Bernard, F.; Guichet, A. The Importance of the Position of the Nucleus in Drosophila Oocyte Development. Cells 2024, 13, 201. https://doi.org/10.3390/cells13020201
Lepesant J-A, Roland-Gosselin F, Guillemet C, Bernard F, Guichet A. The Importance of the Position of the Nucleus in Drosophila Oocyte Development. Cells. 2024; 13(2):201. https://doi.org/10.3390/cells13020201
Chicago/Turabian StyleLepesant, Jean-Antoine, Fanny Roland-Gosselin, Clémentine Guillemet, Fred Bernard, and Antoine Guichet. 2024. "The Importance of the Position of the Nucleus in Drosophila Oocyte Development" Cells 13, no. 2: 201. https://doi.org/10.3390/cells13020201
APA StyleLepesant, J.-A., Roland-Gosselin, F., Guillemet, C., Bernard, F., & Guichet, A. (2024). The Importance of the Position of the Nucleus in Drosophila Oocyte Development. Cells, 13(2), 201. https://doi.org/10.3390/cells13020201