
Identification and Characterization of ATOH7-Regulated Target Genes and Pathways in Human Neuroretinal Development

 ,
,  , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Human iPSC Maintenance
2.2. CRISPR/Cas9 Gene Editing
2.3. Retinal Organoid Differentiation
2.4. Quantitative Reverse-Transcription PCR
2.5. Immunocytochemistry and Axon Sprouting Assay
2.6. Retinal Cell Dissociation
2.7. Cut under Target and Release Using Nuclease Sequencing
2.8. Reporter-Enriched mRNA Sequencing
2.9. Gene Enrichment Analysis
2.10. Fixed Single-Cell RNA Sequencing
3. Results
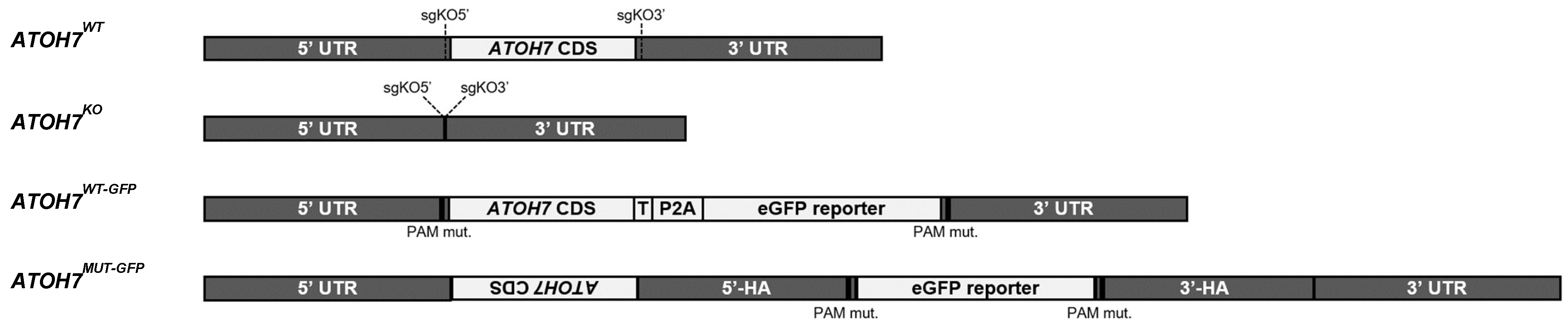
3.1. CRISPR/Cas9 Gene Editing of Human iPSCs
3.2. Mutant Human iPSC-Derived Organoids Recapitulate Axon Deficiency
3.3. The Majority of ATOH7 Targets Are Distal Enhancer Sites
3.4. Top Annotated Genes Are Involved in RGC Differentiation, Axonogenesis, and Notch Signaling
3.5. Motif Analysis Reveals Enrichment of Loci Bound by RPC, RGC, and PR-Associated Factors
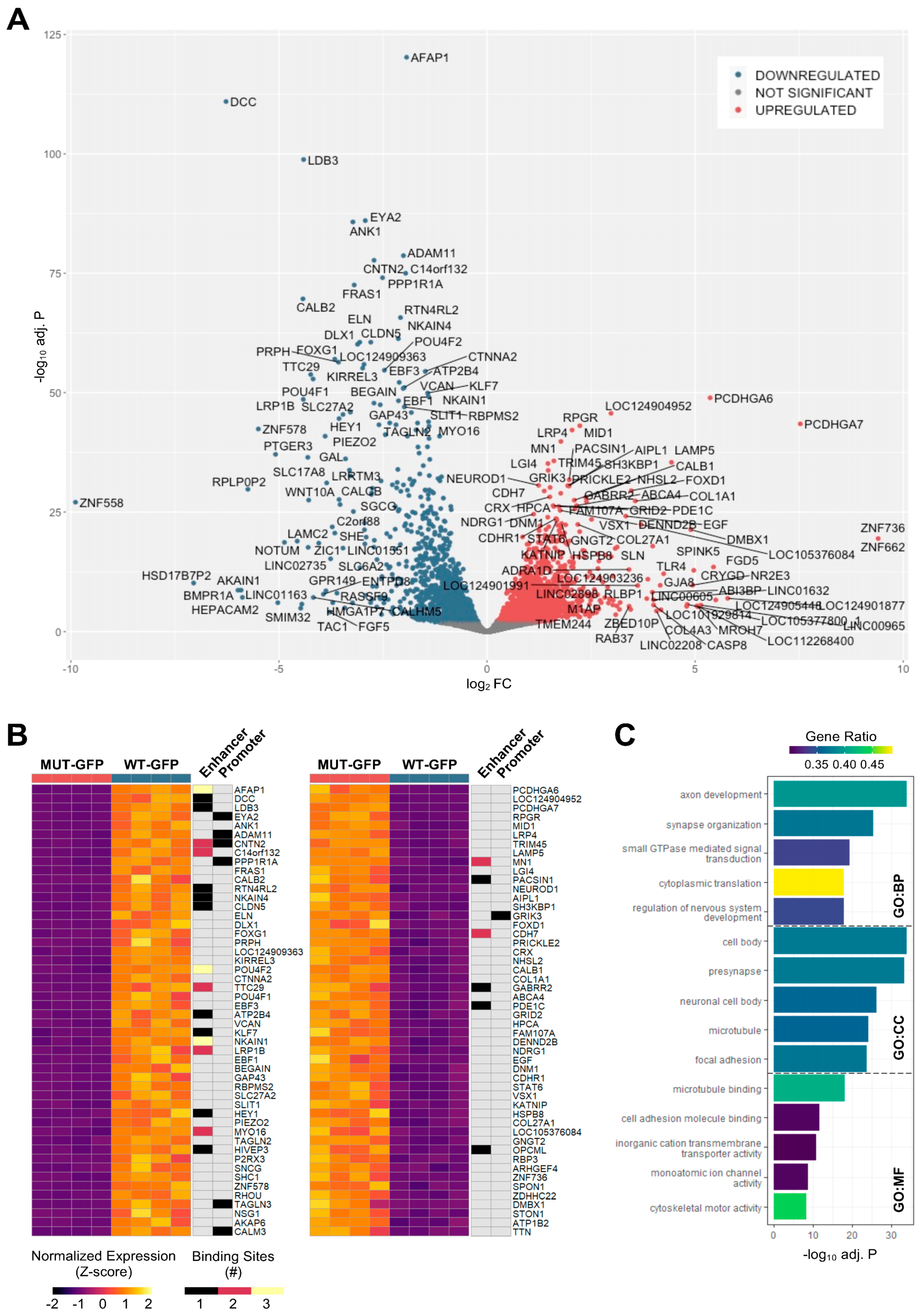
3.6. Differentially Expressed Genes in Reporter Cells Reflect ATOH7 Target Gene Categories
3.7. Loss of ATOH7 Causes Shift in Cell Type Composition in Human Retinal Organoids
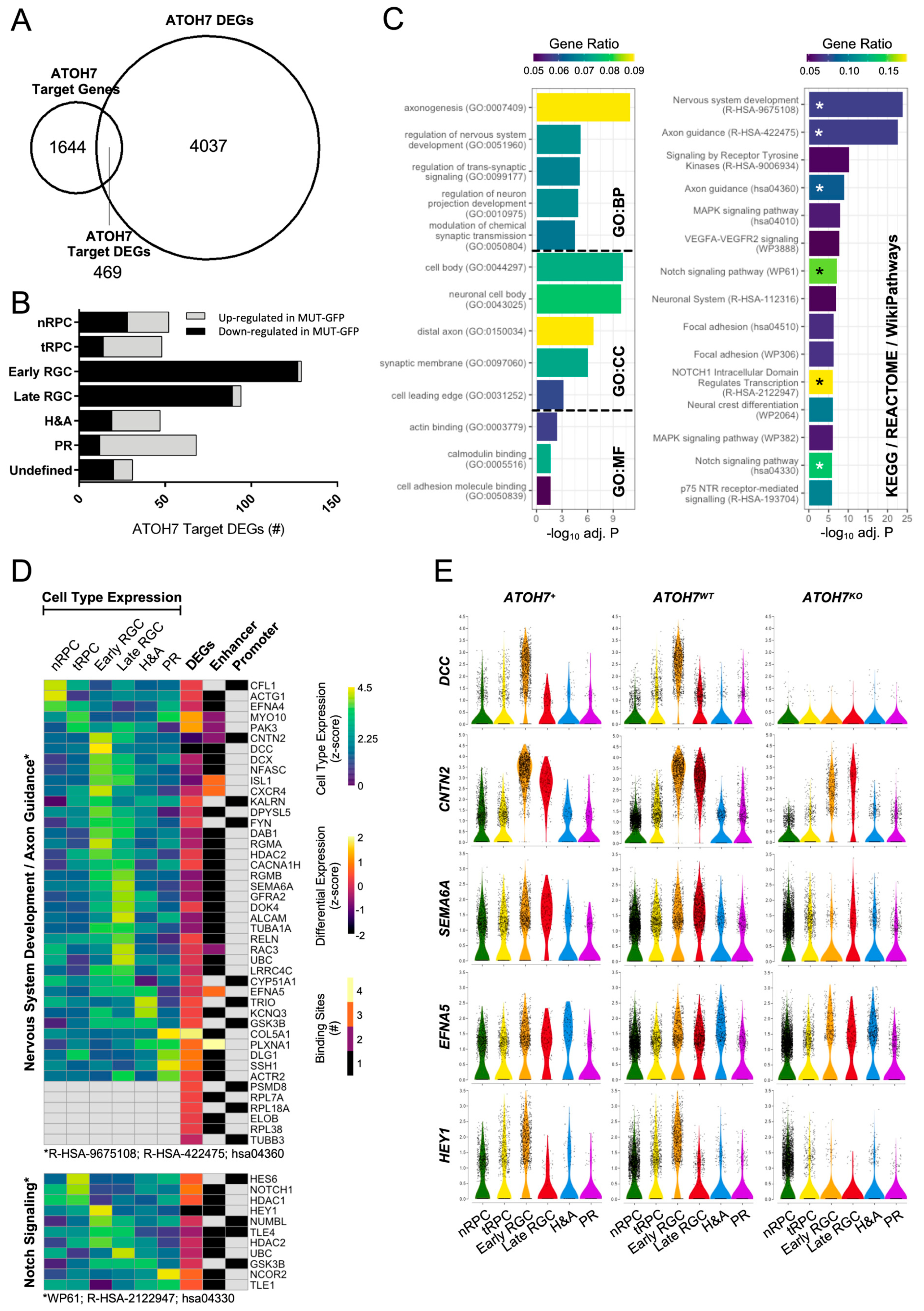
3.8. Differentially Expressed ATOH7 Target Genes and Pathway Enrichment
3.9. Expression of PHPV/EVR-Related Gene NDP Is Reduced in ATOH7 Mutants
4. Discussion
4.1. ATOH7 Is Directly Involved in Axon Development
4.2. ATOH7 Regulates Pluripotency and Differentiation through Notch Signaling
4.3. Cell Fate Shift in ATOH7-Deficient Human Retinal Organoids
4.4. In Search of ATOH7-Dependent Secreted Proteins Regulating Retinal Development
4.5. ATOH7-Associated Disease May Be a Result of Norrin Deficiency
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marquardt, T.; Gruss, P. Generating Neuronal Diversity in the Retina: One for Nearly All. Trends Neurosci. 2002, 25, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Guillemot, F. Vertebrate BHLH Genes and the Determination of Neuronal Fates. Exp. Cell Res. 1999, 253, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Perron, M.; Harris, W.A. Determination of Vertebrate Retinal Progenitor Cell Fate by the Notch Pathway and Basic Helix-Loop-Helix Transcription Factors. Cell. Mol. Life Sci. 2000, 57, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.W. Requirement for Math5 in the Development of Retinal Ganglion Cells. Genes. Dev. 2001, 15, 24–29. [Google Scholar] [CrossRef]
- Brown, N.L.; Kanekar, S.; Vetter, M.L.; Tucker, P.K.; Gemza, D.L.; Glaser, T. Math5 Encodes a Murine Basic Helix-Loop-Helix Transcription Factor Expressed during Early Stages of Retinal Neurogenesis. Development 1998, 125, 4821–4833. [Google Scholar] [CrossRef]
- Kanekar, S.; Perron, M.; Dorsky, R.; Harris, W.A.; Jan, L.Y.; Jan, Y.N.; Vetter, M.L. Xath5 Participates in a Network of BHLH Genes in the Developing Xenopus Retina. Neuron 1997, 19, 981–994. [Google Scholar] [CrossRef]
- Masai, I.; Stemple, D.L.; Okamoto, H.; Wilson, S.W. Midline Signals Regulate Retinal Neurogenesis in Zebrafish. Neuron 2000, 27, 251–263. [Google Scholar] [CrossRef]
- Liu, W.; Mo, Z.; Xiang, M. The Ath5 Proneural Genes Function Upstream of Brn3 POU Domain Transcription Factor Genes to Promote Retinal Ganglion Cell Development. Proc. Natl. Acad. Sci. USA 2001, 98, 1649–1654. [Google Scholar] [CrossRef]
- Matter-Sadzinski, L.; Matter, J.M.; Ong, M.T.; Hernandez, J.; Ballivet, M. Specification of Neurotransmitter Receptor Identity in Developing Retina: The Chick ATH5 Promoter Integrates the Positive and Negative Effects of Several BHLH Proteins. Development 2001, 128, 217–231. [Google Scholar] [CrossRef]
- Ghiasvand, N.M.; Shirzad, E.; Naghavi, M.; Mahdavi, M.R.V. High Incidence of Autosomal Recessive Nonsyndromal Congenital Retinal Nonattachment (NCRNA) in an Iranian Founding Population. Am. J. Med. Genet. 1998, 78, 226–232. [Google Scholar] [CrossRef]
- Ghiasvand, N.M.; Rudolph, D.D.; Mashayekhi, M.; Brzezinski, J.A.; Goldman, D.; Glaser, T. Deletion of a Remote Enhancer near ATOH7 Disrupts Retinal Neurogenesis, Causing NCRNA Disease. Nat. Neurosci. 2011, 14, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.; Logan, C.V.; McKibbin, M.; Sheridan, E.; Elçioglu, N.H.; Yenice, O.; Parry, D.A.; Fernandez-Fuentes, N.; Abdelhamed, Z.I.A.; Al-Maskari, A.; et al. Next Generation Sequencing Identifies Mutations in Atonal Homolog 7 (ATOH7) in Families with Global Eye Developmental Defects. Hum. Mol. Genet. 2012, 21, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Prasov, L.; Masud, T.; Khaliq, S.; Mehdi, S.Q.; Abid, A.; Oliver, E.R.; Silva, E.D.; Lewanda, A.; Brodsky, M.C.; Borchert, M.; et al. ATOH7 Mutations Cause Autosomal Recessive Persistent Hyperplasia of the Primary Vitreous. Hum. Mol. Genet. 2012, 21, 3681–3694. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Matsushita, I.; Tahira, T.; Uchio, E.; Kusaka, S. Mutations in ATOH7 Gene in Patients with Nonsyndromic Congenital Retinal Nonattachment and Familial Exudative Vitreoretinopathy. Ophthalmic Genet. 2016, 37, 462–464. [Google Scholar] [CrossRef] [PubMed]
- Keser, V.; Khan, A.; Siddiqui, S.; Lopez, I.; Ren, H.; Qamar, R.; Nadaf, J.; Majewski, J.; Chen, R.; Koenekoop, R.K. The Genetic Causes of Nonsyndromic Congenital Retinal Detachment: A Genetic and Phenotypic Study of Pakistani Families. Investig. Opthalmol. Vis. Sci. 2017, 58, 1028. [Google Scholar] [CrossRef] [PubMed]
- Atac, D.; Koller, S.; Hanson, J.V.M.; Feil, S.; Tiwari, A.; Bahr, A.; Baehr, L.; Magyar, I.; Kottke, R.; Gerth-Kahlert, C.; et al. Atonal Homolog 7 (ATOH7) Loss-of-Function Mutations in Predominant Bilateral Optic Nerve Hypoplasia. Hum. Mol. Genet. 2020, 29, 132–148. [Google Scholar] [CrossRef] [PubMed]
- Atac, D.; Mohn, L.; Feil, S.; Maggi, K.; Haenni, D.; Seebauer, B.; Koller, S.; Berger, W. Functional Characterization of an In-Frame Deletion in the Basic Domain of the Retinal Transcription Factor ATOH7. Int. J. Mol. Sci. 2022, 23, 1053. [Google Scholar] [CrossRef] [PubMed]
- Warburg, M. Heterogeneity of Congenital Retinal Non-Attachment, Falciform Folds and Retinal Dysplasia. Hum. Hered. 1976, 26, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Warburg, M. Retinal Malformations: Aetiological Heterogeneity and Morphological Similarity in Congenital Retinal Non-Attachment and Falciform Folds. Trans. Ophthalmol. Soc. UK 1979, 99, 272–283. [Google Scholar] [PubMed]
- Criswick, V.G.; Schepens, C.L. Familial Exudative Vitreoretinopathy. Am. J. Ophthalmol. 1969, 68, 578–594. [Google Scholar] [CrossRef]
- Gilmour, D.F. Familial Exudative Vitreoretinopathy and Related Retinopathies. Eye 2015, 29, 1–14. [Google Scholar] [CrossRef]
- Brzezinski, J.A.; Prasov, L.; Glaser, T. Math5 Defines the Ganglion Cell Competence State in a Subpopulation of Retinal Progenitor Cells Exiting the Cell Cycle. Dev. Biol. 2012, 365, 395–413. [Google Scholar] [CrossRef]
- Feng, L.; Xie, Z.; Ding, Q.; Xie, X.; Libby, R.T.; Gan, L. MATH5 Controls the Acquisition of Multiple Retinal Cell Fates. Mol. Brain 2010, 3, 36. [Google Scholar] [CrossRef]
- Brown, N.L.; Patel, S.; Brzezinski, J.; Glaser, T. Math5 Is Required for Retinal Ganglion Cell and Optic Nerve Formation. Development 2001, 128, 2497–2508. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Ding, K.; Pan, L.; Deng, M.; Gan, L. Math5 Determines the Competence State of Retinal Ganglion Cell Progenitors. Dev. Biol. 2003, 264, 240–254. [Google Scholar] [CrossRef]
- Brodie-Kommit, J.; Clark, B.S.; Shi, Q.; Shiau, F.; Kim, D.W.; Langel, J.; Sheely, C.; Ruzycki, P.A.; Fries, M.; Javed, A.; et al. Atoh7-Independent Specification of Retinal Ganglion Cell Identity. Sci. Adv. 2021, 7, eabe4983. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Chen, X.; Nan, N.; Bard, J.; Wu, F.; Yergeau, D.; Liu, T.; Wang, J.; Mu, X. Key Transcription Factors Influence the Epigenetic Landscape to Regulate Retinal Cell Differentiation. Nucleic Acids Res. 2023, 51, 2151–2176. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Deng, M.; Xie, X.; Gan, L. ISL1 and BRN3B Co-Regulate the Differentiation of Murine Retinal Ganglion Cells. Development 2008, 135, 1981–1990. [Google Scholar] [CrossRef]
- Li, X.-L.; Li, G.-H.; Fu, J.; Fu, Y.-W.; Zhang, L.; Chen, W.; Arakaki, C.; Zhang, J.-P.; Wen, W.; Zhao, M.; et al. Highly Efficient Genome Editing via CRISPR–Cas9 in Human Pluripotent Stem Cells Is Achieved by Transient BCL-XL Overexpression. Nucleic Acids Res. 2018, 46, 10195–10215. [Google Scholar] [CrossRef]
- Gonzalez-Cordero, A.; Kruczek, K.; Naeem, A.; Fernando, M.; Kloc, M.; Ribeiro, J.; Goh, D.; Duran, Y.; Blackford, S.J.I.; Abelleira-Hervas, L.; et al. Recapitulation of Human Retinal Development from Human Pluripotent Stem Cells Generates Transplantable Populations of Cone Photoreceptors. Stem Cell Rep. 2017, 9, 820–837. [Google Scholar] [CrossRef]
- Cuevas, E.; Holder, D.L.; Alshehri, A.H.; Tréguier, J.; Lakowski, J.; Sowden, J.C. NRL−/− Gene Edited Human Embryonic Stem Cells Generate Rod-Deficient Retinal Organoids Enriched in S-Cone-like Photoreceptors. Stem Cells 2021, 39, 414–428. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Nordin, A.; Zambanini, G.; Pagella, P.; Cantù, C. The CUT&RUN Suspect List of Problematic Regions of the Genome. Genome Biol. 2023, 24, 185. [Google Scholar] [CrossRef]
- Korsunsky, I.; Millard, N.; Fan, J.; Slowikowski, K.; Zhang, F.; Wei, K.; Baglaenko, Y.; Brenner, M.; Loh, P.; Raychaudhuri, S. Fast, Sensitive and Accurate Integration of Single-Cell Data with Harmony. Nat. Methods 2019, 16, 1289–1296. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, S.-R.; Li, L.-H.; Park, H.-J.; Park, J.-H.; Lee, K.Y.; Kim, M.-K.; Shin, B.A.; Choi, S.-Y. High Cleavage Efficiency of a 2A Peptide Derived from Porcine Teschovirus-1 in Human Cell Lines, Zebrafish and Mice. PLoS ONE 2011, 6, e18556. [Google Scholar] [CrossRef] [PubMed]
- Skene, P.J.; Henikoff, J.G.; Henikoff, S. Targeted in Situ Genome-Wide Profiling with High Efficiency for Low Cell Numbers. Nat. Protoc. 2018, 13, 1006–1019. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Brown, J.B.; Huang, H.; Bickel, P.J. Measuring Reproducibility of High-Throughput Experiments. Ann. Appl. Stat. 2011, 5, 1752–1779. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, T.; Meyer, C.A.; Eeckhoute, J.; Johnson, D.S.; Bernstein, B.E.; Nusbaum, C.; Myers, R.M.; Brown, M.; Li, W.; et al. Model-Based Analysis of ChIP-Seq (MACS). Genome Biol. 2008, 9, R137. [Google Scholar] [CrossRef]
- Welch, R.P.; Lee, C.; Imbriano, P.M.; Patil, S.; Weymouth, T.E.; Smith, R.A.; Scott, L.J.; Sartor, M.A. ChIP-Enrich: Gene Set Enrichment Testing for ChIP-Seq Data. Nucleic Acids Res. 2014, 42, e105. [Google Scholar] [CrossRef]
- Pratt, T.; Conway, C.D.; Tian, N.M.M.-L.; Price, D.J.; Mason, J.O. Heparan Sulphation Patterns Generated by Specific Heparan Sulfotransferase Enzymes Direct Distinct Aspects of Retinal Axon Guidance at the Optic Chiasm. J. Neurosci. 2006, 26, 6911–6923. [Google Scholar] [CrossRef] [PubMed]
- Conway, C.D.; Price, D.J.; Pratt, T.; Mason, J.O. Analysis of Axon Guidance Defects at the Optic Chiasm in Heparan Sulphate Sulphotransferase Compound Mutant Mice. J. Anat. 2011, 219, 734–742. [Google Scholar] [CrossRef] [PubMed]
- Miesfeld, J.B.; Ghiasvand, N.M.; Marsh-Armstrong, B.; Marsh-Armstrong, N.; Miller, E.B.; Zhang, P.; Manna, S.K.; Zawadzki, R.J.; Brown, N.L.; Glaser, T. The Atoh7 Remote Enhancer Provides Transcriptional Robustness during Retinal Ganglion Cell Development. Proc. Natl. Acad. Sci. USA 2020, 117, 21690–21700. [Google Scholar] [CrossRef] [PubMed]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g: Profiler: A Web Server for Functional Enrichment Analysis and Conversions of Gene Lists (2019 Update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed]
- Heinz, S.; Benner, C.; Spann, N.; Bertolino, E.; Lin, Y.C.; Laslo, P.; Cheng, J.X.; Murre, C.; Singh, H.; Glass, C.K. Simple Combinations of Lineage-Determining Transcription Factors Prime Cis-Regulatory Elements Required for Macrophage and B Cell Identities. Mol. Cell 2010, 38, 576–589. [Google Scholar] [CrossRef] [PubMed]
- Wizenmann, A.; Stettler, O.; Moya, K.L. Engrailed Homeoproteins in Visual System Development. Cell Mol. Life Sci. 2015, 72, 1433–1445. [Google Scholar] [CrossRef] [PubMed]
- De Melo, J.; Du, G.; Fonseca, M.; Gillespie, L.-A.; Turk, W.J.; Rubenstein, J.L.R.; Eisenstat, D.D. Dlx1 and Dlx2 Function Is Necessary for Terminal Differentiation and Survival of Late-Born Retinal Ganglion Cells in the Developing Mouse Retina. Development 2005, 132, 311–322. [Google Scholar] [CrossRef]
- Zhang, Q.; Zagozewski, J.; Cheng, S.; Dixit, R.; Zhang, S.; de Melo, J.; Mu, X.; Klein, W.H.; Brown, N.L.; Wigle, J.T.; et al. Regulation of Brn3b by DLX1 and DLX2 Is Required for Retinal Ganglion Cell Differentiation in the Vertebrate Retina. Development 2017, 144, 1698–1711. [Google Scholar] [CrossRef] [PubMed]
- Inoue, J.; Ueda, Y.; Bando, T.; Mito, T.; Noji, S.; Ohuchi, H. The Expression of LIM-Homeobox Genes, Lhx1 and Lhx5, in the Forebrain Is Essential for Neural Retina Differentiation. Dev. Growth Differ. 2013, 55, 668–675. [Google Scholar] [CrossRef]
- Gordon, P.J.; Yun, S.; Clark, A.M.; Monuki, E.S.; Murtaugh, L.C.; Levine, E.M. Lhx2 Balances Progenitor Maintenance with Neurogenic Output and Promotes Competence State Progression in the Developing Retina. J. Neurosci. 2013, 33, 12197–12207. [Google Scholar] [CrossRef]
- Dupacova, N.; Antosova, B.; Paces, J.; Kozmik, Z. Meis Homeobox Genes Control Progenitor Competence in the Retina. Proc. Natl. Acad. Sci. USA 2021, 118, e2013136118. [Google Scholar] [CrossRef]
- Lerner, L.E.; Peng, G.-H.; Gribanova, Y.E.; Chen, S.; Farber, D.B. Sp4 Is Expressed in Retinal Neurons, Activates Transcription of Photoreceptor-Specific Genes, and Synergizes with Crx. J. Biol. Chem. 2005, 280, 20642–20650. [Google Scholar] [CrossRef]
- Mocciaro, E.; Runfola, V.; Ghezzi, P.; Pannese, M.; Gabellini, D. DUX4 Role in Normal Physiology and in FSHD Muscular Dystrophy. Cells 2021, 10, 3322. [Google Scholar] [CrossRef]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon Provides Fast and Bias-Aware Quantification of Transcript Expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Allikmets, R.; Shroyer, N.F.; Singh, N.; Seddon, J.M.; Lewis, R.A.; Bernstein, P.S.; Peiffer, A.; Zabriskie, N.A.; Li, Y.; Hutchinson, A.; et al. Mutation of the Stargardt Disease Gene (ABCR) in Age-Related Macular Degeneration. Science 1997, 277, 1805–1807. [Google Scholar] [CrossRef]
- Meindl, A.; Dry, K.; Herrmann, K.; Manson, E.; Ciccodicola, A.; Edgar, A.; Carvalho, M.R.S.; Achatz, H.; Hellebrand, H.; Lennon, A.; et al. A Gene (RPGR) with Homology to the RCC1 Guanine Nucleotide Exchange Factor Is Mutated in X–Linked Retinitis Pigmentosa (RP3). Nat. Genet. 1996, 13, 35–42. [Google Scholar] [CrossRef]
- Morrow, E.M.; Furukawa, T.; Lee, J.E.; Cepko, C.L. NeuroD Regulates Multiple Functions in the Developing Neural Retina in Rodent. Development 1999, 126, 23–36. [Google Scholar] [CrossRef]
- Haider, N.B.; Jacobson, S.G.; Cideciyan, A.V.; Swiderski, R.; Streb, L.M.; Searby, C.; Beck, G.; Hockey, R.; Hanna, D.B.; Gorman, S.; et al. Mutation of a Nuclear Receptor Gene, NR2E3, Causes Enhanced S Cone Syndrome, a Disorder of Retinal Cell Fate. Nat. Genet. 2000, 24, 127–131. [Google Scholar] [CrossRef]
- Van der Spuy, J.; Kim, J.H.; Yu, Y.S.; Szel, A.; Luthert, P.J.; Clark, B.J.; Cheetham, M.E. The Expression of the Leber Congenital Amaurosis Protein AIPL1 Coincides with Rod and Cone Photoreceptor Development. Investig. Opthalmol. Vis. Sci. 2003, 44, 5396. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, Q.-L.; Nie, Z.; Sun, H.; Lennon, G.; Copeland, N.G.; Gilbert, D.J.; Jenkins, N.A.; Zack, D.J. Crx, a Novel Otx-like Paired-Homeodomain Protein, Binds to and Transactivates Photoreceptor Cell-Specific Genes. Neuron 1997, 19, 1017–1030. [Google Scholar] [CrossRef] [PubMed]
- Carreres, M.I.; Escalante, A.; Murillo, B.; Chauvin, G.; Gaspar, P.; Vegar, C.; Herrera, E. Transcription Factor Foxd1 Is Required for the Specification of the Temporal Retina in Mammals. J. Neurosci. 2011, 31, 5673–5681. [Google Scholar] [CrossRef] [PubMed]
- Chow, R.L.; Snow, B.; Novak, J.; Looser, J.; Freund, C.; Vidgen, D.; Ploder, L.; McInnes, R.R. Vsx1, a Rapidly Evolving Paired -like Homeobox Gene Expressed in Cone Bipolar Cells. Mech. Dev. 2001, 109, 315–322. [Google Scholar] [CrossRef]
- Lekanne Deprez, R.H.; Riegman, P.H.; Groen, N.A.; Warringa, U.L.; van Biezen, N.A.; Molijn, A.C.; Bootsma, D.; de Jong, P.J.; Menon, A.G.; Kley, N.A. Cloning and Characterization of MN1, a Gene from Chromosome 22q11, Which Is Disrupted by a Balanced Translocation in a Meningioma. Oncogene 1995, 10, 1521–1528. [Google Scholar]
- Faulkner-Jones, B.E.; Godinho, L.N.M.; Reese, B.E.; Pasquini, G.F.; Ruefli, A.; Tan, S.-S. Cloning and Expression of Mouse Cadherin-7, a Type-II Cadherin Isolated from the Developing Eye. Mol. Cell. Neurosci. 1999, 14, 1–16. [Google Scholar] [CrossRef]
- D’Souza, S.P.; Upton, B.A.; Eldred, K.C.; Glass, I.; Grover, K.; Ahmed, A.; Ngyuen, M.-T.; Gamlin, P.; Lang, R.A. Developmental Adaptation of Rod Photoreceptor Number via Photoreception in Melanopsin (OPN4) Retinal Ganglion Cells. bioRxiv 2023. [Google Scholar] [CrossRef]
- Sridhar, A.; Hoshino, A.; Finkbeiner, C.R.; Chitsazan, A.; Dai, L.; Haugan, A.K.; Eschenbacher, K.M.; Jackson, D.L.; Trapnell, C.; Bermingham-McDonogh, O.; et al. Single-Cell Transcriptomic Comparison of Human Fetal Retina, HPSC-Derived Retinal Organoids, and Long-Term Retinal Cultures. Cell Rep. 2020, 30, 1644–1659.e4. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, X.; Li, S.; Huang, X.; Qian, H.; Jin, K.; Xiang, M. Foxn4 Is a Temporal Identity Factor Conferring Mid/Late-Early Retinal Competence and Involved in Retinal Synaptogenesis. Proc. Natl. Acad. Sci. USA 2020, 117, 5016–5027. [Google Scholar] [CrossRef]
- Wang, Y.; Li, N.; Zheng, Y.; Wang, A.; Yu, C.; Song, Z.; Wang, S.; Sun, Y.; Zheng, L.; Wang, G.; et al. KIAA1217 Promotes Epithelial-Mesenchymal Transition and Hepatocellular Carcinoma Metastasis by Interacting with and Activating STAT3. Int. J. Mol. Sci. 2021, 23, 104. [Google Scholar] [CrossRef] [PubMed]
- Meindl, A.; Berger, W.; Meitinger, T.; van de Pol, D.; Achatz, H.; Dörner, C.; Haasemann, M.; Hellebrand, H.; Gal, A.; Cremers, F.; et al. Norrie Disease Is Caused by Mutations in an Extracellular Protein Resembling C–Terminal Globular Domain of Mucins. Nat. Genet. 1992, 2, 139–143. [Google Scholar] [CrossRef]
- Berger, W.; Meindl, A.; van de Pol, T.J.R.; Cremers, F.P.M.; Ropers, H.H.; Döerner, C.; Monaco, A.; Bergen, A.A.B.; Lebo, R.; Warburg, M.; et al. Isolation of a Candidate Gene for Norrie Disease by Positional Cloning. Nat. Genet. 1992, 1, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-Y.; Battinelli, E.M.; Fielder, A.; Bundey, S.; Sims, K.; Breakefield, X.O.; Craig, I.W. A Mutation in the Norrie Disease Gene (NDP) Associated with X–Linked Familial Exudative Vitreoretinopathy. Nat. Genet. 1993, 5, 180–183. [Google Scholar] [CrossRef]
- Dhingra, S.; Shears, D.J.; Blake, V.; Stewart, H.; Patel, C.K. Advanced Bilateral Persistent Fetal Vasculature Associated with a Novel Mutation in the Norrie Gene. Br. J. Ophthalmol. 2006, 90, 1324–1325. [Google Scholar] [CrossRef] [PubMed]
- Payabvash, S.; Anderson, J.S.; Nascene, D.R. Bilateral Persistent Fetal Vasculature Due to a Mutation in the Norrie Disease Protein Gene. Neuroradiol. J. 2015, 28, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Toomes, C.; Bottomley, H.M.; Jackson, R.M.; Towns, K.V.; Scott, S.; Mackey, D.A.; Craig, J.E.; Jiang, L.; Yang, Z.; Trembath, R.; et al. Mutations in LRP5 or FZD4 Underlie the Common Familial Exudative Vitreoretinopathy Locus on Chromosome 11q. Am. J. Hum. Genet. 2004, 74, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Robitaille, J.M.; Wallace, K.; Zheng, B.; Beis, M.J.; Samuels, M.; Hoskin-Mott, A.; Guernsey, D.L. Phenotypic Overlap of Familial Exudative Vitreoretinopathy (FEVR) with Persistent Fetal Vasculature (PFV) Caused by FZD4 Mutations in Two Distinct Pedigrees. Ophthalmic Genet. 2009, 30, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Collin, R.W.J.; Nikopoulos, K.; Dona, M.; Gilissen, C.; Hoischen, A.; Boonstra, F.N.; Poulter, J.A.; Kondo, H.; Berger, W.; Toomes, C.; et al. ZNF408 Is Mutated in Familial Exudative Vitreoretinopathy and Is Crucial for the Development of Zebrafish Retinal Vasculature. Proc. Natl. Acad. Sci. USA 2013, 110, 9856–9861. [Google Scholar] [CrossRef] [PubMed]
- Weiner, G.A.; Nudleman, E. Microcornea, Posterior Megalolenticonus, Persistent Fetal Vasculature, and Coloboma Syndrome Associated with a New Mutation in ZNF408. Ophthalmic Surg. Lasers Imaging Retin. 2019, 50, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Nakamura, M.; Amano, E.; Mokuno, K.; Shirai, S.; Terasaki, H. Case of Chromosome 6p25 Terminal Deletion Associated with Axenfeld-Rieger Syndrome and Persistent Hyperplastic Primary Vitreous. Am. J. Med. Genet. A 2006, 140, 503–508. [Google Scholar] [CrossRef]
- Arikawa, A.; Yoshida, S.; Yoshikawa, H.; Ishikawa, K.; Yamaji, Y.; Arita, R.-I.; Ueno, A.; Ishibashi, T. Case of Novel PITX2 Gene Mutation Associated with Peters’ Anomaly and Persistent Hyperplastic Primary Vitreous. Eye 2010, 24, 391–393. [Google Scholar] [CrossRef]
- Hasbrook, M.; Yonekawa, Y.; Van Laere, L.; Shah, A.R.; Capone, A. Bilateral Persistent Fetal Vasculature and a Chromosome 10 Mutation Including COX15. Can. J. Ophthalmol. 2017, 52, e203–e205. [Google Scholar] [CrossRef] [PubMed]
- Williamson, K.A.; Hall, H.N.; Owen, L.J.; Livesey, B.J.; Hanson, I.M.; Adams, G.G.W.; Bodek, S.; Calvas, P.; Castle, B.; Clarke, M.; et al. Recurrent Heterozygous PAX6 Missense Variants Cause Severe Bilateral Microphthalmia via Predictable Effects on DNA-Protein Interaction. Genet. Med. 2020, 22, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xiao, X.; Li, S.; Mai, G.; Zhang, Q. Novel TSPAN12 Mutations in Patients with Familial Exudative Vitreoretinopathy and Their Associated Phenotypes. Mol. Vis. 2011, 17, 1128–1135. [Google Scholar] [PubMed]
- Robitaille, J.M.; Gillett, R.M.; LeBlanc, M.A.; Gaston, D.; Nightingale, M.; Mackley, M.P.; Parkash, S.; Hathaway, J.; Thomas, A.; Ells, A.; et al. Phenotypic Overlap between Familial Exudative Vitreoretinopathy and Microcephaly, Lymphedema, and Chorioretinal Dysplasia Caused by KIF11 Mutations. JAMA Ophthalmol. 2014, 132, 1393–1399. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, J.-H.; Ko, Y.; Wang, C.; Chung, Y.-C.; Chu, K.; Liu, T.; Chao, H.; Jiang, Y.; Chen, S.; et al. Haploinsufficiency of RCBTB1 Is Associated with Coats Disease and Familial Exudative Vitreoretinopathy. Hum. Mol. Genet. 2016, 25, 1637–1647. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, Y.; Yang, L.; Han, S.; Qi, M. Identification of a Novel Splice Mutation in CTNNB1 Gene in a Chinese Family with Both Severe Intellectual Disability and Serious Visual Defects. Neurol. Sci. 2019, 40, 1701–1704. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.M.; Kannabiran, C.; Balasubramanian, D. Identification of Key Genes and Pathways in Persistent Hyperplastic Primary Vitreous of the Eye Using Bioinformatic Analysis. Front. Med. 2021, 8, 690594. [Google Scholar] [CrossRef] [PubMed]
- Freeman-Anderson, N.E.; Zheng, Y.; McCalla-Martin, A.C.; Treanor, L.M.; Zhao, Y.D.; Garfin, P.M.; He, T.-C.; Mary, M.N.; Thornton, J.D.; Anderson, C.; et al. Expression of the Arf Tumor Suppressor Gene Is Controlled by Tgfbeta2 during Development. Development 2009, 136, 2081–2089. [Google Scholar] [CrossRef] [PubMed]
- Rutland, C.S.; Mitchell, C.A.; Nasir, M.; Konerding, M.A.; Drexler, H.C.A. Microphthalmia, Persistent Hyperplastic Hyaloid Vasculature and Lens Anomalies Following Overexpression of VEGF-A188 from the AlphaA-Crystallin Promoter. Mol. Vis. 2007, 13, 47–56. [Google Scholar]
- Lin, S.; Liu, W.; Chen, C.-L.; Sun, D.; Hu, J.-X.; Li, L.; Ye, J.; Mei, L.; Xiong, W.-C. Neogenin-Loss in Neural Crest Cells Results in Persistent Hyperplastic Primary Vitreous Formation. J. Mol. Cell Biol. 2020, 12, 17–31. [Google Scholar] [CrossRef]
- Deiner, M.S.; Kennedy, T.E.; Fazeli, A.; Serafini, T.; Tessier-Lavigne, M.; Sretavan, D.W. Netrin-1 and DCC Mediate Axon Guidance Locally at the Optic Disc: Loss of Function Leads to Optic Nerve Hypoplasia. Neuron 1997, 19, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Mao, C.-A.; Pan, P.; Mu, X.; Klein, W.H. Transcriptome of Atoh7 Retinal Progenitor Cells Identifies New Atoh7 -Dependent Regulatory Genes for Retinal Ganglion Cell Formation. Dev. Neurobiol. 2014, 74, 1123–1140. [Google Scholar] [CrossRef] [PubMed]
- Gurung, S.; Asante, E.; Hummel, D.; Williams, A.; Feldman-Schultz, O.; Halloran, M.C.; Sittaramane, V.; Chandrasekhar, A. Distinct Roles for the Cell Adhesion Molecule Contactin2 in the Development and Function of Neural Circuits in Zebrafish. Mech. Dev. 2018, 152, 1–12. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, S.; Lang, R.A. Retinal Ganglion Cell Interactions Shape the Developing Mammalian Visual System. Development 2020, 147, dev196535. [Google Scholar] [CrossRef] [PubMed]
- Segarra, M.; Ohnuki, H.; Maric, D.; Salvucci, O.; Hou, X.; Kumar, A.; Li, X.; Tosato, G. Semaphorin 6A Regulates Angiogenesis by Modulating VEGF Signaling. Blood 2012, 120, 4104–4115. [Google Scholar] [CrossRef] [PubMed]
- Ciossek, T.; Monschau, B.; Kremoser, C.; Löschinger, J.; Lang, S.; Müller, B.K.; Bonhoeffer, F.; Drescher, U. Eph Receptor-Ligand Interactions Are Necessary for Guidance of Retinal Ganglion Cell Axons in Vitro. Eur. J. Neurosci. 1998, 10, 1574–1580. [Google Scholar] [CrossRef] [PubMed]
- Son, A.I.; Sheleg, M.; Cooper, M.A.; Sun, Y.; Kleiman, N.J.; Zhou, R. Formation of Persistent Hyperplastic Primary Vitreous in Ephrin-A5-/- Mice. Investig. Ophthalmol. Vis. Sci. 2014, 55, 1594–1606. [Google Scholar] [CrossRef] [PubMed]
- Lutty, G.A.; McLeod, D.S. Development of the Hyaloid, Choroidal and Retinal Vasculatures in the Fetal Human Eye. Prog. Retin. Eye Res. 2018, 62, 58–76. [Google Scholar] [CrossRef]
- Du, W.; Yu, W.; Huang, L.; Zhao, M.; Li, X. Ephrin-A4 Is Involved in Retinal Neovascularization by Regulating the VEGF Signaling Pathway. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1990–1998. [Google Scholar] [CrossRef] [PubMed]
- Gharahkhani, P.; Burdon, K.P.; Fogarty, R.; Sharma, S.; Hewitt, A.W.; Martin, S.; Law, M.H.; Cremin, K.; Bailey, J.N.C.; Loomis, S.J.; et al. Common Variants near ABCA1, AFAP1 and GMDS Confer Risk of Primary Open-Angle Glaucoma. Nat. Genet. 2014, 46, 1120–1125. [Google Scholar] [CrossRef]
- Poulopoulos, A.; Murphy, A.J.; Ozkan, A.; Davis, P.; Hatch, J.; Kirchner, R.; Macklis, J.D. Subcellular Transcriptomes and Proteomes of Developing Axon Projections in the Cerebral Cortex. Nature 2019, 565, 356–360. [Google Scholar] [CrossRef]
- Rybnikova, E.; Kärkkäinen, I.; Pelto-Huikko, M.; Huovila, A.-P.J. Developmental Regulation and Neuronal Expression of the Cellular Disintegrin ADAM11 Gene in Mouse Nervous System. Neuroscience 2002, 112, 921–934. [Google Scholar] [CrossRef]
- Blech-Hermoni, Y.; Subedi, K.; Silver, M.; Jensen, L.; Coscia, S.; Kates, M.M.; Zhao, Y.; Raley, C.; Edwards, N.; Tran, B.; et al. Expression of LIM Domain-Binding 3 (LDB3), a Striated Muscle Z-Band Alternatively Spliced PDZ-Motif Protein in the Nervous System. Sci. Rep. 2023, 13, 270. [Google Scholar] [CrossRef]
- Henrique, D.; Hirsinger, E.; Adam, J.; Le Roux, I.; Pourquié, O.; Ish-Horowicz, D.; Lewis, J. Maintenance of Neuroepithelial Progenitor Cells by Delta–Notch Signalling in the Embryonic Chick Retina. Curr. Biol. 1997, 7, 661–670. [Google Scholar] [CrossRef]
- Xie, J.; Lin, L.S.; Huang, X.Y.; Gan, R.H.; Ding, L.C.; Su, B.H.; Zhao, Y.; Lu, Y.G.; Zheng, D.L. The NOTCH1-HEY1 Pathway Regulates Self-Renewal and Epithelial-Mesenchymal Transition of Salivary Adenoid Cystic Carcinoma Cells. Int. J. Biol. Sci. 2020, 16, 598. [Google Scholar] [CrossRef]
- Sakamoto, M.; Hirata, H.; Ohtsuka, T.; Bessho, Y.; Kageyama, R. The Basic Helix-Loop-Helix Genes Hesr1/Hey1 and Hesr2/Hey2 Regulate Maintenance of Neural Precursor Cells in the Brain. J. Biol. Chem. 2003, 278, 44808–44815. [Google Scholar] [CrossRef]
- Hufnagel, R.B.; Riesenberg, A.N.; Quinn, M.; Brzezinski, J.A.; Glaser, T.; Brown, N.L. Heterochronic Misexpression of Ascl1 in the Atoh7 Retinal Cell Lineage Blocks Cell Cycle Exit. Mol. Cell. Neurosci. 2013, 54, 108–120. [Google Scholar] [CrossRef]
- Inoue, T.; Hojo, M.; Bessho, Y.; Tano, Y.; Lee, J.E.; Kageyama, R. Math3 and NeuroD Regulate Amacrine Cell Fate Specification in the Retina. Development 2002, 129, 831–842. [Google Scholar] [CrossRef]
- Yan, R.-T.; Liang, L.; Ma, W.; Li, X.; Xie, W.; Wang, S.-Z. Neurogenin1 Effectively Reprograms Cultured Chick Retinal Pigment Epithelial Cells to Differentiate toward Photoreceptors. J. Comp. Neurol. 2010, 518, 526–546. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.B.; Logan, M.A.; Aldiri, I.; Roberts, J.M.; Steele, M.; Vetter, M.L. C8orf46 Homolog Encodes a Novel Protein Vexin That Is Required for Neurogenesis in Xenopus Laevis. Dev. Biol. 2018, 437, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Anderson, R.E.; Agbaga, M.-P.; Rucker, E.B.; Le, Y.-Z. Loss of BCL-X L in Rod Photoreceptors: Increased Susceptibility to Bright Light Stress. Investig. Opthalmol. Vis. Sci. 2006, 47, 5583. [Google Scholar] [CrossRef] [PubMed]
- Joseph, R.M.; Li, T. Overexpression of Bcl-2 or Bcl-XL Transgenes and Photoreceptor Degeneration. Investig. Ophthalmol. Vis. Sci. 1996, 37, 2434–2446. [Google Scholar]
- Ou, K.; Mertsch, S.; Theodoropoulou, S.; Wu, J.; Liu, J.; Copland, D.A.; Schrader, S.; Liu, L.; Dick, A.D. Restoring Retinal Neurovascular Health via Substance P. Exp. Cell Res. 2019, 380, 115–123. [Google Scholar] [CrossRef]
- Chen, J.; Stahl, A.; Krah, N.M.; Seaward, M.R.; Dennison, R.J.; Sapieha, P.; Hua, J.; Hatton, C.J.; Juan, A.M.; Aderman, C.M.; et al. Wnt Signaling Mediates Pathological Vascular Growth in Proliferative Retinopathy. Circulation 2011, 124, 1871–1881. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Nakamura, R.E.I.; Mohamed, O.; Dufort, D.; Hackam, A.S. Characterization of Wnt Signaling during Photoreceptor Degeneration. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5733–5741. [Google Scholar] [CrossRef] [PubMed]
- Kakugawa, S.; Langton, P.F.; Zebisch, M.; Howell, S.; Chang, T.-H.; Liu, Y.; Feizi, T.; Bineva, G.; O’Reilly, N.; Snijders, A.P.; et al. Notum Deacylates Wnt Proteins to Suppress Signalling Activity. Nature 2015, 519, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, C.-H.; Huang, S.; Chen, J. Wnt Signaling in Vascular Eye Diseases. Prog. Retin. Eye Res. 2019, 70, 110–133. [Google Scholar] [CrossRef]
- Kassumeh, S.; Weber, G.; Nobl, M.; Priglinger, S.; Ohlmann, A. The Neuroprotective Role of Wnt Signaling in the Retina. Neural Regen. Res. 2021, 16, 1524. [Google Scholar] [CrossRef] [PubMed]
- Anchan, R.M.; Reh, T.A.; Angello, J.; Balliet, A.; Walker, M. EGF and TGF-α Stimulate Retinal Neuroepithelial Cell Proliferation in Vitro. Neuron 1991, 6, 923–936. [Google Scholar] [CrossRef]
- Zhang, J.; Cui, C.; Xu, H. Downregulation of MiR-145-5p Elevates Retinal Ganglion Cell Survival to Delay Diabetic Retinopathy Progress by Targeting FGF5. Biosci. Biotechnol. Biochem. 2019, 83, 1655–1662. [Google Scholar] [CrossRef]
- Moreno-Miralles, I.; Ren, R.; Moser, M.; Hartnett, M.E.; Patterson, C. Bone Morphogenetic Protein Endothelial Cell Precursor-Derived Regulator Regulates Retinal Angiogenesis in Vivo in a Mouse Model of Oxygen-Induced Retinopathy. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2216–2222. [Google Scholar] [CrossRef]
- Scruggs, B.A.; Reding, M.Q.; Schimmenti, L.A. NDP-Related Retinopathies; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Meitinger, T.; Meindl, A.; Bork, P.; Rost, B.; Sander, C.; Haasemann, M.; Murken, J. Molecular Modelling of the Norrie Disease Protein Predicts a Cystine Knot Growth Factor Tertiary Structure. Nat. Genet. 1993, 5, 376–380. [Google Scholar] [CrossRef]
- Richter, M.; Gottanka, J.; May, C.A.; Welge-Lüssen, U.; Berger, W.; Lütjen-Drecoll, E. Retinal Vasculature Changes in Norrie Disease Mice. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2450–2457. [Google Scholar]
- Berger, W.; van de Pol, D.; Bachner, D.; Oerlemans, F.; Winkens, H.; Hameister, H.; Wieringa, B.; Hendriks, W.; Ropers, H.-H. An Animal Model for Norrie Disease (ND): Gene Targeting of the Mouse Nd Gene. Hum. Mol. Genet. 1996, 5, 51–59. [Google Scholar] [CrossRef]
- Ohlmann, A.V.; Adamek, E.; Ohlmann, A.; Lütjen-Drecoll, E. Norrie Gene Product Is Necessary for Regression of Hyaloid Vessels. Investig. Ophthalmol. Vis. Sci. 2004, 45, 2384–2390. [Google Scholar] [CrossRef]
- Hartzer, M.K.; Cheng, M.; Liu, X.; Shastry, B.S. Localization of the Norrie Disease Gene MRNA by in Situ Hybridization. Brain Res. Bull. 1999, 49, 355–358. [Google Scholar] [CrossRef]
- Ye, X.; Wang, Y.; Cahill, H.; Yu, M.; Badea, T.C.; Smallwood, P.M.; Peachey, N.S.; Nathans, J. Norrin, Frizzled-4, and Lrp5 Signaling in Endothelial Cells Controls a Genetic Program for Retinal Vascularization. Cell 2009, 139, 285–298. [Google Scholar] [CrossRef]
- McNeill, B.; Mazerolle, C.; Bassett, E.A.; Mears, A.J.; Ringuette, R.; Lagali, P.; Picketts, D.J.; Paes, K.; Rice, D.; Wallace, V.A. Hedgehog Regulates Norrie Disease Protein to Drive Neural Progenitor Self-Renewal. Hum. Mol. Genet. 2013, 22, 1005–1016. [Google Scholar] [CrossRef]
- Ohlmann, A.; Tamm, E.R. Norrin: Molecular and Functional Properties of an Angiogenic and Neuroprotective Growth Factor. Prog. Retin. Eye Res. 2012, 31, 243–257. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | Known Motif | Name | p-Value | FDR | % Targets | % BG |
|---|---|---|---|---|---|---|
| 1 |  | Atoh1(bHLH)/Cerebellum-Atoh1-ChIP-Seq(GSE22111)/Homer | 1 × 10−402 | 0.000 | 66.6 | 15.3 |
| 2 |  | NeuroD1(bHLH)/Islet-NeuroD1-ChIP-Seq(GSE30298)/Homer | 1 × 10−396 | 0.000 | 57.1 | 10.4 |
| 3 |  | BHLHA15(bHLH)/NIH3T3-BHLHB8.HA-ChIP-Seq(GSE119782)/Homer | 1 × 10−368 | 0.000 | 69.9 | 19.0 |
| 4 |  | NeuroG2(bHLH)/Fibroblast-NeuroG2-ChIP-Seq(GSE75910)/Homer | 1 × 10−340 | 0.000 | 68.1 | 19.2 |
| 5 |  | Twist2(bHLH)/Myoblast-Twist2.Ty1-ChIP-Seq(GSE127998)/Homer | 1 × 10−333 | 0.000 | 73.3 | 23.5 |
| Rank | De Novo Motif | p-Value | % Targets | % BG | Best Motif Match | Score | Similar Motif Matches |
|---|---|---|---|---|---|---|---|
| 1 |  | 1 × 10−407 | 61.7 | 12.3 | NeuroD1(bHLH)/Islet-NeuroD1-ChIP-Seq(GSE30298)/Homer | 0.976 | NeuroG2 (0.97); Atoh1 (0.95); NEUROD1 (0.95); NEUROG2 (0.94); Olig2 (0.93); Twist2 (0.91); BHLHA15 (0.91); Ascl1 (0.90) |
| 2 |  | 1 × 10−85 | 47.0 | 22.9 | En1(Homeobox)/SUM149-EN1-ChIP-Seq(GSE120957)/Homer | 0.942 | DLX2 (0.94); Lhx8 (0.93); VSX1 (0.93); Lhx2 (0.92); DLX1 (0.92); DLX5 (0.92); Lhx1 (0.92); DRGX (0.92); LHX9 (0.92) |
| 3 |  | 1 × 10−66 | 35.2 | 16.0 | MEIS2/MA0774.1/Jaspar | 0.922 | Achi (0.90); MEIS3 (0.89); hth (0.88); vis (0.88); Pknox2 (0.87); Vis (0.87); Tgif1 (0.87); Hth (0.87) |
| 4 |  | 1 × 10−49 | 11.5 | 2.80 | Sp2(Zf)/HEK293-Sp2.eGFP-ChIP-Seq(Encode)/Homer | 0.963 | KLF15 (0.94); Sp1 (0.94); GC-box (0.93); KLF1 (0.90); Sp5 (0.90); KLF5 (0.89); btd (0.88); Klf7 (0.88); Bcl6b (0.88) |
| 5 |  | 1 × 10−21 | 8.12 | 2.82 | DUX4(Homeobox)/Myoblasts-DUX4.V5-ChIP-Seq(GSE75791)/Homer | 0.673 | Duxbl (0.67); NFYA (0.66); DUXA (0.65); ONECUT1 (0.65); NFY (0.65); NFYC (0.65); CCAAT-box (0.65); ceh-48 (0.64) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atac, D.; Maggi, K.; Feil, S.; Maggi, J.; Cuevas, E.; Sowden, J.C.; Koller, S.; Berger, W. Identification and Characterization of ATOH7-Regulated Target Genes and Pathways in Human Neuroretinal Development. Cells 2024, 13, 1142. https://doi.org/10.3390/cells13131142
Atac D, Maggi K, Feil S, Maggi J, Cuevas E, Sowden JC, Koller S, Berger W. Identification and Characterization of ATOH7-Regulated Target Genes and Pathways in Human Neuroretinal Development. Cells. 2024; 13(13):1142. https://doi.org/10.3390/cells13131142
Chicago/Turabian StyleAtac, David, Kevin Maggi, Silke Feil, Jordi Maggi, Elisa Cuevas, Jane C. Sowden, Samuel Koller, and Wolfgang Berger. 2024. "Identification and Characterization of ATOH7-Regulated Target Genes and Pathways in Human Neuroretinal Development" Cells 13, no. 13: 1142. https://doi.org/10.3390/cells13131142
APA StyleAtac, D., Maggi, K., Feil, S., Maggi, J., Cuevas, E., Sowden, J. C., Koller, S., & Berger, W. (2024). Identification and Characterization of ATOH7-Regulated Target Genes and Pathways in Human Neuroretinal Development. Cells, 13(13), 1142. https://doi.org/10.3390/cells13131142