
Development of a Highly Permissive Mandarin Fish (Siniperca chuatsi) Kidney Cell Line for Mandarin Fish Ranavirus Using a Single-Cell Cloning Method

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Development of SCK Cells
2.2. Generating Monoclonal Cell Lines
2.3. Virus Strain
2.4. Assessment of Cytopathic Effect
2.5. Virus Titer Assays
2.6. Transmission Electron Microscopy
2.7. MRV Challenge in Fish
2.8. Karyotype Analysis
2.9. Phylogenetic Analysis
- mito16SUnivF-CGCCTGTTTACCAAAAACAT
- mitoCOIUnivF-CGACCAATCACAAAGACATCGGCAC
- mitoCOIUnivR-TAAGAAGCATTGTAATGCCAGCAGC [20].
3. Results
3.1. Establishment and MRV Susceptibility of SCK Cell Line
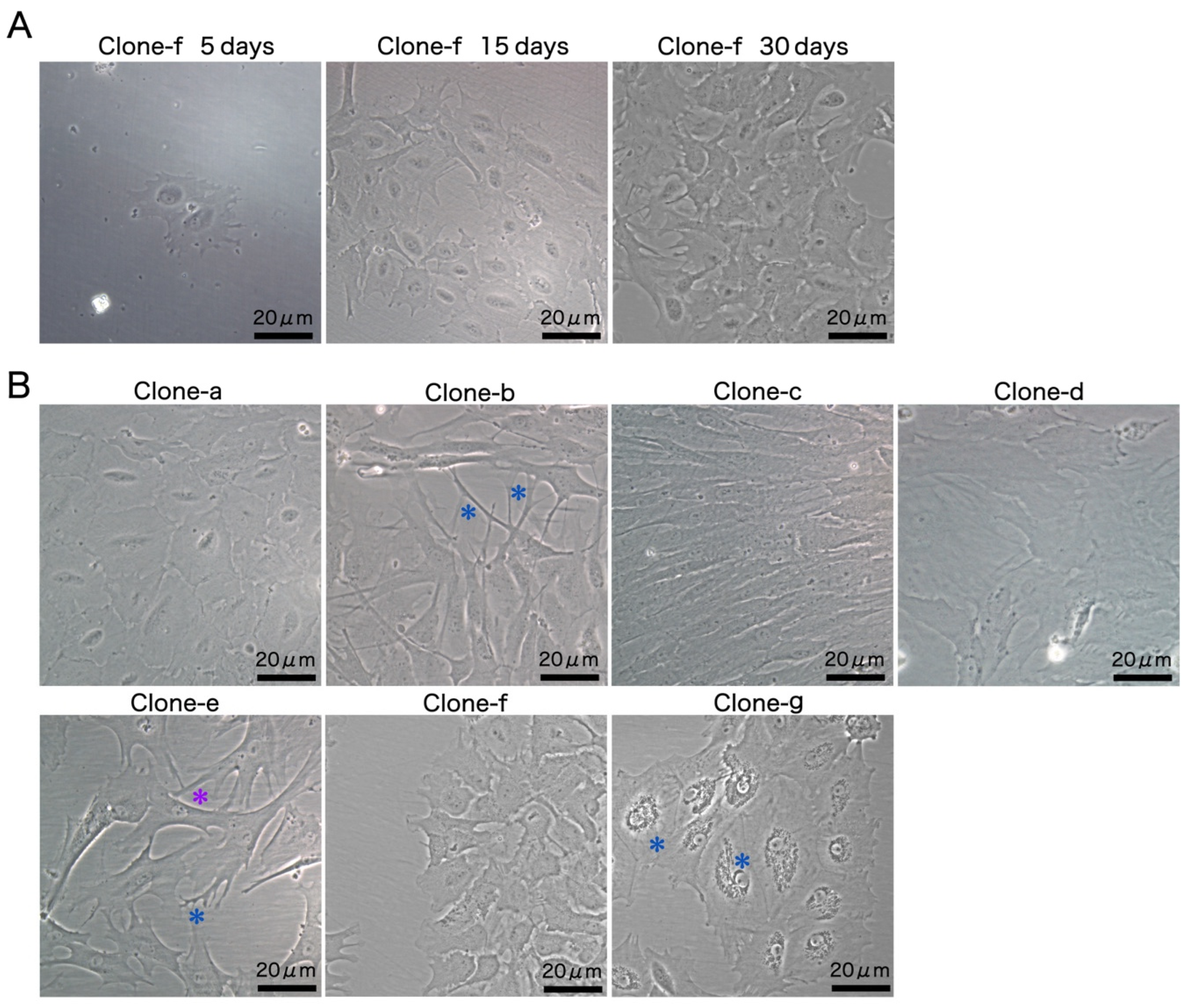
3.2. Development of Monoclonal Cell Lines
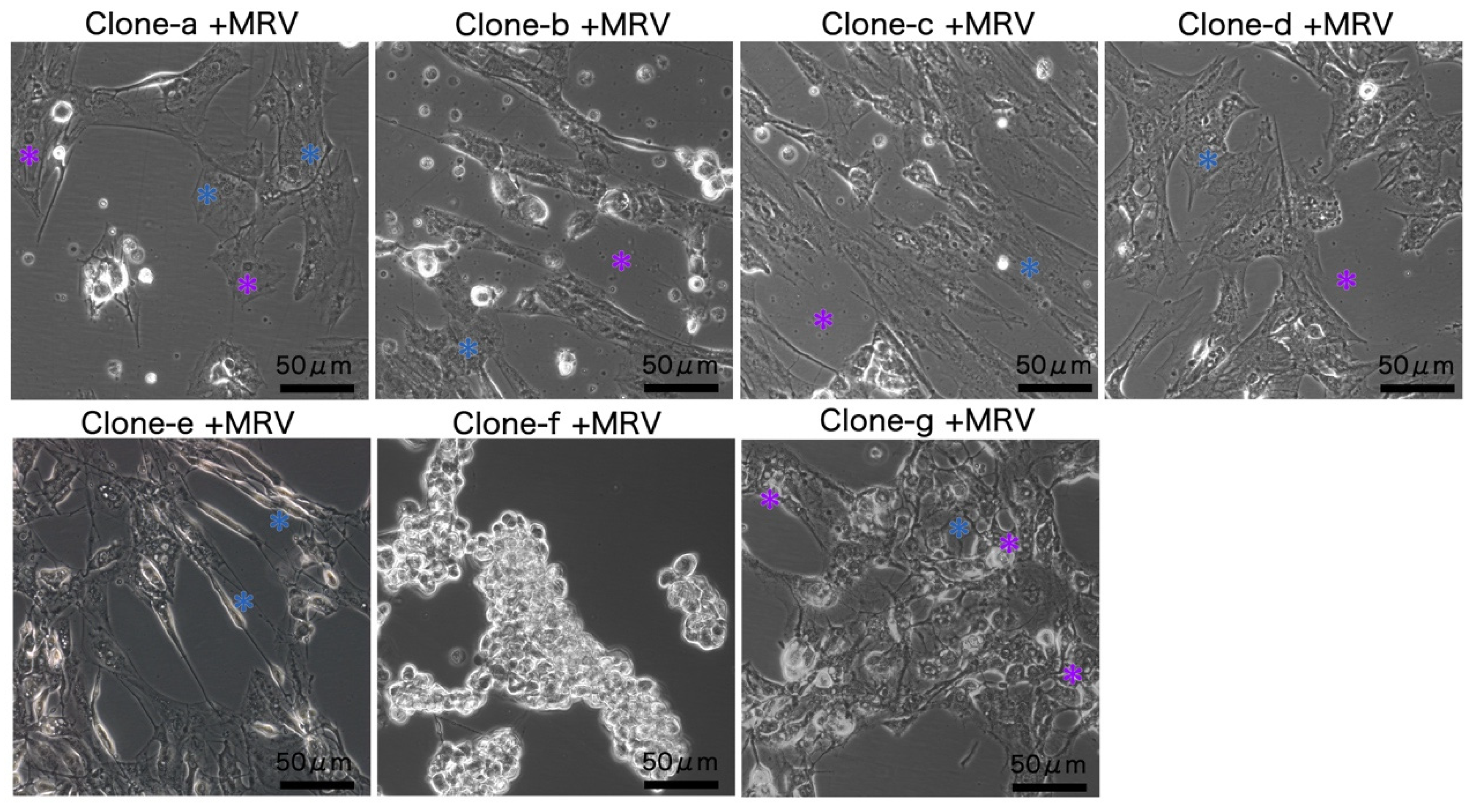
3.3. MRV Sensitivity of Monoclonal Clones
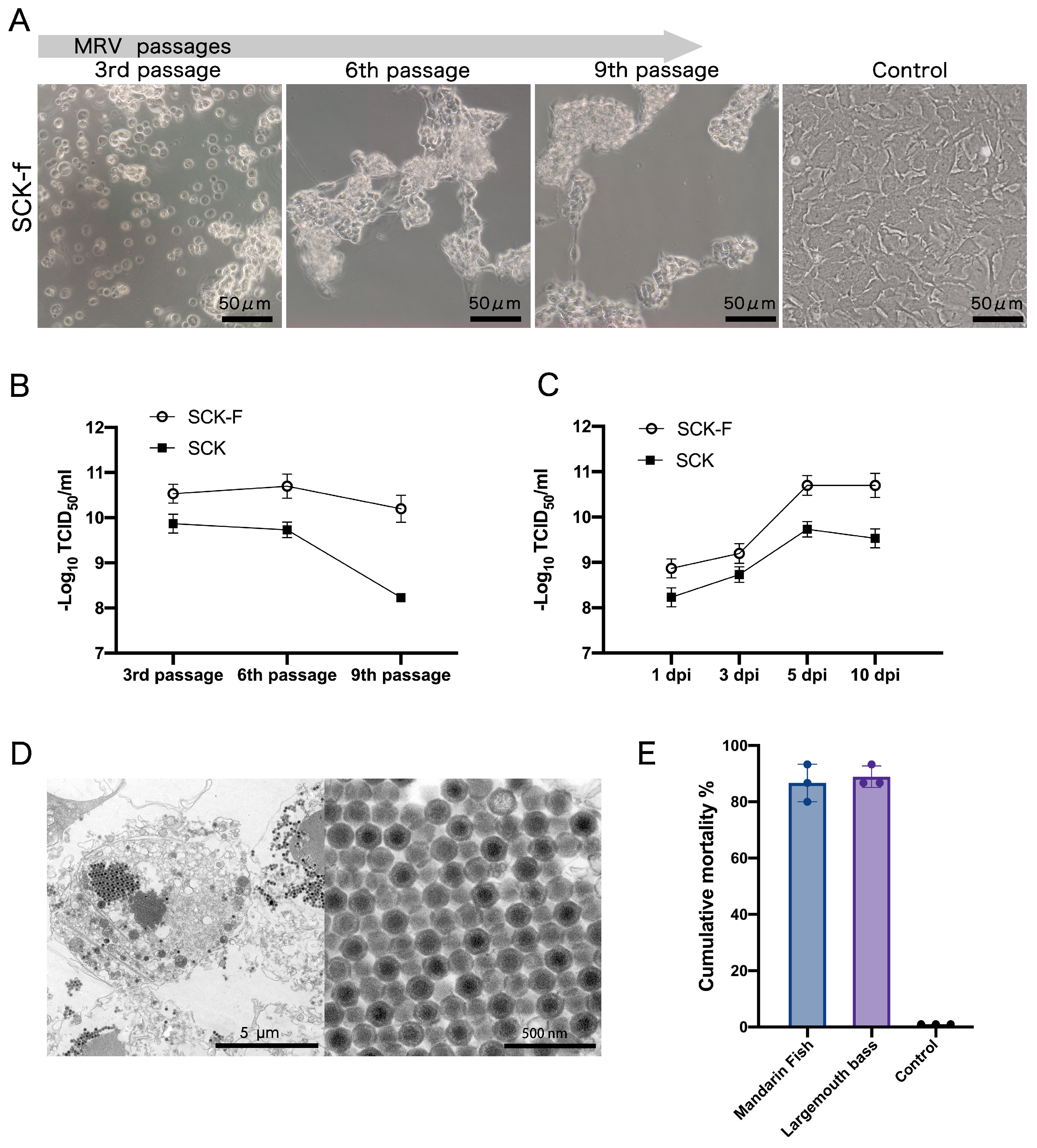
3.4. Characteristics of MRV Infection in SCK-f and In Vivo
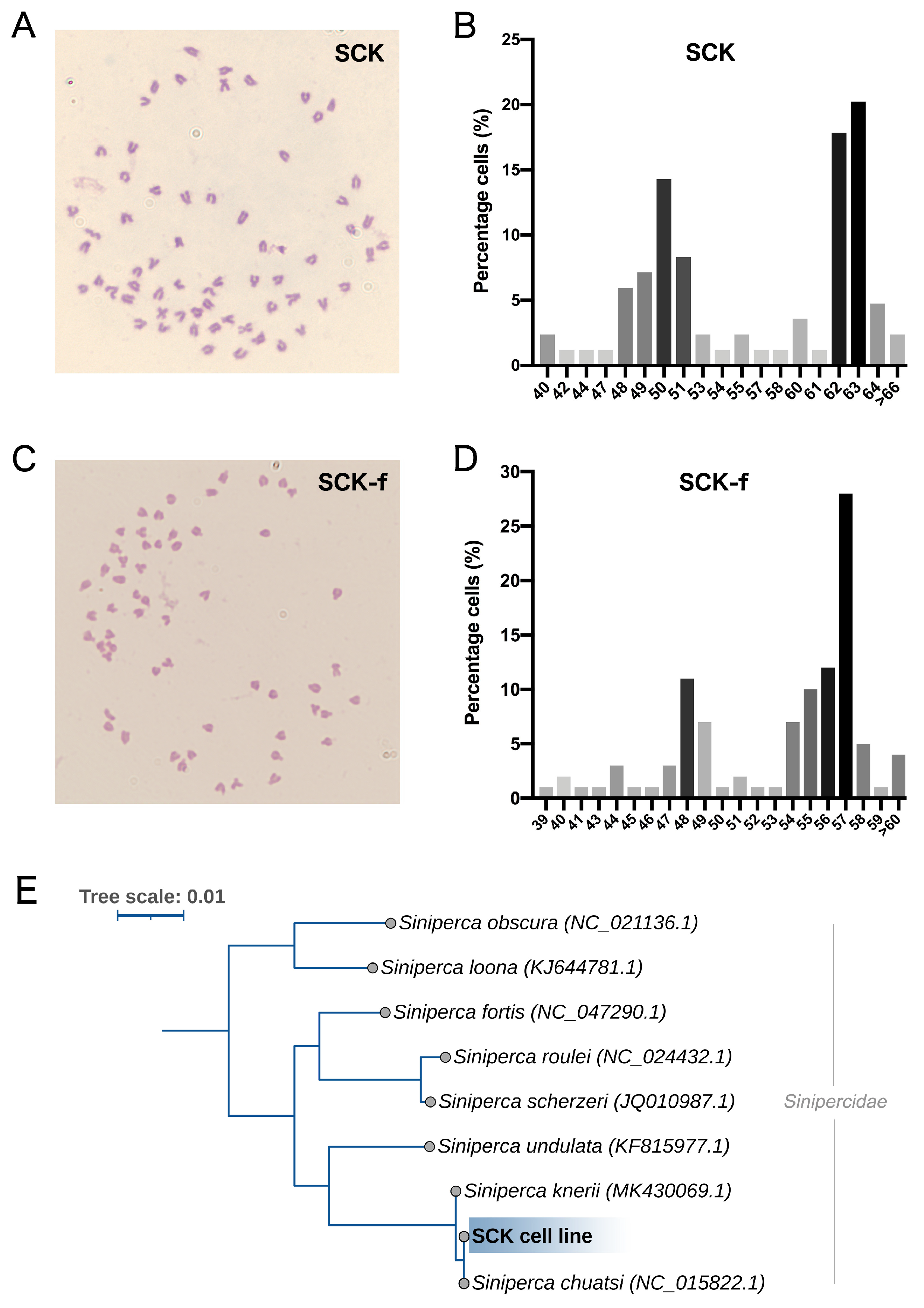
3.5. Karyotype and Phylogenetic Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- John, A.P.; John, M.G.; Helen, E.Y.; Andrew, D.N.; Scott, L. An iridovirus isolated from wild largemouth bass. J. Aquat. Anim. Health 1996, 8, 265–270. [Google Scholar]
- Grizzle, J.M.; Brunner, C.J. Review of largemouth bass virus. Fisheries 2003, 28, 10–14. [Google Scholar] [CrossRef]
- Kayansamruaj, P.; Rangsichol, A.; Dong, H.T.; Rodkhum, C.; Maita, M.; Katagiri, T.; Pirarat, N. Outbreaks of ulcerative disease associated with ranavirus infection in barcoo grunter, Scortum barcoo (McCulloch & Waite). J. Fish Dis. 2017, 40, 1341–1350. [Google Scholar] [PubMed]
- George, M.R.; John, K.R.; Mansoor, M.M.; Saravanakumar, R.; Sundar, P.; Pradeep, V. Isolation and characterization of a ranavirus from koi, Cyprinus carpio L., experiencing mass mortalities in India. J. Fish Dis. 2015, 38, 389–403. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.C.; Li, S.J.; Xie, J.; Bai, J.J.; Chen, K.C.; Ma, D.M.; Jiang, X.Y.; Lao, H.H.; Yu, L.Y. Characterization of a ranavirus isolated from cultured largemouth bass (Micropterus salmoides) in China. Aquaculture 2011, 312, 198–204. [Google Scholar] [CrossRef]
- Dong, C.; Wang, Z.; Weng, S.; He, J. Occurrence of a lethal ranavirus in hybrid mandarin (Siniperca scherzeri×Siniperca chuatsi) in Guangdong, South China. Vet. Microbiol. 2017, 203, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Duan, C.; Zhang, H.; Weng, S.; He, J.; Dong, C. Widespread outbreaks of the emerging mandarinfish ranavirus (MRV) both in natural and ISKNV-FKC vaccinated mandarinfish Siniperca chuatsi in Guangdong, South China, 2017. Aquaculture 2020, 520, 734989. [Google Scholar] [CrossRef]
- Nakajima, K.; Maeno, Y.; Kurita, J.; Inui, Y. Vaccination against red sea bream iridoviral disease in red sea bream. Fish Pathol. 1997, 32, 205–209. [Google Scholar] [CrossRef]
- Dong, C.; Xiong, X.; Luo, Y.; Weng, S.; Wang, Q.; He, J. Efficacy of a formalin-killed cell vaccine against infectious spleen and kidney necrosis virus (ISKNV) and immunoproteomic analysis of its major immunogenic proteins. Vet. Microbiol. 2013, 162, 419–428. [Google Scholar] [CrossRef]
- Piaskoski, T.O.; Plumb, J.A.; Roberts, S.R. Characterization of the largemouth bass virus in cell culture. J. Aquat. Anim. Health 1999, 11, 45–51. [Google Scholar] [CrossRef]
- Getchell, R.G.; Groocock, G.H.; Cornwell, E.R.; Schumacher, V.L.; Glasner, L.I.; Baker, B.J.; Frattini, S.A.; Wooster, G.A.; Bowser, P.R. Development and characterization of a largemouth bass cell line. J. Aquat. Anim. Health 2014, 26, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Lin, Q.; Liu, L.; Liang, H.; Huang, Z.; Li, N. The pathogenicity and biological features of Santee-Cooper Ranaviruses isolated from Chinese perch and snakehead fish. Microb. Pathog. 2017, 112, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Dong, H.; Chen, X.; Bergmann, S.M.; Yang, Y.; Wei, X.; Tong, G.; Li, H.; Yu, H.; Chen, Y. Establishment and characterization of a permanent heart cell line from largemouth bass Micropterus salmoides and its application to fish virology and immunology. Aquaculture 2022, 547, 737427. [Google Scholar] [CrossRef]
- Zhan, Z.; Fu, J.; Chen, H.; Pan, H.; Weng, S.; He, J.; Guo, C. Development and characterization of a spleen cell line from yellowfin seabream Acanthopagrus latus and its susceptibility to mandarinfish ranavirus. J. Fish Dis. 2023, 46, 1173–1181. [Google Scholar] [CrossRef] [PubMed]
- Kärber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Naunyn-Schmiedebergs Arch. Exp. Pathol. Pharmakol. 1931, 162, 480–483. [Google Scholar] [CrossRef]
- Wulff, N.H.; Tzatzaris, M.; Young, P.J. Monte carlo simulation of the spearman-kaerber TCID50. J. Clin. Bioinform. 2012, 2, 5. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.; Orenstein, J.M. Processing tissue and cells for transmission electron microscopy in diagnostic pathology and research. Nat. Protoc. 2007, 2, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
- Palumbi, S.R. The polymerase chain reaction. Mol. Syst. 1996, 205–247. [Google Scholar]
- Händeler, K.; Grzymbowski, Y.P.; Krug, P.J.; Wägele, H. Functional chloroplasts in metazoan cells—A unique evolutionary strategy in animal life. Front. Zool. 2009, 6, 28. [Google Scholar] [CrossRef]
- Zhang, J.; Hanner, R. Molecular approach to the identification of fish in the South China Sea. PLoS ONE 2012, 7, e30621. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Iyer, S.; Lobbardi, R.; Moore, J.C.; Chen, H.; Lareau, C.; Hebert, C.; Shaw, M.L.; Neftel, C.; Suva, M.L.; et al. Dissecting hematopoietic and renal cell heterogeneity in adult zebrafish at single-cell resolution using RNA sequencing. J. Exp. Med. 2017, 214, 2875–2887. [Google Scholar] [CrossRef] [PubMed]
- Mahardika, K.; Miyazaki, T. Electron microscopic features of cultured grunt fin cells infected with Megalocytivirus. Aquac. Sci. 2009, 57, 9–18. [Google Scholar]
- Dong, C.; Shuang, F.; Weng, S.; He, J. Cloning of a new fibroblast cell line from an early primary culture from mandarin fish (Siniperca chuatsi) fry for efficient proliferation of megalocytiviruses. Cytotechnology 2014, 66, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Imajoh, M.; Ikawa, T.; Oshima, S.-I. Characterization of a new fibroblast cell line from a tail fin of red sea bream, Pagrus major, and phylogenetic relationships of a recent RSIV isolate in Japan. Virus Res. 2007, 126, 45–52. [Google Scholar] [CrossRef]
- Chou, H.-Y.; Hsu, C.-C.; Peng, T.-Y. Isolation and characterization of a pathogenic iridovirus from cultured grouper (Epinephelus sp.) in Taiwan. Fish Pathol. 1998, 33, 201–206. [Google Scholar] [CrossRef]
- Iwamoto, T.; Nakai, T.; Mori, K.-i.; Arimoto, M.; Furusawa, I. Cloning of the fish cell line SSN-1 for piscine nodaviruses. Dis. Aquat. Org. 2000, 43, 81–89. [Google Scholar] [CrossRef]
- Lin, Q.; Fu, X.; Li, N.; Wan, Q.; Chen, W.; Huang, Y.; Huang, Z.; Li, J.; Zhao, L.; Lin, L. Co-infections of infectious spleen and kidney necrosis virus and Siniperca chuatsi rhabdovirus in Chinese perch (Siniperca chuatsi). Microb. Pathog. 2017, 111, 422–430. [Google Scholar] [CrossRef]
- He, J.; Zeng, K.; Weng, S.; Chan, S.-M. Experimental transmission, pathogenicity and physical–chemical properties of infectious spleen and kidney necrosis virus (ISKNV). Aquaculture 2002, 204, 11–24. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Dong, J.; Yan, Y.; Liu, S.; Ye, X.; Gao, F.; Sun, C. Development of a Highly Permissive Mandarin Fish (Siniperca chuatsi) Kidney Cell Line for Mandarin Fish Ranavirus Using a Single-Cell Cloning Method. Cells 2024, 13, 18. https://doi.org/10.3390/cells13010018
Zhang H, Dong J, Yan Y, Liu S, Ye X, Gao F, Sun C. Development of a Highly Permissive Mandarin Fish (Siniperca chuatsi) Kidney Cell Line for Mandarin Fish Ranavirus Using a Single-Cell Cloning Method. Cells. 2024; 13(1):18. https://doi.org/10.3390/cells13010018
Chicago/Turabian StyleZhang, Hetong, Junjian Dong, Yunyun Yan, Shanshan Liu, Xing Ye, Fengying Gao, and Chengfei Sun. 2024. "Development of a Highly Permissive Mandarin Fish (Siniperca chuatsi) Kidney Cell Line for Mandarin Fish Ranavirus Using a Single-Cell Cloning Method" Cells 13, no. 1: 18. https://doi.org/10.3390/cells13010018
APA StyleZhang, H., Dong, J., Yan, Y., Liu, S., Ye, X., Gao, F., & Sun, C. (2024). Development of a Highly Permissive Mandarin Fish (Siniperca chuatsi) Kidney Cell Line for Mandarin Fish Ranavirus Using a Single-Cell Cloning Method. Cells, 13(1), 18. https://doi.org/10.3390/cells13010018