

Long-Term Treatment with Gadopentetic Acid or Gadodiamide Increases TRPC5 Expression and Decreases Adriamycin Nuclear Accumulation in Breast Cancer Cells

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Transfection
2.2. Patch-Clamp Electrophysiology
2.3. Solutions
2.4. Cell Counting Kit-8 Assay
2.5. Immunofluorescence
2.6. Quantitative Real-Time Polymerase Chain Reaction
2.7. The ADM Accumulation Assay
2.8. Calcium Imaging
2.9. Western Blot
2.10. Drugs
2.11. Statistical Methods
3. Results
3.1. Gd-DTPA Potentiated TRPC5 Currents in HEK Cells
3.2. Effect of GBCAs on MCF-7, SK-BR-3, and MCF-7 /ADM Cell Viability in the Presence of ADM
3.3. The Nuclear Accumulation of ADM Was Decreased in the MCF-7 Cells Treated with GCBAs for 4 Weeks
3.4. The Expression of TRPC5 Was Increased after Treatment with GCBAs, and the Inhibition of TRPC5 Increased ADM Nuclear Accumulation
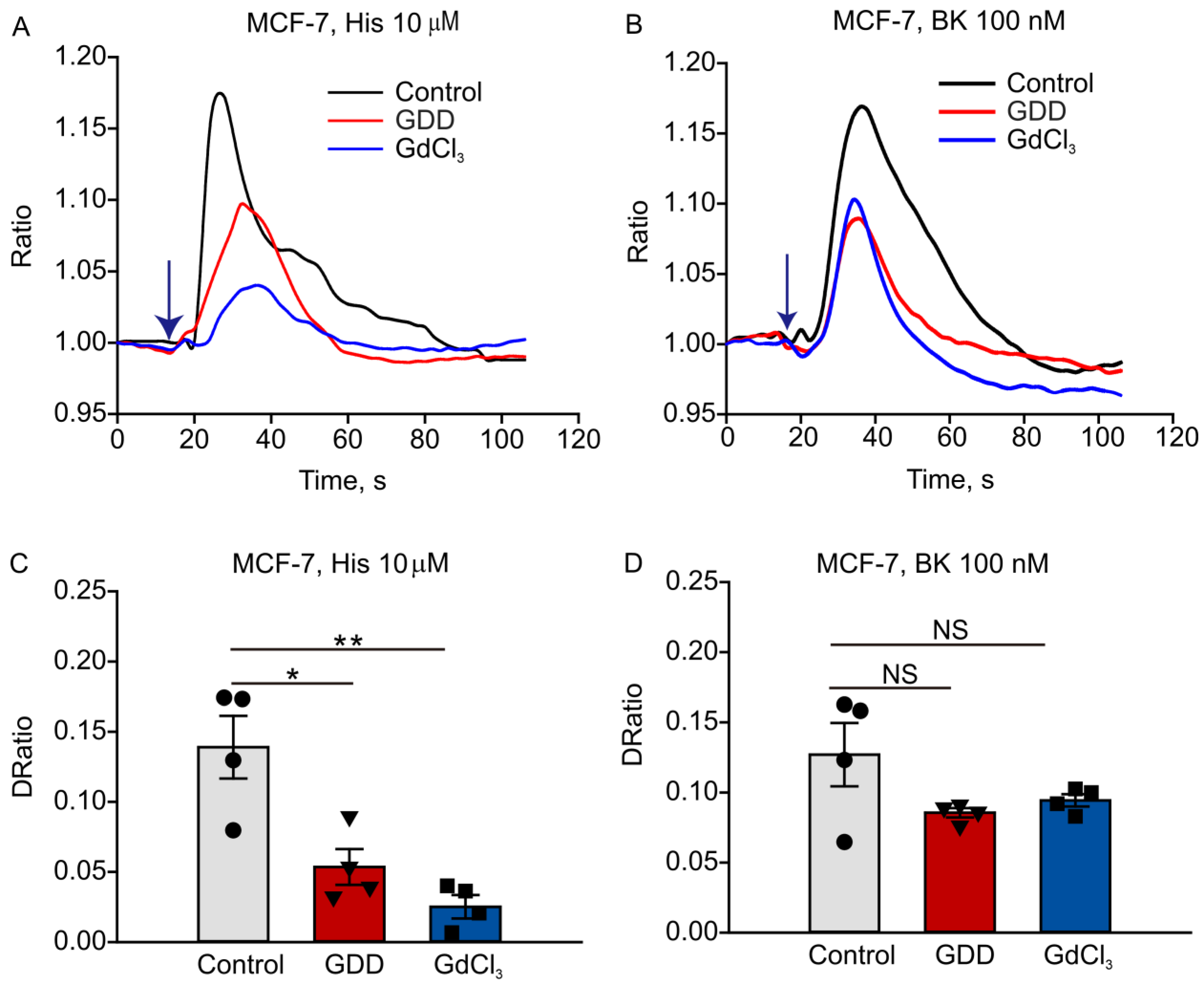
3.5. Long-Term Treatment with GDD Decreased Intracellular Calcium Transients Induced by Histamine and Bradykinin in MCF-7 Cells
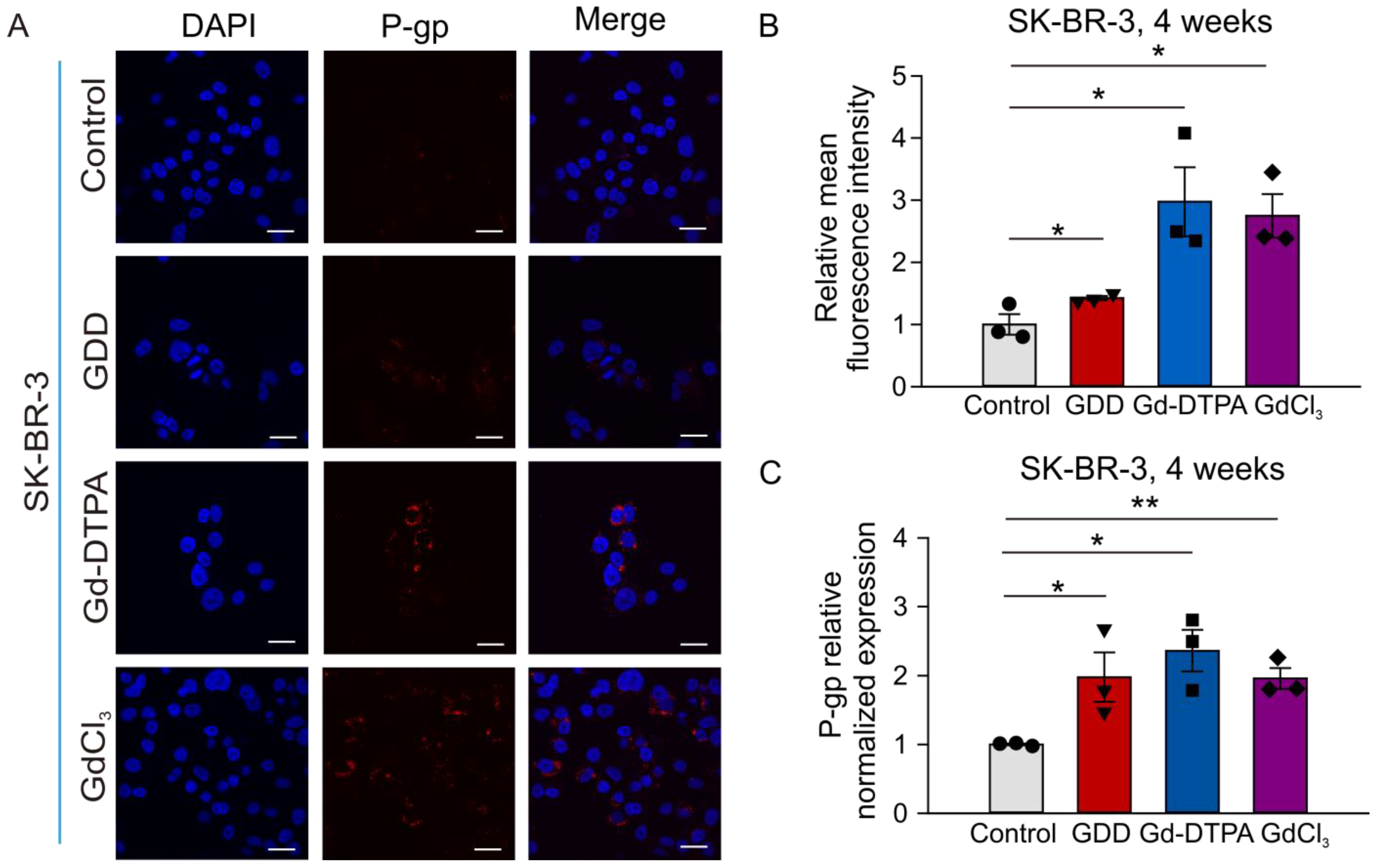
3.6. The Effects of GCBA Treatment on SK-BR-3 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, H.; Zeng, Y.; Zhang, H.; Gu, Z.; Gong, Q.; Luo, K. Functional gadolinium-based nanoscale systems for cancer theranostics. J. Control. Release 2021, 329, 482–512. [Google Scholar] [CrossRef] [PubMed]
- Cheong, B.Y.C.; Wilson, J.M.; Preventza, O.A.; Muthupillai, R. Gadolinium-Based Contrast Agents: Updates and Answers to Typical Questions Regarding Gadolinium Use. Tex. Heart Inst. J. 2022, 49, e217680. [Google Scholar] [CrossRef] [PubMed]
- Rah, Y.C.; Han, E.J.; Park, S.; Rhee, J.; Koun, S.; Park, H.C.; Choi, J. In vivo assay of the potential gadolinium-induced toxicity for sensory hair cells using a zebrafish animal model. J. Appl. Toxicol. JAT 2018, 38, 1398–1404. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Wang, G.; Cao, Q.; Ren, W.; Xu, L.; Bu, S. A risk prediction model for contrast-induced nephropathy associated with gadolinium-based contrast agents. Ren. Fail. 2022, 44, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Turyanskaya, A.; Rauwolf, M.; Pichler, V.; Simon, R.; Burghammer, M.; Fox, O.J.L.; Sawhney, K.; Hofstaetter, J.G.; Roschger, A.; Roschger, P.; et al. Detection and imaging of gadolinium accumulation in human bone tissue by micro- and submicro-XRF. Sci. Rep. 2020, 10, 6301. [Google Scholar] [CrossRef]
- Ramalho, J.; Semelka, R.C.; Ramalho, M.; Nunes, R.H.; AlObaidy, M.; Castillo, M. Gadolinium-Based Contrast Agent Accumulation and Toxicity: An Update. AJNR. Am. J. Neuroradiol. 2016, 37, 1192–1198. [Google Scholar] [CrossRef]
- Ariyani, W.; Miyazaki, W.; Tsushima, Y.; Koibuchi, N. Gadolinium-based contrast agent accelerates the migration of astrocyte via integrin αvβ3 signaling pathway. Sci. Rep. 2022, 12, 5850. [Google Scholar] [CrossRef]
- Funke, S.K.I.; Factor, C.; Rasschaert, M.; Lezius, L.; Sperling, M.; Karst, U.; Robert, P. Long-term Gadolinium Retention in the Healthy Rat Brain: Comparison between Gadopiclenol, Gadobutrol, and Gadodiamide. Radiology 2022, 305, 179–189. [Google Scholar] [CrossRef]
- Nakamura, R.; Takanezawa, Y.; Ohshiro, Y.; Uraguchi, S.; Kiyono, M. Effects of chemical forms of gadolinium on the spleen in mice after single intravenous administration. Biochem. Biophys. Rep. 2022, 29, 101217. [Google Scholar] [CrossRef]
- Richter, H.; Bucker, P.; Martin, L.F.; Dunker, C.; Fingerhut, S.; Xia, A.; Karol, A.; Sperling, M.; Karst, U.; Radbruch, A.; et al. Gadolinium Tissue Distribution in a Large-Animal Model after a Single Dose of Gadolinium-based Contrast Agents. Radiology 2021, 301, 637–642. [Google Scholar] [CrossRef]
- Pasquini, L.; Napolitano, A.; Visconti, E.; Longo, D.; Romano, A.; Toma, P.; Espagnet, M.C.R. Gadolinium-Based Contrast Agent-Related Toxicities. CNS Drugs 2018, 32, 229–240. [Google Scholar] [CrossRef]
- Lee, E.J.; Kim, K.K.; Lee, E.K.; Lee, J.E. Characteristic MRI findings in hyperglycaemia-induced seizures: Diagnostic value of contrast-enhanced fluid-attenuated inversion recovery imaging. Clin. Radiol. 2016, 71, 1240–1247. [Google Scholar] [CrossRef]
- Muldoon, L.L.; Neuwelt, E.A. Dose-Dependent Neurotoxicity (Seizures) Due to Deposition of Gadolinium-based Contrast Agents in the Central Nervous System. Radiology 2015, 277, 925–926. [Google Scholar] [CrossRef]
- Chen, X.; Li, W.; Riley, A.M.; Soliman, M.; Chakraborty, S.; Stamatkin, C.W.; Obukhov, A.G. Molecular Determinants of the Sensitivity to Gq/11-Phospholipase C-dependent Gating, Gd3+ Potentiation, and Ca2+ Permeability in the Transient Receptor Potential Canonical Type 5 (TRPC5) Channel. J. Biol. Chem. 2017, 292, 898–911. [Google Scholar] [CrossRef]
- Semtner, M.; Schaefer, M.; Pinkenburg, O.; Plant, T.D. Potentiation of TRPC5 by protons. J. Biol. Chem. 2007, 282, 33868–33878. [Google Scholar] [CrossRef]
- Strübing, C.; Krapivinsky, G.; Krapivinsky, L.; Clapham, D.E. TRPC1 and TRPC5 form a novel cation channel in mammalian brain. Neuron 2001, 29, 645–655. [Google Scholar] [CrossRef]
- Davare, M.A.; Fortin, D.A.; Saneyoshi, T.; Nygaard, S.; Kaech, S.; Banker, G.; Soderling, T.R.; Wayman, G.A. Transient receptor potential canonical 5 channels activate Ca2+/calmodulin kinase Igamma to promote axon formation in hippocampal neurons. J. Neurosci. 2009, 29, 9794–9808. [Google Scholar] [CrossRef]
- Kumar, S.; Chakraborty, S.; Barbosa, C.; Brustovetsky, T.; Brustovetsky, N.; Obukhov, A.G. Mechanisms controlling neurite outgrowth in a pheochromocytoma cell line: The role of TRPC channels. J. Cell. Physiol. 2012, 227, 1408–1419. [Google Scholar] [CrossRef]
- Zheng, F.; Phelan, K.D. The role of canonical transient receptor potential channels in seizure and excitotoxicity. Cells 2014, 3, 288–303. [Google Scholar] [CrossRef]
- Cai, D.; Li, N.; Jin, L.; Qi, X.; Hua, D.; Wang, T. High CTC-TRPC5 Expression Significantly Associated With Poor Prognosis in Radical Resected Colorectal Cancer Patients. Front. Mol. Biosci. 2021, 8, 727864. [Google Scholar] [CrossRef]
- He, D.X.; Ma, X. Transient receptor potential channel C5 in cancer chemoresistance. Acta Pharmacol. Sin. 2016, 37, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Chen, Z.; Hua, D.; He, D.; Wang, L.; Zhang, P.; Wang, J.; Cai, Y.; Gao, C.; Zhang, X.; et al. Essential role for TrpC5-containing extracellular vesicles in breast cancer with chemotherapeutic resistance. Proc. Natl. Acad. Sci. USA 2014, 111, 6389–6394. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Chen, M.; Zhang, S.; Miao, Z.; Wang, J.; Lu, X.; Zhao, X. TRPC5-induced autophagy promotes the TMZ-resistance of glioma cells via the CAMMKβ/AMPKα/mTOR pathway. Oncol. Rep. 2019, 41, 3413–3423. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Ning, K.; Sun, X.; Zhang, C.; Jin, L.F.; Hua, D. Glycolysis is essential for chemoresistance induced by transient receptor potential channel C5 in colorectal cancer. BMC Cancer 2018, 18, 207. [Google Scholar] [CrossRef]
- Jung, S.; Mühle, A.; Schaefer, M.; Strotmann, R.; Schultz, G.; Plant, T.D. Lanthanides potentiate TRPC5 currents by an action at extracellular sites close to the pore mouth. J. Biol. Chem. 2003, 278, 3562–3571. [Google Scholar] [CrossRef]
- Ma, X.; Cai, Y.; He, D.; Zou, C.; Zhang, P.; Lo, C.Y.; Xu, Z.; Chan, F.L.; Yu, S.; Chen, Y.; et al. Transient receptor potential channel TRPC5 is essential for P-glycoprotein induction in drug-resistant cancer cells. Proc. Natl. Acad. Sci. USA 2012, 109, 16282–16287. [Google Scholar] [CrossRef]
- Thorn, C.F.; Oshiro, C.; Marsh, S.; Hernandez-Boussard, T.; McLeod, H.; Klein, T.E.; Altman, R.B. Doxorubicin pathways: Pharmacodynamics and adverse effects. Pharm. Genom. 2011, 21, 440–446. [Google Scholar] [CrossRef]
- Chen, X.; Sooch, G.; Demaree, I.S.; White, F.A.; Obukhov, A.G. Transient Receptor Potential Canonical (TRPC) Channels: Then and Now. Cells 2020, 9, 1983. [Google Scholar] [CrossRef]
- Santoni, G.; Morelli, M.B.; Marinelli, O.; Nabissi, M.; Santoni, M.; Amantini, C. Calcium Signaling and the Regulation of Chemosensitivity in Cancer Cells: Role of the Transient Receptor Potential Channels. Adv. Exp. Med. Biol. 2020, 1131, 505–517. [Google Scholar] [CrossRef]
- Singh, R.; Adhya, P.; Sharma, S.S. Redox-sensitive TRP channels: A promising pharmacological target in chemotherapy-induced peripheral neuropathy. Expert Opin. Ther. Targets 2021, 25, 529–545. [Google Scholar] [CrossRef]
- Zou, Y.; Liu, Z.; Zhou, Y.; Wang, J.; Xu, Q.; Zhao, X.; Miao, Z. TRPC5 mediates TMZ resistance in TMZ-resistant glioblastoma cells via NFATc3-P-gp pathway. Transl. Oncol. 2021, 14, 101214. [Google Scholar] [CrossRef]
- Perrotta, G.; Metens, T.; Absil, J.; Lemort, M.; Manto, M. Absence of clinical cerebellar syndrome after serial injections of more than 20 doses of gadoterate, a macrocyclic GBCA: A monocenter retrospective study. J. Neurol. 2017, 264, 2277–2283. [Google Scholar] [CrossRef]
- Hylton, N.M.; Blume, J.D.; Bernreuter, W.K.; Pisano, E.D.; Rosen, M.A.; Morris, E.A.; Weatherall, P.T.; Lehman, C.D.; Newstead, G.M.; Polin, S.; et al. Locally advanced breast cancer: MR imaging for prediction of response to neoadjuvant chemotherapy--results from ACRIN 6657/I-SPY TRIAL. Radiology 2012, 263, 663–672. [Google Scholar] [CrossRef]
- Ringler, M.D.; Rhodes, N.G.; Ayers-Ringler, J.R.; Jakaitis, D.R.; McDonald, R.J.; Kallmes, D.F.; McDonald, J.S. Gadolinium retention within multiple rat organs after intra-articular administration of gadolinium-based contrast agents. Skelet. Radiol. 2021, 50, 1419–1425. [Google Scholar] [CrossRef]
- Bezzerides, V.J.; Ramsey, I.S.; Kotecha, S.; Greka, A.; Clapham, D.E. Rapid vesicular translocation and insertion of TRP channels. Nat. Cell Biol. 2004, 6, 709–720. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, X.; Li, H.; Chen, Z.; Yao, X.; Jin, J.; Ma, X. TRPC5-induced autophagy promotes drug resistance in breast carcinoma via CaMKKβ/AMPKα/mTOR pathway. Sci. Rep. 2017, 7, 3158. [Google Scholar] [CrossRef]
- Hittelman, W.N.; Rao, P.N. The nature of adriamycin-induced cytotoxicity in Chinese hamster cells as revealed by premature chromosome condensation. Cancer Res. 1975, 35, 3027–3035. [Google Scholar]
- Machaca, K. Ca(2+) signaling, genes and the cell cycle. Cell Calcium 2010, 48, 243–250. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Wang, M.; Lv, W.; White, F.A.; Chen, X.; Obukhov, A.G. Long-Term Treatment with Gadopentetic Acid or Gadodiamide Increases TRPC5 Expression and Decreases Adriamycin Nuclear Accumulation in Breast Cancer Cells. Cells 2023, 12, 1304. https://doi.org/10.3390/cells12091304
Zhang W, Wang M, Lv W, White FA, Chen X, Obukhov AG. Long-Term Treatment with Gadopentetic Acid or Gadodiamide Increases TRPC5 Expression and Decreases Adriamycin Nuclear Accumulation in Breast Cancer Cells. Cells. 2023; 12(9):1304. https://doi.org/10.3390/cells12091304
Chicago/Turabian StyleZhang, Weiheng, Mengyuan Wang, Weizhen Lv, Fletcher A. White, Xingjuan Chen, and Alexander G. Obukhov. 2023. "Long-Term Treatment with Gadopentetic Acid or Gadodiamide Increases TRPC5 Expression and Decreases Adriamycin Nuclear Accumulation in Breast Cancer Cells" Cells 12, no. 9: 1304. https://doi.org/10.3390/cells12091304
APA StyleZhang, W., Wang, M., Lv, W., White, F. A., Chen, X., & Obukhov, A. G. (2023). Long-Term Treatment with Gadopentetic Acid or Gadodiamide Increases TRPC5 Expression and Decreases Adriamycin Nuclear Accumulation in Breast Cancer Cells. Cells, 12(9), 1304. https://doi.org/10.3390/cells12091304