
Action of 2,6-Dichloro-1,4-benzoquinone on the O2-Evolving Activity of Photosystem II in Chlamydomonas reinhardtii Cells with and without Cell Wall: Inhibitory Effect of Its Oxidized Form


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Confocal Microscopy
2.3. Oxygen Evolution Measurements
2.4. Chlorophyll Concentration
2.5. DCBQ Reduction Measurements
2.6. Statistical Analysis
3. Results
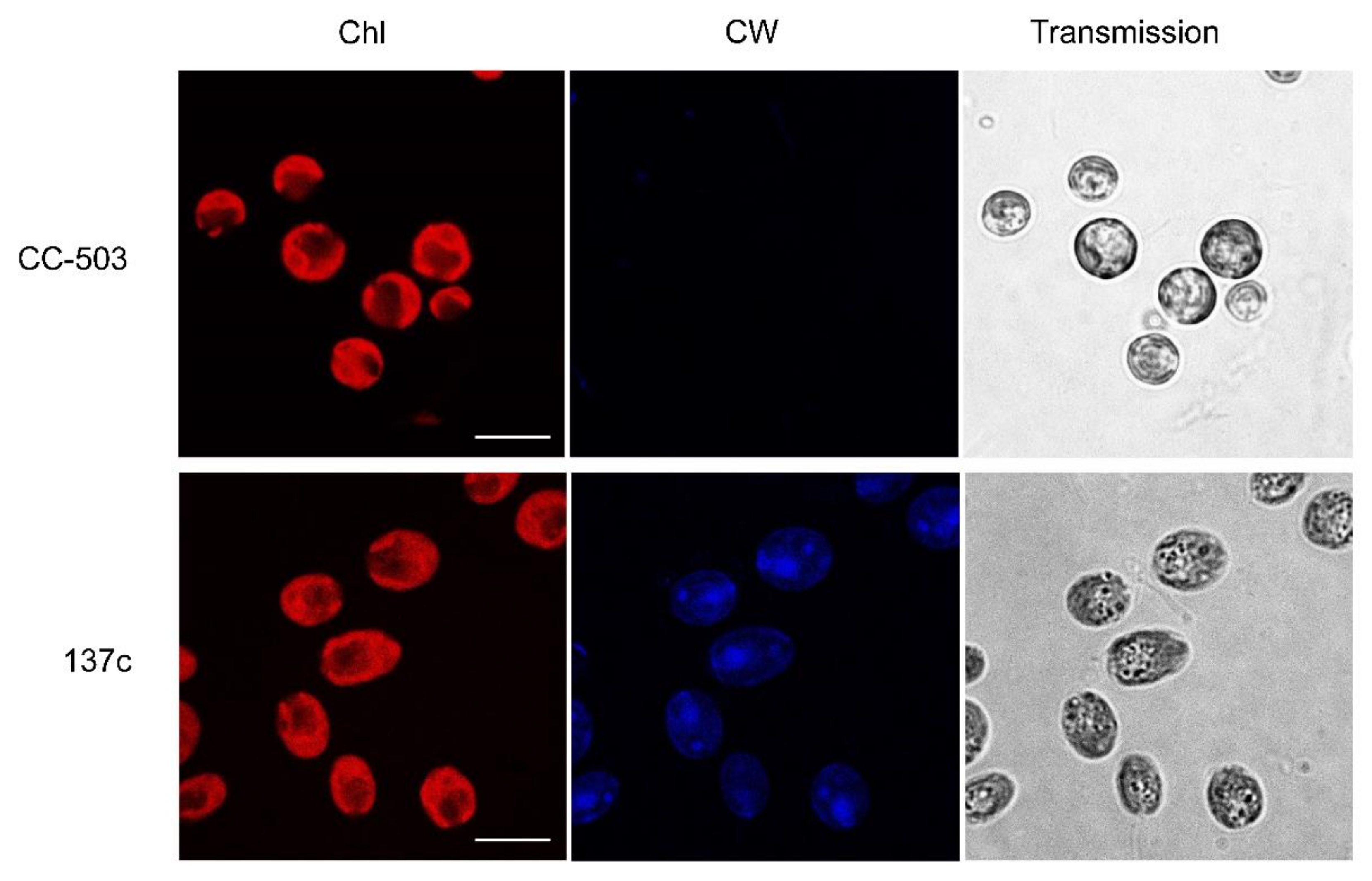
3.1. Presence of Cell Wall
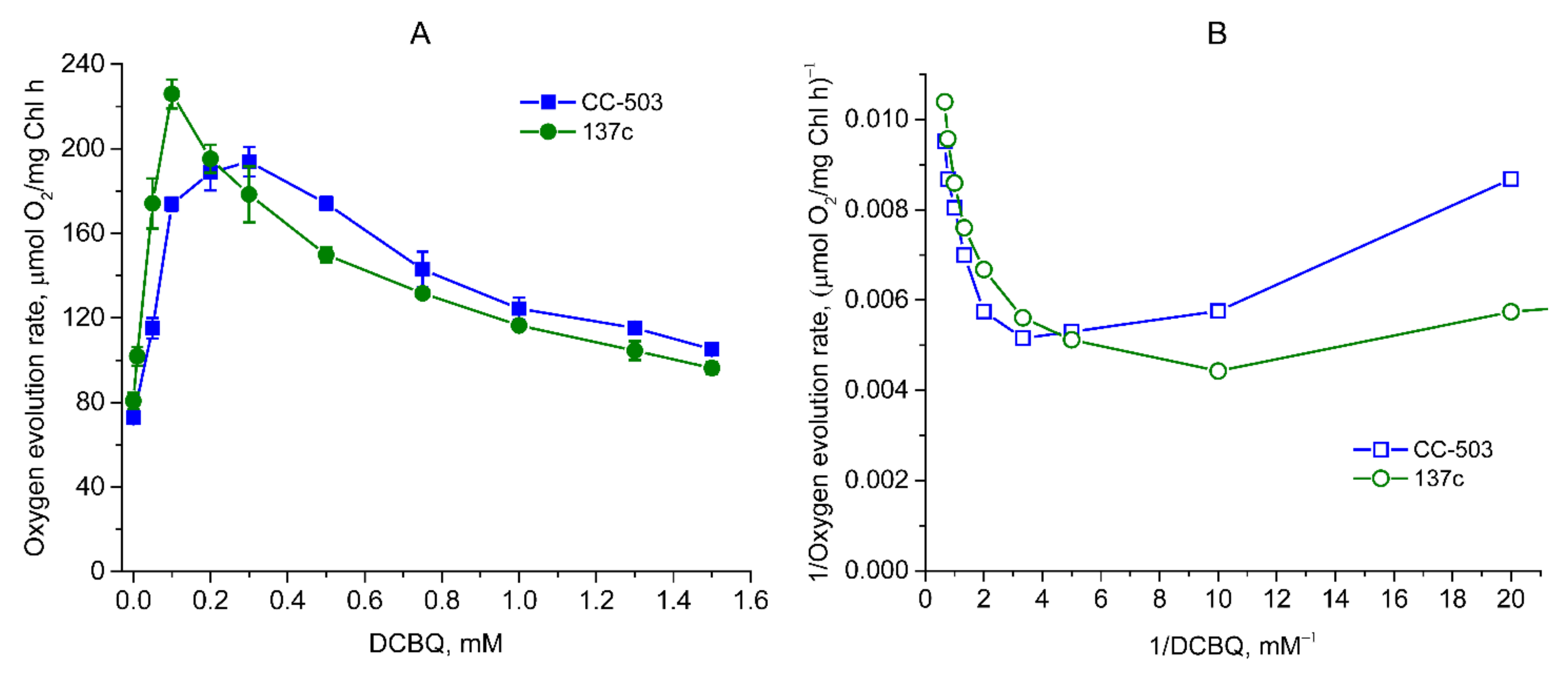
3.2. Influence of DCBQ on the O2 Evolution Rate
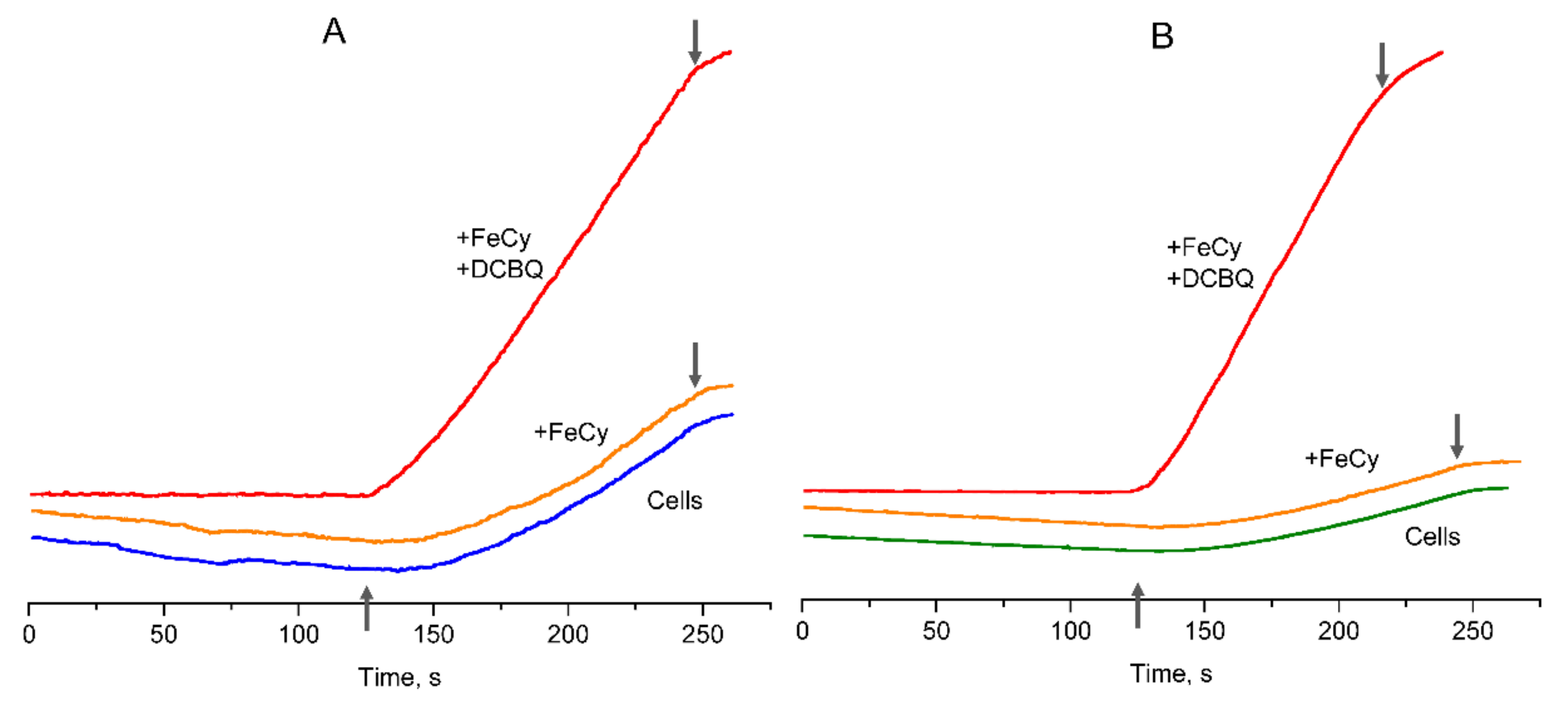
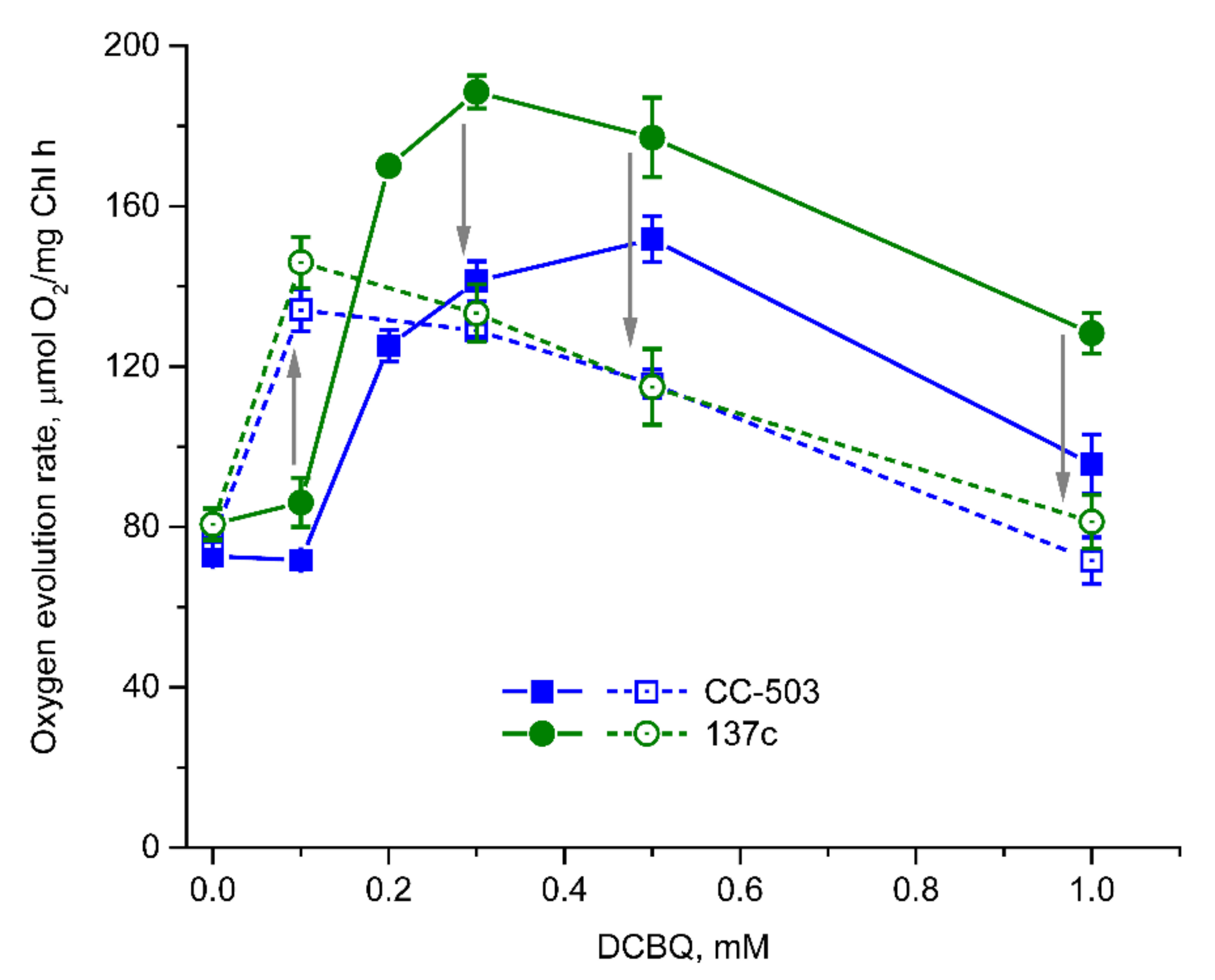
3.3. Involvement of FeCy in the Action of DCBQ
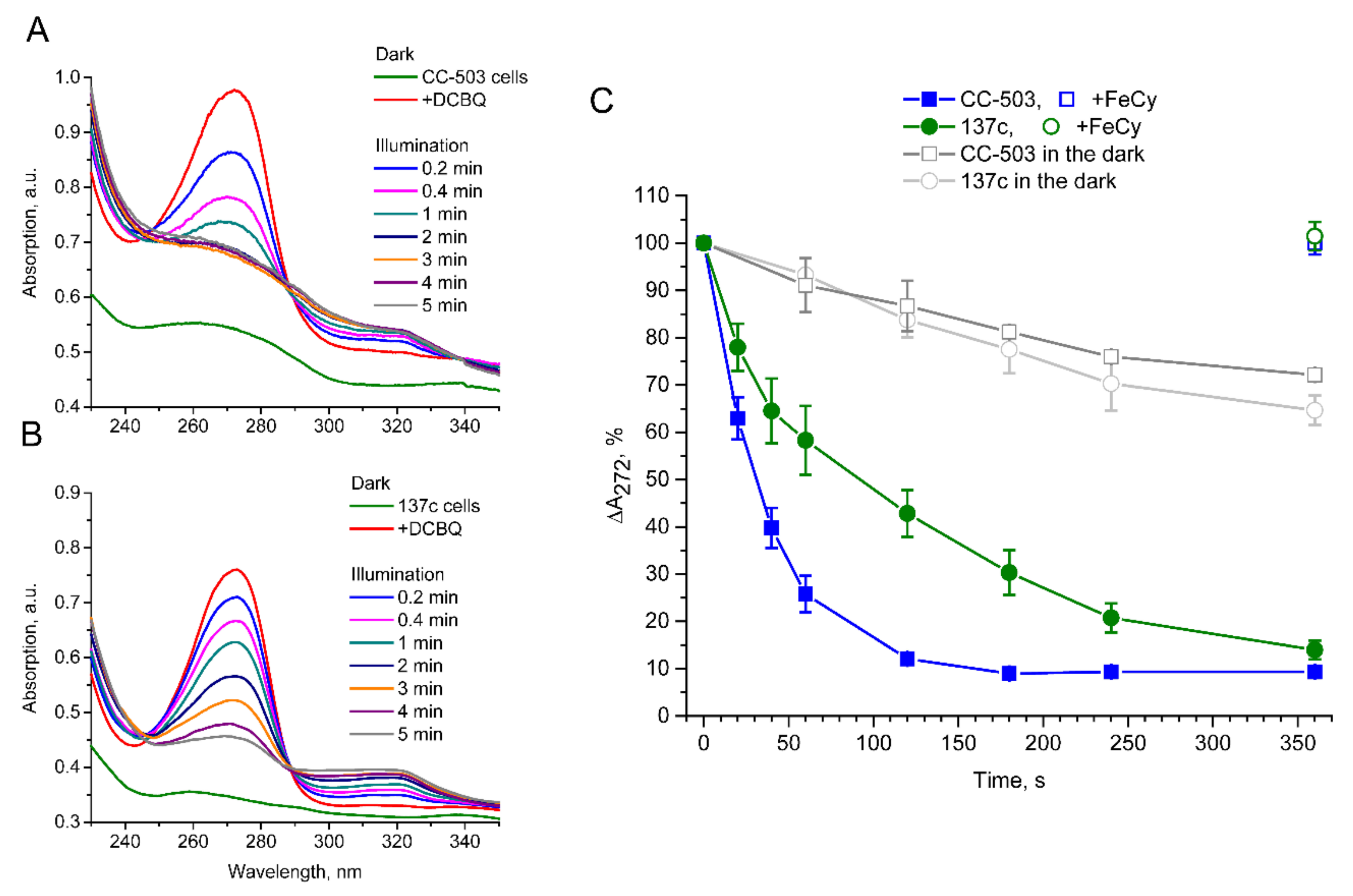
3.4. Light-Induced Reduction in DCBQ by Cells
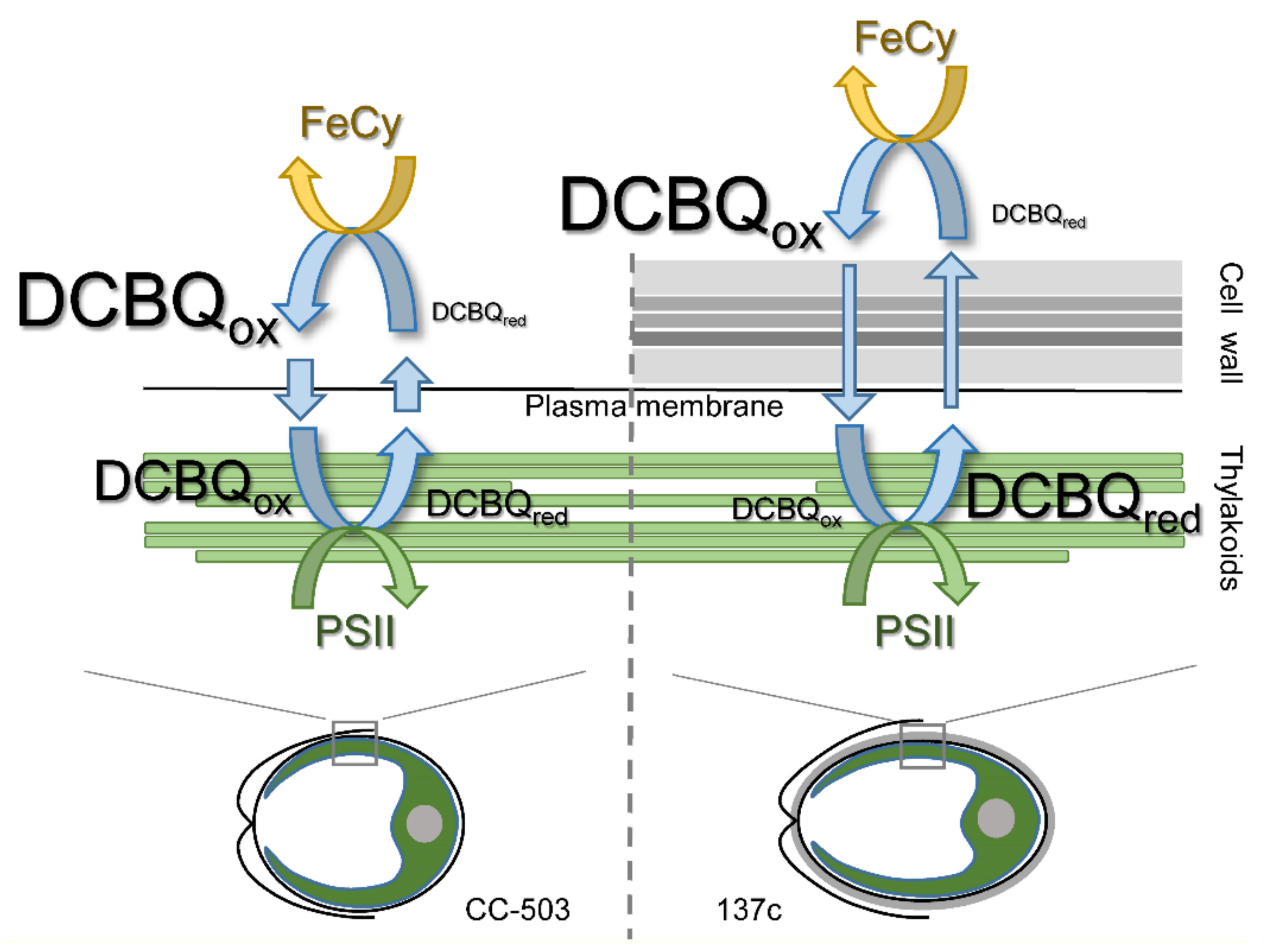
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Müh, F.; Glöckner, C.; Hellmich, J.; Zouni, A. Light-Induced Quinone Reduction in Photosystem II. Biochim. Biophys. Acta-Bioenerg. 2012, 1817, 44–65. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, O.; Khorobrykh, S.; Tyystjärvi, E. Acclimation of Chlamydomonas Reinhardtii to Extremely Strong Light. Photosynth. Res. 2021, 147, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Terentyev, V.V.; Shukshina, A.K.; Shitov, A.V. Carbonic Anhydrase CAH3 Supports the Activity of Photosystem II under Increased PH. Biochim. Biophys. Acta-Bioenerg. 2019, 1860, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Shevela, D.; Messinger, J. Probing the Turnover Efficiency of Photosystem II Membrane Fragments with Different Electron Acceptors. Biochim. Biophys. Acta-Bioenerg. 2012, 1817, 1208–1212. [Google Scholar] [CrossRef]
- Suzuki, T.; Minagawa, J.; Tomo, T.; Sonoike, K.; Ohta, H.; Enami, I. Binding and Functional Properties of the Extrinsic Proteins in Oxygen-Evolving Photosystem II Particle from a Green Alga, Chlamydomonas Reinhardtii Having His-Tagged CP47. Plant Cell Physiol. 2003, 44, 76–84. [Google Scholar] [CrossRef]
- Satoh, K.; Oh-hashi, M.; Kashino, Y.; Koike, H. Mechanism of Electron Flow through the QB Site in Photosystem II. 1. Kinetics of the Reduction of Electron Acceptors at the QB and Plastoquinone Sites in Photosystem II Particles from the Cyanobacterium Synechococcus Vulcanus. Plant Cell Physiol. 1995, 36, 597–605. [Google Scholar] [CrossRef]
- Dudekula, S.; Fragata, M. Investigation of the Electron Transfer Site of P-Benzoquinone in Isolated Photosystem II Particles and Thylakoid Membranes Using α- and β-Cyclodextrins. J. Photochem. Photobiol. B Biol. 2006, 85, 177–183. [Google Scholar] [CrossRef]
- Yanykin, D.V.; Khorobrykh, A.A.; Khorobrykh, S.A.; Klimov, V.V. Photoconsumption of Molecular Oxygen on Both Donor and Acceptor Sides of Photosystem II in Mn-Depleted Subchloroplast Membrane Fragments. Biochim. Biophys. Acta-Bioenerg. 2010, 1797, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Eaton-Rye, J.J. PsbY Is Required for Prevention of Photodamage to Photosystem II in a PsbM-Lacking Mutant of Synechocystis Sp. PCC 6803. Photosynthetica 2018, 56, 200–209. [Google Scholar] [CrossRef]
- Shitov, A.V.; Terentyev, V.V.; Zharmukhamedov, S.K.; Rodionova, M.V.; Karacan, M.; Karacan, N.; Klimov, V.V.; Allakhverdiev, S.I. Is Carbonic Anhydrase Activity of Photosystem II Required for Its Maximum Electron Transport Rate? Biochim. Biophys. Acta-Bioenerg. 2018, 1859, 292–299. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Fedorchuk, T.P.; Terentyev, V.V.; Dymova, O.V.; Naydov, I.A.; Golovko, T.K.; Borisova-Mubarakshina, M.M.; Ivanov, B.N. The Role of Carbonic Anhydrase α-CA4 in the Adaptive Reactions of Photosynthetic Apparatus: The Study with α-CA4 Knockout Plants. Protoplasma 2020, 257, 489–499. [Google Scholar] [CrossRef]
- Roach, T.; Sedoud, A.; Krieger-Liszkay, A. Acetate in Mixotrophic Growth Medium Affects Photosystem II in Chlamydomonas Reinhardtii and Protects against Photoinhibition. Biochim. Biophys. Acta-Bioenerg. 2013, 1827, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M.L.; Franco, E.; Deák, Z.; Schlodder, E.; Vass, I.; Nixon, P.J. Investigating the Photoprotective Role of Cytochrome B-559 in Photosystem II in a Mutant with Altered Ligation of the Haem. Plant Cell Physiol. 2014, 55, 1276–1285. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Sato, F.; Ifuku, K. In Vivo System for Analyzing the Function of the PsbP Protein Using Chlamydomonas Reinhardtii. Photosynth. Res. 2017, 133, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, O.; Valev, D.; Kruse, O.; Wobbe, L.; Tyystjärvi, E. Photoinhibition and Continuous Growth of the Wild-Type and a High-Light Tolerant Strain of Chlamydomonas Reinhardtii. Photosynthetica 2019, 57, 617–626. [Google Scholar] [CrossRef]
- Cecchin, M.; Jeong, J.; Son, W.; Kim, M.; Park, S.; Zuliani, L.; Cazzaniga, S.; Pompa, A.; Young Kang, C.; Bae, S.; et al. LPA2 Protein Is Involved in Photosystem II Assembly in Chlamydomonas Reinhardtii. Plant J. 2021, 107, 1648–1662. [Google Scholar] [CrossRef]
- Longatte, G.; Rappaport, F.; Wollman, F.-A.; Guille-Collignon, M.; Lemaître, F. Electrochemical Harvesting of Photosynthetic Electrons from Unicellular Algae Population at the Preparative Scale by Using 2,6-Dichlorobenzoquinone. Electrochim. Acta 2017, 236, 337–342. [Google Scholar] [CrossRef]
- Sayegh, A.; Longatte, G.; Buriez, O.; Wollman, F.-A.; Guille-Collignon, M.; Labbé, E.; Delacotte, J.; Lemaître, F. Diverting Photosynthetic Electrons from Suspensions of Chlamydomonas Reinhardtii Algae—New Insights Using an Electrochemical Well Device. Electrochim. Acta 2019, 304, 465–473. [Google Scholar] [CrossRef]
- Beauzamy, L.; Beauzamy, L.; Delacotte, J.; Bailleul, B.; Tanaka, K.; Nakanishi, S.; Wollman, F.A.; Lemaître, F. Mediator-Microorganism Interaction in Microbial Solar Cell: A Fluo-Electrochemical Insight. Anal. Chem. 2020, 92, 7532–7539. [Google Scholar] [CrossRef] [PubMed]
- Shlosberg, Y.; Schuster, G.; Adir, N. Harnessing Photosynthesis to Produce Electricity Using Cyanobacteria, Green Algae, Seaweeds and Plants. Front. Plant Sci. 2022, 13, 2603. [Google Scholar] [CrossRef] [PubMed]
- Kirchhoff, H. Diffusion of Molecules and Macromolecules in Thylakoid Membranes. Biochim. Biophys. Acta-Bioenerg. 2014, 1837, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Terentyev, V.V. Loss of Carbonic Anhydrase in the Thylakoid Lumen Causes Unusual Moderate-Light-Induced Rearrangement of the Chloroplast in Chlamydomonas Reinhardtii as a Way of Photosystem II Photoprotection. Plant Physiol. Biochem. 2021, 168, 501–506. [Google Scholar] [CrossRef]
- Polukhina, I.; Fristedt, R.; Dinc, E.; Cardol, P.; Croce, R. Carbon Supply and Photoacclimation Cross Talk in the Green Alga Chlamydomonas Reinhardtii. Plant Physiol. 2016, 172, 1494–1505. [Google Scholar] [CrossRef] [PubMed]
- Jursinic, P.A.; Dennenberg, R.J. Enhanced Oxygen Yields Caused by Double Turnovers of Photosystem II Induced by Dichlorobenzoquinone. Biochim. Biophys. Acta-Bioenerg. 1988, 934, 177–185. [Google Scholar] [CrossRef]
- Halverson, K.M.; Barry, B.A. Sucrose and Glycerol Effects on Photosystem II. Biophys. J. 2003, 85, 1317–1325. [Google Scholar] [CrossRef] [PubMed]
- Iwai, M.; Katoh, H.; Katayama, M.; Ikeuchi, M. PSII-Tc Protein Plays an Important Role in Dimerization of Photosystem II. Plant Cell Physiol. 2004, 45, 1809–1816. [Google Scholar] [CrossRef] [PubMed]
- Kashino, Y.; Yamashita, M.; Okamoto, Y.; Koike, H.; Satoh, K. Mechanisms of Electron Flow through the QB Site in Photosystem II. 3. Effects of the Presence of Membrane Structure on the Redox Reactions at the QB Site. Plant Cell Physiol. 1996, 37, 976–982. [Google Scholar] [CrossRef]
- Goodenough, U.W.; Heuser, J.E. The Chlamydomonas Cell Wall and Its Constituent Glycoproteins Analyzed by the Quick-Freeze, Deep-Etch Technique. J. Cell Biol. 1985, 101, 1550–1568. [Google Scholar] [CrossRef] [PubMed]
- Baudelet, P.-H.; Ricochon, G.; Linder, M.; Muniglia, L. A New Insight into Cell Walls of Chlorophyta. Algal Res. 2017, 25, 333–371. [Google Scholar] [CrossRef]
- Shutova, T.; Kenneweg, H.; Buchta, J.; Nikitina, J.; Terentyev, V.; Chernyshov, S.; Andersson, B.; Allakhverdiev, S.I.; Klimov, V.V.; Dau, H.; et al. The Photosystem II-Associated Cah3 in Chlamydomonas Enhances the O2 Evolution Rate by Proton Removal. EMBO J. 2008, 27, 782–791. [Google Scholar] [CrossRef]
- Terentyev, V.V.; Shukshina, A.K.; Ashikhmin, A.A.; Tikhonov, K.G.; Shitov, A.V. The Main Structural and Functional Characteristics of Photosystem-II-Enriched Membranes Isolated from Wild Type and Cia3 Mutant Chlamydomonas Reinhardtii. Life 2020, 10, 63. [Google Scholar] [CrossRef]
- Shukshina, A.K.; Terentyev, V.V. Involvement of Carbonic Anhydrase CAH3 in the Structural and Functional Stabilization of the Water-Oxidizing Complex of Photosystem II from Chlamydomonas Reinhardtii. Biochemistry 2021, 86, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Work, V.H.; Radakovits, R.; Jinkerson, R.E.; Meuser, J.E.; Elliott, L.G.; Vinyard, D.J.; Laurens, L.M.L.; Dismukes, G.C.; Posewitz, M.C. Increased Lipid Accumulation in the Chlamydomonas Reinhardtii Sta7-10 Starchless Isoamylase Mutant and Increased Carbohydrate Synthesis in Complemented Strains. Eukaryot. Cell 2010, 9, 1251–1261. [Google Scholar] [CrossRef] [PubMed]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of Accurate Extinction Coefficients and Simultaneous Equations for Assaying Chlorophylls a and b Extracted with Four Different Solvents: Verification of the Concentration of Chlorophyll Standards by Atomic Absorption Spectroscopy. Biochim. Biophys. Acta-Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Semin, B.; Davletshina, L.N.; Rubin, A.B. Effect of Sucrose-Bound Polynuclear Iron Oxyhydroxide Nanoparticles on the Efficiency of Electron Transport in the Photosystem II Membranes. Photosynth. Res. 2019, 142, 57–67. [Google Scholar] [CrossRef]
- Cronmiller, E.; Toor, D.; Shao, N.C.; Kariyawasam, T.; Wang, M.H.; Lee, J.-H. Cell Wall Integrity Signaling Regulates Cell Wall-Related Gene Expression in Chlamydomonas Reinhardtii. Sci. Rep. 2019, 9, 12204. [Google Scholar] [CrossRef] [PubMed]
- Shetty, P.; Gitau, M.M.; Maróti, G. Salinity Stress Responses and Adaptation Mechanisms in Eukaryotic Green Microalgae. Cells 2019, 8, 1657. [Google Scholar] [CrossRef]
- Iwai, M.; Roth, M.S.; Niyogi, K.K. Subdiffraction-resolution Live-cell Imaging for Visualizing Thylakoid Membranes. Plant J. 2018, 96, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Samuilov, V.D.; Fedorenko, T.A. Lag Phase of CO2-Dependent O2 Evolution by Illuminated Anabaena Variabilis Cells. Biochemistry 1999, 64, 610–619. [Google Scholar]
- Ilani, A.; Krakover, T. Diffusion- and Reaction Rate-Limited Redox Processes Mediated by Quinones through Bilayer Lipid Membranes. Biophys. J. 1987, 51, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Itoi, H.; Tazawa, S.; Hasegawa, H.; Tanabe, Y.; Iwata, H.; Ohzawa, Y. Study of the Pore Structure and Size Effects on the Electrochemical Capacitor Behaviors of Porous Carbon/Quinone Derivative Hybrids. RSC Adv. 2019, 9, 27602–27614. [Google Scholar] [CrossRef]
- Lennon, A.M.; Prommeenate, P.; Nixon, P.J. Location, Expression and Orientation of the Putative Chlororespiratory Enzymes, Ndh and IMMUTANS, in Higher-Plant Plastids. Planta 2003, 218, 254–260. [Google Scholar] [CrossRef]
- Feilke, K.; Streb, P.; Cornic, G.; Perreau, F.; Kruk, J.; Krieger-Liszkay, A. Effect of Chlamydomonas Plastid Terminal Oxidase 1 Expressed in Tobacco on Photosynthetic Electron Transfer. Plant J. 2016, 85, 219–228. [Google Scholar] [CrossRef]
- Loll, B.; Kern, J.; Saenger, W.; Zouni, A.; Biesiadka, J. Towards Complete Cofactor Arrangement in the 3.0 Å Resolution Structure of Photosystem II. Nature 2005, 438, 1040–1044. [Google Scholar] [CrossRef] [PubMed]
- De Causmaecker, S.; Douglass, J.S.; Fantuzzi, A.; Nitschke, W.; Rutherford, A.W. Energetics of the Exchangeable Quinone, Q B, in Photosystem II. Proc. Natl. Acad. Sci. USA 2019, 116, 19458–19463. [Google Scholar] [CrossRef] [PubMed]
- Huynh, M.T.; Anson, C.W.; Cavell, A.C.; Stahl, S.S.; Hammes-Schiffer, S. Quinone 1 e− and 2 e−/2 H+ Reduction Potentials: Identification and Analysis of Deviations from Systematic Scaling Relationships. J. Am. Chem. Soc. 2016, 138, 15903–15910. [Google Scholar] [CrossRef] [PubMed]
- Guskov, A.; Kern, J.; Gabdulkhakov, A.; Broser, M.; Zouni, A.; Saenger, W. Cyanobacterial Photosystem II at 2.9-Å Resolution and the Role of Quinones, Lipids, Channels and Chloride. Nat. Struct. Mol. Biol. 2009, 16, 334–342. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terentyev, V.V.; Shukshina, A.K.; Chetverkina, A.A. Action of 2,6-Dichloro-1,4-benzoquinone on the O2-Evolving Activity of Photosystem II in Chlamydomonas reinhardtii Cells with and without Cell Wall: Inhibitory Effect of Its Oxidized Form. Cells 2023, 12, 907. https://doi.org/10.3390/cells12060907
Terentyev VV, Shukshina AK, Chetverkina AA. Action of 2,6-Dichloro-1,4-benzoquinone on the O2-Evolving Activity of Photosystem II in Chlamydomonas reinhardtii Cells with and without Cell Wall: Inhibitory Effect of Its Oxidized Form. Cells. 2023; 12(6):907. https://doi.org/10.3390/cells12060907
Chicago/Turabian StyleTerentyev, Vasily V., Anna K. Shukshina, and Angelina A. Chetverkina. 2023. "Action of 2,6-Dichloro-1,4-benzoquinone on the O2-Evolving Activity of Photosystem II in Chlamydomonas reinhardtii Cells with and without Cell Wall: Inhibitory Effect of Its Oxidized Form" Cells 12, no. 6: 907. https://doi.org/10.3390/cells12060907
APA StyleTerentyev, V. V., Shukshina, A. K., & Chetverkina, A. A. (2023). Action of 2,6-Dichloro-1,4-benzoquinone on the O2-Evolving Activity of Photosystem II in Chlamydomonas reinhardtii Cells with and without Cell Wall: Inhibitory Effect of Its Oxidized Form. Cells, 12(6), 907. https://doi.org/10.3390/cells12060907