

Targeting Lung–Gut Axis for Regulating Pollution Particle–Mediated Inflammation and Metabolic Disorders

 ,
,
Abstract
1. The Lung–Gut Axis in Lung and Metabolic Disease
2. CS Exposure Induces Lung Inflammation and Diabetes Mellitus Progression
3. Association of CS Exposure with Gut-Derived Microbiota and Inflammatory Bowel Disease
4. PM Exposure Triggers Lung Inflammation, Gut Microbiota Dysbiosis, and DM Progression
5. CS and PM Exposure Mediates Systemic Inflammation and Metabolic Disorders Associated with Lung–Gut Axis Disruption
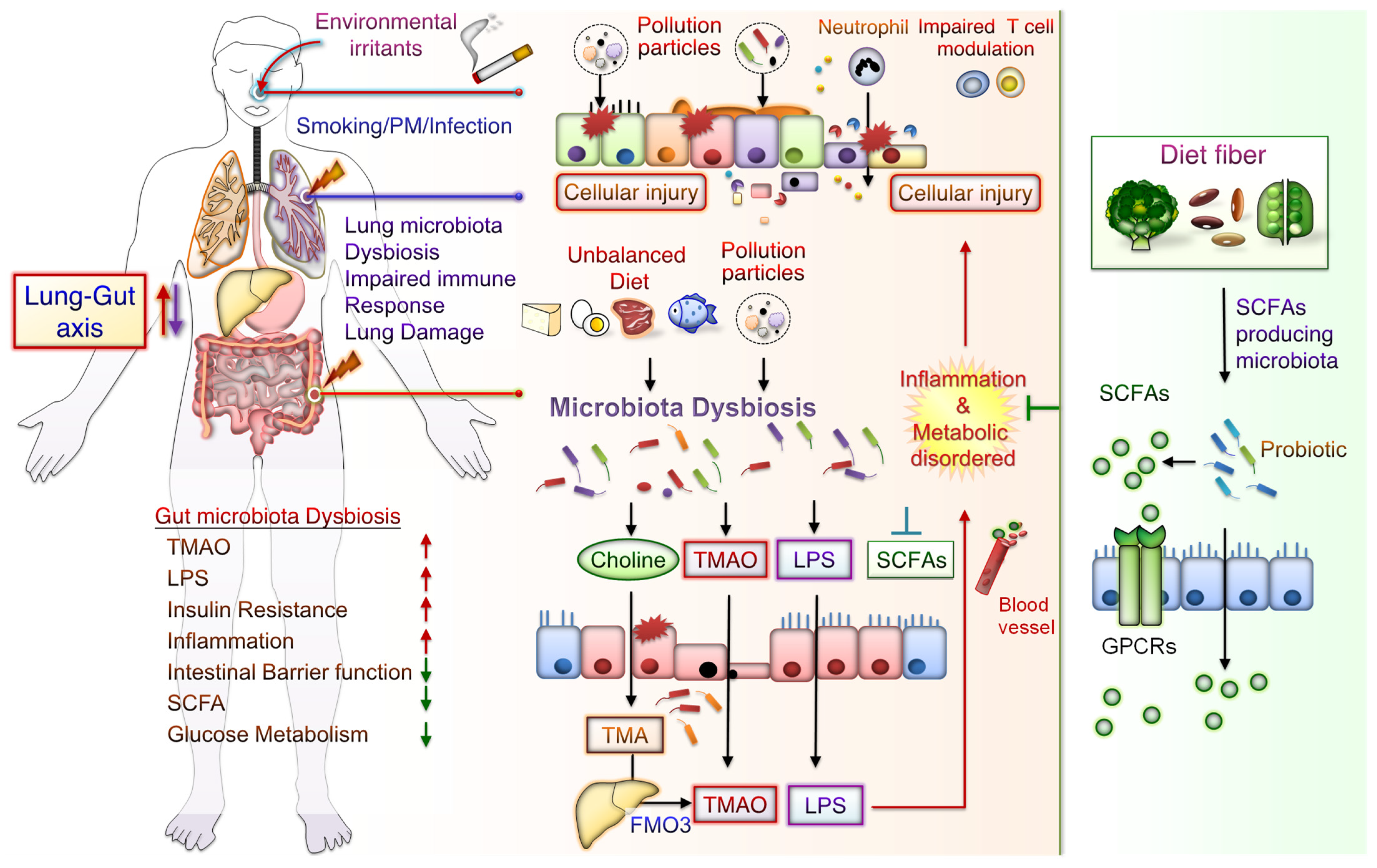
6. Mechanisms Underlying CS and PM Exposure Aggravate Inflammation and Development of Metabolic Disorder through the Lung–Gut Axis
7. Gut Microbiota Dysbiosis in DM
8. Therapeutic Strategies for CS-Mediated Inflammation and Metabolic Disorders
8.1. CS Cessation
8.2. High-Fiber Diet and Probiotics Administration
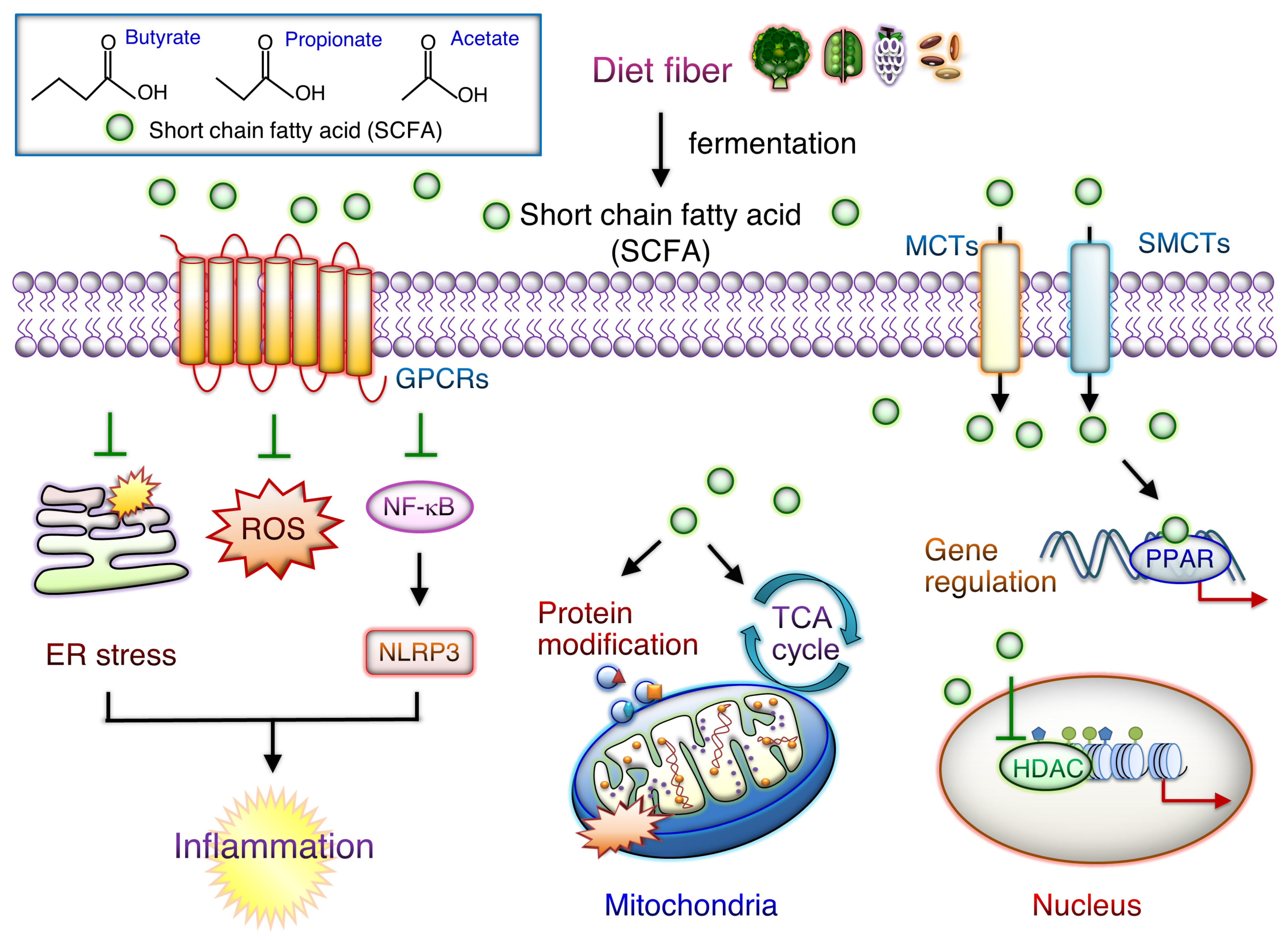
8.3. SCFAs Supplements and Regulation of Systemic Inflammation
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dang, A.T.; Marsland, B.J. Microbes, metabolites, and the gut-lung axis. Mucosal Immunol. 2019, 12, 843–850. Mucosal Immunol. 2019, 12, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Enaud, R.; Prevel, R.; Ciarlo, E.; Beaufils, F.; Wieërs, G.; Guery, B.; Delhaes, L. The Gut-Lung Axis in Health and Respiratory Diseases: A Place for Inter-Organ and Inter-Kingdom Crosstalks. Front. Cell Infect. Microbiol. 2020, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Laiman, V.; Lo, Y.-C.; Chen, H.-C.; Yuan, T.-H.; Hsiao, T.-C.; Chen, J.-K.; Chang, C.-W.; Lin, T.-C.; Li, S.-J.; Chen, Y.-Y.; et al. Effects of antibiotics and metals on lung and intestinal microbiome dysbiosis after sub-chronic lower-level exposure of air pollution in ageing rats. Ecotoxicol. Environ. Saf. 2022, 246, 114164. [Google Scholar] [CrossRef] [PubMed]
- Qing, L.; Xiaoli, T.; Daisuke, M.; Mehrdad, A.; Arun, P. Lung immune tone via gut-lung axis: Gut-derived LPS and short-chain fatty acids’ immunometabolic regulation of lung IL-1beta, FFAR2, and FFAR3 expression. Am. J. Physiol. Lung Cell Mol. Physiol. 2021, 321, L65–L78. [Google Scholar]
- Yongcan, W.; Caixia, P.; Xiaomin, W.; Yilan, W.; Demei, H.; Shihua, S.; Zherui, S.; Shuiqin, L.; Yacong, H.; Zhenxing, W.; et al. Probiotics ameliorates pulmonary inflammation via modulating gut microbiota and rectifying Th17/Treg imbalance in a rat model of PM2.5 induced lung injury. Ecotoxicol. Environ. Saf. 2022, 244, 114060. [Google Scholar]
- Yang, Y.; Zeng, Q.; Gao, J.; Yang, B.; Zhou, J.; Li, K.; Li, L.; Wang, A.; Li, X.; Liu, Z.; et al. High-circulating gut microbiota-dependent metabolite trimethylamine N-oxide is associated with poor prognosis in pulmonary arterial hypertension. Eur. Heart J. Open 2022, 2, oeac021. [Google Scholar] [CrossRef] [PubMed]
- Agirman, G.; Hsiao, E.Y. SnapShot: The microbiota-gut-brain axis. Cell 2021, 184, 2524. [Google Scholar] [CrossRef]
- Sharma, S.; Tripathi, P. Gut microbiome and type 2 diabetes: Where we are and where to go? J. Nutr. Biochem. 2019, 63, 101–108. [Google Scholar] [CrossRef]
- Li, Y.; Xu, L.; Shan, Z.; Teng, W.; Han, C. Association between air pollution and type 2 diabetes: An updated review of the literature. Ther. Adv. Endocrinol. Metab. 2019, 10, 2042018819897046. [Google Scholar] [CrossRef]
- Meo, S.A.; Memon, A.N.; Sheikh, S.A.; Rouq, F.A.; Usmani, A.; Hassan, A.; Arian, S.A. Effect of environmental air pollution on type 2 diabetes mellitus. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 123–128. [Google Scholar]
- Bailey, M.J.; Naik, N.N.; Wild, L.E.; Patterson, W.B.; Alderete, T.L. Exposure to air pollutants and the gut microbiota: A potential link between exposure, obesity, and type 2 diabetes. Gut Microbes 2020, 11, 1188–1202. [Google Scholar] [CrossRef]
- Doiron, D.; De Hoogh, K.; Probst-Hensch, N.; Fortier, I.; Cai, Y.; De Matteis, S.; Hansell, A.L. Air pollution, lung function and COPD: Results from the population-based UK Biobank study. Eur. Respir. J. 2019, 54, 1802140. [Google Scholar] [CrossRef] [PubMed]
- Roemer, E.; Stabbert, R.; Rustemeier, K.; Veltel, D.; Meisgen, T.; Reininghaus, W.; Carchman, R.; Gaworski, C.; Podraza, K. Chemical composition, cytotoxicity and mutagenicity of smoke from US commercial and reference cigarettes smoked under two sets of machine smoking conditions. Toxicology 2003, 195, 31–52. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Shi, G.; Qu, J. Indoor PM2.5, tobacco smoking and chronic lung diseases: A narrative review. Environ. Res. 2019, 181, 108910. [Google Scholar] [CrossRef] [PubMed]
- Lugg, S.T.; Scott, A.; Parekh, D.; Naidu, B.; Thickett, D.R. Cigarette smoke exposure and alveolar macrophages: Mechanisms for lung disease. Thorax 2021, 77, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Laura, A.; Stephanie, S.; Hwang, J.H. Inflammatory Diseases of the Lung Induced by Conventional Cigarette Smoke: A Review. Chest 2015, 148, 1307–1322. [Google Scholar]
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S.; Mehta, J.L. Metabolic syndrome: Pathophysiology, management, and modulation by natural compounds. Ther. Adv. Cardiovasc. Dis. 2017, 11, 215–225. [Google Scholar] [CrossRef]
- Cena, H.; Fonte, M.L.; Turconi, G. Relationship between smoking and metabolic syndrome. Nutr. Rev. 2011, 69, 745–753. [Google Scholar] [CrossRef]
- Baffi, C.W.; Wood, L.; Winnica, D.; Strollo, P.J., Jr.; Gladwin, M.T.; Que, L.G.; Holguin, F. Metabolic Syndrome and the Lung. Chest 2016, 149, 1525–1534. [Google Scholar] [CrossRef]
- Herrero, R.; Sánchez, G.; Asensio, I.; López, E.; Ferruelo, A.; Vaquero, J.; Moreno, L.; de Lorenzo, A.; Bañares, R.; Lorente, J.A. Liver–lung interactions in acute respiratory distress syndrome. Intensiv. Care Med. Exp. 2020, 1, 8–13. [Google Scholar] [CrossRef]
- Mukai, H.; Ming, P.; Lindholm, B.; Heimbürger, O.; Barany, P.; Anderstam, B.; Stenvinkel, P.; Qureshi, A.R. Restrictive lung disorder is common in patients with kidney failure and associates with protein-energy wasting, inflammation and cardiovascular disease. PLoS ONE 2018, 13, e0195585. [Google Scholar] [CrossRef]
- Akter, S.; Goto, A.; Mizoue, T. Smoking and the risk of type 2 diabetes in Japan: A systematic review and meta-analysis. J. Epidemiol. 2017, 27, 553–561. [Google Scholar] [CrossRef]
- Choi, D.-W.; Jeon, J.; Lee, S.A.; Han, K.-T.; Park, E.-C.; Jang, S.-I. Association between Smoking Behavior Patterns and Glycated Hemoglobin Levels in a General Population. Int. J. Environ. Res. Public Health 2018, 15, 2260. [Google Scholar] [CrossRef]
- Campagna, D.; Alamo, A.; Di Pino, A.; Russo, C.; Calogero, A.E.; Purrello, F.; Polosa, R. Smoking and diabetes: Dangerous liaisons and confusing relationships. Diabetol. Metab. Syndr. 2019, 11, 85. [Google Scholar] [CrossRef] [PubMed]
- Eliasson, B.; Attvall, S.; Taskinen, M.R.; Smith, U. The insulin resistance syndrome in smokers is related to smoking habits. Arter. Thromb. A J. Vasc. Biol. 1994, 14, 1946–1950. [Google Scholar] [CrossRef]
- Frati, A.C.; Iniestra, F.; Ariza, C.R. Acute Effect of Cigarette Smoking on Glucose Tolerance and Other Cardiovascular Risk Factors. Diabetes Care 1996, 19, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Eliasson, B.; Attvall, S.; Taskinen, M.-R.; Smith, U. Smoking cessation improves insulin sensitivity in healthy middle-aged men. Eur. J. Clin. Investig. 1997, 27, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Kinney, G.L.; Baker, E.H.; Klein, O.L.; Black-Shinn, J.L.; Wan, E.S.; Make, B.; Regan, E.; Bowler, R.P.; Lutz, S.M.; Young, K.A.; et al. Pulmonary Predictors of Incident Diabetes in Smokers. Chronic Obstr. Pulm. Dis. J. COPD Found. 2016, 3, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Raguraman, R.; Srivastava, A.; Munshi, A.; Ramesh, R. Therapeutic approaches targeting molecular signaling pathways common to diabetes, lung diseases and cancer. Adv. Drug Deliv. Rev. 2021, 178, 113918. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.-W.; Huang, C.-T.; Tsai, Y.-J.; Lien, A.S.-Y.; Lai, F.; Yu, C.-J. Metformin use mitigates the adverse prognostic effect of diabetes mellitus in chronic obstructive pulmonary disease. Respir. Res. 2019, 20, 69. [Google Scholar] [CrossRef]
- Sexton, P.; Metcalf, P.; Kolbe, J. Respiratory Effects of Insulin Sensitisation with Metformin: A Prospective Observational Study. COPD J. Chronic Obstr. Pulm. Dis. 2013, 11, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Khateeb, J.; Fuchs, E.; Khamaisi, M. Diabetes and Lung Disease: A Neglected Relationship. Rev. Diabet. Stud. 2018, 15, 1–15. [Google Scholar] [CrossRef]
- Cade, W.T. Diabetes-related microvascular and macrovascular diseases in the physical therapy setting. Phys. Ther. 2008, 88, 1322–1335. [Google Scholar] [CrossRef] [PubMed]
- Kolahian, S.; Leiss, V.; Nurnberg, B. Diabetic lung disease: Fact or fiction? Rev. Endocr. Metab. Disord. 2019, 20, 303–319. [Google Scholar] [CrossRef]
- Chen, K.-Y.; Wu, S.-M.; Tseng, C.-H.; Lee, K.-Y.; Lin, Y.-H.; Liu, H.-Y.; Chien, L.-N. Combination therapies with thiazolidinediones are associated with a lower risk of acute exacerbations in new-onset COPD patients with advanced diabetic mellitus: A cohort-based case–control study. BMC Pulm. Med. 2021, 21, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, M.; Wang, Z.; Chen, J.; Zhao, J.; Xu, Y.; Wei, X.; Wang, J.; Xie, J. Role of PM2.5 in the development and progression of COPD and its mechanisms. Respir. Res. 2019, 20, 120. [Google Scholar] [CrossRef]
- Håglin, L.M.; Törnkvist, B.; Bäckman, L.O. High serum phosphate and triglyceride levels in smoking women and men with CVD risk and type 2 diabetes. Diabetol. Metab. Syndr. 2014, 6, 39. [Google Scholar] [CrossRef]
- Raftery, A.L.; Tsantikos, E.; Harris, N.L.; Hibbs, M.L. Links Between Inflammatory Bowel Disease and Chronic Obstructive Pulmonary Disease. Front. Immunol. 2020, 11, 2144. [Google Scholar] [CrossRef]
- Long, M.H.; Zhang, C.; Xu, D.Q.; Fu, W.L.; Gan, X.D.; Li, F.; Wang, Q.; Xia, W.; Xu, D.G. PM2.5 aggravates diabetes via the systemically activated IL-6-mediated STAT3/SOCS3 pathway in rats’ liver. Environ. Pollut. 2020, 256, 113342. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Wei, H.; Liu, W.; Coker, O.O.; Gou, H.; Liu, C.; Zhao, L.; Li, C.; Zhou, Y.; Wang, G.; et al. Cigarette smoke promotes colorectal cancer through modulation of gut microbiota and related metabolites. Gut 2022, 71, 2439–2450. [Google Scholar] [CrossRef]
- Chen, X.-Y.; Feng, P.-H.; Han, C.-L.; Jheng, Y.-T.; Wu, C.-D.; Chou, H.-C.; Chen, Y.-Y.; Wu, S.-M.; Lee, K.-Y.; Kuo, H.-P.; et al. Alveolar epithelial inter-alpha-trypsin inhibitor heavy chain 4 deficiency associated with senescence-regulated apoptosis by air pollution. Environ. Pollut. 2021, 278, 116863. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Ho, S.C.; Sun, W.L.; Feng, P.H.; Lin, C.W.; Chen, K.Y.; Chuang, H.C.; Tseng, C.H.; Chen, T.T.; Wu, S.M. Lnc-IL7R alleviates PM2.5-mediated cellular senescence and apoptosis through EZH2 recruitment in chronic obstructive pulmonary disease. Cell Biol. Toxicol. 2022, 38, 1097–1120. [Google Scholar] [CrossRef]
- Wang, J.; Huang, J.; Wang, L.; Chen, C.; Yang, D.; Jin, M.; Bai, C.; Song, Y. Urban particulate matter triggers lung inflammation via the ROS-MAPK-NF-kappaB signaling pathway. J. Thorac. Dis. 2017, 9, 4398–4412. [Google Scholar] [CrossRef]
- Feng, J.; Cavallero, S.; Hsiai, T.; Li, R. Impact of air pollution on intestinal redox lipidome and microbiome. Free Radic. Biol. Med. 2020, 151, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Salim, S.Y.; Kaplan, G.G.; Madsen, K.L. Air pollution effects on the gut microbiota: A link between exposure and inflammatory disease. Gut Microbes. 2014, 5, 215–219. [Google Scholar] [CrossRef]
- Kish, L.; Hotte, N.; Kaplan, G.; Vincent, R.; Tso, R.; Gänzle, M.; Rioux, K.P.; Thiesen, A.; Barkema, H.; Wine, E.; et al. Environmental Particulate Matter Induces Murine Intestinal Inflammatory Responses and Alters the Gut Microbiome. PLoS ONE 2013, 8, e62220. [Google Scholar] [CrossRef] [PubMed]
- Filardo, S.; Di Pietro, M.; Protano, C.; Antonucci, A.; Vitali, M.; Sessa, R. Impact of Air Pollution on the Composition and Diversity of Human Gut Microbiota in General and Vulnerable Populations: A Systematic Review. Toxics 2022, 10, 579. [Google Scholar] [CrossRef]
- Liu, T.; Chen, X.; Xu, Y.; Wu, W.; Tang, W.; Chen, Z.; Ji, G.; Peng, J.; Jiang, Q.; Xiao, J.; et al. Gut microbiota partially mediates the effects of fine particulate matter on type 2 diabetes: Evidence from a population-based epidemiological study. Environ. Int. 2019, 130, 104882. [Google Scholar] [CrossRef]
- Liu, F.; Chen, G.; Huo, W.; Wang, C.; Liu, S.; Li, N.; Mao, S.; Hou, Y.; Lu, Y.; Xiang, H. Associations between long-term exposure to ambient air pollution and risk of type 2 diabetes mellitus: A systematic review and meta-analysis. Environ. Pollut. 2019, 252, 1235–1245. [Google Scholar] [CrossRef]
- Lao, X.Q.; Guo, C.; Chang, L.-Y.; Bo, Y.; Zhang, Z.; Chuang, Y.C.; Jiang, W.K.; Lin, C.; Tam, T.; Lau, A.K.H.; et al. Long-term exposure to ambient fine particulate matter (PM 2.5) and incident type 2 diabetes: A longitudinal cohort study. Diabetologia 2019, 62, 759–769. [Google Scholar] [CrossRef]
- Nassan, F.L.; Kelly, R.S.; Kosheleva, A.; Koutrakis, P.; Vokonas, P.S.; Lasky-Su, J.A.; Schwartz, J.D. Metabolomic signatures of the long-term exposure to air pollution and temperature. Environ. Health 2021, 20, 1–14. [Google Scholar] [CrossRef]
- Wang, N.; Ma, Y.; Liu, Z.; Liu, L.; Yang, K.; Wei, Y.; Liu, Y.; Chen, X.; Sun, X.; Wen, D. Hydroxytyrosol prevents PM 2.5-induced adiposity and insulin resistance by restraining oxidative stress related NF-kappaB pathway and modulation of gut microbiota in a murine model. Free Radic. Biol. Med. 2019, 141, 393–407. [Google Scholar] [CrossRef]
- Xie, S.; Zhang, C.; Zhao, J.; Li, D.; Chen, J. Exposure to concentrated ambient PM 2.5 (CAPM) induces intestinal disturbance via inflammation and alternation of gut microbiome. Environ. Int. 2022, 161, 107138. [Google Scholar] [CrossRef]
- Gui, X.; Yang, Z.; Li, M.D. Effect of Cigarette Smoke on Gut Microbiota: State of Knowledge. Front. Physiol. 2021, 12, 673341. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhou, J.; Chen, M.; Huang, X.; Xie, X.; Li, W.; Cao, Q.; Kan, H.; Xu, Y.; Ying, Z. Exposure to concentrated ambient PM2.5 alters the composition of gut microbiota in a murine model. Part. Fibre Toxicol. 2018, 15, 17. [Google Scholar] [CrossRef] [PubMed]
- Talhout, R.; Schulz, T.; Florek, E.; Van Benthem, J.; Wester, P.; Opperhuizen, A. Hazardous Compounds in Tobacco Smoke. Int. J. Environ. Res. Public Health 2011, 8, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Berkowitz, L.; Pardo-Roa, C.; Salazar, G.; Salazar-Echegarai, F.; Miranda, J.P.; Ramírez, G.; Chávez, J.L.; Kalergis, A.; Bueno, S.M.; Álvarez-Lobos, M. Mucosal Exposure to Cigarette Components Induces Intestinal Inflammation and Alters Antimicrobial Response in Mice. Front. Immunol. 2019, 10, 2289. [Google Scholar] [CrossRef]
- Li, N.; Yang, Z.; Liao, B.; Pan, T.; Pu, J.; Hao, B.; Fu, Z.; Cao, W.; Zhou, Y.; He, F.; et al. Chronic exposure to ambient particulate matter induces gut microbial dysbiosis in a rat COPD model. Respir. Res. 2020, 21, 271. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, S.; Cui, J.; Ding, Z.; Meng, Q.; Sun, H.; Li, B.; Teng, J.; Dong, Y.; Aschner, M.; et al. The influences of ambient fine particulate matter constituents on plasma hormones, circulating TMAO levels and blood pressure: A panel study in China. Environ. Pollut. 2022, 296, 118746. [Google Scholar] [CrossRef]
- Lambertz, J.; Weiskirchen, S.; Landert, S.; Weiskirchen, R. Fructose: A Dietary Sugar in Crosstalk with Microbiota Contributing to the Development and Progression of Non-Alcoholic Liver Disease. Front. Immunol. 2017, 8, 1159. [Google Scholar] [CrossRef]
- Varela-Trinidad, G.U.; Domínguez-Díaz, C.; Solórzano-Castanedo, K.; Íñiguez-Gutiérrez, L.; Hernández-Flores, T.D.J.; Fafutis-Morris, M. Probiotics: Protecting Our Health from the Gut. Microorganisms 2022, 10, 1428. [Google Scholar] [CrossRef]
- Ding, K.; Chen, J.; Zhan, W.; Zhang, S.; Chen, Y.; Long, S.; Lei, M. Microbiome Links Cigarette Smoke-Induced Chronic Obstructive Pulmonary Disease and Dietary Fiber via the Gut-Lung Axis: A Narrative Review. COPD J. Chronic Obstr. Pulm. Dis. 2021, 19, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.G.; Vaughan, A.; Dent, A.G.; O’Hare, P.; Goh, F.; Bowman, R.V.; Fong, K.; Yang, I.A. Biomarkers of progression of chronic obstructive pulmonary disease (COPD). J. Thorac. Dis. 2014, 6, 1532–1547. [Google Scholar]
- Shukla, S.D.; Budden, K.F.; Neal, R.; Hansbro, P.M. Microbiome effects on immunity, health and disease in the lung. Clin. Transl. Immunol. 2017, 6, e133. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Yun, Y.; Kim, S.J.; Lee, E.-J.; Chang, Y.; Ryu, S.; Shin, H.; Kim, H.-L.; Kim, H.-N.; Lee, J.H. Association between Cigarette Smoking Status and Composition of Gut Microbiota: Population-Based Cross-Sectional Study. J. Clin. Med. 2018, 7, 282. [Google Scholar] [CrossRef] [PubMed]
- Biedermann, L.; Zeitz, J.; Mwinyi, J.; Sutter-Minder, E.; Rehman, A.; Ott, S.J.; Steurer-Stey, C.; Frei, A.; Frei, P.; Scharl, M.; et al. Smoking Cessation Induces Profound Changes in the Composition of the Intestinal Microbiota in Humans. PLoS ONE 2013, 8, e59260. [Google Scholar] [CrossRef]
- Antinozzi, M.; Giffi, M.; Sini, N.; Gallè, F.; Valeriani, F.; De Vito, C.; Liguori, G.; Spica, V.R.; Cattaruzza, M.S. Cigarette Smoking and Human Gut Microbiota in Healthy Adults: A Systematic Review. Biomedicines 2022, 10, 510. [Google Scholar] [CrossRef]
- Savin, Z.; Kivity, S.; Yonath, H.; Yehuda, S. Smoking and the intestinal microbiome. Arch. Microbiol. 2018, 200, 677–684. [Google Scholar] [CrossRef]
- Benjamin, J.L.; Hedin, C.R.; Koutsoumpas, A.; Ng, S.C.; McCarthy, N.E.; Prescott, N.J.; Pessoa-Lopes, P.; Mathew, C.G.; Sanderson, J.; Hart, A.L.; et al. Smokers with active Crohn’s disease have a clinically relevant dysbiosis of the gastrointestinal microbiota. Inflamm. Bowel Dis. 2012, 18, 1092–1100. [Google Scholar] [CrossRef]
- Biedermann, L.; Brülisauer, K.; Zeitz, J.; Frei, P.; Scharl, M.; Vavricka, S.R.; Fried, M.; Loessner, M.J.; Rogler, G.; Schuppler, M. Tu1771 Smoking Cessation Alters Intestinal Microbiota: Further Insights From Quantitative Investigations on Human Fecal Samples Using FISH and qPCR. Gastroenterology 2014, 146, 1496–1501. [Google Scholar] [CrossRef]
- Nolan-Kenney, R.; Wu, F.; Hu, J.; Yang, L.; Kelly, D.; Li, H.; Jasmine, F.; Kibriya, M.G.; Parvez, F.; Shaheen, I.; et al. The Association Between Smoking and Gut Microbiome in Bangladesh. Nicotine Tob. Res. 2019, 22, 1339–1346. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Shi, G. Smoking and microbiome in oral, airway, gut and some systemic diseases. J. Transl. Med. 2019, 17, 225. [Google Scholar] [CrossRef]
- Zeller, I.; Malovichko, M.V.; Hurst, H.E.; Renaud, D.E.; Scott, D.A. Cigarette smoke reduces short chain fatty acid production by a Porphyromonas gingivalis clinical isolate. J. Periodontal. Res. 2019, 54, 566–571. [Google Scholar] [CrossRef]
- Jang, Y.O.; Kim, O.-H.; Kim, S.J.; Lee, S.H.; Yun, S.; Lim, S.E.; Yoo, H.J.; Shin, Y.; Lee, S.W. High-fiber diets attenuate emphysema development via modulation of gut microbiota and metabolism. Sci. Rep. 2021, 11, 7008. [Google Scholar] [CrossRef] [PubMed]
- Tomoda, K.; Kubo, K.; Asahara, T.; Andoh, A.; Nomoto, K.; Nishii, Y.; Yamamoto, Y.; Yoshikawa, M.; Kimura, H. Cigarette smoke decreases organic acids levels and population of bifidobacterium in the caecum of rats. J. Toxicol. Sci. 2011, 36, 261–266. [Google Scholar] [CrossRef]
- Meehan, C.; Beiko, R.G. A Phylogenomic View of Ecological Specialization in the Lachnospiraceae, a Family of Digestive Tract-Associated Bacteria. Genome Biol. Evol. 2014, 6, 703–713. [Google Scholar] [CrossRef]
- Fluhr, L.; Mor, U.; Kolodziejczyk, A.A.; Dori-Bachash, M.; Leshem, A.; Itav, S.; Cohen, Y.; Suez, J.; Zmora, N.; Moresi, C.; et al. Gut microbiota modulates weight gain in mice after discontinued smoke exposure. Nature 2021, 600, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Brule, S.V.D.; Rappe, M.; Ambroise, J.; Bouzin, C.; Dessy, C.; Paquot, A.; Muccioli, G.G.; Lison, D. Diesel exhaust particles alter the profile and function of the gut microbiota upon subchronic oral administration in mice. Part. Fibre Toxicol. 2021, 18, 7. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, T.; Si, B.; Du, H.; Liu, Y.; Waqas, A.; Huang, S.; Zhao, G.; Chen, S.; Xu, A. Intratracheally instillated diesel PM2.5 significantly altered the structure and composition of indigenous murine gut microbiota. Ecotoxicol. Environ. Saf. 2021, 210, 111903. [Google Scholar] [CrossRef]
- Zhong, H.; Ren, H.; Lu, Y.; Fang, C.; Hou, G.; Yang, Z.; Chen, B.; Yang, F.; Zhao, Y.; Shi, Z.; et al. Distinct gut metagenomics and metaproteomics signatures in prediabetics and treatment-naïve type 2 diabetics. Ebiomedicine 2019, 47, 373–383. [Google Scholar] [CrossRef]
- Wang, J.; Li, W.; Wang, C.; Wang, L.; He, T.; Hu, H.; Song, J.; Cui, C.; Qiao, J.; Qing, L.; et al. Enterotype Bacteroides Is Associated with a High Risk in Patients with Diabetes: A Pilot Study. J. Diabetes Res. 2020, 2020, 6047145. [Google Scholar] [CrossRef] [PubMed]
- James, M.M.; Pal, N.; Sharma, P.; Kumawat, M.; Shubham, S.; Verma, V.; Tiwari, R.R.; Singh, B.; Nagpal, R.; Sarma, D.K.; et al. Role of butyrogenic Firmicutes in type-2 diabetes. J. Diabetes Metab. Disord. 2022, 21, 1873–1882. [Google Scholar] [CrossRef] [PubMed]
- Bhute, S.S.; Suryavanshi, M.V.; Joshi, S.M.; Yajnik, C.S.; Shouche, Y.S.; Ghaskadbi, S.S. Gut Microbial Diversity Assessment of Indian Type-2-Diabetics Reveals Alterations in Eubacteria, Archaea, and Eukaryotes. Front. Microbiol. 2017, 8, 214. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Stoeva, M.K.; Justice, N.; Nemchek, M.; Sieber, C.M.K.; Tyagi, S.; Gines, J.; Skennerton, C.T.; Souza, M.; Kolterman, O.; et al. Increased circulating butyrate and ursodeoxycholate during probiotic intervention in humans with type 2 diabetes. BMC Microbiol. 2022, 22, 19. [Google Scholar] [CrossRef] [PubMed]
- Walicka, M.; Russo, C.; Baxter, M.; John, I.; Caci, G.; Polosa, R. Impact of stopping smoking on metabolic parameters in diabetes mellitus: A scoping review. World J. Diabetes 2022, 13, 422–433. [Google Scholar] [CrossRef]
- Ohkuma, T.; Iwase, M.; Fujii, H.; Kaizu, S.; Ide, H.; Jodai, T.; Kikuchi, Y.; Idewaki, Y.; Hirakawa, Y.; Nakamura, U.; et al. Dose- and Time-Dependent Association of Smoking and Its Cessation with Glycemic Control and Insulin Resistance in Male Patients with Type 2 Diabetes Mellitus: The Fukuoka Diabetes Registry. PLoS ONE 2015, 10, e0122023. [Google Scholar] [CrossRef]
- Chen, H.-J.; Huang, W.-H.; Chan, H.-L.; Hwang, L.-C. Improvement in Cardiometabolic Risk Factors During Smoking Cessation Treatment in Patients with Type 2 Diabetes: A Retrospective Cohort Study. Diabetes Metab. Syndr. Obes. 2021, 14, 1695–1702. [Google Scholar] [CrossRef]
- Bergamino, M.; Rullan, A.J.; Saigí, M.; Peiró, I.; Montanya, E.; Palmero, R.; Ruffinelli, J.C.; Navarro, A.; Arnaiz, M.D.; Brao, I.; et al. Fasting plasma glucose is an independent predictor of survival in patients with locally advanced non-small cell lung cancer treated with concurrent chemoradiotherapy. BMC Cancer 2019, 19, 165. [Google Scholar] [CrossRef]
- Fu, Z.; Shrubsole, M.J.; Smalley, W.E.; Ness, R.M.; Zheng, W. Associations between Dietary Fiber and Colorectal Polyp Risk Differ by Polyp Type and Smoking Status. J. Nutr. 2014, 144, 592–598. [Google Scholar] [CrossRef]
- Kocsis, T.; Molnár, B.; Németh, D.; Hegyi, P.; Szakács, Z.; Bálint, A.; Garami, A.; Soós, A.; Márta, K.; Solymár, M. Probiotics have beneficial metabolic effects in patients with type 2 diabetes mellitus: A meta-analysis of randomized clinical trials. Sci. Rep. 2020, 10, 11787. [Google Scholar] [CrossRef] [PubMed]
- Kotlyarov, S. Role of Short-Chain Fatty Acids Produced by Gut Microbiota in Innate Lung Immunity and Pathogenesis of the Heterogeneous Course of Chronic Obstructive Pulmonary Disease. Int. J. Mol. Sci. 2022, 23, 4768. [Google Scholar] [CrossRef] [PubMed]
- Simpson, H.L.; Campbell, B.J. Review article: Dietary fibre-microbiota interactions. Aliment. Pharmacol. Ther. 2015, 42, 158–179. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Jang, Y.O.; Lee, S.H.; Choi, J.J.; Kim, D.-H.; Choi, J.-M.; Kang, M.-J.; Oh, Y.-M.; Park, Y.-J.; Shin, Y.; Lee, S.W. Fecal microbial transplantation and a high fiber diet attenuates emphysema development by suppressing inflammation and apoptosis. Exp. Mol. Med. 2020, 52, 1128–1139. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, F.M.; Silva, H.L.; Poso, S.M.; Barroso, M.V.; Lanzetti, M.; Rocha, R.S.; Graça, J.S.; Esmerino, E.A.; Freitas, M.Q.; Silva, M.C.; et al. Probiotic Prato cheese attenuates cigarette smoke-induced injuries in mice. Food Res. Int. 2019, 123, 697–703. [Google Scholar] [CrossRef]
- Carvalho, J.L.; Miranda, M.; Fialho, A.K.; Castro-Faria-Neto, H.; Anatriello, E.; Keller, A.C.; Aimbire, F. Oral feeding with probiotic Lactobacillus rhamnosus attenuates cigarette smoke-induced COPD in C57Bl/6 mice: Relevance to inflammatory markers in human bronchial epithelial cells. PLoS ONE 2020, 15, e0225560. [Google Scholar] [CrossRef]
- Levi, Y.L.D.A.S.; Picchi, R.N.; Silva, E.K.T.; Neto, H.B.; Prado, R.L.D.; Neves, A.D.P.; Messora, M.R.; Maia, L.P. Probiotic Administration Increases Mandibular Bone Mineral Density on Rats Exposed to Cigarette Smoke Inhalation. Braz. Dent. J. 2019, 30, 634–640. [Google Scholar] [CrossRef]
- Naruszewicz, M.; Johansson, M.-L.; Zapolska-Downar, D.; Bukowska, H. Effect of Lactobacillus plantarum 299v on cardiovascular disease risk factors in smokers. Am. J. Clin. Nutr. 2002, 76, 1249–1255. [Google Scholar] [CrossRef]
- Caballero-Franco, C.; Keller, K.; De Simone, C.; Chadee, K. The VSL#3 probiotic formula induces mucin gene expression and secretion in colonic epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G315–G322. [Google Scholar]
- Wang, H.; Chen, X.; Gao, Q.; Liu, K.; Bi, G.; Deng, J.; Zhang, X. Smoking induces the occurrence of colorectal cancer via changing the intestinal permeability. J. BUON J. Balk. Union Oncol. 2021, 26, 1009–1015. [Google Scholar]
- Wang, X.; Hui, Y.; Zhao, L.; Hao, Y.; Guo, H.; Ren, F. Oral administration of Lactobacillus paracasei L9 attenuates PM2.5-induced enhancement of airway hyperresponsiveness and allergic airway response in murine model of asthma. PLoS ONE 2017, 12, e0171721. [Google Scholar] [CrossRef] [PubMed]
- Panebianco, C.; Eddine, F.B.N.; Forlani, G.; Palmieri, G.; Tatangelo, L.; Villani, A.; Xu, L.; Accolla, R.; Pazienza, V. Probiotic Bifidobacterium lactis, anti-oxidant vitamin E/C and anti-inflammatory dha attenuate lung inflammation due to pm2.5 exposure in mice. Benef. Microbes 2019, 10, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Reale, M.; Boscolo, P.; Bellante, V.; Tarantelli, C.; Di Nicola, M.; Forcella, L.; Li, Q.; Morimoto, K.; Muraro, R. Daily intake of Lactobacillus casei Shirota increases natural killer cell activity in smokers. Br. J. Nutr. 2011, 108, 308–314. [Google Scholar] [CrossRef]
- Phillippi, D.T.; Daniel, S.; Nguyen, K.N.; Penaredondo, B.A.; Lund, A.K. Probiotics Function as Immunomodulators in the Intestine in C57Bl/6 Male Mice Exposed to Inhaled Diesel Exhaust Particles on a High-Fat Diet. Cells 2022, 11, 1445. [Google Scholar] [CrossRef]
- Zeng, Z.; Yuan, Q.; Yu, R.; Zhang, J.; Ma, H.; Chen, S. Ameliorative Effects of Probiotic Lactobacillus paracasei NL41 on Insulin Sensitivity, Oxidative Stress, and Beta-Cell Function in a Type 2 Diabetes Mellitus Rat Model. Mol. Nutr. Food Res. 2019, 63, e1900457. [Google Scholar] [CrossRef]
- Svingen, G.F.; Zuo, H.; Ueland, P.M.; Seifert, R.; Løland, K.H.; Pedersen, E.R.; Schuster, P.M.; Karlsson, T.; Tell, G.S.; Schartum-Hansen, H.; et al. Increased plasma trimethylamine- N -oxide is associated with incident atrial fibrillation. Int. J. Cardiol. 2018, 267, 100–106. [Google Scholar] [CrossRef]
- Strassheim, D.; Sullivan, T.; Irwin, D.C.; Gerasimovskaya, E.; Lahm, T.; Klemm, D.J.; Dempsey, E.C.; Stenmark, K.R.; Karoor, V. Metabolite G-Protein Coupled Receptors in Cardio-Metabolic Diseases. Cells 2021, 10, 3347. [Google Scholar] [CrossRef]
- Li, M.; van Esch, B.C.A.M.; Henricks, P.A.J.; Folkerts, G.; Garssen, J. The Anti-inflammatory Effects of Short Chain Fatty Acids on Lipopolysaccharide- or Tumor Necrosis Factor alpha-Stimulated Endothelial Cells via Activation of GPR41/43 and Inhibition of HDACs. Front. Pharmacol. 2018, 9, 533. [Google Scholar] [CrossRef]
- Mao, J.; Li, Y.; Bian, Q.; Xuan, Y.; Li, J.; Wang, Z.; Feng, S.; Liu, X.; Tian, Y.; Li, S. The Bufei Jianpi Formula Improves Mucosal Immune Function by Remodeling Gut Microbiota Through the SCFAs/GPR43/NLRP3 Pathway in Chronic Obstructive Pulmonary Disease Rats. Int. J. Chron. Obstruct. Pulmon. Dis. 2022, 17, 1285–1298. [Google Scholar] [CrossRef] [PubMed]
- Vieira, R.D.S.; Castoldi, A.; Basso, P.J.; Hiyane, M.I.; Câmara, N.O.S.; Almeida, R.R. Butyrate Attenuates Lung Inflammation by Negatively Modulating Th9 Cells. Front. Immunol. 2019, 10, 67. [Google Scholar] [CrossRef] [PubMed]
- Ying, X.D.; Wei, G.; An, H. Sodium butyrate relieves lung ischemia-reperfusion injury by inhibiting NF-kappaB and JAK2/STAT3 signaling pathways. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 413–422. [Google Scholar]
- Prihandoko, R.; Kaur, D.; Wiegman, C.H.; Alvarez-Curto, E.; Donovan, C.; Chachi, L.; Ulven, T.; Tyas, M.R.; Euston, E.; Dong, Z.; et al. Pathophysiological regulation of lung function by the free fatty acid receptor FFA4. Sci. Transl. Med. 2020, 12, 557. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef]
- Müller, M.; Hernández, M.A.G.; Goossens, G.H.; Reijnders, D.; Holst, J.J.; Jocken, J.W.E.; Van Eijk, H.; Canfora, E.E.; Blaak, E.E. Circulating but not faecal short-chain fatty acids are related to insulin sensitivity, lipolysis and GLP-1 concentrations in humans. Sci. Rep. 2019, 9, 12515. [Google Scholar] [CrossRef]
- Huang, W.; Man, Y.; Gao, C.; Zhou, L.; Gu, J.; Xu, H.; Wan, Q.; Long, Y.; Chai, L.; Xu, Y.; et al. Short-Chain Fatty Acids Ameliorate Diabetic Nephropathy via GPR43-Mediated Inhibition of Oxidative Stress and NF-kappaB Signaling. Oxidative Med. Cell. Longev. 2020, 2020, 4074832. [Google Scholar] [CrossRef] [PubMed]
- Michael, O.; Dibia, C.; Soetan, O.; Adeyanju, O.; Oyewole, A.; Badmus, O.; Adetunji, C.; Soladoye, A. Sodium acetate prevents nicotine-induced cardiorenal dysmetabolism through uric acid/creatine kinase-dependent pathway. Life Sci. 2020, 257, 118127. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Probiotic | Study Design | Tissue/Outcomes | Proposed Mechanisms | Ref. |
|---|---|---|---|---|
| L. casei 01 | Male C57BL/6 mice were grouped as CS (12 cigarettes per day) + regular chow, CS + conventional cheese (daily), CS + probiotic cheese (including L. casei 01, 3 × 108 to 3 × 109 CFU/30 g/day), all for 5 days. | Lung: Oxidative stress ↓ Lipid peroxidation ↓ (lung and BAL) Leukocyte count (BAL) ↓ Gut: Oxidative stress ↓ Liver: Oxidative stress ↓ | Probiotic cheese reduces CS-induced oxidative stress in the lungs, gut, and liver. | [97] |
| L. rhamnosus | C57Bl/6 mice were grouped as control (pure air inhalation for 60 days), COPD (14 CS inhalations for 60 days), L. rhamnosus (109 CFU/3 days/week) + COPD. | Lung: Lung remodeling ↓ Pro-Inflammatory mediators ↓ Anti-inflammatory mediator ↑ | L. rhamnosus mediates the balance between pro-inflammatory and anti-inflammatory cytokines to control airway and lung remodeling in COPD. | [98] |
| L. acidophilus, E. faecium, Bacillus subtilis, and B. bifidum | Rats were divided into control (without CS or probiotic), probiotic, CS, and CS + probiotic groups. The probiotic included Lactobacillus acidophilus, Enterococcus faecium, Bacillus subtilis, and Bifidobacterium bifidum at 1 × 109, 2.1 × 109, 2.9 × 109, and 2 × 109 CFU, respectively, incorporated in 1 kg of feed. It was administered for 6 months. | Bone: Bone mineral density ↑ | Probiotic intervention increased bone mineral density and consequently exerted a protective effect on mandibular bone structures in rats exposed to CS. | [99] |
| L. plantarum 299v | In a controlled, double-blind clinical trial, 36 healthy smokers were administered 400 mL of a rose-hip drink with L. plantarum 299v (5 × 107 CFU/mL) daily for 6 weeks; the control group also received the same volume of drink but without the probiotic for 6 weeks. | Peripheral Blood: Systolic blood pressure ↓ Plasma IL-6 ↓ Lipid peroxidation and oxidative stress ↓ | In smokers, L. plantarum 299v treatment reduces systolic blood pressure, inflammation, and lipid oxidation, thus alleviating cardiovascular risk factors. | [100] |
| VSL#3 (L. plantarum, L. casei, L. acidophilus, L. delbrueckii subsp. Bulgaricus, B. infantis, B. longum, B. breve, and S. salivarius subsp. Thermophilus) | APCmin/+ or wild-type mice were exposed to smoke from five cigarettes per day for 4, 8, or 12 weeks. Subsequently, 15 mg of VSL#3 (Grifols, Barcelona, Spain [101]) was dissolved in 200 µL of PBS and administered through oral gavage. Intestinal permeability and APCmin/+-associated colorectal cancer cell growth were analyzed. | Gut: Colorectal cancer tissue numbers ↓ Intestinal permeability ↓ | VSL#3 reduces CRC cell population and intestinal permeability in APCmin/+ mice exposed to CS. | [102] |
| L. paracasei L9 | Mice with asthma [induced through 21 days of ovalbumin (OVA) sensitization and challenge model] were exposed to PM2.5 (from an urban area of Beijing) twice on the last challenge, followed by orally Lactobacillus paracasei L9 administration at 4 × 109 CFU/mouse daily. | Lung: Airway hyperresponsiveness (AHR) ↓ Eosinophil/neutrophil ↓ Th2-related cytokines ↓ Th1 related IFN-γ ↑ IL-17A ↓ | Oral administration of L. paracasei L9 diminishes PM2.5-mediated enhancement of the airway allergic response and hyperresponsiveness in a mouse model of asthma. | [103] |
| B. lactis BB-12 | Female BALB/c mice received pharyngeal aspiration with either sham treatment or PM2.5-containing aerosols. Before treatment, mice were fed either a regular chow or a supplemental diet (including docosahexaenoic acid, vitamins C and E, and B. lactis BB-12). | Lung: Inflammation ↓ | Supplemental diet reduces PM2.5-induced lung inflammation. | [104] |
| LcS | In a placebo-controlled, double-blind clinical trial, 72 healthy male smokers were randomly divided for the intake of LcS powder (4 × 1010 CFU/daily) or placebo for 3 weeks. | Peripheral blood: NK cell activity ↑ CD16+ cell number ↑ | LcS intake in male smokers is associated with decreased cytotoxic activity and CD16+ cell numbers. | [105] |
| Ecologic® Barrier 849 (B. bifidum W23, B. lactis W51, B. lactis W52, L. casei W56, L. salvarius W24, L. acidophilus W37, L. brevis W63, Lc. W19, and Lc. lactis W58) | Mice with a high-fat diet were grouped as control (OA exposure of sterile saline 2 times a week for 28 days), PM (OA exposure of 35 µg diesel exhaust particles 2 times a week for 28 days), PM+ Ecologic® Barrier 849 (~7.5 × 108 CFU/days/28 days). | Gut: Intestinal integrity ↑ Inflammation ↓ | Probiotics-Ecologic® Barrier 849 with the protective effect on regulating inflammatory responses and intestines integrity following inhaled diesel exhaust particles. | [106] |
| LGG | Rats were grouped as control (normal air exposure for 112 days), PM2.5 (exposure in PM2.5 chamber for 112 days), PM2.5+ LGG group (109 CFU/day from day 84 to day 112). | Lung: Inflammation ↓ Th17/Treg balance ↑ Gut: Beneficial bacteria ↑ Bacteria associated with inflammation ↓ | LGG intervention with preventive effects on the PM2.5-induced inflammatory response, Th17/Treg imbalance and intestinal internal environment instability. | [5] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, T.-Y.; Chang, C.-C.; Luo, C.-S.; Chen, K.-Y.; Yeh, Y.-K.; Zheng, J.-Q.; Wu, S.-M. Targeting Lung–Gut Axis for Regulating Pollution Particle–Mediated Inflammation and Metabolic Disorders. Cells 2023, 12, 901. https://doi.org/10.3390/cells12060901
Cheng T-Y, Chang C-C, Luo C-S, Chen K-Y, Yeh Y-K, Zheng J-Q, Wu S-M. Targeting Lung–Gut Axis for Regulating Pollution Particle–Mediated Inflammation and Metabolic Disorders. Cells. 2023; 12(6):901. https://doi.org/10.3390/cells12060901
Chicago/Turabian StyleCheng, Tzu-Yu, Chih-Cheng Chang, Ching-Shan Luo, Kuan-Yuan Chen, Yun-Kai Yeh, Jing-Quan Zheng, and Sheng-Ming Wu. 2023. "Targeting Lung–Gut Axis for Regulating Pollution Particle–Mediated Inflammation and Metabolic Disorders" Cells 12, no. 6: 901. https://doi.org/10.3390/cells12060901
APA StyleCheng, T.-Y., Chang, C.-C., Luo, C.-S., Chen, K.-Y., Yeh, Y.-K., Zheng, J.-Q., & Wu, S.-M. (2023). Targeting Lung–Gut Axis for Regulating Pollution Particle–Mediated Inflammation and Metabolic Disorders. Cells, 12(6), 901. https://doi.org/10.3390/cells12060901