Abstract
Blood–brain barrier (BBB) dysfunction emerges as one of the mechanisms underlying the induction of seizures and epileptogenesis. There is growing evidence that seizures also affect BBB, yet only scarce data is available regarding serum levels of BBB-associated proteins in chronic epilepsy. In this study, we aimed to assess serum levels of molecules associated with BBB in patients with epilepsy in the interictal period. Serum levels of MMP-9, MMP-2, TIMP-1, TIMP-2, S100B, CCL-2, ICAM-1, P-selectin, and TSP-2 were examined in a group of 100 patients who were seizure-free for a minimum of seven days and analyzed by ELISA. The results were compared with an age- and sex-matched control group. Serum levels of MMP-9, MMP-2, TIMP-1, TIMP-2 and S100B were higher in patients with epilepsy in comparison to control group (p < 0.0001; <0.0001; 0.001; <0.0001; <0.0001, respectively). Levels of CCL-2, ICAM-1, P-selectin and TSP-2 did not differ between the two groups. Serum levels of MMP-9, MMP-2, TIMP-1, TIMP-2 and S100B are elevated in patients with epilepsy in the interictal period, which suggests chronic processes of BBB disruption and restoration. The pathological process initiating epilepsy, in addition to seizures, is probably the factor contributing to the elevation of serum levels of the examined molecules.
Keywords:
MMP-9; MMP-2; TIMP-1; TIMP-2; S100B; blood–brain barrier; inflammation; ECM; epilepsy; seizure 1. Introduction
Epilepsy is a chronic neurological disorder affecting 1% of the world population and the most common neurological disease [1]. Causes of epilepsy comprise a variety of structural and functional changes of the central nervous system (CNS). Moreover, seizures can also occur in the course of other diseases. Diverse mechanisms underlie the initiation of seizures and development of epilepsy, and although epilepsy has been researched for a long time, new hypotheses regarding its pathogenesis are still emerging.
There is growing evidence indicating the importance of BBB dysfunction in the development of seizures and epilepsy. Increased permeability of BBB plays an important role in brain damage and leads to the modification of the extracellular matrix (ECM) [2], influx of blood cells, endo and exogenic substances and pathogens, causing local inflammation and angiogenesis [3]. Proinflammatory mediators increase even further BBB permeability in a positive feedback loop [4]. In the situation of dysfunctional BBB, endothelial cells are activated and interact with circulating leukocytes, which results in leukocyte transmigration through BBB and the following generation of seizures [3,5]. Long-lasting BBB disruption leads to the development of epilepsy—epileptogenesis, causing chronic recurrent epileptic seizures [6].
Regulation of BBB permeability is complex and depends not only on the state of cells that comprise BBB (endothelial cells, pericytes, astrocytic end-feet), but also neighboring and circulating molecules and cells (local and circulating mediators of inflammation, local neurons, myocytes, astrocytes and ECM, blood-borne leukocytes, etc.) [7,8,9,10]. Matrix metalloproteinases (MMPs) modify ECM, modulate inflammation and increase BBB permeability [9,11]. Increased activity of MMPs was observed in epileptogenic zones in patients with focal cortical dysplasia (FCD) [12] and tuberous sclerosis (TSC) [13], two types of epilepsy characterized with high activity of disease. Serum level of MMP-9 was elevated after generalized tonic-clonic seizures (GTCS) [14]. Tissue inhibitors of metalloproteinases (TIMPs) counteract the actions of MMPs and are associated with neuroprotection [15]. Their serum level is increased after GTCS [16,17]. TSP-2 is another protein that plays a restorative role, able to non-specifically inhibit MMPs. Loss of TSP-2 is associated with slower BBB repair and intensification of inflammation [18]. Patients with temporal lobe epilepsy (TLE) had higher serum levels of TSP-2 than the control group [19]. Serum S100B is known as a biomarker of brain damage [20] as it correlates with the albumin quotient. In high concentrations, S100B exacerbates inflammation [21]. S100B is overexpressed in TLE [22] and its level in serum is elevated after seizures [23,24]. Another proinflammatory protein is CCL-2, which participates in leukocyte adhesion [25], modifies ECM [26] and induces microglia [27]. CCL-2 is increased after seizures [28]. ICAM-1 and P-selectin (P-sel) are molecules that participate in leukocyte rolling and binding [29,30], thus enabling leukocyte trafficking through BBB. Serum levels of these molecules were elevated in patients with status epilepticus [31], which might suggest increased BBB permeability and intensification of leukocyte influx to the CNS.
A set of molecules associated with BBB permeability (MMP-2, MMP-9, S100B), restoration (TIMP-1, TIMP-2, TSP-2), neuroinflammation (CCL-2), and endothelial activation (ICAM-1, P-sel) that are affected in epilepsy and can be measured in blood was selected in this study. The aim of the present study was to evaluate serum concentrations of BBB-associated proteins in patients with epilepsy in the interictal period with a control group.
2. Materials and Methods
The patient group consisted of 100 successive adult patients with epilepsy who attended the outpatient clinic of the Institute of Psychiatry and Neurology in Warsaw, Poland. Strict inclusion and exclusion criteria were applied for the recruitment of the study participants (Table 1). We included patients after a 7-day period of seizure freedom based on our previous results [24,32]. The exclusion criteria included factors which could influence the examined markers with the exception of disease activity, which did not affect qualification to the study. The same exclusion criteria were applied for control group. The qualification took place in the period from 20 July 2015 to 21 July 2017. All the patients confirmed their willingness to participate in the study by signing informed consent.

Table 1.
Inclusion and exclusion criteria.
Detailed characteristics of patients were obtained, including demographic data, etiology of epilepsy, type of seizures, age of epilepsy onset, disease duration, anti-seizure medication, and time of seizure freedom. The etiology of epilepsy (genetic, structural-metabolic, unknown) was diagnosed according to the classification proposed by ILAE [34].
Patients were recommended to fast, refrain from smoking, using alcohol and undertaking vigorous physical activities for 24 h before the examination. At the day of examination, between 8 and 9 a.m., blood was collected; afterwards, we performed standard EEG in a 10–20 montage for 30 min. To ascertain similar conditions, patients were asked to rest for 5 to 15 min before blood collection. Samples were centrifuged and the supernatant was divided into two portions. One portion was examined with standard laboratory tests for CRP and the second one was frozen and stored in closed test tubes at −80 °C for further analysis, when it was thawed before the immunoenzymatic testing.
We analyzed a panel of nine BBB-associated proteins (MMP-2, MMP-9, TIMP-1, TIMP-2, S100B, TSP-2, CCL-2, ICAM-1, P-sel) using sandwich-type ELISA kits following the manufacturers’ instructions. For the S100B kit, the producer was Merck Millipore, Darmstadt, Germany, and all the remaining proteins were analyzed using kits by R&D Systems, Minneapolis, MN, USA. Multiskan Go microplate reader (Thermo Scientific, Waltham, MA, USA) was used to acquire data from the reactions. Calculations of protein concentrations were carried out following the manufacturers’ instructions.
Preliminarily, descriptive statistics (means, standard deviation, percentiles, and correlations) were used to analyze the data. Depending on the distributions of probability, one-dimensional calculations were performed using Student’s or Wilcoxon’s tests for continuous variables, and chi-square test or Fisher’s exact test were used for nominal variables. We used the generalized linear mixed models (GLMMs) to identify variables influencing the serum levels of BBB-associated proteins. For the optimal model selection, Akaike information criterion (AIC) statistics were used, as this method is asymptotically equivalent to resampling methods [35]. Model selection was performed according to the practical information–theoretic approach [36]. The majority of the variables were modeled with the assumption of normal distribution, gamma distribution was optimal in few cases and variables with over 10% of zero values were analyzed using the Tweedie distribution. SAS package (SAS/STAT rel. 15.1) was used for calculations. The p value of 0.05 was considered statistically significant. There was no missing data. We performed the data analysis from 5 May 2016 to 13 August 2021.
The preparation of the manuscript followed the STROBE checklist [37].
3. Results
3.1. Research Group
The research group consisted primarily of 115 patients, 15 of which were excluded (see Figure 1), yielding 100 patients, 52 women and 48 men. The mean age of the patients was 43.01 ± 1.53 years, and the median was 39 years. There were no differences in age between men and women. All 100 patients completed follow-up and, afterwards, were analyzed in the study. The patients were examined after a minimum 7-day period of seizure freedom. Mean duration of seizure freedom was 432.64 ± 66.50 days, median was 140 days (range 7–4036 days) and 35% of patients did not experience seizures during the last year.
Figure 1.
Study flow chart.
As far as epilepsy etiology is concerned, there were 35 patients with genetic epilepsy (juvenile myoclonic epilepsy and idiopathic genetic epilepsy), 50 patients with structural-metabolic epilepsy and 15 patients with epilepsy of unknown etiology. In the group of patients with structural-metabolic epilepsy, the largest subgroup were patients with post-traumatic epilepsy (14 patients) and post-stroke epilepsy (12 patients). The remaining subgroups were 10 patients with mesial temporal lobe epilepsy (MTLE), two of whom had confirmed hippocampal sclerosis (HS), six patients with neonatal hypoxic-ischemic encephalopathy, five patients with congenital malformations of CNS, and three patients with epilepsy developed after CNS infection.
A total of 45 patients had GTCS (both primary and secondary), 17 patients had focal motor seizures and 38 patients had both focal seizures and secondary GTCS. EEG examination carried out after blood collection was normal in 45% of patients. In 17%, abnormal background activity was observed. Focal slow waves were recorded in 49% of patients and 17% had epileptiform discharges without clinical manifestation. Out of 100 patients, 99 were treated and one patient had newly diagnosed epilepsy and was analyzed before introduction of medication. A total of 65 patients were on monotherapy and 34 patients needed polytherapy (31 with two, and three with three drugs).
The demographic characteristics of the research and control group are shown in Table 2.
Table 2.
Demographic characteristics of research and control group.
3.2. Control Group
The control group consisted of 100 sex- and age-matched subjects, 52 women and 48 men, and did not differ from research group in terms of sex and age. The mean age was 42.16 ± 1.58 years, the median 39 years, min–max was 17–84 years. There were no differences in age between the men and women in control group.
3.3. Serum Levels of BBB-Associated Proteins
Serum levels of MMP-9, MMP-2, TIMP-1, TIMP-2, S100B, proteins associated with increased BBB permeability, were higher in the research group than in the control group (Table 3). MMP-2/TIMP-2 ratio was also higher in the research group. Serum levels of the proteins associated with activation of endothelium (P-sel, ICAM-1), inflammation (CCL-2), and synaptogenesis (TSP-2) did not differ between the two groups.
Table 3.
Means and SEM values of examined proteins. Wilcoxon’s test.
3.4. Influence of Demographic Factors on the Serum Levels of BBB-Associated Proteins
We did not observe significant correlations between age and serum levels of BBB-associated proteins in the research group. Interestingly, there was a weak positive correlation between age and serum CCL-2 level in the control group (r = 0.24, p = 0.0151; Figure 2).
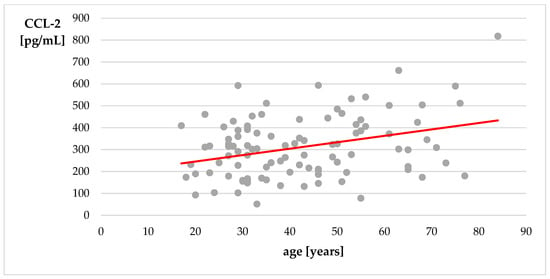
Figure 2.
Correlation of age with level of CCL-2—control group. Spearman’s correlation coefficient.
Serum levels of TIMP-2 were higher in women both in the research and in the control group. Mean serum levels of TIMP-2 in women with epilepsy were 155.91 ± 5.52 ng/mL, median 149.97 ng/mL vs. 142.13 ± 5.46 ng/mL and 133.84 ng/mL in men (p = 0.0172). In the control group, mean serum levels of TIMP-2 in women were 128.03 ± 4.93 ng/mL, median 124.84 ng/mL and in men 111.54 ± 5.03 ng/mL, median 111.14 ng/mL (p = 0.0015; Figure 3).
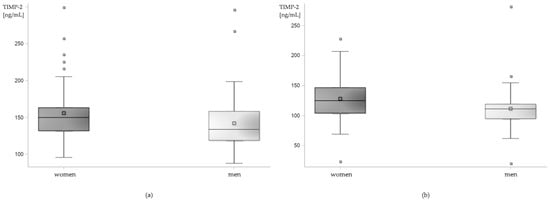
Figure 3.
Sex-related differences in serum level of TIMP-2 in (a) patients with epilepsy and (b) control group. Box plot.
The GLMMs confirmed the sex-related differences in TIMP-2 levels in the research group (least squares means; women: 155.30 ± 4.65 ng/mL, men: 143.91 ± 4.53 ng/mL, p = 0.0397). Additionally, higher levels of S100B were observed in men. Mean level of S100B in men was 77.62 ± 13.13 pg/mL, in women 40.89 ± 6.58 pg/mL (p = 0.0061, least squares means).
3.5. Correlation between the Serum Levels of BBB-Associated Proteins
There was a strong positive correlation between the serum level of MMP-9 and MMP-9/TIMP-1 ratio, and between the level of MMP-2 and MMP-2/TIMP-2 ratio. The level of TIMP-1 showed a strong negative correlation with the MMP-9/TIMP-1 ratio. A moderate positive correlation was observed between the levels of MMP-9 and P-selectin as well as between levels of MMP-2 and TIMP-2. The level of MMP-2 showed a moderate negative correlation with the level of TSP-2. Remaining correlations between the examined proteins were weaker (Table 4).
Table 4.
Correlations between the levels of the BBB-associated proteins. Upper line—correlation coefficient; lower line—level of significance. The values are given only for correlation coefficients ≥ 0.2. Cells are colored according to the color legend depending on the correlation coefficient.
4. Discussion
4.1. Proteins Associated with BBB Disruption and Restoration—MMP-9, MMP-2, TIMP-1, TIMP-2, S100B
We observed higher serum levels of MMP-9, MMP-2, TIMP-1, TIMP-2, and S100B in the research group in comparison to the control group. The examination took place after a 7-day period of seizure freedom to exclude the influence of recent seizures on serum levels of the examined proteins. Elevation of serum level of S100B, which is a well-known biomarker of BBB activation/disruption, suggests that this process is present in patients with epilepsy in the interictal period. Increased activity of MMP-9 [38], overexpression of MMP-2 [12], TIMP-1 [39], TIMP-2 [38], and S100B [22] were found within the epileptogenic zone where repeating epileptic discharges lead to BBB dysfunction. In our study, we showed that these proteins can also be measured in serum and that their levels are higher in patients with epilepsy, which might indicate chronic BBB dysfunction.
4.1.1. Short Characteristics of Proteins Associated with BBB Disruption and Restoration
MMPs are proteolytic enzymes responsible for ECM degradation and an increase in BBB permeability. MMPs participate in various physiological processes, such as cell differentiation and migration, tissue remodeling, cytokine secretion, regulation of trophic factors and maintaining balance between pro- and anti-inflammatory factors [11,40]. They also play an important role in neuroinflammation, neurotoxicity, carcinogenesis and lysis of BBB basal lamina [41,42].
MMP-9, also known as gelatinase B, is a complex protein able to bind a variety of substrates such as collagen type I and IV [43], chemokines [44], interleukins [45], precursors of growth factors [46], tight junctions [47], synaptic proteins [48], and TIMPs. MMP-9-dependent lysis of dystroglycan facilitates leukocyte transmigration through ECM [49] and exacerbates local inflammation. MMP-9 modifies synaptic transmission through changes in ECM, and modulation of glutamatergic transmission by the increasing activity of NMDAR [50] and decreasing the activity of AMPAR [51].
MMP-2 (gelatinase A) is another MMP associated with BBB dysfunction. Similarly to MMP-9, MMP-2 degrades BBB basal lamina and tight junctions [52]. Other substrates of MMP-2 are ECM components, such as laminins, aggrecan, collagen [40], growth factors [53], and IL-1β [54]. An important difference between MMP-2 and MMP-9 lies in their expression. In physiological conditions, expression of MMP-9 is low, but it rises significantly after neuronal depolarization and receptor activation [55,56], and in the presence of proinflammatory factors [57]. MMP-2 is expressed constitutively at higher levels than MMP-9 [58,59], and inflammatory mediators cause a small increase in its expression [60].
TIMPs act as antagonists to MMPs, with TIMP-1 binding preferentially with MMP-9 and TIMP-2 with MMP-2. TIMPs prevent excessive cell damage and cell death [61], influence cell differentiation, growth and migration [61,62,63], impact activation of growth factors [64] and regulate long-term potentiation [65]. Higher expression of TIMPs is associated with lower BBB disruption, which suggests a neuroprotective role of TIMPs within the CNS [66]. The balance between MMPs and TIMPs is essential for maintaining CNS in health [67].
S100B is a calcium-binding protein known as a biomarker of BBB disruption [68]. S100B influences cell proliferation, differentiation and growth [69,70], calcium homeostasis [71] and enzyme activity within the cell [72]. Low amount of S100B is secreted in a constitutive manner and its secretion is increased after CNS damage [73]. S100B acts differently depending on its concentration. At low concentration, it might be associated with increased permeability of BBB without CNS damage [20] and promote nerve growth [74], neuronal survival [75], and long-term potentiation [76]. High concentration of S100B is correlated with neurotoxicity and inflammation [77,78] mediated by binding with RAGE [21], activation of microglia and astroglia and increased expression of proinflammatory mediators [79,80].
4.1.2. Proteins Associated with BBB Disruption and Restoration in Epilepsy
There is growing evidence on the BBB-associated proteins in animal models and specimens obtained from patients during epilepsy surgery. Higher MMP-9 expression in the hippocampus correlates with increased susceptibility to seizures [81] and BBB permeability [82]. In a mouse model of PTZ-induced kindling, degradation of one of ECM components, perineuronal nets, through MMPs was associated with an increased number of seizures [83]. In a rat model of electrically induced epilepsy, increased expression of MMP-2 was found a week after seizures [84].
MMP-9 activity was higher in surgical specimens of patients with MTLE with HS [38], FCD [12] and TSC [13]. Increased MMP-2 expression was found in surgical specimens of adult patients with FCD, but not in children [12].
MMP-9 was increased after GTCS in CSF [85] and serum [14,23]. Serum MMP-9 was also showed to be increased in a general population of patients with epilepsy [86,87], but the precise time from the last seizure was either not mentioned or fitted in a broad range, starting from 0 days. Increased serum level of MMP-9 was found in patients with HS, yet the period of seizure freedom before examination was also unknown and it cannot be excluded that recent seizures influenced these results [88]. Our study shows that the serum level of MMP-9 is increased in patients with epilepsy even without recent seizures, which suggests chronic BBB disruption and ECM alterations in patients with epilepsy.
The study examining CSF MMP-2 after GTCS showed no differences between the level of MMP-2 in the research group and the control group [85]; yet, the control group consisted of patients diagnosed due do other neurologic diseases. There were no statistically significant differences in serum MMP-2 at 1 h after GTCS (although a tendency for higher levels of MMP-2 could be observed), and at 72 h serum MMP-2 was lower than in the control group [16]. In patients with epilepsy examined regardless of the time from last seizure, the serum level of MMP-2 was lower than in control group [89,90,91]. In our study, serum MMP-2 level was higher in patients with epilepsy. As the results are contradictory, further research is needed. The differences might be a result of examining the serum at different time points from last seizure.
In a rat model of electrically induced epilepsy, authors observed increased expression of TIMP-1 and TIMP-2 [84]. A sequential increase in TIMP-1 expression, first in neurons and after 3 days in astrocytes, was showed in a model of kainate-induced epilepsy [92]. In this model, TIMP-1 activity was correlated with neuroprotection. Increased expression of TIMP-2 in microglia was found in dogs with epilepsy [93]. In patients with MTLE with HS, low TIMP-1 activity was found in CA1 and CA2 and high TIMP-1 activity was observed in the neocortex [38]. In the same study, moderate TIMP-2 activity was found in CA1 and high TIMP-2 activity in the neocortex and mossy fibers [38]. Increased levels of TIMP-2 was observed in adult patients with FCD [12].
The serum level of TIMP-1 was higher in children with febrile seizures in course of HHV infection than in children without febrile seizures [94], but lower in children with prolonged febrile seizures with encephalopathy than in children with febrile seizures without encephalopathy, status epilepticus, West syndrome, and control group [95]. A study of serum TIMP-1 kinetics after GTCS showed its increase [17]. In our study, patients examined in the interictal period had increased serum levels of TIMP-1, which suggests long-term activation of BBB restorative processes.
There is little evidence concerning CSF and serum levels of TIMP-2. CSF level of TIMP-2 was not increased in patients with meningitis complicated with epilepsy [96]. Serum TIMP-2 level did not differ between children with febrile seizures, infectious disease with fever, and GTCS [97]; however, the results were not compared with the control group of healthy children. Serum TIMP-2 increased after GTCS [16]. In our study, the serum level of TIMP-2 was increased, which might indicate its persistent neuroprotective activity; nevertheless, more studies are needed to confirm that hypothesis.
Tissue S100B level correlated with seizure frequency and loss of nerve cells in a rat model of epilepsy [98]. In a model of PTZ-induced seizures CSF, the level of S100B was increased during the first 10–30 min from seizures [99]. Increased expression of S100B was shown in patients with temporal lobe epilepsy (TLE) [22]. The serum level of S100B correlates with albumin quotient [100]. After osmotic disruption of BBB, the level of its damage measured by S100B correlated with the occurrence of seizures [101]. Increased S100B levels were observed in the serum of patients with MTLE and TLE [102,103]. Serum S100B level was increased at 30 min, 2, 6, and 24 h after GTCS [23]. However, some studies report different results, indicating no differences in S100B levels between patients with idiopathic epilepsy and control group [104] and no differences after GTCS [105]. Nevertheless, the results of a recent meta-analysis indicate that patients with epilepsy have higher serum levels of S100B [106], which is in concordance with our results and suggests chronic increase of BBB permeability in patients with epilepsy.
The data concerning serum and CSF levels of the examined proteins concentrate mainly on their level shortly after seizures. As existing data indicate that seizures influence levels of the examined proteins, there was a substantial need to fill in this gap with a carefully planned study in which the patients were examined in the interictal period and the duration of period from last seizure was observed. In this study, we tried to approach this task, which resulted in identifying MMP-9, MMP-2, TIMP-1, TIMP-2, and S100B as BBB-associated proteins whose levels are increased regardless of seizure occurrence.
4.2. Other Proteins Associated with BBB—ICAM-1, P-sel, CCL-2, TSP-2
In our study, levels of ICAM-1, P-sel, CCL-2, and TSP-2 did not differ between the patients with epilepsy and control group. ICAM-1 and P-sel are molecules participating in leukocyte transmigration through BBB and their expression is increased after seizures [107]. Expression of ICAM-1 is increased in patients with MTLE [108,109] and TSC [110]. Increased serum levels of ICAM-1 were observed after more than 24 h from seizures, while CSF levels of ICAM-1 remained unchanged [111]. Patients in status epilepticus also had increased serum levels of ICAM-1 and P-sel, although the study did not exclude patients with comorbidities which could affect levels of the examined proteins [31]. Studies assessing the kinetics of ICAM-1 after seizures either showed its increase after 1 h after GTCS [112] or did not show any differences [23]. Serum level of P-sel was increased at 1 h after GTCS [24]. In our study, serum levels of ICAM-1 and P-sel did not differ between the patients with epilepsy in the interictal period and the control group. It is plausible that the activation of endothelium associated with the increase of ICAM-1 and P-sel is a transient process, occurring shortly after seizures, and at that time increasing leukocyte rolling and arrest, while in the interictal period level of endothelial activation is low.
Chemokine CCL-2 (previously known as monocyte chemotactic protein-1, MCP-1) acts as a leukocyte chemoattractant [113], plays a role in leukocyte adhesion and activation [25,114], modifies ECM [115] and tight junction proteins [116], modulates synaptic transmission [117] and plasticity [118] and, by activating CCL-2/CCR2 pathway, leads to the promotion of seizures [119]. CCL-2 expression is increased in brain specimens taken from patients with MTLE, HS and FCD type II [120,121,122]. Level of CCL-2 was increased in CSF of patients with status epilepticus and did not differ between patients with chronic epilepsy and the control group [123]. Serum CCL-2 was increased in patients with HHV infection complicated with acute encephalopathy and seizures [124] and in patients with West syndrome it was either increased [125] or did not differ [126]. CCL-2 might be an important seizure-exacerbating factor by intensifying leukocyte migration, increasing BBB permeability, and tipping the pro-/anti-inflammatory balance in favor of inflammation. The fact that its serum level did not differ between the patients with epilepsy in the interictal period and control group might suggest that main activity of CCL-2 takes place during seizures and that local neuroinflammation in the interictal period might not be enough to influence serum levels of CCL-2.
TSP-2 is a molecule acting as an ECM modulator, modifying cell migration and proliferation [127], promoting excitatory synaptogenesis [128], and also non-specifically inhibiting MMPs [129]. Loss of TSP-2 was associated with the slowing of BBB repair and exacerbation of inflammation [18]. Increased activity of TSP-2 was observed in a model of cortical malformations [130] and serum levels of TSP-2 were higher in patients undergoing epilepsy surgery [19]. In our study, serum TSP-2 did not differ between the research group and the control group, which might be caused by lower disease activity in our patients. Yet, the data regarding serum TSP-2 level is scarce and more research is needed.
4.3. Influence of Demographic Factors on the Serum Levels of BBB-Associated Proteins
4.3.1. Age
Serum levels of CCL-2 showed a weak positive correlation with age in the control group. No other BBB-associated proteins showed correlation with age in control group and patients with epilepsy. Higher serum levels of CCL-2 were observed in patients with frailty syndrome, which was considered to support the connection between CCL-2 and aging [131]. CCL-2 is suggested to promote modification of cell phenotype to the senescence-associated secretory phenotype (SASP) [132] and induce inflammaging, chronic inflammation associated with aging [133]. Positive correlation of CCL-2 with age was also shown in two studies on a healthy population [134,135]. The results of our study replicate previous findings and are in line with the theory of inflammaging.
Studies concerning correlations of serum levels of MMP-9 [136,137,138,139], MMP-2 [136,137,140], TIMP-1 [136,138,141], TIMP-2 [136,138,140] with age are either contradictory or show weak correlations with this factor. In the majority of these studies, serum S100B did not correlate with age in adult patients [32,101,142], while a negative correlation was observed in patients under 20 years of age [142]. Serum levels of P-sel [31,32,143], ICAM-1 [31,32,143,144], and TSP-2 [32,145,146] showed no correlation with age. The data suggests the lack of strong association between the serum levels of the examined proteins and age and is in line with the results of our study.
4.3.2. Sex
We observed sex-dependent differences in serum levels of TIMP-2 and S100B. She serum level of TIMP-2 was higher in women both in control and in research group and the serum level of S100B was higher in men with epilepsy, while no differences were observed in the control group. Interestingly, existing data did not show differences in the serum levels of TIMP-2 of men and women [32,136,138,147]. This might suggest that factors other than sex influenced TIMP-2 level in our patients and control group, but also that a meta-analysis of a larger amount of data would be helpful to shed more light on this topic.
In our study, the serum level of S100B did not differ between women and men in the control group, which is in line with the results obtained in the literature [32,142,148,149]. Yet, in patients with epilepsy, the serum S100B level was higher in men. This finding is in contrast with previous results in which either no differences in S100B level were found between women and men with epilepsy [32,150] or higher levels of S100B were observed in women with MTLE after seizures [102]. Elevated serum S100B in men in our group of patients might be a result of different etiology of disease between the examined men and women, as there was a majority of men in the subgroup of patients with structural-metabolic epilepsy; however, a study in a larger group of patients is needed to determine possible differences in S100B level in women and men.
Both our study and previous studies examining the serum level of MMP-9 [14,136,138], MMP-2 [32,136,138], TIMP-1 [32,136,138,151], ICAM-1 [32,143], P-sel [32,143], and CCL-2 [152,153] with respect to sex-related differences did not show differences between the serum levels of these proteins in men and women. The serum level of TSP-2 in the literature either did not differ [145,146] or was higher in women [32]. Our study did not show differences in the serum TSP-2 level between women and men.
4.4. Correlation between the Serum Levels of BBB-Associated Proteins
We analyzed correlations between serum levels of the examined molecules and found a strong positive correlation between the serum level of MMP-9 and MMP-9/TIMP-1 ratio and between MMP-2 and the MMP-2/TIMP-2 ratio. Concurrently, TIMP-1 showed a strong negative correlation with the MMP-9/TIMP-1 ratio, but TIMP-2 showed no significant correlation with the MMP-2/TIMP-2 ratio. These results indicate that while the MMP-9/TIMP-1 ratio depends on the levels of MMP-9 and TIMP-1 in a comparable way, the MMP-2/TIMP-2 ratio is mainly influenced by serum MMP-2.
A weak negative correlation was observed between the serum level of MMP-9 and TIMP-1. MMP-9 is inhibited by TIMP-1, but it also indirectly influences TIMP-1 expression [154] via IL-1β and TNF-α [155,156]. In previous studies, a positive correlation was shown between CSF TIMP-1 and MMP-9 in patients with bacterial meningitis [96]; however, disease activity was higher in these patients and proteins were measured in CSF, which may explain the contradictory results.
The serum MMP-2 level showed a moderate positive correlation with the level of TIMP-2. A similar correlation was observed in the literature in glioblastoma cells [157]. In a study on the kinetics of serum MMP-2 and TIMP-2 after GTCS, a moderate positive correlation was observed at 72 h from seizures and weaker correlations were found at earlier time points [32]. TIMP-2 is an inhibitor of MMP-2, but it also enables activation of proMMP-2 to active MMP-2 [158]. The lack of correlation between serum level of TIMP-2 and MMP-2/TIMP-2 ratio might result from constitutive expression of TIMP-2 and less pronounced differences between the serum TIMP-2 levels in patients with epilepsy and control group.
We observed a moderate positive correlation between the serum level of MMP-9 and P-sel. A possible explanation for this correlation is MMP-9-dependent release of proinflammatory mediators, which can increase redistribution of P-sel from granules to cell surface [159]. After GTCS, the serum levels of MMP-9 and P-sel showed a weak positive correlation [32].
A moderate negative correlation was observed between the serum level of MMP-2 and TSP-2. TSP-2 is a non-specific inhibitor of MMPs [129] and its loss is associated with higher levels of MMPs [18] which may explain this correlation, especially in light of a weak negative correlation observed between serum level of TSP-2 and MMP-9. To date, only one study has analyzed the correlation between serum MMPs and TSP-2, in which no correlation was observed between serum levels of those proteins after GTCS [32]. To our knowledge, the present study is the first one that analyzes the correlation between serum levels of TSP-2 and MMPs in patients with stable epilepsy.
4.5. Strengths and Limitations
Our study addresses the important question of circulating proteins associated with BBB in patients with epilepsy in the interictal period. To date, most of the studies analyzed serum levels of BBB-associated proteins in a short period of time after seizures or in specific clinical situations where seizures were a co-symptom of other diseases. In these patients, changes in serum levels of BBB-associated proteins might reflect recent seizures. The design of our study focused on patients in the interictal period to avoid the influence of seizures. The period of seizure freedom was based on our previous studies [24,32]. Our study shows that serum levels of proteins associated with BBB disruption and restoration are increased in patients with epilepsy even if they have not recently experienced seizures. This observation is in line with the hypothesis of BBB dysfunction as one of the pathogenetic mechanisms of epilepsy and might indicate that serum levels of proteins associated with BBB disruption and restoration reflect chronic processes occurring at the level of BBB. A panel of BBB disruption and restoration proteins could also act as a diagnostic tool in unclear cases suspected of epilepsy.
One of the limitations of the study is its small sample size and the fact that the data are observational. Therefore, it is suggested to consider the results carefully and, whenever possible, compare them with other studies. Accordingly, we discussed our results in comparison with other studies; yet, as some of the observations are novel, it is necessary to carry out further research, preferably in larger patient groups. We selected proteins that are proven to act at the level of BBB, participating in its disruption or restoration, neuroinflammation and activation of endothelial cells, and that are clinically correlated with seizures; however, they also have many more physiological and pathological functions. We are also aware that the proteins that we evaluated may derive from peripheral vascular system and other tissues. Therefore, we designed an extensive list of exclusion criteria to minimize the impact of these factors. Additionally, blood was always collected at the same time of the day to avoid diurnal changes. Taking into consideration short half-life of some of the examined molecules, blood samples were quickly transported to the laboratory with the observance of the same procedures. Patients were advised to avoid strenuous physical activities for 24 h before blood collection to reduce the possible impact of physical exertion on molecule levels.
5. Conclusions
BBB dysfunction and ECM modulation are processes which are increasingly studied in the light of epilepto- and ictogenesis, yet the majority of research focuses on the impact of seizures on serum molecules. Our study addressed patients with epilepsy in the interictal period and tried to answer the question of whether serum levels of BBB-associated proteins are elevated regardless of seizure occurrence. We showed that serum levels of molecules associated with BBB disruption (MMP-2, MMP-9, S100B) and restoration (TIMP-1, TIMP-2) are increased in patients with epilepsy in the interictal period, which might reflect chronic processes taking place at the BBB, even in the absence of seizures. In our study, serum levels of molecules associated with increased activation of endothelium and leukocyte transmigration (ICAM-1, P-sel) did not differ between patients with epilepsy and the control group, which might indicate that activation of endothelial cells occurs only shortly after seizures. The serum level of CCL-2 did not differ between the research and control groups, which might suggest that the intensity of local inflammation is low. There were also no differences in the serum level of TSP-2 that might indicate that its role in chronic BBB restoration in patients with epilepsy is scarce.
The molecules whose levels are chronically elevated in serum of patients with epilepsy (MMP-2, MMP-9, S100B, TIMP-1, TIMP-2) might be considered diagnostic biomarkers of epilepsy. This finding might be useful in uncertain cases under the condition of carefully considering the factors which can influence their levels. As seizures impact serum levels of the BBB-associated molecules, we underline the importance of at least a 7-day period of seizure freedom to ascertain that seizures do not affect levels of the examined molecules in patients in the interictal period [24,32].
Our study underlines the relationship between chronic disruption and restoration of BBB and epilepsy and indicates that BBB is affected in patients with epilepsy regardless of seizure occurrence. Yet, further research in larger cohorts of patients is needed to fully understand the role of BBB in epilepsy.
Author Contributions
Conceptualization, E.B. and I.K.-J.; methodology, E.B. and I.K.-J., funding acquisition, I.K.-J.; data curation, E.B., A.C.; investigation, E.B., A.W.. and A.C.; supervision I.K.-J.; validation, E.B., A.W. and A.C.; visualization, E.B. and I.K.-J.; writing—original draft, E.B.; writing—review and editing, E.B. and I.K.-J. All authors have read and agreed to the published version of the manuscript.
Funding
The analyses performed for this study were financed by an internal grant financed from the Institute of Psychiatry and Neurology under project number 501-15-021-20016 within statutory activity. No additional external funding and support was received for this study.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Ethics Committee of Institute of Psychiatry and Neurology in Warsaw, Poland (protocol code 01/2012, approved on 23 February 2012 and prolonged on 27 February 2015, and 9 February 2018).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data set can be obtained at https://docs.google.com/spreadsheets/d/1k0U_4ND0F45Wp2wqNxFSg4O4G7WUQhZg/edit?usp=share_link&ouid=108322422041402375271&rtpof=true&sd=true (accessed on 6 December 2022).
Acknowledgments
The authors are grateful to all the patients who spent their time participating in this study and whose cooperation made it possible.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Löscher, W.; Klitgaard, H.; Twyman, R.E.; Schmidt, D. New avenues for anti-epileptic drug discovery and development. Nat. Rev. Drug Discov. 2013, 12, 757–776. [Google Scholar] [CrossRef] [PubMed]
- Baeten, K.M.; Akassoglou, K. Extracellular matrix and matrix receptors in blood-brain barrier formation and stroke. Dev. Neurobiol. 2011, 71, 1018–1039. [Google Scholar] [CrossRef] [PubMed]
- Fabene, P.F.; Mora, G.N.; Martinello, M.; Rossi, B.; Ottoboni, L.; Bach, S.; Angiari, S.; Benati, D.; Zanetti, L.; Schio, F.; et al. A role for leukocyte-endothelial adhesion mechanisms in epilepsy. Nat. Med. 2008, 14, 1377–1383. [Google Scholar] [CrossRef]
- Vezzani, A.; French, J.; Bartfai, T.; Baram, T.Z. The role of inflammation in epilepsy. Nat. Rev. Neurol. 2011, 7, 31–40. [Google Scholar] [CrossRef]
- Fabene, P.F.; Laudanna, C.; Constantin, G. Leukocyte trafficking mechanisms in epilepsy. Mol. Immunol. 2013, 55, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Seiffert, E.; Dreier, J.P.; Ivens, S.; Bechmann, I.; Tomkins, O.; Heinemann, U.; Friedman, A. Lasting blood-brain barrier disruption induces epileptic focus in the rat somatosensory cortex. J. Neurosci. 2004, 24, 7829–7836. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, C.J.D.; LaRivière, C.G.; Young, J.D.; Cass, C.E.; Baldwin, S.A.; Parkinson, F.E. Purine uptake and release in rat C6 glioma cells: Nucleoside transport and purine metabolism under ATP-depleting conditions. J. Neurochem. 2000, 75, 1528–1538. [Google Scholar] [CrossRef]
- Stamatovic, S.M.; Keep, R.F.; Kunkel, S.L.; Andjelkovic, A. V Potential role of MCP-1 in endothelial cell tight junction “opening”: Signaling via Rho and Rho kinase. J. Cell Sci. 2003, 116, 4615–4628. [Google Scholar] [CrossRef]
- Haorah, J.; Ramirez, S.H.; Schall, K.; Smith, D.; Pandya, R.; Persidsky, Y. Oxidative stress activates protein tyrosine kinase and matrix metalloproteinases leading to blood-brain barrier dysfunction. J. Neurochem. 2007, 101, 566–576. [Google Scholar] [CrossRef]
- da Fonseca, A.C.C.; Matias, D.; Garcia, C.; Amaral, R.; Geraldo, L.H.; Freitas, C.; Lima, F.R.S. The impact of microglial activation on blood-brain barrier in brain diseases. Front. Cell Neurosci. 2014, 8, 1–13. [Google Scholar] [CrossRef]
- Verslegers, M.; Lemmens, K.; Van Hove, I.; Moons, L. Matrix metalloproteinase-2 and -9 as promising benefactors in development, plasticity and repair of the nervous system. Prog. Neurobiol. 2013, 105, 60–78. [Google Scholar] [CrossRef] [PubMed]
- Konopka, A.; Grajkowska, W.; Ziemiańska, K.; Roszkowski, M.; Daszkiewicz, P.; Rysz, A.; Marchel, A.; Koperski, L.; Wilczyński, G.M.; Dzwonek, J. Matrix metalloproteinase-9 (MMP-9) in human intractable epilepsy caused by focal cortical dysplasia. Epilepsy Res. 2013, 104, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yu, S.; Zhang, C.; Shu, H.; Liu, S.; An, N.; Yang, M.; Yin, Q.; Yang, H. Increased expression of matrix metalloproteinase 9 in cortical lesions from patients with focal cortical dysplasia type IIb and tuberous sclerosis complex. Brain Res. 2012, 1453, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Cudna, A.; Jopowicz, A.; Mierzejewski, P.; Kurkowska-Jastrzębska, I. Serum metalloproteinase 9 levels increase after generalized tonic-clonic seizures. Epilepsy Res. 2017, 129, 33–36. [Google Scholar] [CrossRef]
- Tan, H.K.; Heywood, D.; Ralph, G.S.; Bienemann, A.; Baker, A.H.; Uney, J.B. Tissue inhibitor of metalloproteinase 1 inhibits excitotoxic cell death in neurons. Mol. Cell Neurosci. 2003, 22, 98–106. [Google Scholar] [CrossRef]
- Cudna, A.; Bronisz, E.; Jopowicz, A.; Kurkowska-Jastrzębska, I. Metalloproteinase 2 and its Inhibitors Expression in Serum after Tonic-Clonic Seizures. In Proceedings of the 13th European Congress on Epileptology, Vienna, Austria, 26–30 August 2018. [Google Scholar]
- Cudna, A.; Jopowicz, A.; Mierzejewski, P.; Kurkowska-Jastrzębska, I. Serum MMP-9 and TIMP-1 level increases after generalised tonic-clonic seizures. In Proceedings of the 2nd Congress of the European Academy of Neurology, Copenhagen, Denmark, 28–31 May 2016. [Google Scholar]
- Tian, W.; Sawyer, A.; Kocaoglu, F.B.; Kyriakides, T.R. Astrocyte-derived thrombospondin-2 is critical for the repair of the blood-brain barrier. Am. J. Pathol. 2011, 179, 860–868. [Google Scholar] [CrossRef]
- Alizada, O.; Akgun, M.Y.; Ozdemir, A.F.; Toklu, S.; Kemerdere, R.; Orhan, B.; Inal, B.B.; Yeni, S.N.; Tanriverdi, T. Circulating Levels of Thrombospondin-1 and Thrombospondin-2 in Patients with Temporal Lobe Epilepsy Before and After Surgery. Turk. Neurosurg. 2021, 31, 228–232. [Google Scholar] [CrossRef]
- Marchi, N.; Cavaglia, M.; Fazio, V.; Bhudia, S.; Hallene, K.; Janigro, D. Peripheral markers of blood-brain barrier damage. Clin. Chim. Acta 2004, 342, 1–12. [Google Scholar] [CrossRef]
- Hofmann, M.A.; Drury, S.; Fu, C.; Qu, W.; Taguchi, A.; Lu, Y.; Avila, C.; Kambham, N.; Bierhaus, A.; Nawroth, P.; et al. RAGE mediates a novel proinflammatory axis: A central cell surface receptor for S100/calgranulin polypeptides. Cell 1999, 97, 889–901. [Google Scholar] [CrossRef]
- Griffin, W.S.; Yeralan, O.; Sheng, J.G.; Boop, F.A.; Mrak, R.E.; Rovnaghi, C.R.; Burnett, B.A.; Feoktistova, A.; Van Eldik, L.J. Overexpression of the neurotrophic cytokine S100 beta in human temporal lobe epilepsy. J. Neurochem. 1995, 65, 228–233. [Google Scholar] [CrossRef]
- Nass, R.D.; Wagner, M.; Surges, R.; Holdenrieder, S. Time courses of HMGB1 and other inflammatory markers after generalized convulsive seizures. Epilepsy Res. 2020, 162, 106301. [Google Scholar] [CrossRef] [PubMed]
- Cudna, A.; Jopowicz, A.; Bronisz, E.; Kurkowska-Jastrzębska, I. Blood-brain barrier markers after tonic-clonic seizures. In Proceedings of the 12th European Congress on Epileptology, Prague, Czech Republic, 11–15 September 2016. [Google Scholar]
- Jiang, Y.; Zhu, J.F.; Luscinskas, F.W.; Graves, D.T. MCP-1-stimulated monocyte attachment to laminin is mediated by beta 2-integrins. Am. J. Physiol. 1994, 267, C1112–C1118. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-S.; Kang, S.I.; Lee, S.-Y.; Noh, K.-T.; Kim, E.-C. Involvement of SDF-1 and monocyte chemoattractant protein-1 in hydrogen peroxide-induced extracellular matrix degradation in human dental pulp cells. Int. Endod. J. 2014, 47, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Lv, R.; Xu, X.; Luo, Z.; Shen, N.; Wang, F.; Zhao, Y. Pyrrolidine dithiocarbamate (PDTC) inhibits the overexpression of MCP-1 and attenuates microglial activation in the hippocampus of a pilocarpine-induced status epilepticus rat model. Exp. Ther. Med. 2013, 7, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Arisi, G.M.; Foresti, M.L.; Katki, K.; Shapiro, L.A. Increased CCL2, CCL3, CCL5, and IL-1β cytokine concentration in piriform cortex, hippocampus, and neocortex after pilocarpine-induced seizures. J. Neuroinflammation 2015, 12, 129. [Google Scholar] [CrossRef] [PubMed]
- Lawson, C.; Wolf, S. ICAM-1 signaling in endothelial cells. Pharmacol. Reports 2009, 61, 22–32. [Google Scholar] [CrossRef]
- Takeshita, Y.; Ransohoff, R.M. Inflammatory cell trafficking across the blood-brain barrier: Chemokine regulation and in vitro models. Immunol. Rev. 2012, 248, 228–239. [Google Scholar] [CrossRef]
- Atmaca, M.M.; Telci, A.; Dirican, A.; Gurses, C. Could sP-Selectin and sICAM-1 be potential biomarkers in status epilepticus? Med. Sci. Discov. 2019, 90, 32–40. [Google Scholar] [CrossRef]
- Cudna, A. Activation of Blood-Brain Barrier in Epilepsy-Dynamics of the Expression of Blood-Brain Barrier Markers after Seizures. Doctoral Thesis, Institute of Psychiatry and Neurology, Warsaw, Poland, 2020. [Google Scholar]
- Fisher, R.S.; Acevedo, C.; Arzimanoglou, A.; Bogacz, A.; Helen Cross, J.; Elger, C.E.; Engel Jr, J.; Forsgren, L.; French, J.A.; Glynn, M.; et al. A practical clinical definition of epilepsy. Epilepsia 2014, 55, 475–482. [Google Scholar] [CrossRef]
- Berg, A.T.; Berkovic, S.F.; Brodie, M.J.; Buchhalter, J.; Cross, J.H.; van Emde Boas, W.; Engel, J.; French, J.; Glauser, T.A.; Mathern, G.W.; et al. Revised terminology and concepts for organization of seizures and epilepsies: Report of the ILAE Commission on Classification and Terminology, 2005–2009. Epilepsia 2010, 51, 676–685. [Google Scholar] [CrossRef]
- Konishi, S.; Kitagawa, G. Information Criteria and Statistical Modeling; Springer: New York, NY, USA, 2008. [Google Scholar]
- Burnham, K.P.; Anderson, D. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- von Elm, E.; Altman, D.G.; Egger, M.; Pocock, S.J.; Gøtzsche, P.C.; Vandenbroucke, J.P. The Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) statement: Guidelines for reporting observational studies. J. Clin. Epidemiol. 2008, 61, 344–349. [Google Scholar] [CrossRef]
- Acar, G.; Tanriover, G.; Acar, F.; Demir, R. Increased Expression of Matrix Metalloproteinase-9 in Patients with temporal Lobe Epilepsy. Turk. Neurosurg. 2015, 25, 749–756. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gupta, T.; Kaur, M.; Singla, N.; Radotra, B.; Gupta, K.S.; Sahni, D. Mesial Temporal Lobe Epilepsy: Role of Reelin Signalling Pathway. Oral Present. 2019 Annu. Meet. Congr. Neurol. Surg. 2019, 66, 2019. [Google Scholar] [CrossRef]
- Ethell, I.M.; Ethell, D.W. Matrix metalloproteinases in brain development and remodeling: Synaptic functions and targets. J. Neurosci. Res. 2007, 85, 2813–2823. [Google Scholar] [CrossRef]
- Rempe, R.G.; Hartz, A.M.S.; Bauer, B. Matrix metalloproteinases in the brain and blood-brain barrier: Versatile breakers and makers. J. Cereb. Blood Flow Metab. 2016, 36, 1481–1507. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, F.L.; Brites, D.; Brito, M.A. Looking at the blood-brain barrier: Molecular anatomy and possible investigation approaches. Brain Res. Rev. 2010, 64, 328–363. [Google Scholar] [CrossRef] [PubMed]
- Bigg, H.F.; Rowan, A.D.; Barker, M.D.; Cawston, T.E. Activity of matrix metalloproteinase-9 against native collagen types I and III. FEBS J. 2007, 274, 1246–1255. [Google Scholar] [CrossRef] [PubMed]
- Denney, H.; Clench, M.R.; Woodroofe, M.N. Cleavage of chemokines CCL2 and CXCL10 by matrix metalloproteinases-2 and -9: Implications for chemotaxis. Biochem. Biophys. Res. Commun. 2009, 382, 341–347. [Google Scholar] [CrossRef]
- Van Den Steen, P.E.; Proost, P.; Wuyts, A.; Van Damme, J.; Opdenakker, G. Neutrophil gelatinase B potentiates interleukin-8 tenfold by aminoterminal processing, whereas it degrades CTAP-III, PF-4, and GRO-α and leaves RANTES and MCP-2 intact. Blood 2000, 96, 2673–2681. [Google Scholar] [CrossRef]
- Van den Steen, P.E.; Dubois, B.; Nelissen, I.; Rudd, P.M.; Dwek, R.A.; Opdenakker, G. Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9). Crit. Rev. Biochem. Mol. Biol. 2002, 37, 375–536. [Google Scholar] [CrossRef] [PubMed]
- Asahi, M.; Wang, X.; Mori, T.; Sumii, T.; Jung, J.C.; Moskowitz, M.A.; Fini, M.E.; Lo, E.H. Effects of matrix metalloproteinase-9 gene knock-out on the proteolysis of blood-brain barrier and white matter components after cerebral ischemia. J. Neurosci. 2001, 21, 7724–7732. [Google Scholar] [CrossRef] [PubMed]
- Michaluk, P.; Wawrzyniak, M.; Alot, P.; Szczot, M.; Wyrembek, P.; Mercik, K.; Medvedev, N.; Wilczek, E.; De Roo, M.; Zuschratter, W.; et al. Influence of matrix metalloproteinase MMP-9 on dendritic spine morphology. J. Cell Sci. 2011, 124, 3369–3380. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.; Anderson, P.; Durbeej, M.; Van Rooijen, N.; Ivars, F.; Opdenakker, G.; Sorokin, L.M. Dystroglycan is selectively cleaved at the parenchymal basement membrane at sites of leukocyte extravasation in experimental autoimmune encephalomyelitis. J. Exp. Med. 2006, 203, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Gorkiewicz, T.; Szczuraszek, K.; Wyrembek, P.; Michaluk, P.; Kaczmarek, L.; Mozrzymas, J.W. Matrix Metalloproteinase-9 reversibly affects the time course of NMDA-induced currents in cultured rat hippocampal neurons. Hippocampus 2010, 20, 1105–1108. [Google Scholar] [CrossRef] [PubMed]
- Michaluk, P.; Mikasova, L.; Groc, L.; Frischknecht, R.; Choquet, D.; Kaczmarek, L. Matrix Metalloproteinase-9 Controls NMDA Receptor Surface Diffusion through Integrin 1 Signaling. J. Neurosci. 2009, 29, 6007–6012. [Google Scholar] [CrossRef]
- Qin, L.H.; Huang, W.; Mo, X.A.; Chen, Y.L.; Wu, X.H. LPS induces occludin dysregulation in cerebral microvascular endothelial cells via MAPK signaling and augmenting mmp-2 levels. Oxid. Med. Cell Longev. 2015, 2015, 120641. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Estrada, E.Y.; Thompson, J.F.; Liu, W.; Rosenberg, G.A. Matrix Metalloproteinase-Mediated Disruption of Tight Junction Proteins in Cerebral Vessels is Reversed by Synthetic Matrix Metalloproteinase Inhibitor in Focal Ischemia in Rat. J. Cereb. Blood Flow Metab. 2007, 27, 697–709. [Google Scholar] [CrossRef]
- Overall, C. Molecular Determinants of Metalloproteinase Substrate Specificity: Matrix Metalloproteinase Substrate Binding Domains, Modules, and Exosites. Mol. Biotechnol. 2002, 22, 51–86. [Google Scholar] [CrossRef]
- Rylski, M.; Amborska, R.; Zybura, K.; Michaluk, P.; Bielinska, B.; Konopacki, F.A.; Wilczynski, G.M.; Kaczmarek, L. JunB is a repressor of MMP-9 transcription in depolarized rat brain neurons. Mol. Cell Neurosci. 2009, 40, 98–110. [Google Scholar] [CrossRef]
- Meighan, S.E.; Meighan, P.C.; Choudhury, P.; Davis, C.J.; Olson, M.L.; Zornes, P.A.; Wright, J.W.; Harding, J.W. Effects of extracellular matrix-degrading proteases matrix metalloproteinases 3 and 9 on spatial learning and synaptic plasticity. J. Neurochem. 2006, 96, 1227–1241. [Google Scholar] [CrossRef]
- Bartholomé, E.J.; Van Aelst, I.; Koyen, E.; Kiss, R.; Willems, F.; Goldman, M.; Opdenakker, G. Human Monocyte-Derived Dendritic Cells Produce Bioactive Gelatinase B: Inhibition by IFN-β. J. Interf. Cytokine Res. 2001, 21, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Planas, A.M.; Solé, S.; Justicia, C. Expression and Activation of Matrix Metalloproteinase-2 and -9 in Rat Brain after Transient Focal Cerebral Ischemia. Neurobiol. Dis. 2001, 8, 834–846. [Google Scholar] [CrossRef]
- Takacs, E.; Nyilas, R.; Szpesei, Z.; Baracskay, P.; Karlsen, B.; Rosvold, T.; Bjørkum, A.A.; Czurko, A.; Kovacs, Z.; Kekesi, A.K.; et al. Matrix metalloproteinase-9 activity increased by two different types of epileptic seizures that do not induce neuronal death: A possible role in homeostatic synaptic plasticity. Neurochem. Int. 2010, 56, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Wu, C.; Korpos, E.; Zhang, X.; Agrawal, S.M.; Wang, Y.; Faber, C.; Schäfers, M.; Körner, H.; Opdenakker, G.; et al. Focal MMP-2 and MMP-9 Activity at the Blood-Brain Barrier Promotes Chemokine-Induced Leukocyte Migration. Cell Rep. 2015, 10, 1040–1054. [Google Scholar] [CrossRef]
- Stetler-Stevenson, W.G. Tissue inhibitors of metalloproteinases in cell signaling: Metalloproteinase-independent biological activities. Sci. Signal. 2008, 1, re6. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.-H.; Park, S.-Y.; Rho, S.B.; Lee, J.-H. Tissue inhibitor of metalloproteinases-3 interacts with angiotensin II type 2 receptor and additively inhibits angiogenesis. Cardiovasc. Res. 2008, 79, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Jaworski, D.M.; Pérez-Martínez, L. Tissue inhibitor of metalloproteinase-2 (TIMP-2) expression is regulated by multiple neural differentiation signals. J. Neurochem. 2006, 98, 234–247. [Google Scholar] [CrossRef]
- Kopitz, C.; Gerg, M.; Bandapalli, O.R.; Ister, D.; Pennington, C.J.; Hauser, S.; Flechsig, C.; Krell, H.-W.; Antolovic, D.; Brew, K.; et al. Tissue Inhibitor of Metalloproteinases-1 Promotes Liver Metastasis by Induction of Hepatocyte Growth Factor Signaling. Cancer Res. 2007, 67, 8615–8623. [Google Scholar] [CrossRef]
- Magnowska, M.; Gorkiewicz, T.; Suska, A.; Wawrzyniak, M.; Rutkowska-Wlodarczyk, I.; Kaczmarek, L.; Wlodarczyk, J. Transient ECM protease activity promotes synaptic plasticity. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef]
- Tejima, E.; Guo, S.; Murata, Y.; Arai, K.; Lok, J.; van Leyen, K.; Rosell, A.; Wang, X.; Lo, E.H. Neuroprotective effects of overexpressing tissue inhibitor of metalloproteinase TIMP-1. J. Neurotrauma 2009, 26, 1935–1941. [Google Scholar] [CrossRef]
- Moore, C.S.; Crocker, S.J. An Alternate Perspective on the Roles of TIMPs and MMPs in Pathology. Am. J. Pathol. 2012, 180, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Murcko, R.; Marchi, N.; Bailey, D.; Janigro, D. Diagnostic biomarker kinetics: How brain-derived biomarkers distribute through the human body, and how this affects their diagnostic significance: The case of S100B. Fluids Barriers CNS 2022, 19, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Arcuri, C.; Bianchi, R.; Brozzi, F.; Donato, R. S100B increases proliferation in PC12 neuronal cells and reduces their responsiveness to nerve growth factor via Akt activation. J. Biol. Chem. 2005, 280, 4402–4414. [Google Scholar] [CrossRef] [PubMed]
- Riuzzi, F.; Sorci, G.; Donato, R. S100B protein regulates myoblast proliferation and differentiation by activating FGFR1 in a bFGF-dependent manner. J. Cell Sci. 2011, 124, 2389–2400. [Google Scholar] [CrossRef]
- Xiong, Z.G.; O’Hanlon, D.; Becker, L.E.; Roder, J.; MacDonald, J.F.; Marks, A. Enhanced calcium transients in glial cells in neonatal cerebellar cultures derived from S100B null mice. Exp. Cell Res. 2000, 257, 281–289. [Google Scholar] [CrossRef]
- Heierhorst, J.; Kobe, B.; Feil, S.C.; Parker, M.W.; Benian, G.M.; Weiss, K.R.; Kemp, B.E. Ca2+/S100 regulation of giant protein kinases. Nature 1996, 380, 636–639. [Google Scholar] [CrossRef]
- Hajduková, L.; Sobek, O.; Prchalová, D.; Bílková, Z.; Koudelková, M.; Lukášková, J.; Matuchová, I. Biomarkers of Brain Damage: S100B and NSE Concentrations in Cerebrospinal Fluid-A Normative Study. Biomed Res. Int. 2015, 2015, 379071. [Google Scholar] [CrossRef]
- Kligman, D.; Marshak, D.R. Purification and characterization of a neurite extension factor from bovine brain. Proc. Natl. Acad. Sci. USA 1985, 82, 7136–7139. [Google Scholar] [CrossRef]
- Winningham-Major, F.; Staecker, J.L.; Barger, S.W.; Coats, S.; Van Eldik, L.J. Neurite extension and neuronal survival activities of recombinant S100β proteins that differ in the content and position of cysteine residues. J. Cell Biol. 1989, 109, 3063–3071. [Google Scholar] [CrossRef]
- Nishiyama, H.; Knöpfel, T.; Endo, S.; Itohara, S. Glial protein S100B modulates long-term neuronal synaptic plasticity. Proc. Natl. Acad. Sci. USA 2002, 99, 4037–4042. [Google Scholar] [CrossRef] [PubMed]
- Businaro, R.; Leone, S.; Fabrizi, C.; Sorci, G.; Donato, R.; Lauro, G.M.; Fumagalli, L. S100B protects LAN-5 neuroblastoma cells against Abeta amyloid-induced neurotoxicity via RAGE engagement at low doses but increases Abeta amyloid neurotoxicity at high doses. J. Neurosci. Res. 2006, 83, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Koppal, T.; Lam, A.G.; Guo, L.; Van Eldik, L.J. S100B proteins that lack one or both cysteine residues can induce inflammatory responses in astrocytes and microglia. Neurochem. Int. 2001, 39, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Villarreal, A.; Seoane, R.; Torres, A.G.; Rosciszewski, G.; Angelo, M.F.; Rossi, A.; Barkert, P.A.; Ramos, A.J. S100B protein activates a RAGE-dependent autocrine loop in astrocytes: Implications for its role in the propagation of reactive gliosis. J. Neurochem. 2014, 131, 190–205. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, R.; Giambanco, I.; Donato, R. S100B/RAGE-dependent activation of microglia via NF-kappaB and AP-1 Co-regulation of COX-2 expression by S100B, IL-1beta and TNF-alpha. Neurobiol. Aging 2010, 31, 665–677. [Google Scholar] [CrossRef]
- Szklarczyk, A.; Lapinska, J.; Rylski, M.; McKay, R.D.G.; Kaczmarek, L. Matrix metalloproteinase-9 undergoes expression and activation during dendritic remodeling in adult hippocampus. J. Neurosci. 2002, 22, 920–930. [Google Scholar] [CrossRef]
- Gorter, J.A.; van Vliet, E.A.; Aronica, E. Status epilepticus, blood–brain barrier disruption, inflammation, and epileptogenesis. Epilepsy Behav. 2015, 49, 13–16. [Google Scholar] [CrossRef]
- Rankin-Gee, E.K.; McRae, P.A.; Baranov, E.; Rogers, S.; Wandrey, L.; Porter, B.E. Perineuronal net degradation in epilepsy. Epilepsia 2015, 56, 1124–1133. [Google Scholar] [CrossRef]
- Gorter, J.A.; Van Vliet, E.A.; Rauwerda, H.; Breit, T.; Stad, R.; Van Schaik, L.; Vreugdenhil, E.; Redeker, S.; Hendriksen, E.; Aronica, E.; et al. Dynamic changes of proteases and protease inhibitors revealed by microarray analysis in CA3 and entorhinal cortex during epileptogenesis in the rat. Epilepsia 2007, 48, 53–64. [Google Scholar] [CrossRef]
- Li, Y.-J.; Wang, Z.-H.; Zhang, B.; Zhe, X.; Wang, M.-J.; Shi, S.-T.; Bai, J.; Lin, T.; Guo, C.-J.; Zhang, S.-J.; et al. Disruption of the blood-brain barrier after generalized tonic-clonic seizures correlates with cerebrospinal fluid MMP-9 levels. J. Neuroinflammation 2013, 10, 80. [Google Scholar] [CrossRef]
- Tao, H.; Gong, Y.; Yu, Q.; Zhou, H.; Liu, Y. Elevated Serum Matrix Metalloproteinase-9, Interleukin-6, Hypersensitive C-Reactive Protein, and Homocysteine Levels in Patients with Epilepsy. J. Interf. Cytokine Res. 2020, 40, 152–158. [Google Scholar] [CrossRef]
- Wang, R.; Zeng, G.Q.; Liu, X.; Tong, R.Z.; Zhou, D.; Hong, Z. Evaluation of serum matrix metalloproteinase-3 as a biomarker for diagnosis of epilepsy. J. Neurol. Sci. 2016, 367, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Quirico-Santos, T.; Nascimento Mello, A.; Casimiro Gomes, A.; de Carvalho, L.P.; de Souza, J.M.; Alves-Leon, S. Increased metalloprotease activity in the epileptogenic lesion—Lobectomy reduces metalloprotease activity and urokinase-type uPAR circulating levels. Brain Res. 2013, 1538, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zeng, G.Q.; Tong, R.Z.; Zhou, D.; Hong, Z. Serum matrix metalloproteinase-2: A potential biomarker for diagnosis of epilepsy. Epilepsy Res. 2016, 122, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Salih, K.S.; Hamdan, F.B.; Al-Mayah, Q.S. Diagnostic value of matrix metalloproteinase-2 and high mobility group box 1 in patients with refractory epilepsy. Egypt. J. Neurol. Psychiatry Neurosurg. 2020, 56, 102. [Google Scholar] [CrossRef]
- Soliman, A.M.; Ebeary, M.E.S.E.; Gouda, T.A.R.; Ebied, A.A. Serum matrix metalloproterinse-2 as a biomarker for diagnosis of idiopathic epilepsy. Zagazig Univ. Med. J. 2019, 25, 155–163. [Google Scholar] [CrossRef]
- Rivera, S.; Tremblay, E.; Timsit, S.; Canals, O.; Ben-Ari, Y.; Khrestchatisky, M. Tissue inhibitor of metalloproteinases-1 (TIMP-1) is differentially induced in neurons and astrocytes after seizures: Evidence for developmental, immediate early gene, and lesion response. J. Neurosci. 1997, 17, 4223–4235. [Google Scholar] [CrossRef]
- Stein, V.M.; Genini, S.; Puff, C.; Baumgärtner, W.; Tipold, A. Seizure activity in dogs is associated with enhanced TIMP-2 expression of microglia. Vet. Immunol. Immunopathol. 2012, 146, 101–105. [Google Scholar] [CrossRef]
- Kittaka, S.; Hasegawa, S.; Ito, Y.; Ohbuchi, N.; Suzuki, E.; Kawano, S.; Aoki, Y.; Nakatsuka, K.; Kudo, K.; Wakiguchi, H.; et al. Serum levels of matrix metalloproteinase-9 and tissue inhibitor of metalloproteinases-1 in human herpesvirus-6–infected infants with or without febrile seizures. J. Infect. Chemother. 2014, 20, 716–721. [Google Scholar] [CrossRef]
- Suenaga, N.; Ichiyama, T.; Kubota, M.; Isumi, H.; Tohyama, J.; Furukawa, S. Roles of matrix metalloproteinase-9 and tissue inhibitors of metalloproteinases 1 in acute encephalopathy following prolonged febrile seizures. J. Neurol. Sci. 2008, 266, 126–130. [Google Scholar] [CrossRef]
- Leppert, D.; Leib, S.L.; Grygar, C.; Miller, K.M.; Schaad, U.B.; Holländer, G. a Matrix metalloproteinase (MMP)-8 and MMP-9 in cerebrospinal fluid during bacterial meningitis: Association with blood-brain barrier damage and neurological sequelae. Clin. Infect. Dis. 2000, 31, 80–84. [Google Scholar] [CrossRef]
- Haberlandt, E.; Rauchenzauner, M.; Morass, M.; Wondrak, P.; Bürgi, S.S.; Rostásy, K.; Karall, D. Matrix-metalloproteinases and proinflammatory cytokines in children with febrile convulsions and epilepsy-Cause or consequence? Epilepsy Res. 2013, 105, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Filibian, M.; Frasca, A.; Maggioni, D.; Micotti, E.; Vezzani, A.; Ravizza, T. In vivo imaging of glia activation using 1H-magnetic resonance spectroscopy to detect putative biomarkers of tissue epileptogenicity. Epilepsia 2012, 53, 1907–1916. [Google Scholar] [CrossRef] [PubMed]
- Oses, J.P.; Leke, R.; Portela, L.V.; Lara, D.R.; Schmidt, A.P.; Casali, E.A.; Wofchuk, S.; Souza, D.O.; Sarkis, J.J.F. Biochemical brain markers and purinergic parameters in rat CSF after seizure induced by pentylenetetrazol. Brain Res. Bull. 2004, 64, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Blyth, B.J.; Farhavar, A.; Gee, C.; Hawthorn, B.; He, H.; Nayak, A.; Stöcklein, V.; Bazarian, J.J. Validation of serum markers for blood-brain barrier disruption in traumatic brain injury. J. Neurotrauma 2009, 26, 1497–1507. [Google Scholar] [CrossRef] [PubMed]
- Marchi, N.; Angelov, L.; Masaryk, T.; Fazio, V.; Granata, T.; Hernandez, N.; Hallene, K.; Diglaw, T.; Franic, L.; Najm, I.; et al. Seizure-promoting effect of blood-brain barrier disruption. Epilepsia 2007, 48, 732–742. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Li, J.; Sun, W.; Feng, L.; Li, L.; Liu, A.; Li, J.; Mao, W.; Wei, H.; Gao, L.; et al. Elevated plasma S100B concentration is associated with mesial temporal lobe epilepsy in Han Chinese: A case-control study. Neurosci. Lett. 2010, 484, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Calik, M.; Abuhandan, M.; Sonmezler, A.; Kandemir, H.; Oz, I.; Taskin, A.; Selek, S.; Iscan, A. Elevated serum S-100B levels in children with temporal lobe epilepsy. Seizure 2013, 22, 99–102. [Google Scholar] [CrossRef]
- Portela, L.V.C.; Tort, A.B.L.; Walz, R.; Bianchin, M.; Trevisol-Bittencourt, P.C.; Wille, P.R.; Cardoso, R.C.; Ishida, M.M.I.; VonWangenheim, A.; Grisard, E.C.; et al. Interictal serum S100B levels in chronic neurocysticercosis and idiopathic epilepsy. Acta Neurol. Scand. 2003, 108, 424–427. [Google Scholar] [CrossRef]
- Palmio, J.; Peltola, J.; Vuorinen, P.; Laine, S.; Suhonen, J.; Keränen, T. Normal CSF neuron-specific enolase and S-100 protein levels in patients with recent non-complicated tonic-clonic seizures. J. Neurol. Sci. 2001, 183, 27–31. [Google Scholar] [CrossRef]
- Liang, K.G.; Mu, R.Z.; Liu, Y.; Jiang, D.; Jia, T.T.; Huang, Y.J. Increased serum S100B levels in patients with epilepsy: A systematic review and meta-analysis study. Front. Neurosci. 2019, 13, 456. [Google Scholar] [CrossRef]
- Librizzi, L.; Regondi, M.C.; Pastori, C.; Frigerio, S.; Frassoni, C.; De Curtis, M. Expression of adhesion factors induced by epileptiform activity in the endothelium of the isolated guinea pig brain in vitro. Epilepsia 2007, 48, 743–751. [Google Scholar] [CrossRef]
- Kan, A.A.; de Jager, W.; de Wit, M.; Heijnen, C.; van Zuiden, M.; Ferrier, C.; van Rijen, P.; Gosselaar, P.; Hessel, E.; van Nieuwenhuizen, O.; et al. Protein expression profiling of inflammatory mediators in human temporal lobe epilepsy reveals co-activation of multiple chemokines and cytokines. J. Neuroinflammation 2012, 9, 207. [Google Scholar] [CrossRef] [PubMed]
- Strauss, K.I.; Elisevich, K.V. Brain region and epilepsy-associated differences in inflammatory mediator levels in medically refractory mesial temporal lobe epilepsy. J. Neuroinflammation 2016, 13, 270. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, M.; Baybis, M.; Newman, D.; Kolson, D.L.; Chen, W.; McKhann, G.; Gutmann, D.H.; Crino, P.B. Expression of ICAM-1, TNF-α, NFκB, and MAP kinase in tubers of the tuberous sclerosis complex. Neurobiol. Dis. 2003, 14, 279–290. [Google Scholar] [CrossRef]
- Luo, J.; Wang, W.; Xi, Z.; Dan, C.; Wang, L.; Xiao, Z.; Wang, X. Concentration of Soluble Adhesion Molecules in Cerebrospinal Fluid and Serum of Epilepsy Patients. J. Mol. Neurosci. 2014, 54, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Cudna, A.; Kurkowska-Jastrzebska, I. Blood–brain barrier markers after acute epileptic seizures. J. Neuroimmunol. 2014, 275, 28. [Google Scholar] [CrossRef]
- Carr, M.W.; Roth, S.J.; Luther, E.; Rose, S.S.; Springer, T.A. Monocyte chemoattractant protein 1 acts as a T-lymphocyte chemoattractant. Proc. Natl. Acad. Sci. USA 1994, 91, 3652–3656. [Google Scholar] [CrossRef]
- Allavena, P.; Bianchi, G.; Zhou, D.; van Damme, J.; Jílek, P.; Sozzani, S.; Mantovani, A. Induction of natural killer cell migration by monocyte chemotactic protein-1, -2 and -3. Eur. J. Immunol. 1994, 24, 3233–3236. [Google Scholar] [CrossRef]
- Park, J.; Ryu, D.R.; Li, J.J.; Jung, D.S.; Kwak, S.J.; Lee, S.H.; Yoo, T.H.; Han, S.H.; Lee, J.E.; Kim, D.K.; et al. MCP-1/CCR2 system is involved in high glucose-induced fibronectin and type IV collagen expression in cultured mesangial cells. Am. J. Physiol. Ren. Physiol. 2008, 295, F749–F757. [Google Scholar] [CrossRef]
- Song, L.; Pachter, J.S. Monocyte chemoattractant protein-1 alters expression of tight junction-associated proteins in brain microvascular endothelial cells. Microvasc. Res. 2004, 67, 78–89. [Google Scholar] [CrossRef]
- Nelson, T.E.; Hao, C.; Manos, J.; Ransohoff, R.M.; Gruol, D.L. Altered hippocampal synaptic transmission in transgenic mice with astrocyte-targeted enhanced CCL2 expression. Brain. Behav. Immun. 2011, 25 (Suppl. 1), S106–S119. [Google Scholar] [CrossRef] [PubMed]
- Kalehua, A.N.; Nagel, J.E.; Whelchel, L.M.; Gides, J.J.; Pyle, R.S.; Smith, R.J.; Kusiak, J.W.; Taub, D.D. Monocyte chemoattractant protein-1 and macrophage inflammatory protein-2 are involved in both excitotoxin-induced neurodegeneration and regeneration. Exp. Cell Res. 2004, 297, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Cerri, C.; Genovesi, S.; Allegra, M.; Pistillo, F.; Püntener, U.; Guglielmotti, A.; Perry, V.H.; Bozzi, Y.; Caleo, M. The Chemokine CCL2 Mediates the Seizure-enhancing Effects of Systemic Inflammation. J. Neurosci. 2016, 37, 1–5. [Google Scholar] [CrossRef]
- Lee, T.-S.; Mane, S.; Eid, T.; Zhao, H.; Lin, A.; Guan, Z.; Kim, J.H.; Schweitzer, J.; King-Stevens, D.; Weber, P.; et al. Gene Expression in Temporal Lobe Epilepsy is Consistent with Increased Release of Glutamate by Astrocytes. Mol. Med. 2007, 13, 1–13. [Google Scholar] [CrossRef]
- Iyer, A.; Zurolo, E.; Spliet, W.G.M.; Van Rijen, P.C.; Baayen, J.C.; Gorter, J.A.; Aronica, E. Evaluation of the innate and adaptive immunity in type I and type II focal cortical dysplasias. Epilepsia 2010, 51, 1763–1773. [Google Scholar] [CrossRef] [PubMed]
- Aalbers, M.W.; Rijkers, K.; Majoie, H.J.M.; Dings, J.T.; Schijns, O.E.M.G.; Schipper, S.; De Baets, M.H.; Kessels, A.; Vles, J.S.H.; Hoogland, G. The influence of neuropathology on brain inflammation in human and experimental temporal lobe epilepsy. J. Neuroimmunol. 2014, 271, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Kothur, K.; Bandodkar, S.; Wienholt, L.; Chu, S.; Pope, A.; Gill, D.; Dale, R.C. Etiology is the key determinant of neuroinflammation in epilepsy: Elevation of cerebrospinal fluid cytokines and chemokines in febrile infection-related epilepsy syndrome and febrile status epilepticus. Epilepsia 2019, 1678–1688. [Google Scholar] [CrossRef]
- Kawamura, Y.; Yamazaki, Y.; Ohashi, M.; Ihira, M.; Yoshikawa, T. Monocyte chemoattractant protein-1 alters expression of tight junction-associated proteins in brain microvascular endothelial cells. J. Med. Virol. 2014, 86, 512–518. [Google Scholar] [CrossRef]
- Yamanaka, G.; Morishita, N.; Morichi, S.; Takeshita, M.; Tomomi, U.; Ishida, Y.; Tomoko, T.; Oana, S.; Watanabe, Y.; Go, S.; et al. Serial Analysis of Multiple Serum Cytokine Responses to Adrenocorticotropic Hormone Therapy in Patients With West Syndrome. J. Child Neurol. 2018, 33, 528–533. [Google Scholar] [CrossRef]
- Shiihara, T.; Miyashita, M.; Yoshizumi, M.; Watanabe, M.; Yamada, Y.; Kato, M. Peripheral lymphocyte subset and serum cytokine profiles of patients with West syndrome. Brain Dev. 2010, 32, 695–702. [Google Scholar] [CrossRef]
- Jayakumar, A.R.; Apeksha, A.; Norenberg, M.D. Role of Matricellular Proteins in Disorders of the Central Nervous System. Neurochem. Res. 2017, 42, 858–875. [Google Scholar] [CrossRef] [PubMed]
- Lau, L.A.; Noubary, F.; Wang, D.; Dulla, C.G. α2δ-1 signaling drives cell death, synaptogenesis, circuit reorganization, and gabapentin-mediated neuroprotection in a model of insult-induced cortical malformation. eNeuro 2017, 4, ENEURO.0316-17.2017. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Kyriakides, T.R.; Bornstein, P. Matricellular proteins as modulators of cell-matrix interactions: Adhesive defect in thrombospondin 2-null fibroblasts is a consequence of increased levels of matrix metalloproteinase-2. Mol. Biol. Cell 2000, 11, 3353–3364. [Google Scholar] [CrossRef] [PubMed]
- Andresen, L.; Hampton, D.; Taylor, A.; Morel, L.; Yang, Y.; Maguire, J.; Dulla, C.G.; Dulla, C. Gabapentin attenuates hyperexcitability in the freeze-lesion model of developmental cortical malformation. Neurobiol. Dis. 2014, 71, 305–316. [Google Scholar] [CrossRef]
- Yousefzadeh, M.J.; Schafer, M.J.; Hooten, N.N.; Atkinson, E.J.; Evans, M.K.; Baker, D.J.; Quarles, E.K.; Robbins, P.D.; Ladiges, W.C.; Lebrasseur, N.K.; et al. Circulating levels of monocyte chemoattractant protein-1 as a potential measure of biological age in mice and frailty in humans. Aging Cell 2017, 17, e12706. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.J.; Lee, H.J.; Heo, J.; Lim, J.; Kim, M.; Kim, M.K.; Nam, H.Y.; Hong, G.H.; Cho, Y.S.; Choi, S.J.; et al. Senescence-Associated MCP-1 Secretion Is Dependent on a Decline in BMI1 in Human Mesenchymal Stromal Cells. Antioxid. Redox Signal. 2016, 24, 471–485. [Google Scholar] [CrossRef]
- Franceschi, C.; Campisi, J. Chronic Inflammation (Inflammaging) and Its Potential Contribution to Age-Associated Diseases. J. Gerontol. Ser. A: Biomed. Sci. Med. Sci. 2014, 69 (Suppl. 1), S4–S9. [Google Scholar] [CrossRef]
- Inadera, H.; Egashira, K.; Takemoto, M.; Ouchi, Y.; Matsushima, K. Increase in circulating levels of monocyte chemoattractant protein-1 with aging. J. Interf. Cytokine Res. 1999, 19, 1179–1182. [Google Scholar] [CrossRef]
- Seidler, S.; Zimmermann, H.W.; Bartneck, M.; Trautwein, C.; Tacke, F. Age-dependent alterations of monocyte subsets and monocyte-related chemokine pathways in healthy adults. BMC Immunol. 2010, 11, 30. [Google Scholar] [CrossRef]
- Tayebjee, M.H.; Lip, G.Y.H.; Blann, A.D.; MacFadyen, R.J. Effects of age, gender, ethnicity, diurnal variation and exercise on circulating levels of matrix metalloproteinases (MMP)-2 and -9, and their inhibitors, tissue inhibitors of matrix metalloproteinases (TIMP)-1 and -2. Thromb. Res. 2005, 115, 205–210. [Google Scholar] [CrossRef]
- Cancemi, P.; Aiello, A.; Accardi, G.; Caldarella, R.; Candore, G.; Caruso, C.; Ciaccio, M.; Cristaldi, L.; Di Gaudio, F.; Siino, V.; et al. The Role of Matrix Metalloproteinases (MMP-2 and MMP-9) in Ageing and Longevity: Focus on Sicilian Long-Living Individuals (LLIs). Mediators Inflamm. 2020, 2020, 8635158. [Google Scholar] [CrossRef] [PubMed]
- Beaudeux, J.L.; Giral, P.; Bruckert, E.; Bernard, M.; Foglietti, M.J.; Chapman, M.J. Serum matrix metalloproteinase-3 and tissue inhibitor of metalloproteinases-1 as potential markers of carotid atherosclerosis in infraclinical hyperlipidemia. Atherosclerosis 2003, 169, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.L.; Mcdonald, V.M.; Baines, K.J.; Oreo, K.M.; Wang, F.; Hansbro, P.M.; Gibson, P.G. Influence of Age, Past Smoking, and Disease Severity on TLR2, Neutrophilic Inflammation, and MMP-9 Levels in COPD. Mediators Inflamm. 2013, 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- Bonnema, D.D.; Webb, C.S.; Pennington, W.R.; Stroud, R.E.; Leonardi, A.E.; Clark, L.L.; McClure, C.D.; Finklea, L.; Spinale, F.G.; Zile, M.R. Effects of age on plasma matrix metalloproteinases (MMPs) and tissue inhibitor of metalloproteinases (TIMPs). J Card. Fail 2007, 13, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Komosinska-Vassev, K.; Olczyk, P.; Winsz-Szczotka, K.; Kuznik-Trocha, K.; Klimek, K.; Olczyk, K. Age- and gender-dependent changes in connective tissue remodeling: Physiological differences in circulating MMP-3, MMP-10, TIMP-1 and TIMP-2 level. Gerontology 2010, 57, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Portela, L.V.C.; Tort, A.B.L.; Schaf, D.V.; Ribeiro, L.; Nora, D.B.; Walz, R.; Rotta, L.N.; Silva, C.T.; Busnello, J.V.; Kapczinski, F.; et al. The serum S100B concentration is age dependent. Clin. Chem. 2002, 48, 950–952. [Google Scholar] [CrossRef]
- Deneva-Koycheva, T.I.; Vladimirova-Kitova, L.G.; Angelova, E.A.; Tsvetkova, T.Z. Serum levels of siCAM-1, sVCAM-1, sE-selectin, sP-selectin in healthy Bulgarian people. Folia Med. (Plovdiv) 2011, 53, 22–28. [Google Scholar] [CrossRef]
- Grothey, A.; Heistermann, P.; Philippou, S.; Voigtmann, R. Serum levels of soluble intercellular adhesion molecule-1 (ICAM-1, CD54) in patients with non-small cell lung cancer: Correlation with histological expression of ICAM-1 and tumour stage. Br. J. Cancer 1998, 77, 801–807. [Google Scholar] [CrossRef]
- Morikawa, N.; Adachi, H.; Enomoto, M.; Fukami, A.; Kumagai, E.; Nakamura, S.; Nohara, Y.; Nakao, E.; Kono, S.; Tsuru, T.; et al. Thrombospondin-2 as a potential risk factor in a general population. Int. Heart J. 2019, 60, 310–317. [Google Scholar] [CrossRef]
- Hanatani, S.; Izumiya, Y.; Takashio, S.; Kimura, Y.; Araki, S.; Rokutanda, T.; Tsujita, K.; Yamamoto, E.; Tanaka, T.; Yamamuro, M.; et al. Circulating thrombospondin-2 reflects disease severity and predicts outcome of heart failure with reduced ejection fraction. Circ. J. 2014, 78, 903–910. [Google Scholar] [CrossRef]
- Young-Min, S.A.; Beeton, C.; Laughton, R.; Plumpton, T.; Bartram, S.; Murphy, G.; Black, C.; Cawston, T.E. Serum TIMP-1, TIMP-2, and MMP-1 in patients with systemic sclerosis, primary Raynaud’s phenomenon, and in normal controls. Ann. Rheum. Dis. 2001, 60, 846–851. [Google Scholar] [PubMed]
- Wiesmann, M.; Missler, U.; Gottmann, D.; Gehring, S. Plasma S-100b protein concentration in healthy adults is age- and sex- independent. Clin. Chem. 1998, 44, 1056–1058. [Google Scholar] [CrossRef] [PubMed]
- Lara, D.R.; Gama, C.S.; Belmonte-De-Abreu, P.; Portela, L.V.C.; Gonçalves, C.A.; Fonseca, M.; Hauck, S.; Souza, D.O. Increased serum S100B protein in schizophrenia: A study in medication-free patients. J. Psychiatr. Res. 2001, 35, 11–14. [Google Scholar] [CrossRef]
- Calik, M.; Abuhandan, M.; Kandemir, H.; Güzel, B.; Solmaz, A.; Celik, H.; Taskin, A.; Iscan, A. Interictal serum S-100B protein levels in intractable epilepsy: A case-control study. Neurosci. Lett. 2014, 558, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Irani, S.R.; Bera, K.; Waters, P.; Zuliani, L.; Maxwell, S.; Zandi, M.S.; Friese, M.A.; Galea, I.; Kullmann, D.M.; Beeson, D.; et al. N-methyl-d-aspartate antibody encephalitis: Temporal progression of clinical and paraclinical observations in a predominantly non-paraneoplastic disorder of both sexes. Brain 2010, 133, 1655–1667. [Google Scholar] [CrossRef] [PubMed]
- Murea, M.; Register, T.C.; Divers, J.; Bowden, D.W.; Carr, J.J.; Hightower, C.R.; Xu, J.; Smith, S.C.; Hruska, K.A.; Langefeld, C.D.; et al. Relationships between serum MCP-1 and subclinical kidney disease: African American-Diabetes Heart Study. BMC Nephrol. 2012, 13, 6–8. [Google Scholar] [CrossRef]
- Mansfield, A.S.; Nevala, W.K.; Dronca, R.S.; Leontovich, A.A.; Shuster, L.; Markovic, S.N. Normal ageing is associated with an increase in Th2 cells, MCP-1 (CCL1) and RANTES (CCL5), with differences in sCD40L and PDGF-AA between sexes. Clin. Exp. Immunol. 2012, 170, 186–193. [Google Scholar] [CrossRef]
- Brew, K.; Dinakarpandian, D.; Nagase, H. Tissue inhibitors of metalloproteinases: Evolution, structure and function. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 2000, 1477, 267–283. [Google Scholar] [CrossRef]
- Van Lint, P.; Libert, C. Chemokine and cytokine processing by matrix metalloproteinases and its effect on leukocyte migration and inflammation. J. Leukoc. Biol. 2007, 82, 1375–1381. [Google Scholar] [CrossRef]
- Schönbeck, U.; Mach, F.; Libby, P. Generation of biologically active IL-1 beta by matrix metalloproteinases: A novel caspase-1-independent pathway of IL-1 beta processing. J. Immunol. 1998, 161, 3340–3346. [Google Scholar] [CrossRef]
- Munaut, C.; Noël, A.; Hougrand, O.; Foidart, J.M.; Boniver, J.; Deprez, M. Vascular endothelial growth factor expression correlates with matrix metalloproteinases MT1-MMP, MMP-2 and MMP-9 in human glioblastomas. Int. J. Cancer 2003, 106, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Juttermann, R.; Soloway, P.D. TIMP-2 is required for efficient activation of proMMP-2 in vivo. J. Biol. Chem. 2000, 275, 26411–26415. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, T.; McEver, R.P. The cytoplasmic domain of P-selectin is phosphorylated on serine and threonine residues. Blood 1993, 82, 1758–1766. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).