.png)
FastCellpose: A Fast and Accurate Deep-Learning Framework for Segmentation of All Glomeruli in Mouse Whole-Kidney Microscopic Optical Images


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Tissue Preparation
2.3. Cryo-fMOST for Whole-Kidney Imaging
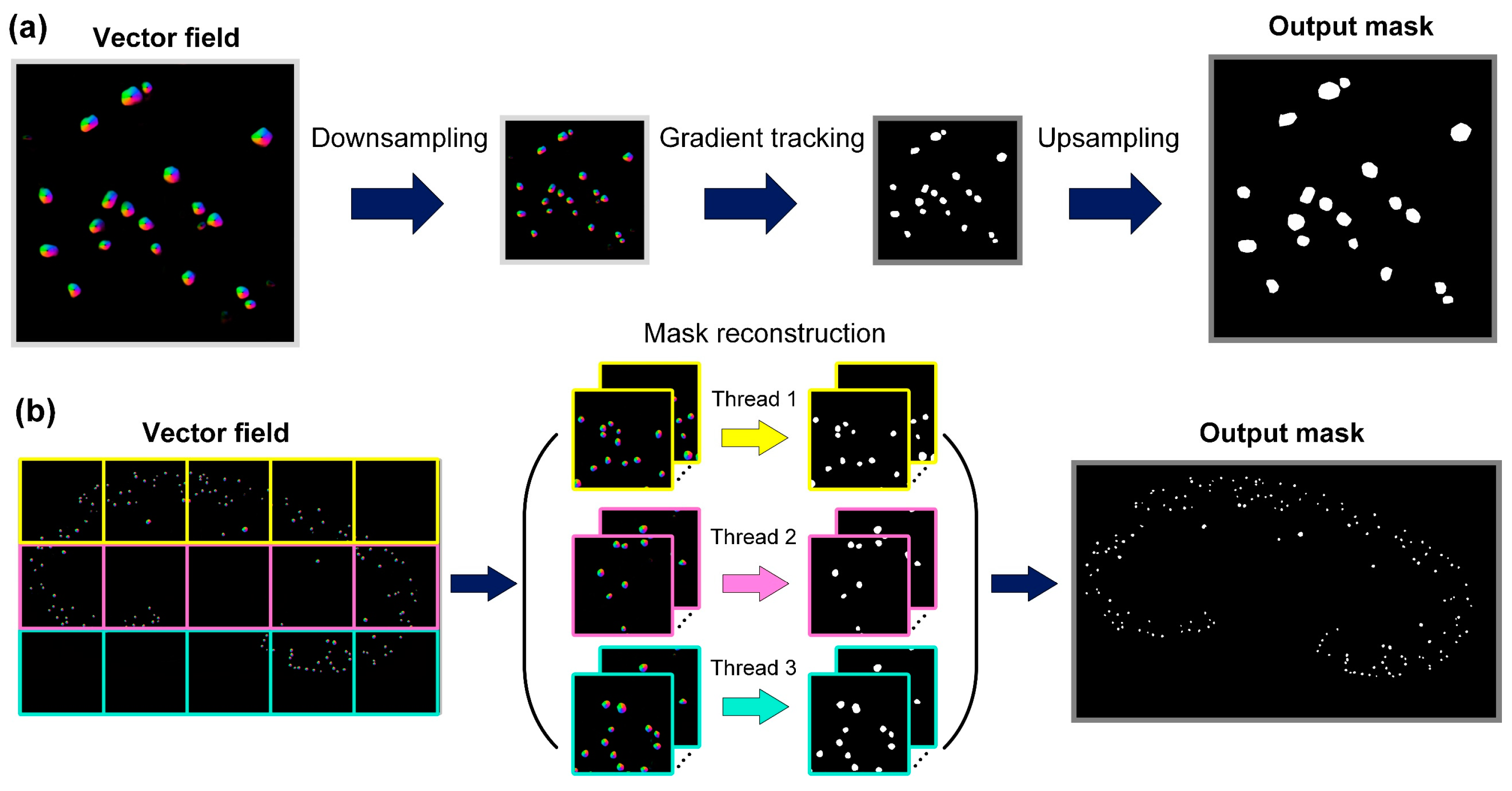
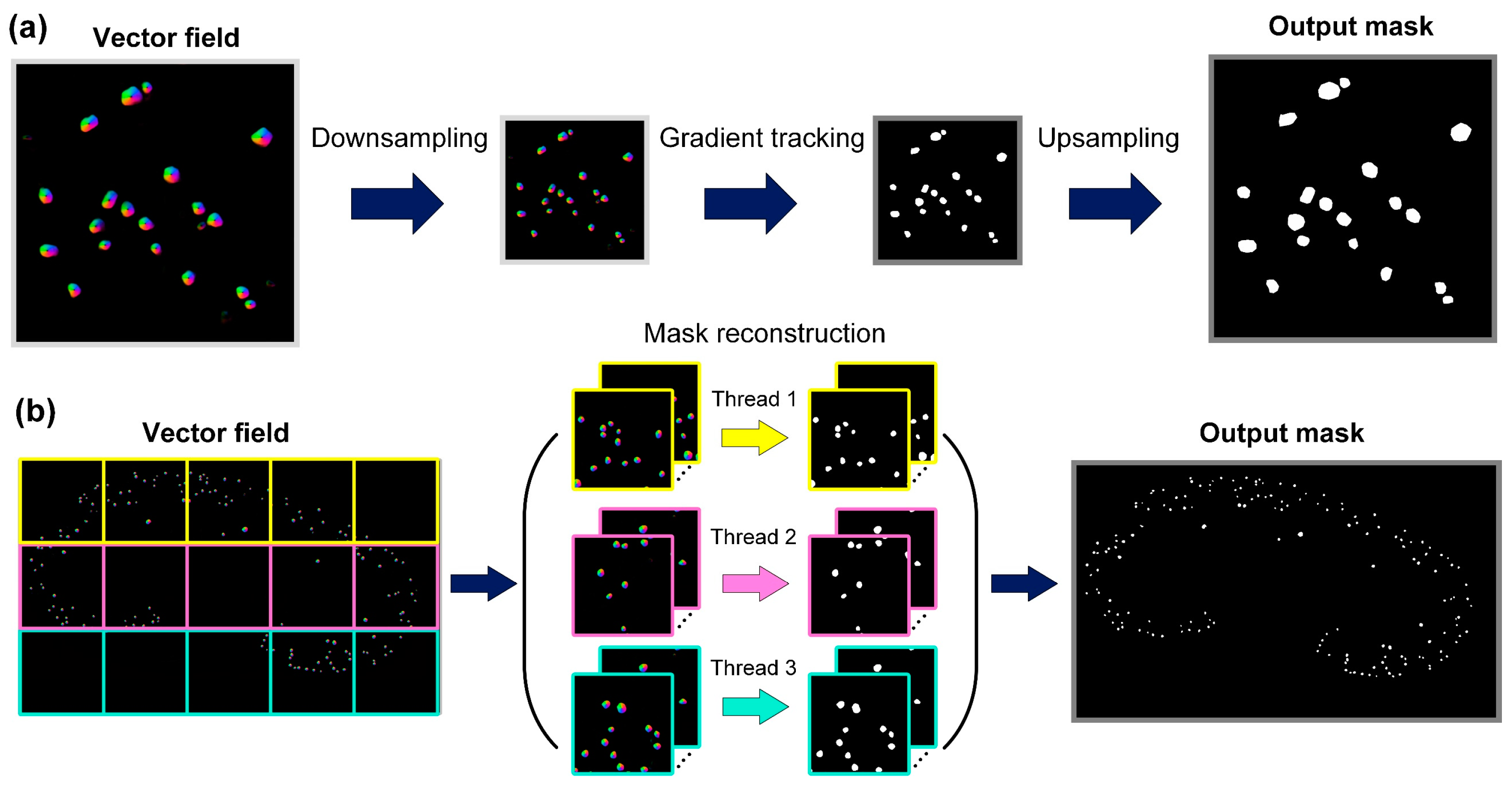
2.4. FastCellpose Framework
2.5. Training Data Preparation and Implementation Details
2.6. Performance Criteria
3. Results
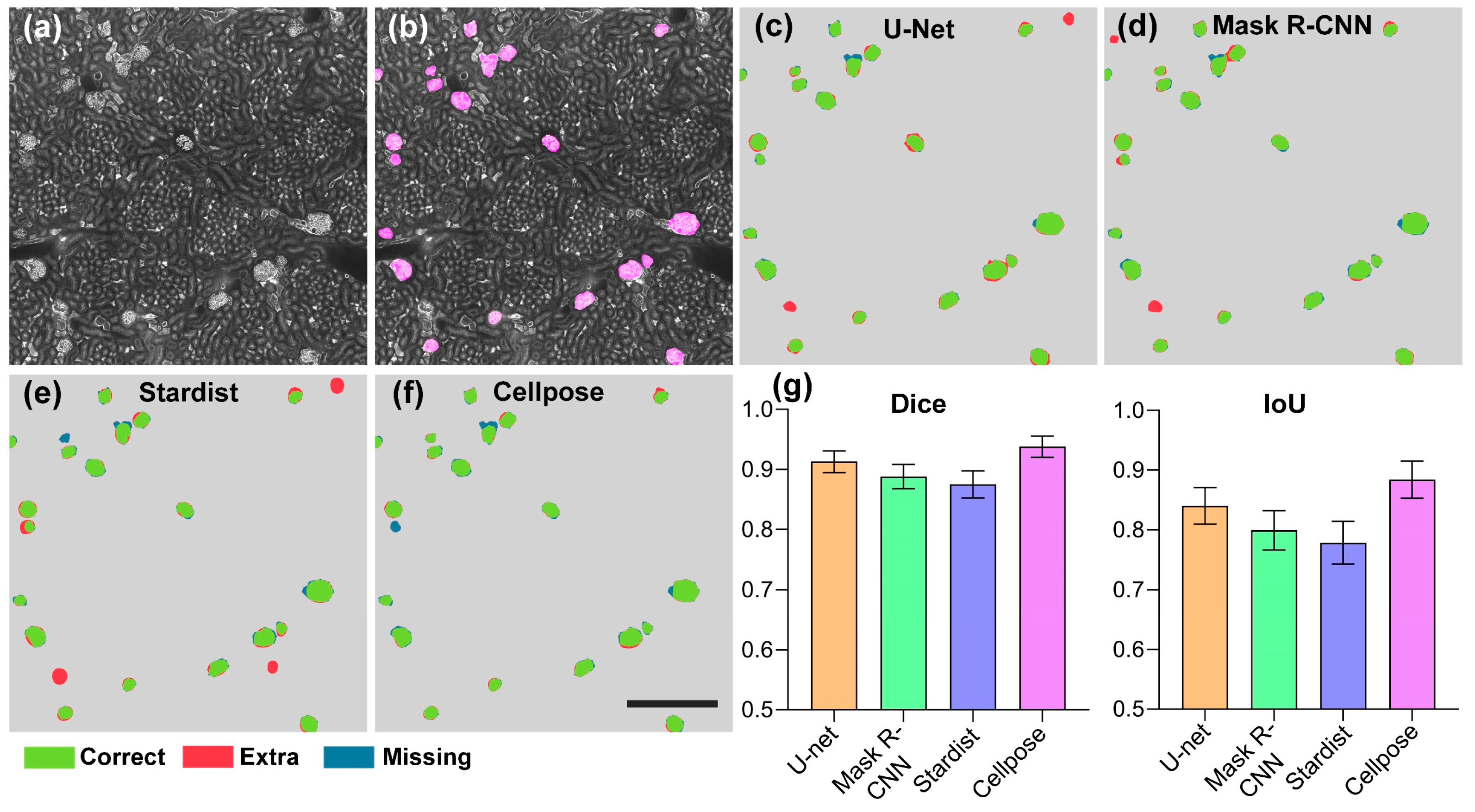
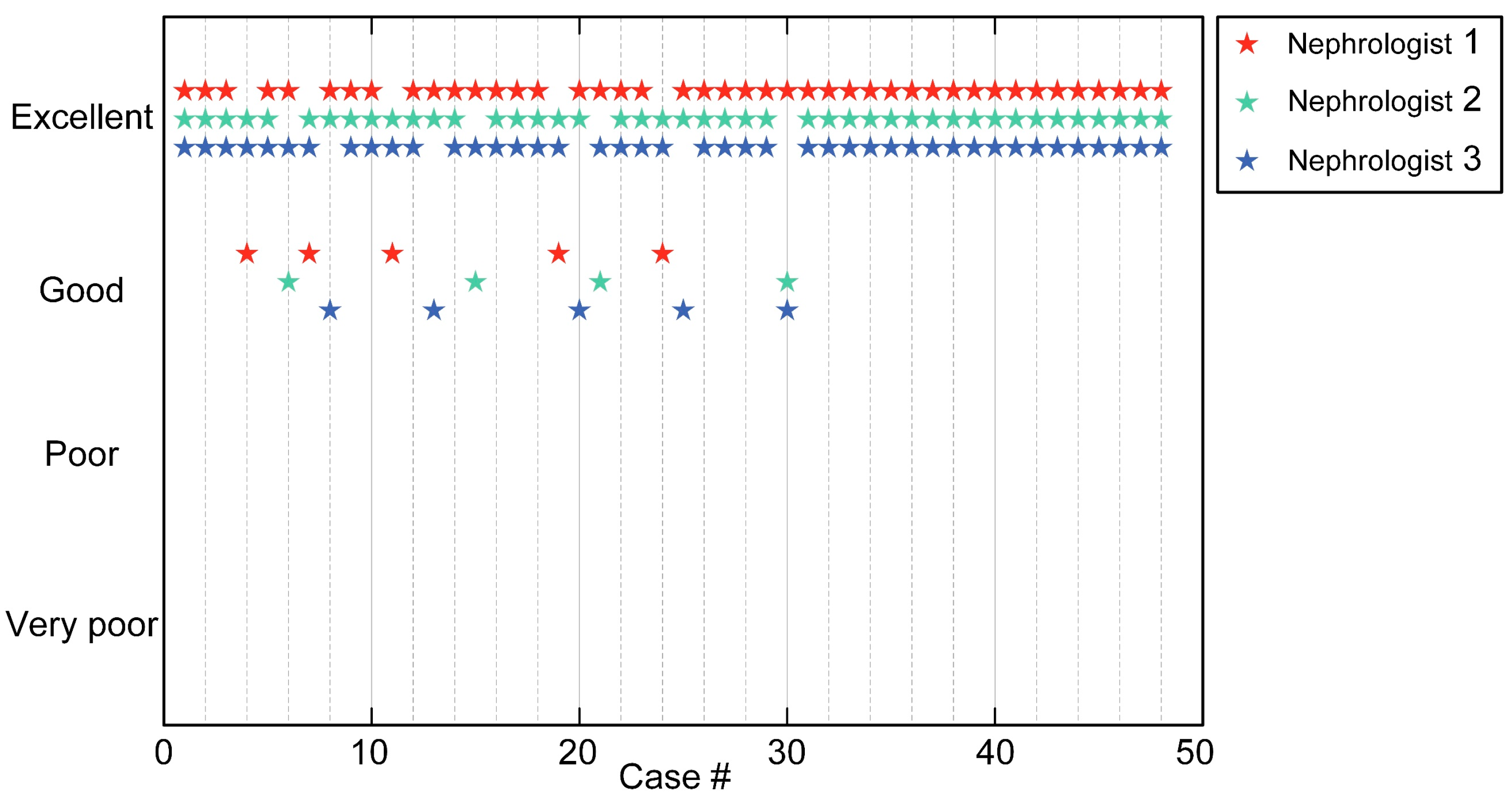
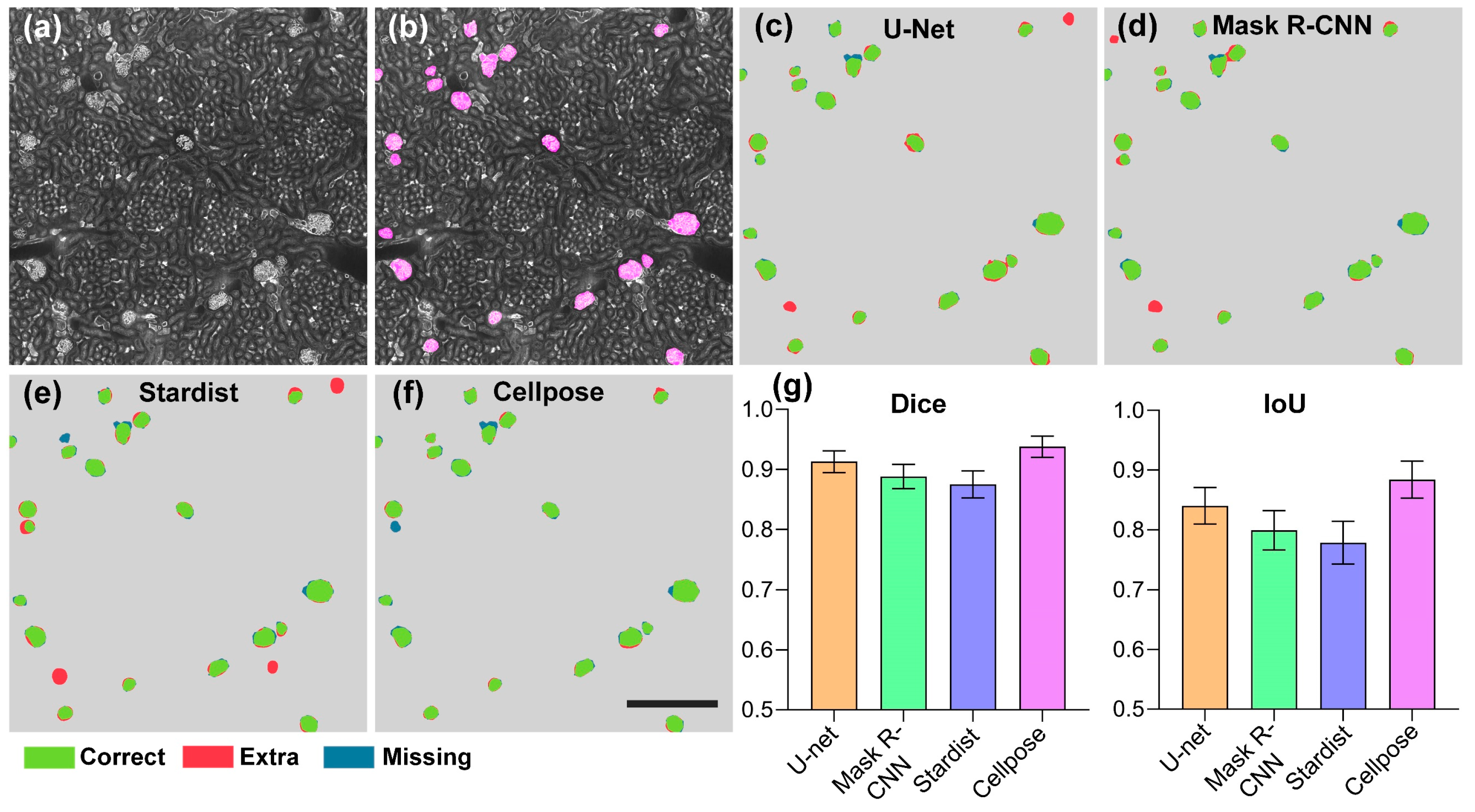
3.1. Evaluation of Segmentation Algorithms for Glomeruli Segmentation
3.2. Comprehensive Optimization of FastCellpose for Rapid Image Segmentation
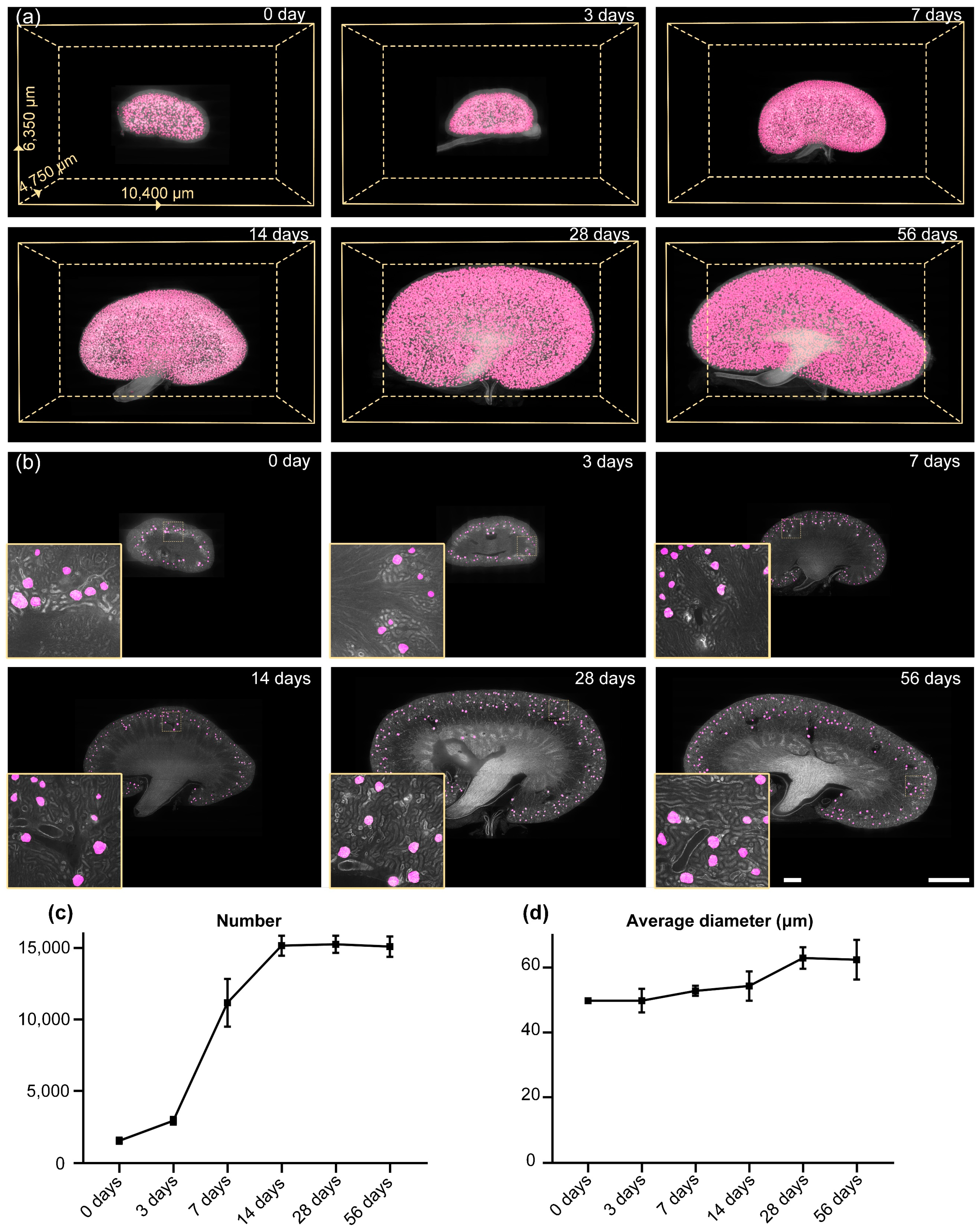
3.3. Quantitative Analysis of Glomerular Development
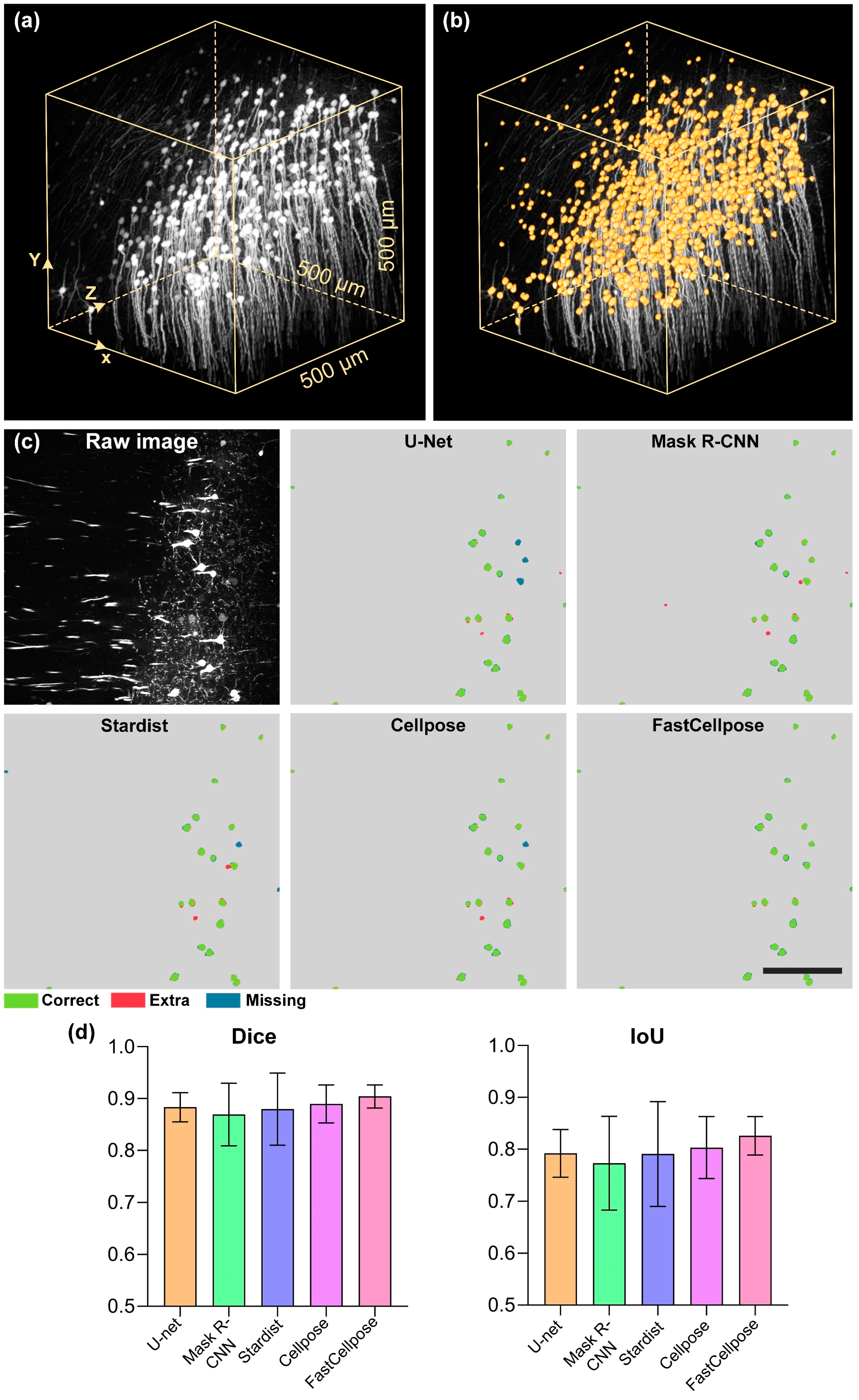
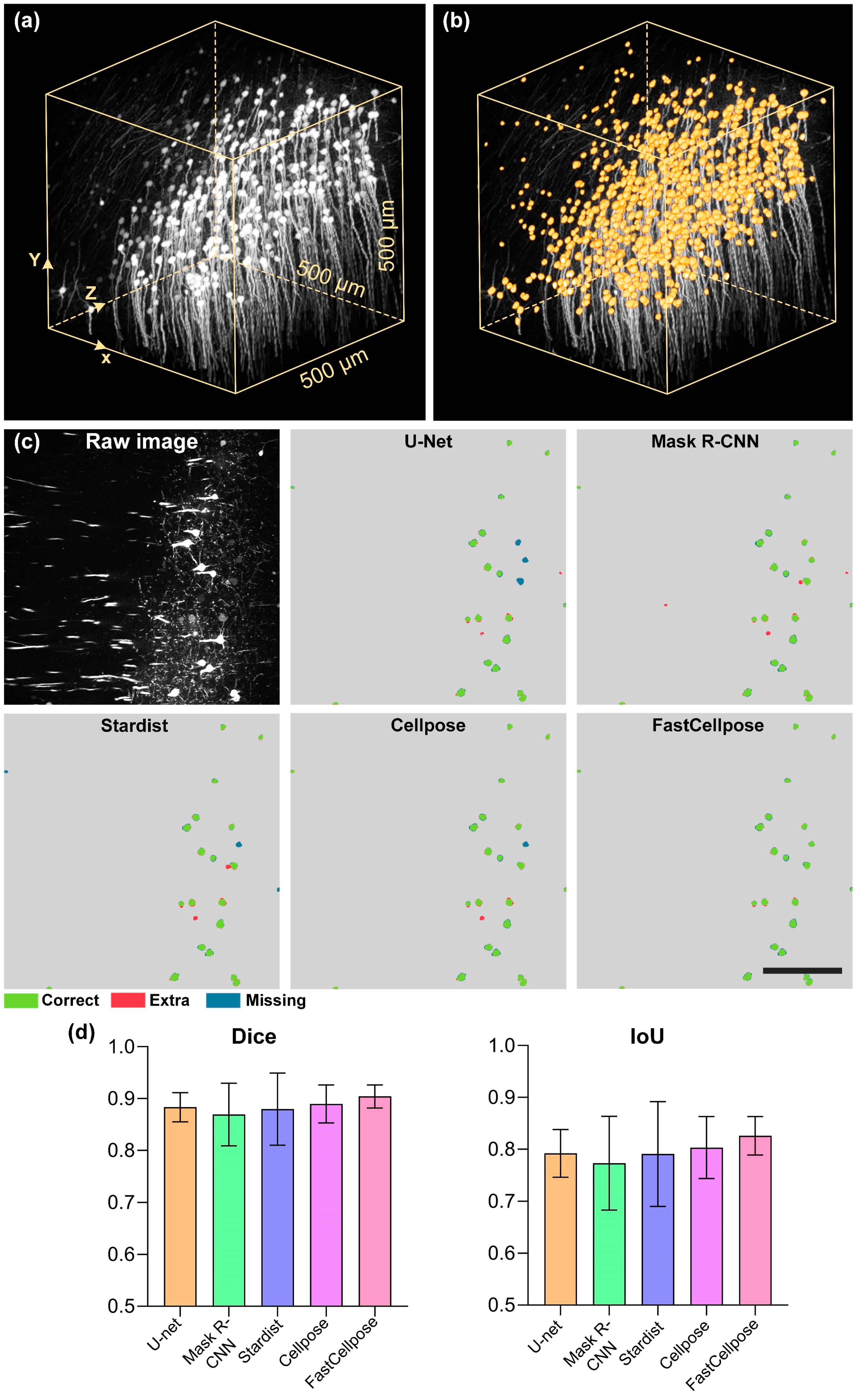
3.4. FastCellpose for Neuronal Soma Segmentation
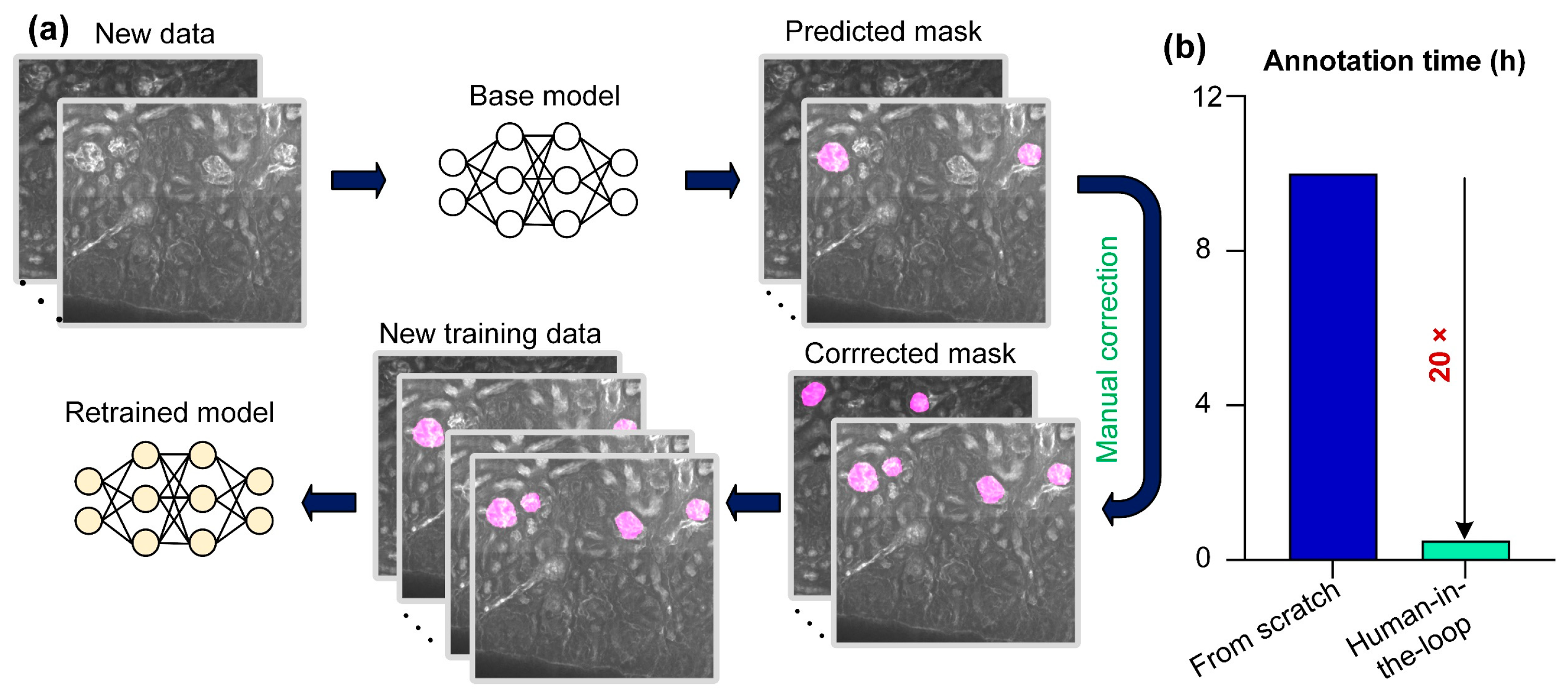
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pollak, M.R.; Quaggin, S.E.; Hoenig, M.P.; Dworkin, L.D. The glomerulus: The sphere of influence. Clin. J. Am. Soc. Nephrol. 2014, 9, 1461–1469. [Google Scholar] [CrossRef] [PubMed]
- Puelles, V.G.; Hoy, W.E.; Hughson, M.D.; Diouf, B.; Douglas-Denton, R.N.; Bertram, J.F. Glomerular number and size variability and risk for kidney disease. Curr. Opin. Nephrol. Hypertens. 2011, 20, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Ruggenenti, P.; Cravedi, P.; Remuzzi, G. Mechanisms and treatment of CKD. J. Am. Soc. Nephrol. 2012, 23, 1917–1928. [Google Scholar] [CrossRef] [PubMed]
- Chaabane, W.; Praddaude, F.; Buleon, M.; Jaafar, A.; Vallet, M.; Rischmann, P.; Galarreta, C.I.; Chevalier, R.L.; Tack, I. Renal functional decline and glomerulotubular injury are arrested but not restored by release of unilateral ureteral obstruction (UUO). Am. J. Physiol.-Ren. Physiol. 2013, 304, F432–F439. [Google Scholar] [CrossRef]
- Tonneijck, L.; Muskiet, M.H.A.; Smits, M.M.; van Bommel, E.J.; Heerspink, H.J.L.; van Raalte, D.H.; Joles, J.A. Glomerular hyperfiltration in diabetes: Mechanisms, clinical significance, and treatment. J. Am. Soc. Nephrol. 2017, 28, 1023–1039. [Google Scholar] [CrossRef]
- Cullen-McEwen, L.A.; Armitage, J.A.; Nyengaard, J.R.; Moritz, K.M.; Bertram, J.F. A design-based method for estimating glomerular number in the developing kidney. Am. J. Physiol.-Ren. Physiol. 2011, 300, F1448–F1453. [Google Scholar] [CrossRef]
- Cullen-McEwen, L.A.; Kett, M.M.; Dowling, J.; Anderson, W.P.; Bertram, J.F. Nephron number, renal function, and arterial pressure in aged GDNF heterozygous mice. Hypertension 2003, 41, 335–340. [Google Scholar] [CrossRef]
- Murawski, I.J.; Maina, R.W.; Gupta, I.R. The relationship between nephron number, kidney size and body weight in two inbred mouse strains. Organogenesis 2010, 6, 189–194. [Google Scholar] [CrossRef]
- Baldelomar, E.J.; Charlton, J.R.; Beeman, S.C.; Hann, B.D.; Cullen-McEwen, L.; Pearl, V.M.; Bertram, J.F.; Wu, T.; Zhang, M.; Bennett, K.M. Phenotyping by magnetic resonance imaging nondestructively measures glomerular number and volume distribution in mice with and without nephron reduction. Kidney Int. 2016, 89, 498–505. [Google Scholar] [CrossRef]
- Richardson, D.S.; Lichtman, J.W. Clarifying tissue clearing. Cell 2015, 162, 246–257. [Google Scholar] [CrossRef]
- Ueda, H.R.; Ertürk, A.; Chung, K.; Gradinaru, V.; Chédotal, A.; Tomancak, P.; Keller, P.J. Tissue clearing and its applications in neuroscience. Nat. Rev. Neurosci. 2020, 21, 61–79. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, N.; Nicolas, N.; Roux, E. Computational identification and 3D morphological characterization of renal glomeruli in optically cleared murine kidneys. Sensors 2021, 21, 7440. [Google Scholar] [CrossRef] [PubMed]
- Ragan, T.; Kadiri, L.R.; Venkataraju, K.U.; Bahlmann, K.; Sutin, J.; Taranda, J.; Arganda-Carreras, I.; Kim, Y.; Seung, H.S.; Osten, P. Serial two-photon tomography for automated ex vivo mouse brain imaging. Nat. Methods 2012, 9, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zeng, S.; Yan, C.; Lv, X.; Yang, Z.; Xu, T.; Feng, Z.; Ding, W.; Qi, X.; Li, A.; et al. Continuously tracing brain-wide long-distance axonal projections in mice at a one-micron voxel resolution. NeuroImage 2013, 74, 87–98. [Google Scholar] [CrossRef]
- Economo, M.N.; Clack, N.G.; Lavis, L.D.; Gerfen, C.R.; Svoboda, K.; Myers, E.W.; Chandrashekar, J. A platform for brain-wide imaging and reconstruction of individual neurons. eLife 2016, 5, e10566. [Google Scholar] [CrossRef]
- Zhong, Q.; Li, A.; Jin, R.; Zhang, D.; Li, X.; Jia, X.; Ding, Z.; Luo, P.; Zhou, C.; Jiang, C.; et al. High-definition imaging using line-illumination modulation microscopy. Nat. Methods 2021, 18, 309–315. [Google Scholar] [CrossRef]
- Jiang, T.; Gong, H.; Yuan, J. Whole-brain optical imaging: A powerful tool for precise brain mapping at the mesoscopic level. Neurosci. Bull. 2023, 39, 1840–1858. [Google Scholar] [CrossRef]
- Deng, L.; Chen, J.; Li, Y.; Han, Y.; Fan, G.; Yang, J.; Cao, D.; Lu, B.; Ning, K.; Nie, S.; et al. Cryo-fluorescence micro-optical sectioning tomography for volumetric imaging of various whole organs with subcellular resolution. iScience 2022, 25, 104805. [Google Scholar] [CrossRef]
- Wu, H.S.; Berba, J.; Gil, J. Iterative thresholding for segmentation of cells from noisy images. J. Microsc. 2000, 197, 296–304. [Google Scholar] [CrossRef]
- Bolte, S.; Cordelieres, F.P. A guided tour into subcellular colocalization analysis in light microscopy. J. Microsc. 2006, 224, 213–232. [Google Scholar] [CrossRef]
- Carpenter, A.E.; Jones, T.R.; Lamprecht, M.R.; Clarke, C.; Kang, I.H.; Friman, O.; Guertin, D.A.; Chang, J.H.; Lindquist, R.A.; Moffat, J.; et al. CellProfiler: Image analysis software for identifying and quantifying cell phenotypes. Genome Biol. 2006, 7, R100. [Google Scholar] [CrossRef]
- Li, G.; Liu, T.; Nie, J.; Guo, L.; Chen, J.; Zhu, J.; Xia, W.; Mara, A.; Holley, S.; Wong, S.T.C. Segmentation of touching cell nuclei using gradient flow tracking. J. Microsc. 2008, 231, 47–58. [Google Scholar] [CrossRef]
- McQuin, C.; Goodman, A.; Chernyshev, V.; Kamentsky, L.; Cimini, B.A.; Karhohs, K.W.; Doan, M.; Ding, L.; Rafelski, S.M.; Thirstrup, D.; et al. CellProfiler 3.0: Next-generation image processing for biology. PLoS Biol. 2018, 16, e2005970. [Google Scholar] [CrossRef]
- Berg, S.; Kutra, D.; Kroeger, T.; Straehle, C.N.; Kausler, B.X.; Haubold, C.; Schiegg, M.; Ales, J.; Beier, T.; Rudy, M.; et al. ilastik: Interactive machine learning for (bio)image analysis. Nat. Methods 2019, 16, 1226–1232. [Google Scholar] [CrossRef]
- Moen, E.; Bannon, D.; Kudo, T.; Graf, W.; Covert, M.; Van Valen, D. Deep learning for cellular image analysis. Nat. Methods 2019, 16, 1233–1246. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, H.; Jin, L.; Chen, J.; Nedzved, A.; Ablameyko, S.; Ma, Q.; Yu, J.; Xu, Y. U-Net-based deep learning for tracking and quantitative analysis of intracellular vesicles in time-lapse microscopy images. J. Innov. Opt. Health Sci. 2022, 15, 2250031. [Google Scholar] [CrossRef]
- Wang, H.; Wang, Z.; Wang, J.; Li, K.; Geng, G.; Kang, F.; Cao, X. ICA-Unet: An improved U-net network for brown adipose tissue segmentation. J. Innov. Opt. Health Sci. 2022, 15, 2250018. [Google Scholar] [CrossRef]
- Yin, J.; Yang, G.; Qin, X.; Li, H.; Wang, L. Optimized U-Net model for 3D light-sheet image segmentation of zebrafish trunk vessels. Biomed. Opt. Express 2022, 13, 2896–2908. [Google Scholar] [CrossRef]
- Park, J.; Bae, S.; Seo, S.; Park, S.; Bang, J.-I.; Han, J.H.; Lee, W.W.; Lee, J.S. Measurement of glomerular filtration rate using quantitative SPECT/CT and deep-learning-based kidney segmentation. Sci. Rep. 2019, 9, 4223. [Google Scholar] [CrossRef]
- Klepaczko, A.; Strzelecki, M.; Kociołek, M.; Eikefjord, E.; Lundervold, A. A multi-layer perceptron network for perfusion parameter estimation in DCE-MRI studies of the healthy kidney. Appl. Sci. 2020, 10, 5525. [Google Scholar] [CrossRef]
- Silva, J.; Souza, L.; Chagas, P.; Calumby, R.; Souza, B.; Pontes, I.; Duarte, A.; Pinheiro, N.; Santos, W.; Oliveira, L. Boundary-aware glomerulus segmentation: Toward one-to-many stain generalization. Comput. Med. Imaging Graph. 2022, 100, 102104. [Google Scholar] [CrossRef]
- Saikia, F.N.; Iwahori, Y.; Suzuki, T.; Bhuyan, M.K.; Wang, A.; Kijsirikul, B. MLP-UNet: Glomerulus segmentation. IEEE Access 2023, 11, 53034–53047. [Google Scholar] [CrossRef]
- Singh Samant, S.; Chauhan, A.; Dn, J.; Singh, V. Glomerulus detection using segmentation neural networks. J. Digit. Imaging 2023, 36, 1633–1642. [Google Scholar] [CrossRef]
- Ronneberger, O.; Fischer, P.; Brox, T. U-net: Convolutional networks for biomedical image segmentation. In Proceedings of the Medical Image Computing and Computer-Assisted Intervention, Munich, Germany, 5–9 October 2015; pp. 234–241. [Google Scholar]
- He, K.; Gkioxari, G.; Dollár, P.; Girshick, R. Mask r-cnn. In Proceedings of the IEEE International Conference on Computer Vision, Venice, Italy, 22–29 October 2017; pp. 2961–2969. [Google Scholar]
- Schmidt, U.; Weigert, M.; Broaddus, C.; Myers, G. Cell detection with star-convex polygons. In Proceedings of the Medical Image Computing and Computer Assisted Intervention, Granada, Spain, 16–20 September 2018; pp. 265–273. [Google Scholar]
- Stringer, C.; Wang, T.; Michaelos, M.; Pachitariu, M. Cellpose: A generalist algorithm for cellular segmentation. Nat. Methods 2021, 18, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Pachitariu, M.; Stringer, C. Cellpose 2.0: How to train your own model. Nat. Methods 2022, 19, 1634–1641. [Google Scholar] [CrossRef]
- Kleinberg, G.; Wang, S.; Comellas, E.; Monaghan, J.R.; Shefelbine, S.J. Usability of deep learning pipelines for 3D nuclei identification with Stardist and Cellpose. Cells Dev. 2022, 172, 203806. [Google Scholar] [CrossRef]
- Ioffe, S.; Szegedy, C. Batch normalization: Accelerating deep network training by reducing internal covariate shift. arXiv 2015, arXiv:1502.03167. [Google Scholar]
- Zhang, C.; Yan, C.; Ren, M.; Li, A.; Quan, T.; Gong, H.; Yuan, J. A platform for stereological quantitative analysis of the brain-wide distribution of type-specific neurons. Sci. Rep. 2017, 7, 14334. [Google Scholar] [CrossRef]
- Zhao, J.; Chen, X.; Xiong, Z.; Liu, D.; Zeng, J.; Xie, C.; Zhang, Y.; Zha, Z.J.; Bi, G.; Wu, F. Neuronal population reconstruction from ultra-scale optical microscopy images via progressive learning. IEEE Trans. Med. Imaging 2020, 39, 4034–4046. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Segmentation Model | Architecture Parameters | Performance | ||||
|---|---|---|---|---|---|---|
| K | N | Trainable Parameters | Dice | IoU | Inference Time | |
| Model 1 | 32 | 4 | 6.60 × 106 | 0.938 ± 0.018 | 0.884 ± 0.031 | 1.50 |
| Model 2 | 32 | 2 | 3.06 × 106 | 0.942 ± 0.015 | 0.891 ± 0.026 | 0.98 |
| Model 3 | 16 | 4 | 1.62 × 106 | 0.936 ± 0.021 | 0.881 ± 0.037 | 0.91 |
| Model 4 | 32 | 1 | 1.49 × 106 | 0.926 ± 0.017 | 0.863 ± 0.030 | 0.84 |
| Model 5 | 16 | 2 | 0.56 × 106 | 0.947 ± 0.017 | 0.899 ± 0.031 | 0.68 |
| Model 6 | 16 | 1 | 0.37 × 106 | 0.922 ± 0.017 | 0.856 ± 0.029 | 0.57 |
| Downsampling Scale Factor | Segmentation Performance | Acceleration | |
|---|---|---|---|
| Dice | IoU | ||
| 1 | 0.947 ± 0.017 | 0.899 ± 0.031 | \ |
| 2 | 0.945 ± 0.016 | 0.894 ± 0.031 | 3.5-fold |
| 3 | 0.931 ± 0.015 | 0.871 ± 0.026 | 8.0-fold |
| 4 | 0.896 ± 0.016 | 0.812 ± 0.020 | 11.4-fold |
| Segmentation Methods | Trainable Parameters | Performance | |||
|---|---|---|---|---|---|
| Dice | IoU | Network Inference Time (s) | Mask Reconstruction Time (s) | ||
| U-Net | 2.96 × 106 | 0.913 ± 0.018 | 0.840 ± 0.031 | 0.88 | \ |
| Mask R-CNN | 4.39 × 107 | 0.888 ± 0.020 | 0.799 ± 0.033 | 3.45 | \ |
| Stardist | 1.43 × 106 | 0.875 ± 0.022 | 0.778 ± 0.036 | 0.81 | 4.05 |
| Cellpose | 6.60 × 106 | 0.938 ± 0.018 | 0.884 ± 0.031 | 1.50 | 4.68 |
| FastCellpose | 0.56 × 106 | 0.945 ± 0.016 | 0.894 ± 0.028 | 0.22 | 0.28 |
| Mouse Age (Days) | Number of Datasets | Number of Images per Dataset | Image Size |
|---|---|---|---|
| 0 | 3 | 736 | 8497 × 11,712 |
| 3 | 3 | 852 | 10,997 × 15,616 |
| 7 | 3 | 1020 | 19,993 × 11,712 |
| 14 | 3 | 1248 | 22,993 × 17,568 |
| 28 | 3 | 1527 | 31,993 × 17,568 |
| 56 | 3 | 1584 | 33,996 × 19,520 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Y.; Zhang, Z.; Li, Y.; Fan, G.; Liang, M.; Liu, Z.; Nie, S.; Ning, K.; Luo, Q.; Yuan, J. FastCellpose: A Fast and Accurate Deep-Learning Framework for Segmentation of All Glomeruli in Mouse Whole-Kidney Microscopic Optical Images. Cells 2023, 12, 2753. https://doi.org/10.3390/cells12232753
Han Y, Zhang Z, Li Y, Fan G, Liang M, Liu Z, Nie S, Ning K, Luo Q, Yuan J. FastCellpose: A Fast and Accurate Deep-Learning Framework for Segmentation of All Glomeruli in Mouse Whole-Kidney Microscopic Optical Images. Cells. 2023; 12(23):2753. https://doi.org/10.3390/cells12232753
Chicago/Turabian StyleHan, Yutong, Zhan Zhang, Yafeng Li, Guoqing Fan, Mengfei Liang, Zhijie Liu, Shuo Nie, Kefu Ning, Qingming Luo, and Jing Yuan. 2023. "FastCellpose: A Fast and Accurate Deep-Learning Framework for Segmentation of All Glomeruli in Mouse Whole-Kidney Microscopic Optical Images" Cells 12, no. 23: 2753. https://doi.org/10.3390/cells12232753
APA StyleHan, Y., Zhang, Z., Li, Y., Fan, G., Liang, M., Liu, Z., Nie, S., Ning, K., Luo, Q., & Yuan, J. (2023). FastCellpose: A Fast and Accurate Deep-Learning Framework for Segmentation of All Glomeruli in Mouse Whole-Kidney Microscopic Optical Images. Cells, 12(23), 2753. https://doi.org/10.3390/cells12232753