
Life-Cycle-Dependent Toxicities of Mono- and Bifunctional Alkylating Agents in the 3R-Compliant Model Organism C. elegans


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. C. elegans Maintenance
2.2. Treatment with Mustards
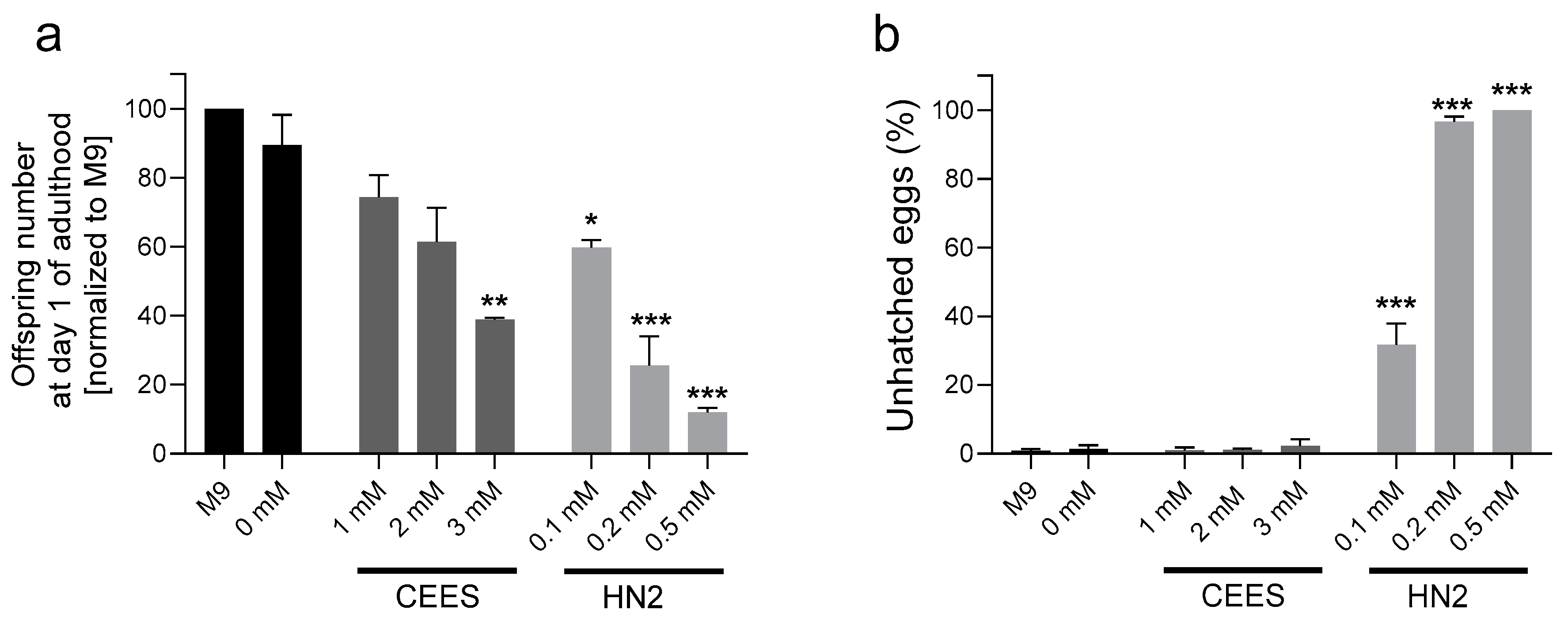
2.3. Survival and Reproduction
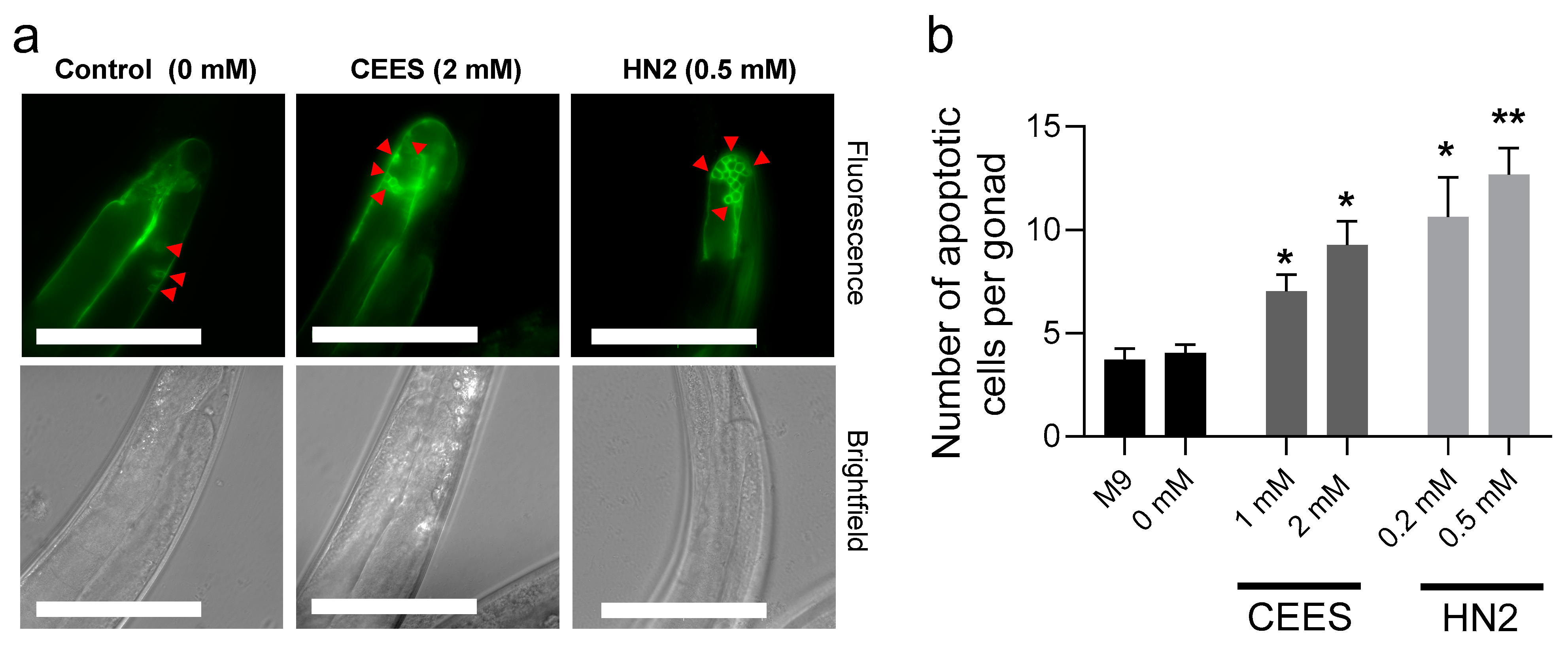
2.4. Germline Apoptosis
2.5. Neurotoxicity
2.6. NAD+ Levels
2.7. Statistical Analysis
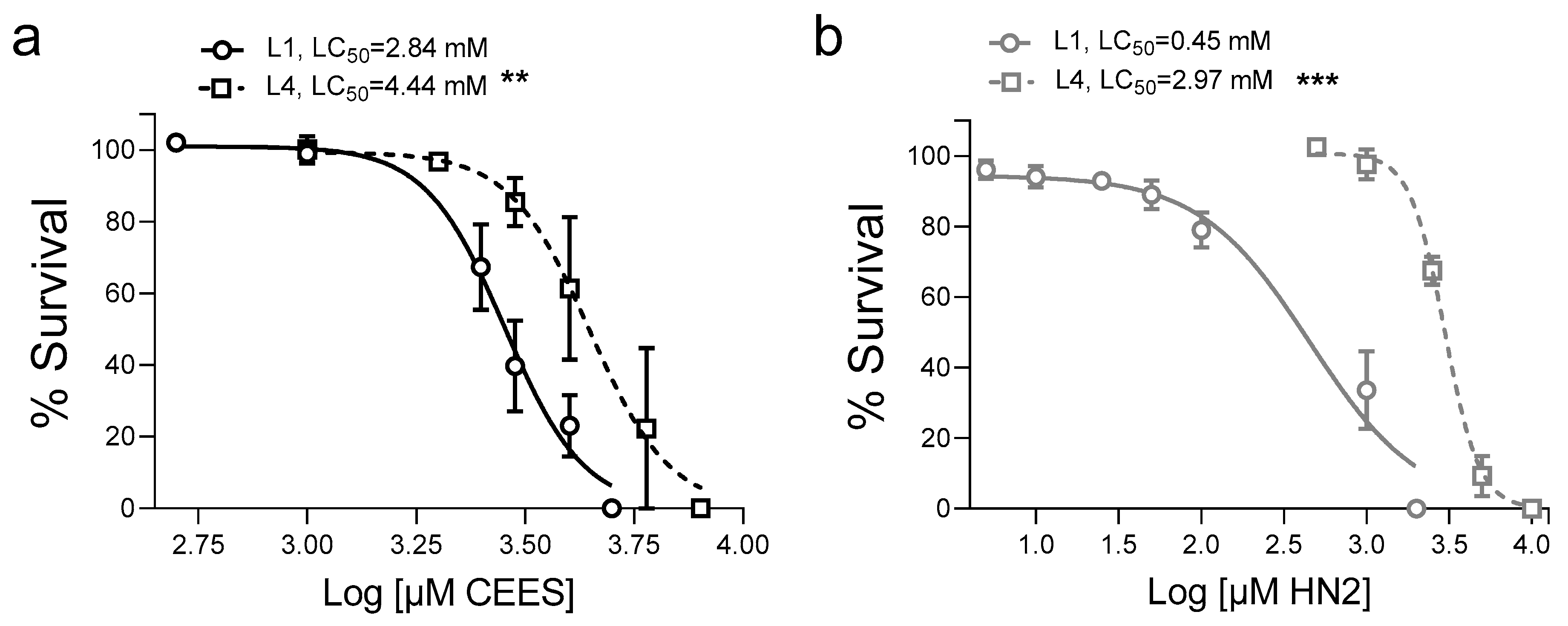
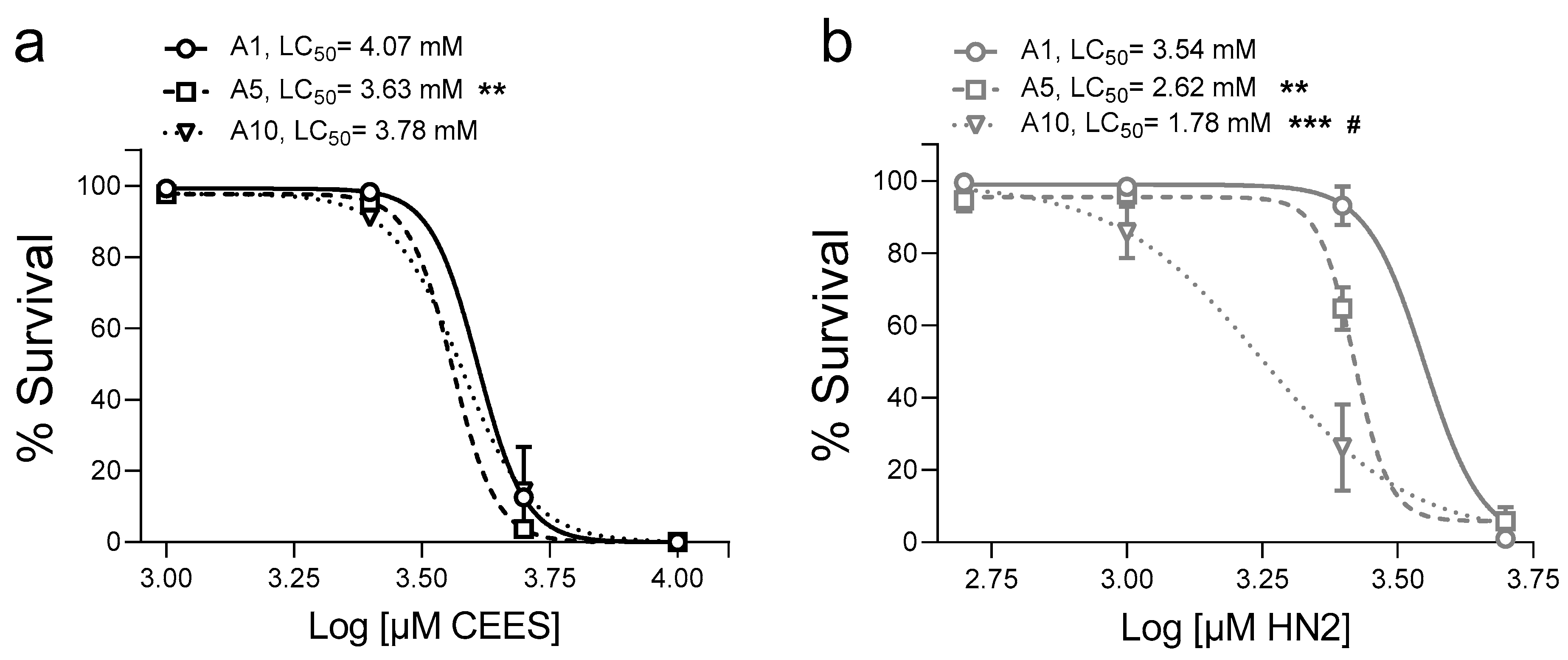
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mack, H.I.D.; Heimbucher, T.; Murphy, C.T. The nematode Caenorhabditis elegans as a model for aging research. Drug Discov. Today Dis. Models 2018, 27, 3–13. [Google Scholar] [CrossRef]
- Corsi, A.K.; Wightman, B.; Chalfie, M. A Transparent Window into Biology: A Primer on Caenorhabditis elegans. Genetics 2015, 200, 387–407. [Google Scholar] [CrossRef]
- Honnen, S. Caenorhabditis elegans as a powerful alternative model organism to promote research in genetic toxicology and biomedicine. Arch. Toxicol. 2017, 91, 2029–2044. [Google Scholar] [CrossRef]
- Hunt, P.R. The C. elegans model in toxicity testing. J. Appl. Toxicol. 2017, 37, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Tejeda-Benitez, L.; Olivero-Verbel, J. Caenorhabditis elegans, a Biological Model for Research in Toxicology. Rev. Environ. Contam. Toxicol. 2016, 237, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Long, N.P.; Kang, J.S.; Kim, H.M. Caenorhabditis elegans: A model organism in the toxicity assessment of environmental pollutants. Environ. Sci. Pollut. Res. Int. 2023, 30, 39273–39287. [Google Scholar] [CrossRef] [PubMed]
- Ruszkiewicz, J.A.; Pinkas, A.; Miah, M.R.; Weitz, R.L.; Lawes, M.J.A.; Akinyemi, A.J.; Ijomone, O.M.; Aschner, M.C. elegans as a model in developmental neurotoxicology. Toxicol. Appl. Pharmacol. 2018, 354, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Mangerich, A.; Esser, C. Chemical warfare in the First World War: Reflections 100 years later. Arch. Toxicol. 2014, 88, 1909–1911. [Google Scholar] [CrossRef] [PubMed]
- Panahi, Y.; Abdolghaffari, A.H.; Sahebkar, A. A review on symptoms, treatments protocols, and proteomic profile in sulfur mustard-exposed victims. J. Cell. Biochem. 2018, 119, 197–206. [Google Scholar] [CrossRef]
- Etemad, L.; Moshiri, M.; Balali-Mood, M. Advances in treatment of acute sulfur mustard poisoning—A critical review. Crit. Rev. Toxicol. 2019, 49, 191–214. [Google Scholar] [CrossRef] [PubMed]
- Neidle, S.; Thurston, D.E. Chemical approaches to the discovery and development of cancer therapies. Nat. Rev. Cancer 2005, 5, 285–296. [Google Scholar] [CrossRef]
- Singh, R.K.; Kumar, S.; Prasad, D.N.; Bhardwaj, T.R. Therapeutic journery of nitrogen mustard as alkylating anticancer agents: Historic to future perspectives. Eur. J. Med. Chem. 2018, 151, 401–433. [Google Scholar] [CrossRef]
- Ludlum, D.B.; Austin-Ritchie, P.; Hagopian, M.; Niu, T.Q.; Yu, D. Detection of sulfur mustard-induced DNA modifications. Chem. Biol. Interact. 1994, 91, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Kehe, K.; Balszuweit, F.; Steinritz, D.; Thiermann, H. Molecular toxicology of sulfur mustard-induced cutaneous inflammation and blistering. Toxicology 2009, 263, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Povirk, L.F.; Shuker, D.E. DNA damage and mutagenesis induced by nitrogen mustards. Mutat. Res. /Rev. Genet. Toxicol. 1994, 318, 205–226. [Google Scholar] [CrossRef] [PubMed]
- Zubel, T.; Hochgesand, S.; John, H.; Steinritz, D.; Schmidt, A.; Burkle, A.; Mangerich, A. A mass spectrometric platform for the quantitation of sulfur mustard-induced nucleic acid adducts as mechanistically relevant biomarkers of exposure. Arch. Toxicol. 2019, 93, 61–79. [Google Scholar] [CrossRef] [PubMed]
- Dębiak, M.; Panas, A.; Steinritz, D.; Kehe, K.; Bürkle, A. High-throughput analysis of DNA interstrand crosslinks in human peripheral blood mononuclear cells by automated reverse FADU assay. Toxicology 2011, 280, 53–60. [Google Scholar] [CrossRef]
- Debiak, M.; Lex, K.; Ponath, V.; Burckhardt-Boer, W.; Thiermann, H.; Steinritz, D.; Schmidt, A.; Mangerich, A.; Bürkle, A. Immunochemical analysis of poly(ADP-ribosyl)ation in HaCaT keratinocytes induced by the mono-alkylating agent 2-chloroethyl ethyl sulfide (CEES): Impact of experimental conditions. Toxicol. Lett. 2016, 244, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Covarrubias, A.J.; Perrone, R.; Grozio, A.; Verdin, E. NAD(+) metabolism and its roles in cellular processes during ageing. Nat. Rev. Mol. Cell Biol. 2021, 22, 119–141. [Google Scholar] [CrossRef]
- Lüscher, B.; Ahel, I.; Altmeyer, M.; Ashworth, A.; Bai, P.; Chang, P.; Cohen, M.; Corda, D.; Dantzer, F.; Daugherty, M.D.; et al. ADP-ribosyltransferases, an update on function and nomenclature. FEBS J. 2022, 289, 7399–7410. [Google Scholar] [CrossRef]
- Mol, M.E.; de Vries, R.; Kluivers, A.W. Effects of nicotinamide on biochemical changes and microblistering induced by sulfur mustard in human skin organ cultures. Toxicol. Appl. Pharmacol. 1991, 107, 439–449. [Google Scholar] [CrossRef]
- Yourick, J.J.; Dawson, J.S.; Mitcheltree, L.W. Sulfur mustard-induced microvesication in hairless guinea pigs: Effect of short-term niacinamide administration. Toxicol. Appl. Pharmacol. 1992, 117, 104–109. [Google Scholar] [CrossRef]
- Yourick, J.J.; Dawson, J.S.; Benton, C.D.; Craig, M.E.; Mitcheltree, L.W. Pathogenesis of 2,2′-dichlorodiethyl sulfide in hairless guinea pigs. Toxicology 1993, 84, 185–197. [Google Scholar] [CrossRef]
- Mol, M.A.; van de Ruit, A.M.; Kluivers, A.W. NAD+ levels and glucose uptake of cultured human epidermal cells exposed to sulfur mustard. Toxicol. Appl. Pharmacol. 1989, 98, 159–165. [Google Scholar] [CrossRef]
- Smith, W.J.; Gross, C.L.; Chan, P.; Meier, H.L. The use of human epidermal keratinocytes in culture as a model for studying the biochemical mechanisms of sulfur mustard toxicity. Cell Biol. Toxicol. 1990, 6, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Ruszkiewicz, J.; Papatheodorou, Y.; Jäck, N.; Melzig, J.; Eble, F.; Pirker, A.; Thomann, M.; Haberer, A.; Rothmiller, S.; Bürkle, A.; et al. NAD+ Acts as a Protective Factor in Cellular Stress Response to DNA Alkylating Agents. Cells 2023, 12, 2396. [Google Scholar] [CrossRef]
- Papirmeister, B.; Gross, C.L.; Meier, H.L.; Petrali, J.P.; Johnson, J.B. Molecular Basis for Mustard-Induced Vesication12. Toxicol. Sci. 1985, 5, 134–149. [Google Scholar] [CrossRef]
- Ruszkiewicz, J.A.; Burkle, A.; Mangerich, A. NAD(+) in sulfur mustard toxicity. Toxicol. Lett. 2020, 324, 95–103. [Google Scholar] [CrossRef]
- Ray Chaudhuri, A.; Nussenzweig, A. The multifaceted roles of PARP1 in DNA repair and chromatin remodelling. Nat. Rev. Mol. Cell Biol. 2017, 18, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Cortes, U.; Tong, W.M.; Coyle, D.L.; Meyer-Ficca, M.L.; Meyer, R.G.; Petrilli, V.; Herceg, Z.; Jacobson, E.L.; Jacobson, M.K.; Wang, Z.Q. Depletion of the 110-kilodalton isoform of poly(ADP-ribose) glycohydrolase increases sensitivity to genotoxic and endotoxic stress in mice. Mol. Cell. Biol. 2004, 24, 7163–7178. [Google Scholar] [CrossRef]
- St-Laurent, J.F.; Desnoyers, S. Poly(ADP-ribose) metabolism analysis in the nematode Caenorhabditis elegans. Methods Mol. Biol. 2011, 780, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Jeong, H.; Park, K.H.; Kim, K.W. Effects of NAD(+) in Caenorhabditis elegans Models of Neuronal Damage. Biomolecules 2020, 10, 993. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, S.; Santelli, J.; Mitchell, D.H.; Stiles, J.W.; Sanadi, D.R. A simple method for maintaining large, aging populations of Caenorhabditis elegans. Mech. Ageing Dev. 1980, 12, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Jacques, M.T.; Bornhorst, J.; Soares, M.V.; Schwerdtle, T.; Garcia, S.; Ávila, D.S. Reprotoxicity of glyphosate-based formulation in Caenorhabditis elegans is not due to the active ingredient only. Environ Pollut. 2019, 252, 1854–1862. [Google Scholar] [CrossRef] [PubMed]
- de la Guardia, Y.; Gilliat, A.F.; Hellberg, J.; Rennert, P.; Cabreiro, F.; Gems, D. Run-on of germline apoptosis promotes gonad senescence in C. elegans. Oncotarget 2016, 7, 39082–39096. [Google Scholar] [CrossRef]
- Zhou, Z.; Hartwieg, E.; Horvitz, H.R. CED-1 is a transmembrane receptor that mediates cell corpse engulfment in C. elegans. Cell 2001, 104, 43–56. [Google Scholar] [CrossRef]
- Ijomone, O.M.; Gubert, P.; Okoh, C.O.A.; Varão, A.M.; Amara, L.O.; Aluko, O.M.; Aschner, M. Application of Fluorescence Microscopy and Behavioral Assays to Demonstrating Neuronal Connectomes and Neurotransmitter Systems in C. elegans. Neuromethods 2021, 172, 399–426. [Google Scholar]
- Ijomone, O.M.; Miah, M.R.; Peres, T.V.; Nwoha, P.U.; Aschner, M. Null allele mutants of trt-1, the catalytic subunit of telomerase in Caenorhabditis elegans, are less sensitive to Mn-induced toxicity and DAergic degeneration. NeuroToxicology 2016, 57, 54–60. [Google Scholar] [CrossRef]
- Nass, R.; Hall, D.H.; Miller, D.M., 3rd; Blakely, R.D. Neurotoxin-induced degeneration of dopamine neurons in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2002, 99, 3264–3269. [Google Scholar] [CrossRef]
- Doitsidou, M.; Flames, N.; Lee, A.C.; Boyanov, A.; Hobert, O. Automated screening for mutants affecting dopaminergic-neuron specification in C. elegans. Nat. Methods 2008, 5, 869–872. [Google Scholar] [CrossRef]
- Jacobson, E.L.; Jacobson, M.K. Pyridine nucleotide levels as a function of growth in normal and transformed 3T3 cells. Arch. Biochem. Biophys. 1976, 175, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Mangerich, A.; Debiak, M.; Birtel, M.; Ponath, V.; Balszuweit, F.; Lex, K.; Martello, R.; Burckhardt-Boer, W.; Strobelt, R.; Siegert, M.; et al. Sulfur and nitrogen mustards induce characteristic poly(ADP-ribosyl)ation responses in HaCaT keratinocytes with distinctive cellular consequences. Toxicol. Lett. 2016, 244, 56–71. [Google Scholar] [CrossRef][Green Version]
- Wang, Q.Q.; Begum, R.A.; Day, V.W.; Bowman-James, K. Sulfur, oxygen, and nitrogen mustards: Stability and reactivity. Org. Biomol. Chem. 2012, 10, 8786–8793. [Google Scholar] [CrossRef]
- Bailly, A.P.; Freeman, A.; Hall, J.; Déclais, A.C.; Alpi, A.; Lilley, D.M.; Ahmed, S.; Gartner, A. The Caenorhabditis elegans homolog of Gen1/Yen1 resolvases links DNA damage signaling to DNA double-strand break repair. PLoS Genet. 2010, 6, e1001025. [Google Scholar] [CrossRef]
- Hartman, J.H.; Widmayer, S.J.; Bergemann, C.M.; King, D.E.; Morton, K.S.; Romersi, R.F.; Jameson, L.E.; Leung, M.C.K.; Andersen, E.C.; Taubert, S.; et al. Xenobiotic metabolism and transport in Caenorhabditis elegans. J. Toxicol. Environ. Health B Crit. Rev. 2021, 24, 51–94. [Google Scholar] [CrossRef]
- Meier, B.; Cooke, S.L.; Weiss, J.; Bailly, A.P.; Alexandrov, L.B.; Marshall, J.; Raine, K.; Maddison, M.; Anderson, E.; Stratton, M.R.; et al. C. elegans whole-genome sequencing reveals mutational signatures related to carcinogens and DNA repair deficiency. Genome Res. 2014, 24, 1624–1636. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.T.; Mohideen, F.; Meyer, K.; Harper, J.W.; Colaiácovo, M.P. SLX-1 is required for maintaining genomic integrity and promoting meiotic noncrossovers in the Caenorhabditis elegans germline. PLoS Genet. 2012, 8, e1002888. [Google Scholar] [CrossRef]
- Bu, M.; Farrer, M.J.; Khoshbouei, H. Dynamic control of the dopamine transporter in neurotransmission and homeostasis. NPJ Park. Dis. 2021, 7, 22. [Google Scholar] [CrossRef]
- Jiang, N.; Zhang, P.; Shen, W.; Zhang, Y.; Zhou, W. Clinical and experimental research progress on neurotoxicity of sulfur mustard and its possible mechanisms. Toxicology 2023, 483, 153372. [Google Scholar] [CrossRef] [PubMed]
- Darchini-Maragheh, E.; Nemati-Karimooy, H.; Hasanabadi, H.; Balali-Mood, M. Delayed neurological complications of sulphur mustard and tabun poisoning in 43 Iranian veterans. Basic Clin. Pharmacol. Toxicol. 2012, 111, 426–432. [Google Scholar] [CrossRef]
- Balali-Mood, M.; Hefazi, M.; Mahmoudi, M.; Jalali, E.; Attaran, D.; Maleki, M.; Razavi, M.E.; Zare, G.; Tabatabaee, A.; Jaafari, M.R. Long-term complications of sulphur mustard poisoning in severely intoxicated Iranian veterans. Fundam. Clin. Pharmacol. 2005, 19, 713–721. [Google Scholar] [CrossRef]
- Shoeibi, N.; Mousavi, M.N.; Balali-Mood, M.; Moshiri, M.; Darchini-Maragheh, E.; Mousavi, S.R.; Abrishami, M. Long-term complications of sulfur mustard poisoning: Retinal electrophysiological assessment in 40 severely intoxicated Iranian veterans. Int. J. Retin. Vitr. 2017, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, A.B.; Eriksen, J.; Smidt-Nielsen, K. Chronic neuropathic symptoms after exposure to mustard gas: A long-term investigation. J. Am. Acad. Dermatol. 1998, 39, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Roshan, R.; Rahnama, P.; Ghazanfari, Z.; Montazeri, A.; Soroush, M.R.; Naghizadeh, M.M.; Melyani, M.; Tavoli, A.; Ghazanfari, T. Long-term effects of sulfur mustard on civilians’ mental health 20 years after exposure (The Sardasht-Iran Cohort Study). Health Qual. Life Outcomes 2013, 11, 69. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Hernandez, A.; Roney, A.; Goswami, D.G.; Tewari-Singh, N.; Brown, J.M. A review of chemical warfare agents linked to respiratory and neurological effects experienced in Gulf War Illness. Inhal. Toxicol. 2022, 34, 412–432. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, T.W.; Wang, Y.; Song, Y.; Villanueva, M.; Jimenez, A. Sulphur mustard induces progressive toxicity and demyelination in brain cell aggregate culture. Neurotoxicology 2021, 84, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Kisby, G.E.; Springer, N.; Spencer, P.S. In vitro neurotoxic and DNA-damaging properties of nitrogen mustard. J. Appl. Toxicol. 2000, 20 (Suppl. S1), S35–S41. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.R.; Sunkaria, A.; Bal, A.; Bhutia, Y.D.; Vijayaraghavan, R.; Flora, S.J.S.; Gill, K.D. Neurobehavioral impairments, generation of oxidative stress and release of pro-apoptotic factors after chronic exposure to sulphur mustard in mouse brain. Toxicol. Appl. Pharmacol. 2009, 240, 208–218. [Google Scholar] [CrossRef]
- Tekiner, A.; Yucel, O.; Sargin, A.K.; Genc, O.; Can, B.; Karayilanoglu, T.; Karakaya, J.; Bayar, M.A. The effect of nitrogen mustard on the rat brain and the therapeutic value of proanthocyanidin. Turk. Neurosurg. 2009, 19, 360–366. [Google Scholar]
- Tekiner, A.; Yucel, D.; Bayar, M.A.; Yucel, O.; Erdem, Y.; Karakaya, J. The effect of Nitrogen Mustard on the enzymatic antioxidant defense of rat brain tissue and the therapeutic value of proanthocyanidin. Turk. Neurosurg. 2011, 21, 461–466. [Google Scholar] [CrossRef]
- Jafari, M. Dose- and time-dependent effects of sulfur mustard on antioxidant system in liver and brain of rat. Toxicology 2007, 231, 30–39. [Google Scholar] [CrossRef]
- Gilardoni, M.; Léonço, D.; Caffin, F.; Gros-Désormeaux, F.; Eldin, C.; Béal, D.; Ouzia, S.; Junot, C.; Fenaille, F.; Piérard, C.; et al. Evidence for the systemic diffusion of (2-chloroethyl)-ethyl-sulfide, a sulfur mustard analog, and its deleterious effects in brain. Toxicology 2021, 462, 152950. [Google Scholar] [CrossRef]
- Gadsden-Gray, J.; Mukherjee, S.; Ogunkua, O.; Das, S.K. Induction of neuronal damage in guinea pig brain by intratracheal infusion of 2-chloroethyl ethyl sulfide, a mustard gas analog. J. Biochem. Mol. Toxicol. 2012, 26, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Spanoudakis, E.; Tavernarakis, N. Age-associated anatomical and physiological alterations in Caenorhabditis elegans. Mech. Ageing Dev. 2023, 213, 111827. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, Q.; Song, Y.; He, Z.; Zhang, H.; Song, M.; Zhang, X.; Dai, Y.; Karalay, O.; Dieterich, C.; et al. Ageing induces tissue-specific transcriptomic changes in Caenorhabditis elegans. EMBO J. 2022, 41, e109633. [Google Scholar] [CrossRef]
- Copes, N.; Edwards, C.; Chaput, D.; Saifee, M.; Barjuca, I.; Nelson, D.; Paraggio, A.; Saad, P.; Lipps, D.; Stevens, S.M., Jr.; et al. Metabolome and proteome changes with aging in Caenorhabditis elegans. Exp. Gerontol. 2015, 72, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Son, H.G.; Altintas, O.; Kim, E.J.E.; Kwon, S.; Lee, S.V. Age-dependent changes and biomarkers of aging in Caenorhabditis elegans. Aging Cell 2019, 18, e12853. [Google Scholar] [CrossRef] [PubMed]
- Fang, E.F.; Scheibye-Knudsen, M.; Brace, L.E.; Kassahun, H.; SenGupta, T.; Nilsen, H.; Mitchell, J.R.; Croteau, D.L.; Bohr, V.A. Defective mitophagy in XPA via PARP-1 hyperactivation and NAD(+)/SIRT1 reduction. Cell 2014, 157, 882–896. [Google Scholar] [CrossRef]
- Hyun, M.; Lee, J.; Lee, K.; May, A.; Bohr, V.A.; Ahn, B. Longevity and resistance to stress correlate with DNA repair capacity in Caenorhabditis elegans. Nucleic Acids Res. 2008, 36, 1380–1389. [Google Scholar] [CrossRef] [PubMed]
- Marchal, L.; Hamsanathan, S.; Karthikappallil, R.; Han, S.; Shinglot, H.; Gurkar, A.U. Analysis of representative mutants for key DNA repair pathways on healthspan in Caenorhabditis elegans. Mech. Ageing Dev. 2021, 200, 111573. [Google Scholar] [CrossRef]
- Debès, C.; Papadakis, A.; Grönke, S.; Karalay, Ö.; Tain, L.S.; Mizi, A.; Nakamura, S.; Hahn, O.; Weigelt, C.; Josipovic, N.; et al. Ageing-associated changes in transcriptional elongation influence longevity. Nature 2023, 616, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Bernstein, I.A.; Vaughan, F.L. Failure to observe a relationship between bis-(beta-chloroethyl)sulfide-induced NAD depletion and cytotoxicity in the rat keratinocyte culture. J. Toxicol. Environ. Health 1994, 42, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Dequen, F.; Gagnon, S.N.; Desnoyers, S. Ionizing radiations in Caenorhabditis elegans induce poly(ADP-ribosyl)ation, a conserved DNA-damage response essential for survival. DNA Repair (Amst.) 2005, 4, 814–825. [Google Scholar] [CrossRef]
- Crone, B.; Aschner, M.; Schwerdtle, T.; Karst, U.; Bornhorst, J. Elemental bioimaging of Cisplatin in Caenorhabditis elegans by LA-ICP-MS. Metallomics 2015, 7, 1189–1195. [Google Scholar] [CrossRef]
- St-Laurent, J.F.; Gagnon, S.N.; Dequen, F.; Hardy, I.; Desnoyers, S. Altered DNA damage response in Caenorhabditis elegans with impaired poly(ADP-ribose) glycohydrolases genes expression. DNA Repair (Amst.) 2007, 6, 329–343. [Google Scholar] [CrossRef]
- Bae, W.; Park, J.H.; Lee, M.H.; Park, H.W.; Koo, H.S. Hypersensitivity to DNA double-strand breaks associated with PARG deficiency is suppressed by exo-1 and polq-1 mutations in Caenorhabditis elegans. FEBS J. 2020, 287, 1101–1115. [Google Scholar] [CrossRef]
- Janisiw, E.; Raices, M.; Balmir, F.; Paulin, L.F.; Baudrimont, A.; von Haeseler, A.; Yanowitz, J.L.; Jantsch, V.; Silva, N. Poly(ADP-ribose) glycohydrolase coordinates meiotic DNA double-strand break induction and repair independent of its catalytic activity. Nat Commun 2020, 11, 4869. [Google Scholar] [CrossRef]
- Trivedi, S.; Blazícková, J.; Silva, N. PARG and BRCA1-BARD1 cooperative function regulates DNA repair pathway choice during gametogenesis. Nucleic Acids Res. 2022, 50, 12291–12308. [Google Scholar] [CrossRef]
- Byrne, A.B.; McWhirter, R.D.; Sekine, Y.; Strittmatter, S.M.; Miller, D.M.; Hammarlund, M. Inhibiting poly(ADP-ribosylation) improves axon regeneration. eLife 2016, 5, e12734. [Google Scholar] [CrossRef] [PubMed]
- Lagunas-Rangel, F.A. Current role of mammalian sirtuins in DNA repair. DNA Repair (Amst.) 2019, 80, 85–92. [Google Scholar] [CrossRef]
- Singh, C.K.; Chhabra, G.; Ndiaye, M.A.; Garcia-Peterson, L.M.; Mack, N.J.; Ahmad, N. The Role of Sirtuins in Antioxidant and Redox Signaling. Antioxid. Redox Signal. 2018, 28, 643–661. [Google Scholar] [CrossRef] [PubMed]
- Tissenbaum, H.A.; Guarente, L. Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans. Nature 2001, 410, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.A.; Arango, M.; Abderrahmane, S.; Lambert, E.; Tourette, C.; Catoire, H.; Néri, C. Resveratrol rescues mutant polyglutamine cytotoxicity in nematode and mammalian neurons. Nat. Genet. 2005, 37, 349–350. [Google Scholar] [CrossRef] [PubMed]
- Sangaletti, R.; D’Amico, M.; Grant, J.; Della-Morte, D.; Bianchi, L. Knock-out of a mitochondrial sirtuin protects neurons from degeneration in Caenorhabditis elegans. PLoS Genet. 2017, 13, e1006965. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruszkiewicz, J.; Endig, L.; Güver, E.; Bürkle, A.; Mangerich, A. Life-Cycle-Dependent Toxicities of Mono- and Bifunctional Alkylating Agents in the 3R-Compliant Model Organism C. elegans. Cells 2023, 12, 2728. https://doi.org/10.3390/cells12232728
Ruszkiewicz J, Endig L, Güver E, Bürkle A, Mangerich A. Life-Cycle-Dependent Toxicities of Mono- and Bifunctional Alkylating Agents in the 3R-Compliant Model Organism C. elegans. Cells. 2023; 12(23):2728. https://doi.org/10.3390/cells12232728
Chicago/Turabian StyleRuszkiewicz, Joanna, Lisa Endig, Ebru Güver, Alexander Bürkle, and Aswin Mangerich. 2023. "Life-Cycle-Dependent Toxicities of Mono- and Bifunctional Alkylating Agents in the 3R-Compliant Model Organism C. elegans" Cells 12, no. 23: 2728. https://doi.org/10.3390/cells12232728
APA StyleRuszkiewicz, J., Endig, L., Güver, E., Bürkle, A., & Mangerich, A. (2023). Life-Cycle-Dependent Toxicities of Mono- and Bifunctional Alkylating Agents in the 3R-Compliant Model Organism C. elegans. Cells, 12(23), 2728. https://doi.org/10.3390/cells12232728