
Towards the Identification and Characterization of Putative Adult Human Lens Epithelial Stem Cells



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Human Tissue Preparation
Whole Mount of the Human Anterior Lens Epithelium
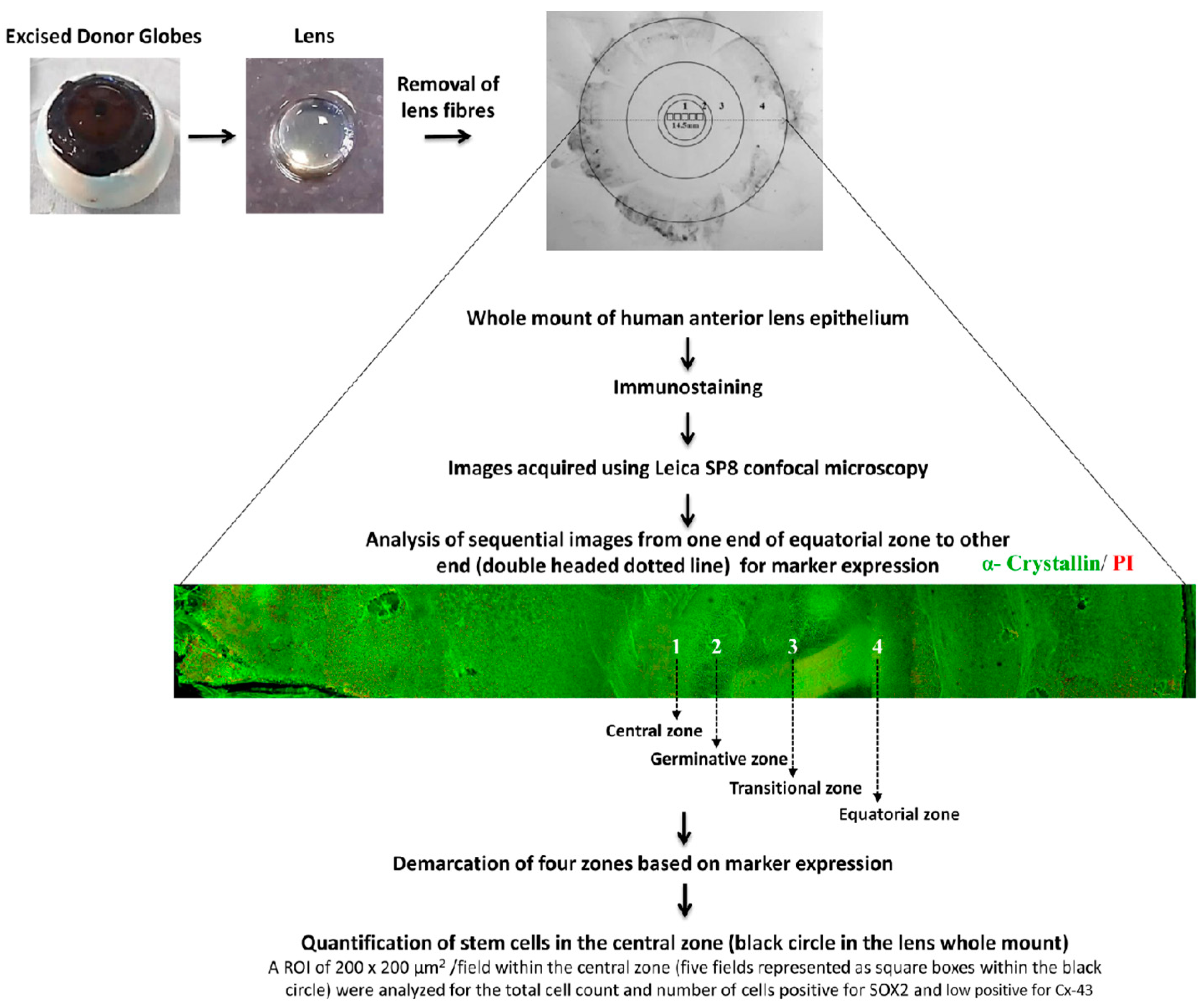

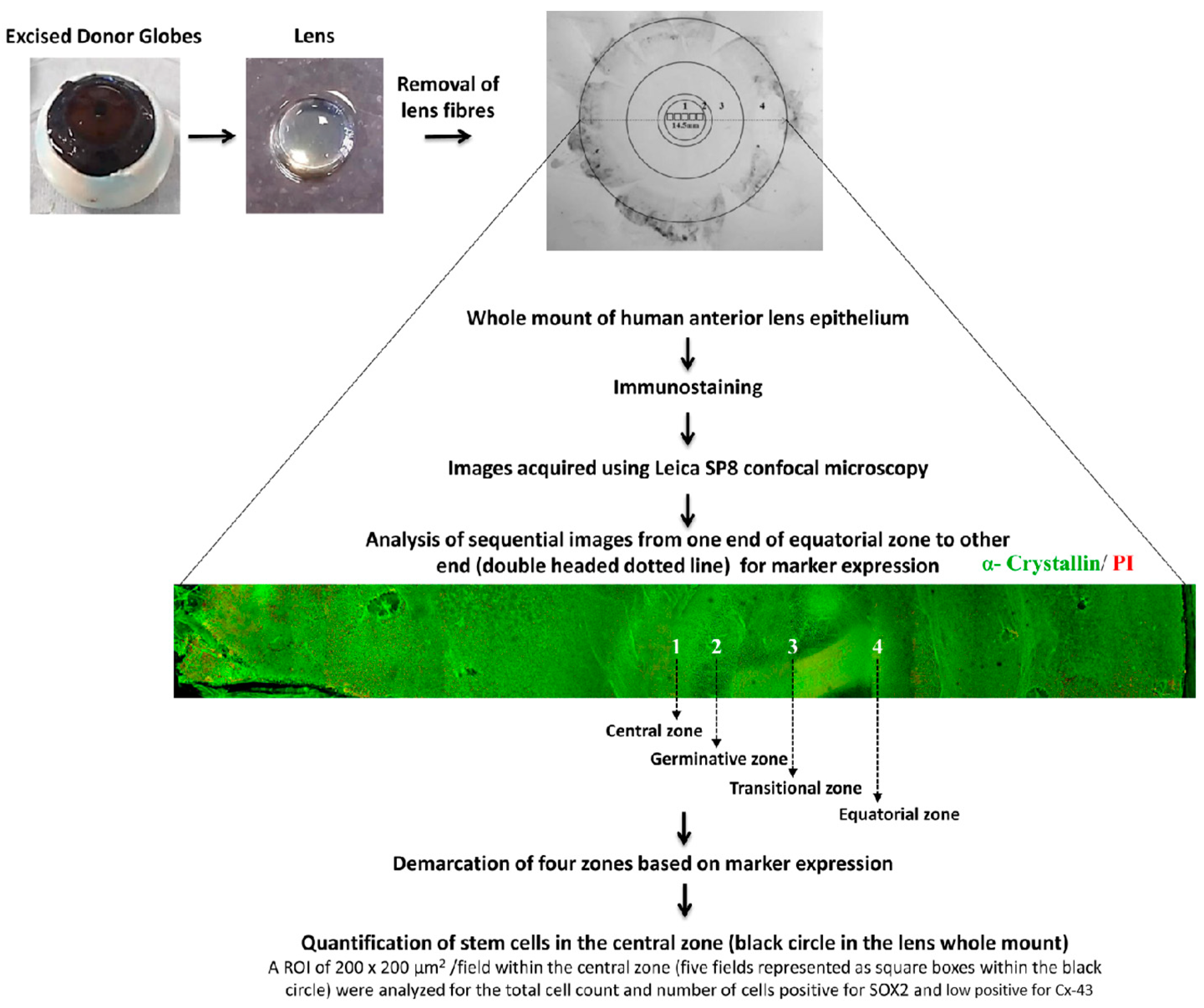{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
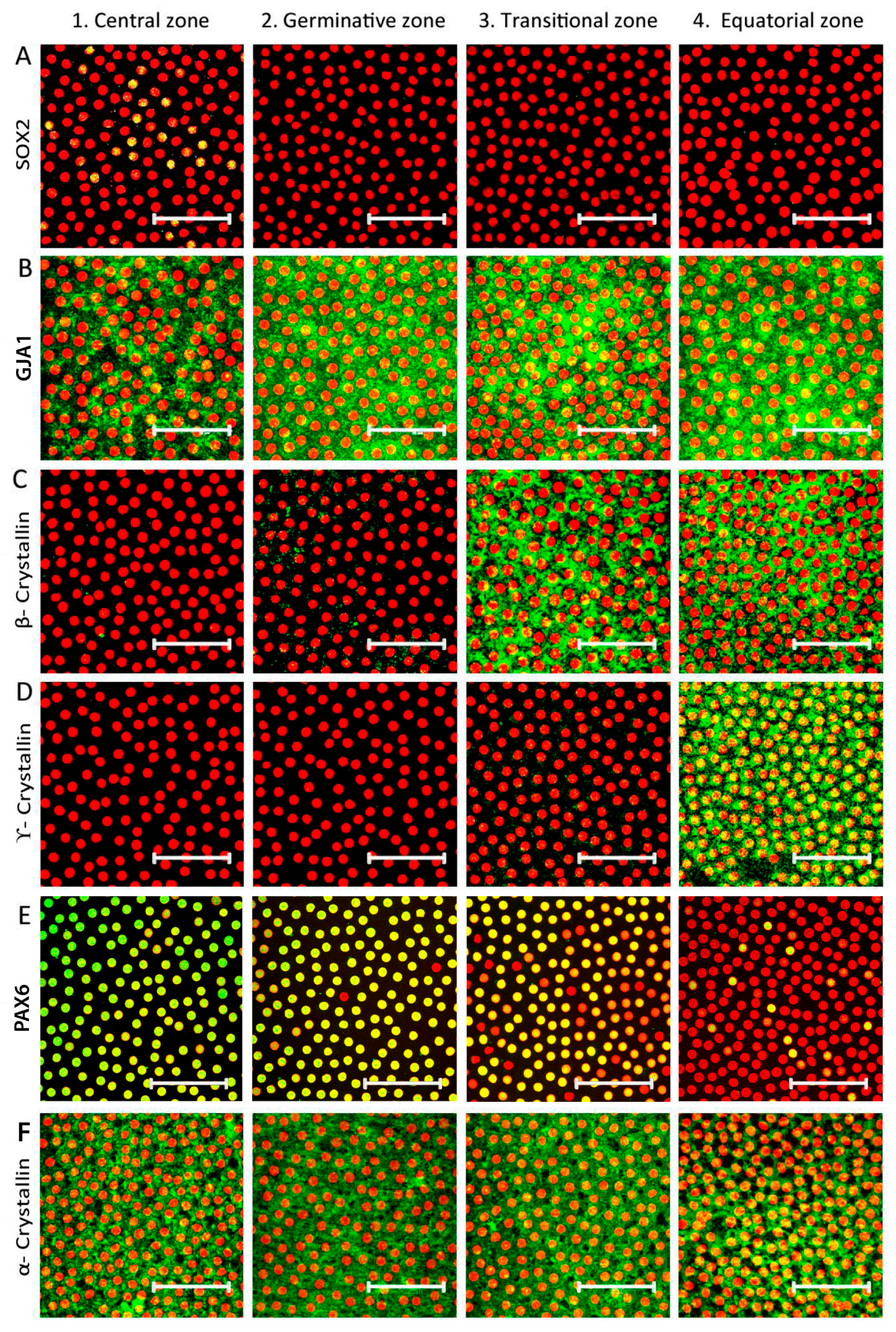
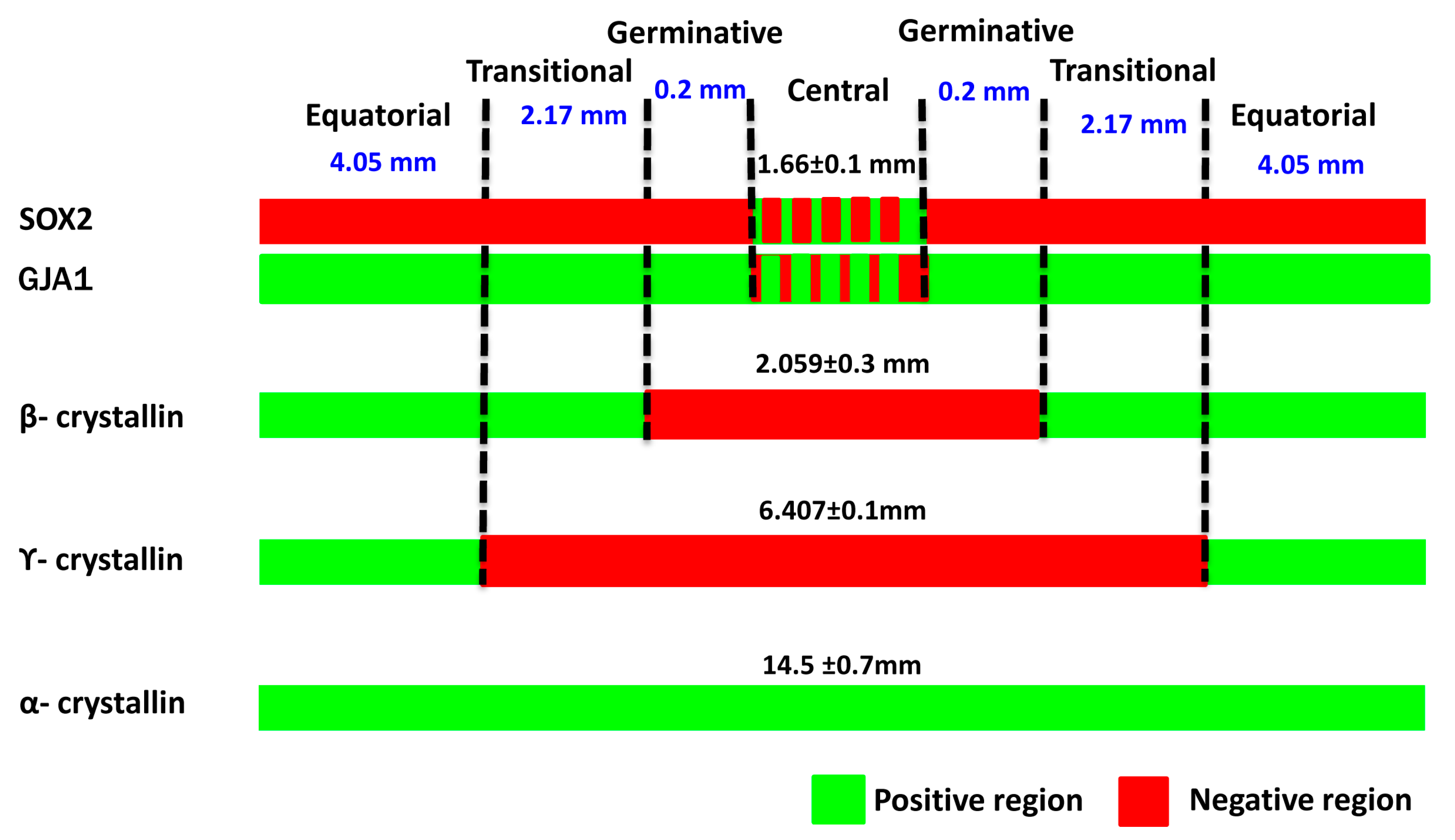
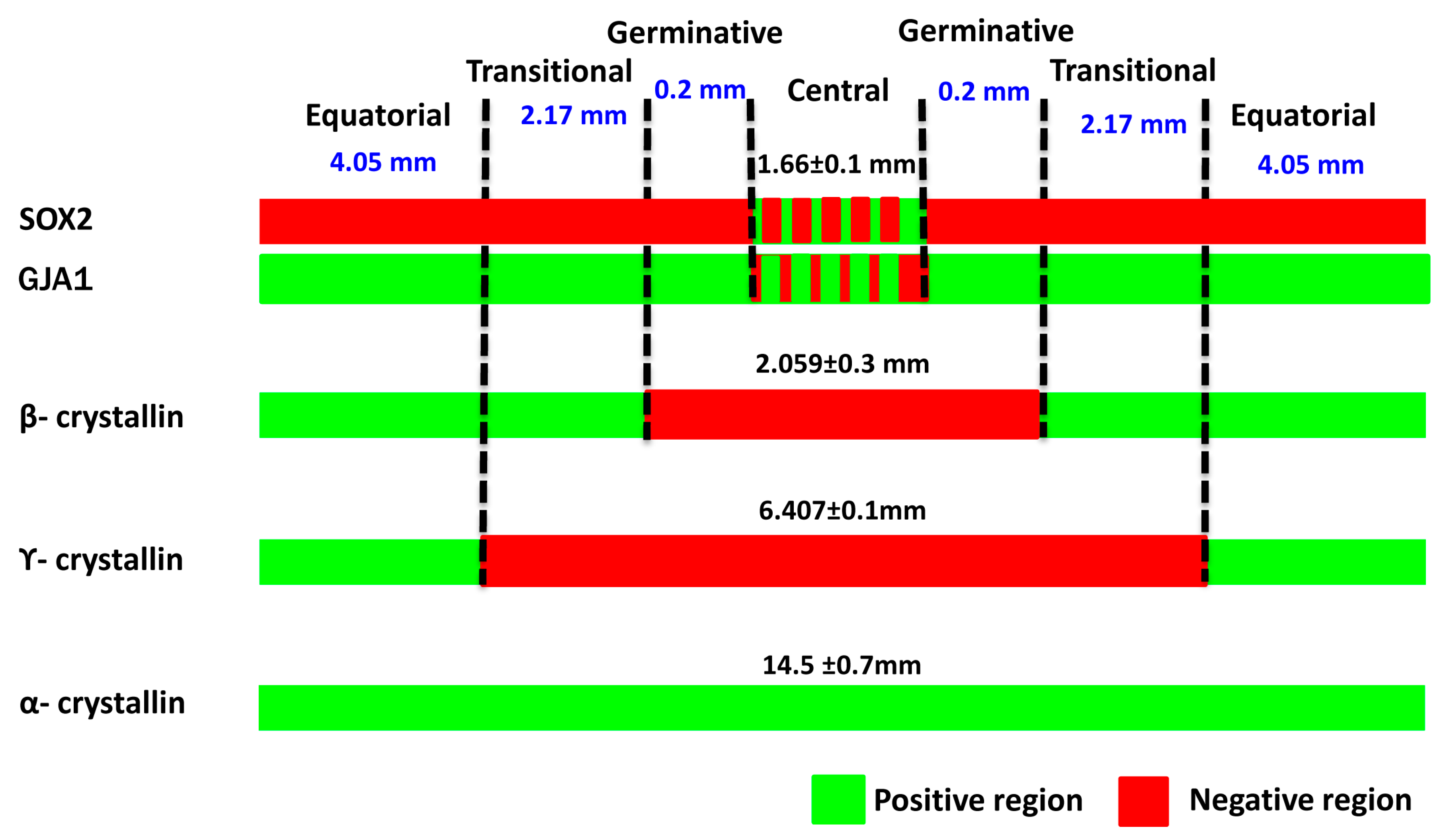
| Zones | Marker Expression | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Based on the Literature | Present Study | |||||||||
| SOX2 [16] | GJA1 [21] | β-cry [2] | γ-cry [2] | α-cry [22] | SOX2 | GJA1 | β-cry | γ-cry | α-cry | |
| Central zone | − | + | − | − | + | +/− | −/+ | − | − | + |
| Germinative zone | − | + | − | − | + | − | + | − | − | + |
| Transitional zone | − | + | + | − | + | − | + | + | − | + |
| Equatorial zone | + | + | + | + | + | − | + | + | + | + |
2.3. Immunostaining
2.4. Confocal Microscopy Imaging
2.5. Image Analysis
2.6. Lens Explant Cultures
2.7. Establishment of Lens Sphere Cultures
3. Results
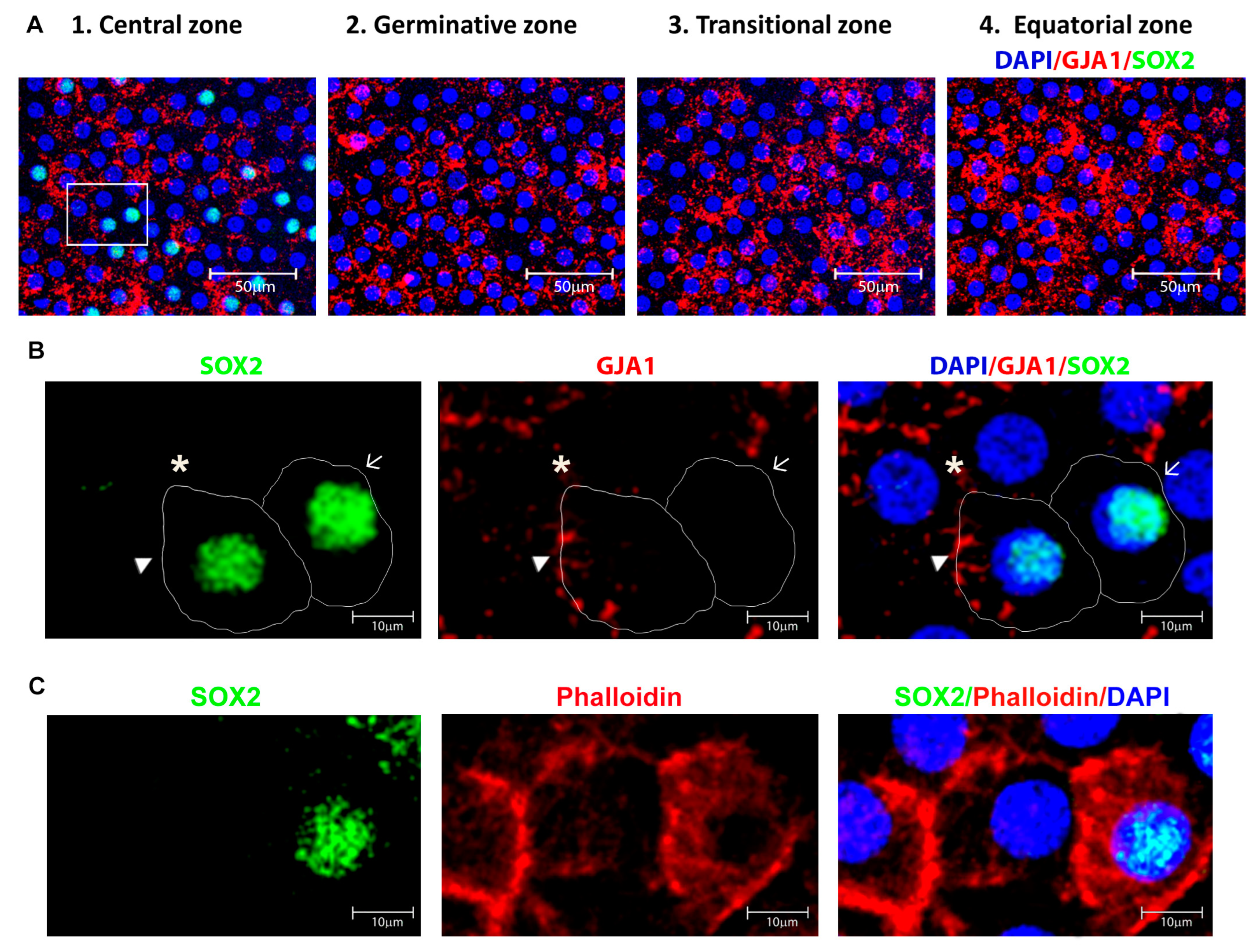
3.1. Identification of Four Zones in Human Anterior Lens Epithelium
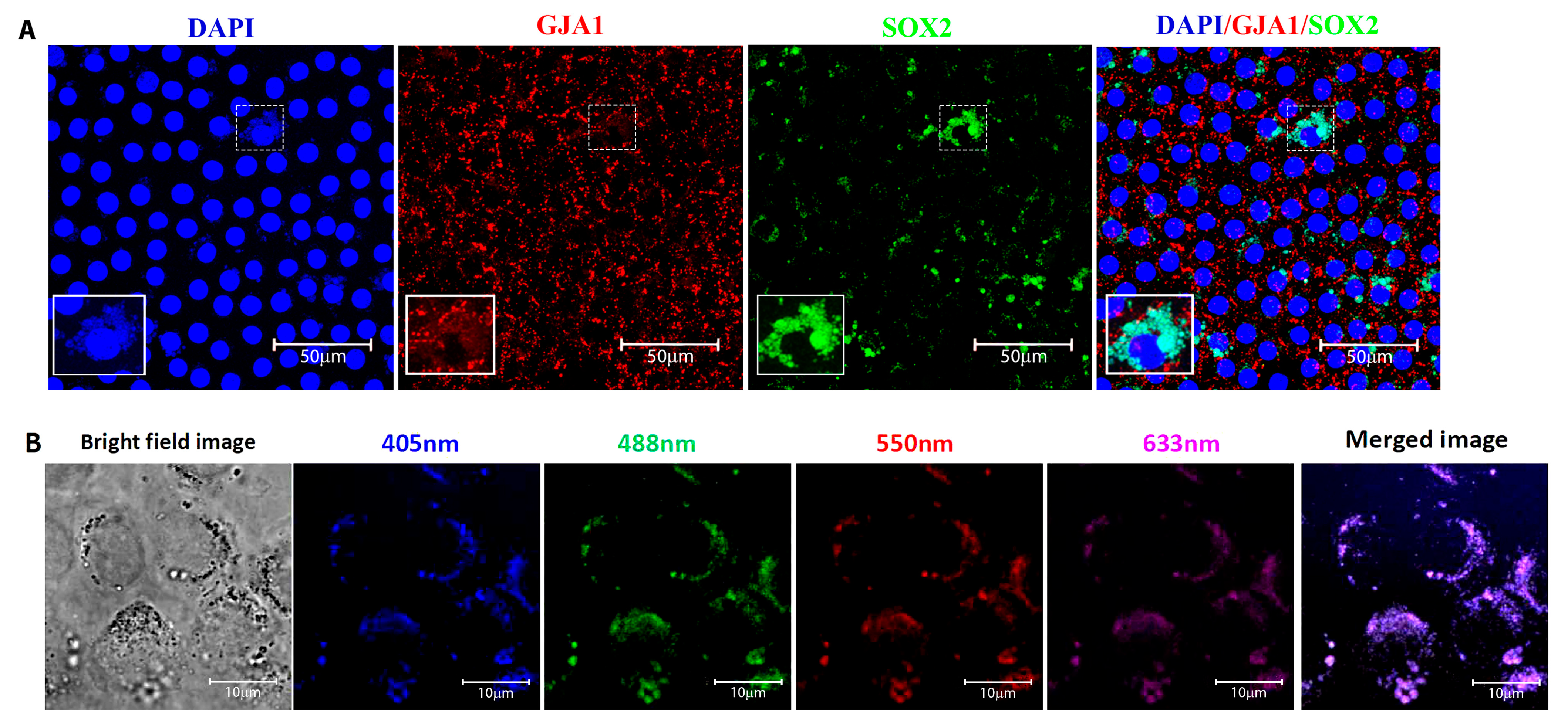
3.2. Identification and Location of Putative Adult Stem Cells in the Human Anterior Lens Epithelium
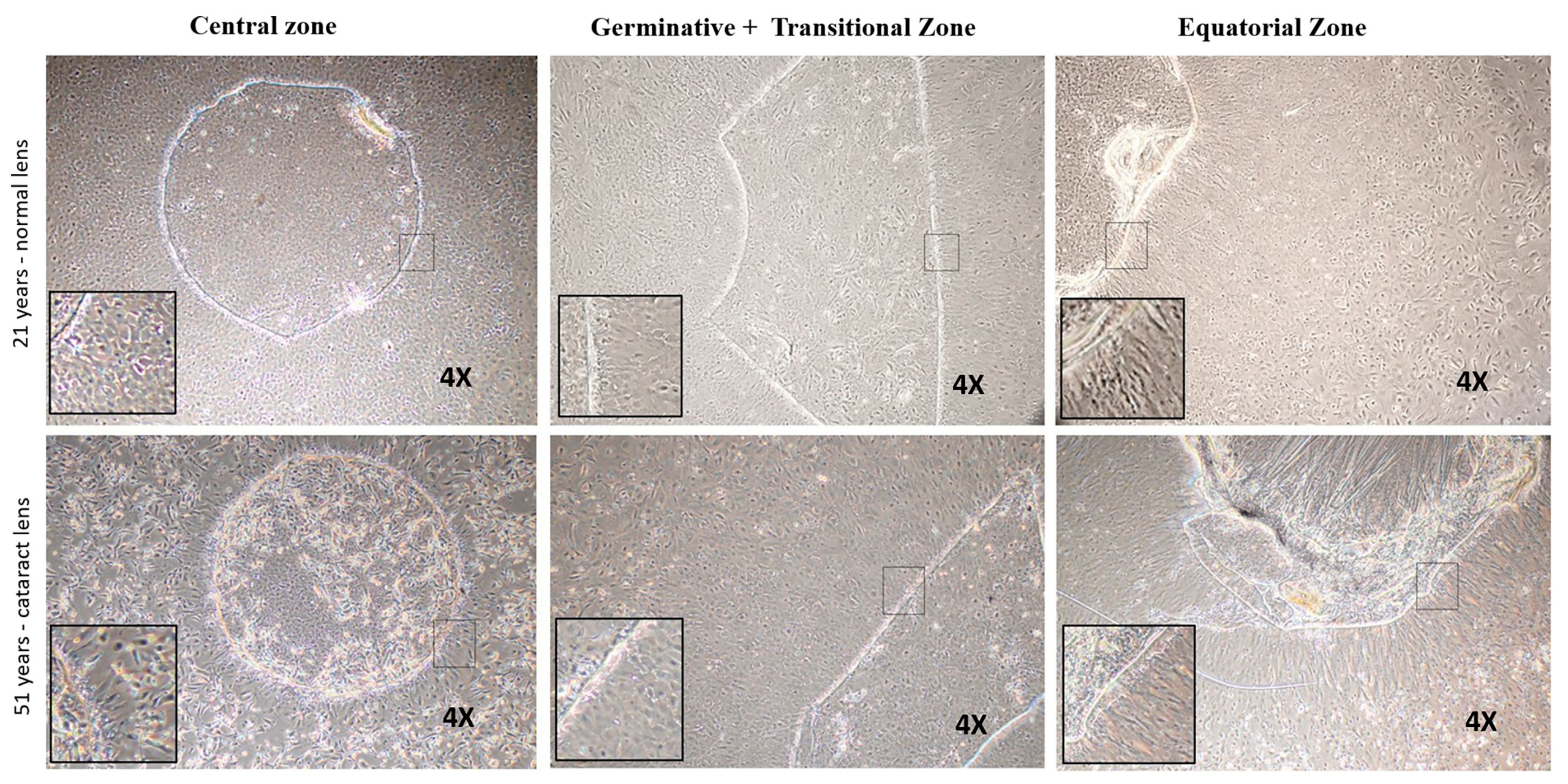
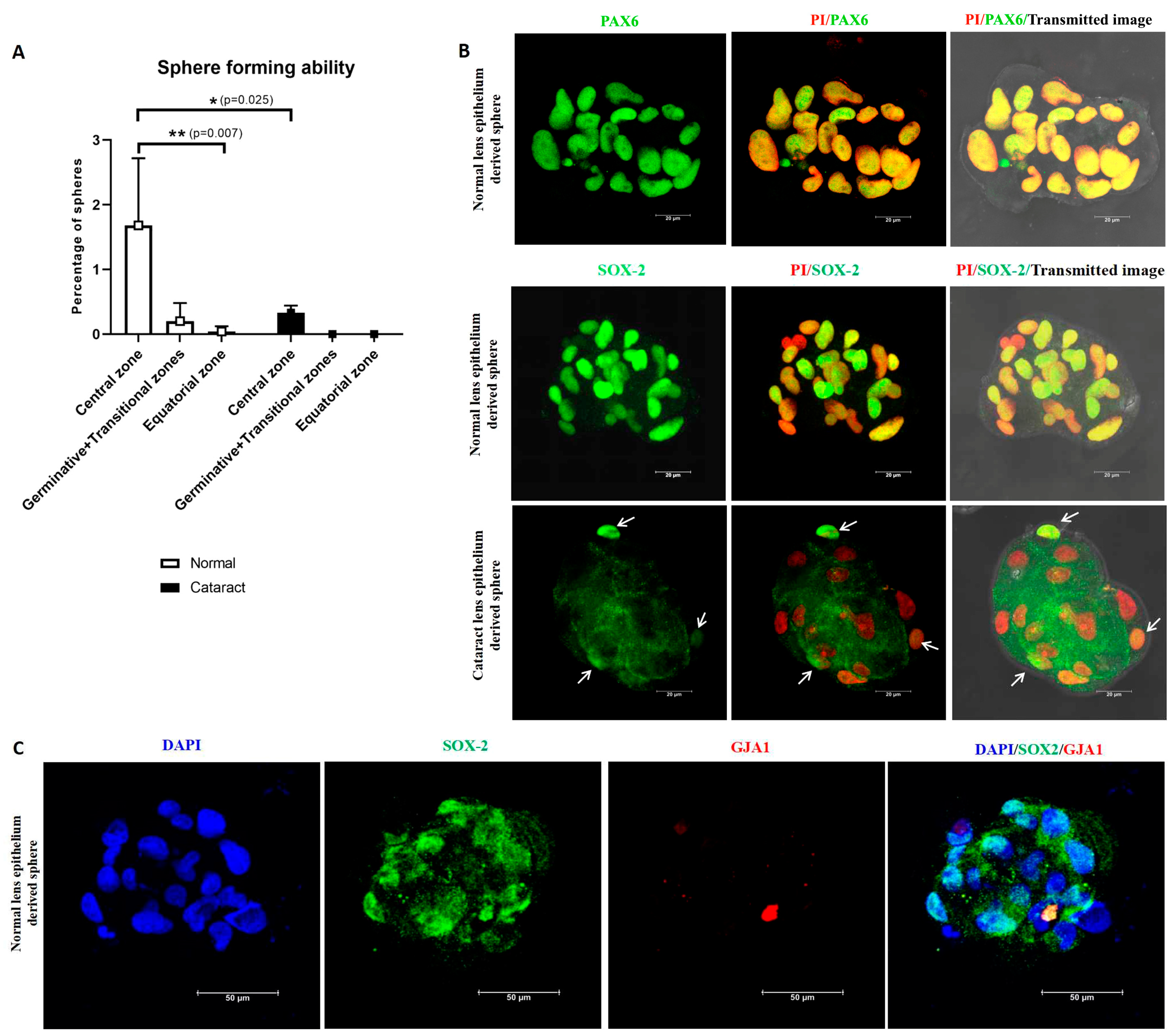
3.3. Functional Characterization of Lens Epithelial Cells—Sphere Forming Ability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SOX-2 | SRY-Box transcription factor 2 |
| GJA1 | Connexin 43 |
| LRCs | Label retaining cells |
| LESCs | Human lens epithelial stem cells |
References
- Muthukkaruppan, V. Inductive Tissue Interaction in the Development of the Mouse Lens in Vitro. J. Exp. Zool. 1965, 159, 269–287. [Google Scholar] [CrossRef] [PubMed]
- Ong, M.D.; Payne, D.M.; Garner, M.H. Differential Protein Expression in Lens Epithelial Whole-Mounts and Lens Epithelial Cell Cultures. Exp. Eye Res. 2003, 77, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; De Maria, A.; Lubura, S.; Šikić, H.; Bassnett, S. The penny pusher: A cellular model of lens growth. Investig. Ophthalmol. Vis. Sci. 2015, 56, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Leiberman, J.; Xu, J.; Lavker, R.M. A Hierarchy of Proliferative Cells Exists in Mouse Lens Epithelium: Implications for Lens Maintenance. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2997–3003. [Google Scholar] [CrossRef] [PubMed]
- Bassnett, S.; Šikić, H. The lens growth process. Prog. Retin. Eye Res. 2017, 60, 181–200. [Google Scholar] [CrossRef] [PubMed]
- Tsonis, P.A.; Del Rio-Tsonis, K. Lens and retina regeneration: Transdifferentiation, stem cells and clinical applications. Exp. Eye Res. 2004, 78, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Gwon, A. Lens regeneration in mammals: A review. Surv. Ophthalmol. 2006, 51, 51–62. [Google Scholar] [CrossRef]
- Lin, H.; Ouyang, H.; Zhu, J.; Huang, S.; Liu, Z.; Chen, S.; Cao, G.; Li, G.; Signer, R.A.J.; Xu, Y.; et al. Lens Regeneration Using Endogenous Stem Cells with Gain of Visual Function. Nature 2016, 531, 323–328. [Google Scholar] [CrossRef]
- Arpitha, P.; Prajna, N.V.; Srinivasan, M.; Muthukkaruppan, V. A Method to Isolate Human Limbal Basal Cells Enriched for a Subset of Epithelial Cells with a Large Nucleus/Cytoplasm Ratio Expressing High Levels of P63. Microsc. Res. Tech. 2008, 71, 469–476. [Google Scholar] [CrossRef]
- Yu, W.Y.; Grierson, I.; Sheridan, C.; Lo, A.C.-Y.; Wong, D.S.-H. Bovine Posterior Limbus: An Evaluation of an Alternative Source for Corneal Endothelial and Trabecular Meshwork Stem/Progenitor Cells. Stem Cells Dev. 2015, 24, 624–639. [Google Scholar] [CrossRef]
- Fu, Q.; Qin, Z.; Yu, J.; Yu, Y.; Tang, Q.; Lyu, D.; Zhang, L.; Chen, Z.; Yao, K. Effects of Senescent Lens Epithelial Cells on the Severity of Age-Related Cortical Cataract in Humans: A Case-Control Study. Medicine 2016, 95, e3869. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Majima, K.; Marunouchi, T. A Study of the Proliferating Activity in Lens Epithelium and the Identification of Tissue-Type Stem Cells. Med. Mol. Morphol. 2008, 41, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Oka, M.; Toyoda, C.; Kaneko, Y.; Nakazawa, Y.; Aizu-Yokota, E.; Takehana, M. Characterization and Localization of Side Population Cells in the Lens. Mol. Vis. 2010, 16, 945–953. [Google Scholar] [PubMed]
- Stern, J.H.; Tian, Y.; Funderburgh, J.; Pellegrini, G.; Zhang, K.; Goldberg, J.L.; Ali, R.R.; Young, M.; Xie, Y.; Temple, S. Regenerating Eye Tissues to Preserve and Restore Vision. Cell Stem Cell 2018, 22, 834–849. [Google Scholar] [CrossRef]
- Kamachi, Y.; Uchikawa, M.; Collignon, J.; Lovell-Badge, R.; Kondoh, H. Involvement of Sox1, 2 and 3 in the early and subsequent molecular events of lens induction. Development 1998, 125, 2521–2532. [Google Scholar] [CrossRef] [PubMed]
- Arnold, K.; Sarkar, A.; Yram, M.A.; Polo, J.M.; Bronson, R.; Sengupta, S.; Seandel, M.; Geijsen, N.; Hochedlinger, K. Sox2+ Adult Stem and Progenitor Cells Are Important for Tissue Regeneration and Survival of Mice. Cell Stem Cell 2011, 9, 317–329. [Google Scholar] [CrossRef]
- Jaenisch, R.; Young, R. Stem cells, the molecular circuitry of pluripotency and nuclear reprogramming. Cell 2008, 32, 567–582. [Google Scholar] [CrossRef]
- Matic, M.; Evans, W.H.; Brink, P.R.; Simon, M. Epidermal Stem Cells Do Not Communicate through Gap Junctions. J. Investig. Dermatol. 2002, 118, 110–116. [Google Scholar] [CrossRef]
- Chen, Z.; Evans, W.H.; Pflugfelder, S.C.; Li, D.-Q. Gap Junction Protein Connexin 43 Serves as a Negative Marker for a Stem Cell-Containing Population of Human Limbal Epithelial Cells. Stem Cells 2006, 24, 1265–1273. [Google Scholar] [CrossRef]
- West-Mays, J.A.; Pino, G.; Lovicu, F.J. Development and Use of the Lens Epithelial Explant System to Study Lens Differentiation and Cataractogenesis. Prog. Retin. Eye Res. 2010, 29, 135–143. [Google Scholar] [CrossRef]
- Berthoud, V.M.; Minogue, P.J.; Osmolak, P.; Snabb, J.I.; Beyer, E.C. Roles and Regulation of Lens Epithelial Cell Connexins. FEBS Lett. 2014, 588, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- McAvoy, J.W. Cell Division, Cell Elongation and Distribution of Alpha-, Beta- and Gamma-Crystallins in the Rat Lens. J. Embryol. Exp. Morphol. 1978, 44, 149–165. [Google Scholar] [PubMed]
- Sundaresan, Y.; Veerappan, M.; Ramasamy, K.S.; Chidambaranathan, G.P. Identification, Quantification and Age-Related Changes of Human Trabecular Meshwork Stem Cells. Eye Vis. 2019, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Sundaresan, Y.; Ramasamy, K.S.; Veerappan, M.; Chidambaranathan, G.P. Functional characterization of adult human trabecular meshwork stem cells. Exp. Cell Res. 2021, 405, 112709. [Google Scholar] [CrossRef] [PubMed]
- Van Velthoven, C.T.J.; Rando, T.A. Stem Cell Quiescence: Dynamism, Restraint, and Cellular Idling. Cell Stem Cell 2019, 24, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Arpitha, P.; Prajna, N.V.; Srinivasan, M.; Muthukkaruppan, V. High Expression of P63 Combined with a Large N/C Ratio Defines a Subset of Human Limbal Epithelial Cells: Implications on Epithelial Stem Cells. Investig. Ophthalmol. Vis. Sci. 2005, 46, 3631–3636. [Google Scholar] [CrossRef]
- Meyer-Blazejewska, E.A.; Kruse, F.E.; Bitterer, K.; Meyer, C.; Hofmann-Rummelt, C.; Wünsch, P.H.; Schlötzer-Schrehardt, U. Preservation of the limbal stem cell phenotype by appropriate culture techniques. Investig. Ophthalmol. Vis. Sci. 2010, 51, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, R.; Yamato, M.; Saito, T.; Oshima, T.; Okano, T.; Tano, Y.; Nishida, K. Enrichment of corneal epithelial stem/progenitor cells using cell surface markers, integrin alpha6 and CD71. Biochem. Biophys. Res. Commun. 2008. [Google Scholar] [CrossRef]
- Shinin, V.; Gayraud-Morel, B.; Tajbakhsh, S. Template DNA-strand co-segregation and asymmetric cell division in skeletal muscle stem cells. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2009; pp. 259–317. [Google Scholar] [CrossRef]
- Pellegrini, G.; Golisano, O.; Paterna, P.; Lambiase, A.; Bonini, S.; Rama, P.; De Luca, M. Location and clonal analysis of stem cells and their differentiated progeny in the human ocular surface. J. Cell Biol. 1999, 145, 769–782. [Google Scholar] [CrossRef]
- Sekiya, I.; Larson, B.L.; Smith, J.R.; Pochampally, R.; Cui, J.G.; Prockop, D.J. Expansion of human adult stem cells from bone marrow stroma: Conditions that maximize the yields of early progenitors and evaluate their quality. Stem Cells 2002, 20, 530–541. [Google Scholar] [CrossRef]
- Isern, J.; Martín-Antonio, B.; Ghazanfari, R.; Martín, A.M.; López, J.A.; del Toro, R.; Sánchez-Aguilera, A.; Arranz, L.; Martín-Pérez, D.; Suárez-Lledó, M.; et al. Self-renewing human bone marrow mesenspheres promote hematopoietic stem cell expansion. Cell Rep. 2013, 3, 1714–1724. [Google Scholar] [CrossRef] [PubMed]
- Goodell, M.A.; Brose, K.; Paradis, G.; Conner, A.S.; Mulligan, R.C. Isolation and functional properties of murine hematopoietic stem cells that are replicating in vivo. J. Exp. Med. 1996, 183, 1797–1806. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.; Helm, K.; Ruegg, P.; Varella-Garcia, M.; Burnham, E.; Majka, S. Adult lung side population cells have mesenchymal stem cell potential. Cytotherapy 2008, 10, 140–151. [Google Scholar] [CrossRef]
- Watanabe, K.; Nishida, K.; Yamato, M.; Umemoto, T.; Sumide, T.; Yamamoto, K.; Maeda, N.; Watanabe, H.; Okano, T.; Tano, Y. Human limbal epithelium contains side population cells expressing the ATP-binding cassette transporter ABCG2. FEBS Lett. 2004. [CrossRef]
- Rama, P.; Matuska, S.; Paganoni, G.; Spinelli, A.; De Luca, M.; Pellegrini, G. Limbal stem-cell therapy and long-term corneal regeneration. N. Engl. J. Med. 2010, 363, 147–155. [Google Scholar] [CrossRef]
- Cheng, X.; Yu, Z.; Song, Y.; Zhang, Y.; Du, J.; Su, Y.; Ma, X. Hair follicle bulge-derived stem cells promote tissue regeneration during skin expansion. Biomed. Pharmacother. 2020, 132, 110805. [Google Scholar] [CrossRef]
- Dorà, N.J.; Hill, R.E.; Collinson, J.M.; West, J.D. Lineage tracing in the adult mouse corneal epithelium supports the limbal epithelial stem cell hypothesis with intermittent periods of stem cell quiescence. Stem Cell Res. 2015, 15, 665–677. [Google Scholar] [CrossRef]
- Konturek-Ciesla, A.; Bryder, D. Stem Cells, Hematopoiesis and Lineage Tracing: Transplantation-Centric Views and Beyond. Front. Cell Dev. Biol. 2022, 10, 903528. [Google Scholar] [CrossRef]
- Kenmotsu, M.; Matsuzaka, K.; Kokubu, E.; Azuma, T.; Inoue, T. Analysis of side population cells derived from dental pulp tissue. Int. Endod. J. 2010, 43, 1132–1142. [Google Scholar] [CrossRef]
- Liang, Y.; Van Zant, G.; Szilvassy, S.J. Effects of Aging on the Homing and Engraftment of Murine Hematopoietic Stem and Progenitor Cells. Blood 2005, 106, 1479–1487. [Google Scholar] [CrossRef]
- Keyes, B.E.; Segal, J.P.; Heller, E.; Lien, W.-H.; Chang, C.-Y.; Guo, X.; Oristian, D.S.; Zheng, D.; Fuchs, E. Nfatc1 Orchestrates Aging in Hair Follicle Stem Cells. Proc. Natl. Acad. Sci. USA 2013, 110, E4950–E4959. [Google Scholar] [CrossRef] [PubMed]
- Mikulicich, A.G.; Young, R.W. Cell proliferation and displacement in the lens epithelium of young rats injected with tritiated thymidine. Investig. Ophthalmol. Vis. Sci. 1963, 2, 344–354. [Google Scholar]
- Rafferty, N.S.; Rafferty, K.A. Cell Population Kinetics of the Mouse Lens Epithelium. J. Cell Physiol. 1981, 107, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Griep, A.E. Cell Cycle Regulation in the Developing Lens. Semin. Cell Dev. Biol. 2006, 17, 686–697. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Cveklova, K.; Oppermann, B.; Kantorow, M.; Cvekl, A. Quantitation of PAX6 and PAX6(5a) transcript levels in adult human lens, cornea, and monkey retina. Mol. Vis. 2001, 7, 1–5. [Google Scholar]
- Bhattacharya, S.; Serror, L.; Nir, E.; Dhiraj, D.; Altshuler, A.; Khreish, M.; Tiosano, B.; Hasson, P.; Panman, L.; Luxenburg, C.; et al. SOX2 Regulates P63 and Stem/Progenitor Cell State in the Corneal Epithelium. Stem Cells 2019, 37, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, B.; Singhal, S.; Lawrence, J.M.; Khaw, P.T.; Limb, G.A. Distribution of Müller Stem Cells within the Neural Retina: Evidence for the Existence of a Ciliary Margin-like Zone in the Adult Human Eye. Exp. Eye Res. 2009, 89, 373–382. [Google Scholar] [CrossRef]
- Carrasco-Garcia, E.; Moreno-Cugnon, L.; Garcia, I.; Borras, C.; Revuelta, M.; Izeta, A.; Lopez-Lluch, G.; de Pancorbo, M.M.; Vergara, I.; Vina, J.; et al. SOX2 Expression Diminishes with Ageing in Several Tissues in Mice and Humans. Mech. Ageing Dev. 2019, 177, 30–36. [Google Scholar] [CrossRef]
- Nishiguchi, S.; Wood, H.; Kondoh, H.; Lovell-Badge, R.; Episkopou, V. Sox1 directly regulates the gamma-crystallin genes and is essential for lens development in mice. Genes Dev. 1998, 12, 776–781. [Google Scholar] [CrossRef]
- Priya, C.G.; Arpitha, P.; Vaishali, S.; Prajna, N.V.; Usha, K.; Sheetal, K.; Muthukkaruppan, V. Adult Human Buccal Epithelial Stem Cells: Identification, Ex-Vivo Expansion, and Transplantation for Corneal Surface Reconstruction. Eye 2011, 25, 1641–1649. [Google Scholar] [CrossRef]
- Shortt, A.J.; Secker, G.A.; Munro, P.M.; Khaw, P.T.; Tuft, S.J.; Daniels, J.T. Characterization of the Limbal Epithelial Stem Cell Niche: Novel Imaging Techniques Permit in Vivo Observation and Targeted Biopsy of Limbal Epithelial Stem Cells. Stem Cells 2007, 25, 1402–1409. [Google Scholar] [CrossRef] [PubMed]
- Di Iorio, E.; Barbaro, V.; Ruzza, A.; Ponzin, D.; Pellegrini, G.; De Luca, M. Isoforms of DeltaNp63 and the Migration of Ocular Limbal Cells in Human Corneal Regeneration. Stem Cells 2005, 23, 63–73. [Google Scholar]
- Priya, C.G.; Prasad, T.; Prajna, N.V.; Muthukkaruppan, V. Identification of Human Corneal Epithelial Stem Cells on the Basis of High ABCG2 Expression Combined with a Large N/C Ratio. Microsc. Res. Tech. 2013, 76, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Priya, C.G.; Waseema, A.; Narendran, S.; Babu, K.N.; Kim, R.; Muthukkaruppan, V. Decline in the function of adult human retinal pigment epithelial stem cells with ageing. Investig. Ophthalmol. Vis. Sci. 2023, 64, 5387. [Google Scholar]
- Turan, G.; Turan, M. The Evaluation of TUNEL, PCNA and SOX2 Expressions in Lens Epithelial Cells of Cataract Patients with Pseudoexfoliation Syndrome. Curr. Eye Res. 2020, 45, 12–16. [Google Scholar] [CrossRef]
- Heslop, L.; Morgan, J.E.; Partridge, T.A. Evidence for a myogenic stem cell that is exhausted in dystrophic muscle. J. Cell Sci. 2000, 113, 2299–2308. [Google Scholar] [CrossRef]





| Antibody | Species and Type | Manufacturer, City/State, Country | Catalogue Number | Dilution Factor |
|---|---|---|---|---|
| SOX2 | Rabbit monoclonal | Abcam, Waltham, MA, USA | ab92494 | 1:100 |
| Connexin-43 (GJA1) | Mouse monoclonal | BD Biosciences, Franklin Lakes, NJ, USA | 610062 | 1:200 |
| PAX6 | Rabbit polyclonal | Proteintech, Manchester, UK | 12323-1-AP | 1:250 |
| α-crystallin | Mouse monoclonal | SantaCruz Biotechnology, Dallas, TX, USA | sc-28306 | 1:50 |
| β-crystallin | Mouse monoclonal | SantaCruz Biotechnology, Dallas, TX, USA | sc-374496 | 1:50 |
| γ-crystallin | Mouse monoclonal | SantaCruz Biotechnology, Dallas, TX, USA | sc-365256 | 1:50 |
| Phalloidin-TRITC | Synthetic: Peptide Sequence | Sigma, Saint Louis, MO, USA | P1951 | 1:400 |
| anti-mouse | Goat polyclonal | Agilent, Santa Clara, CA, USA | E0433 | 1:200 |
| anti-rabbit | Mouse monoclonal | SantaCruz Biotechnology, Dallas, TX, USA | sc-2491 | 1:200 |
| S. No | Zone | Percentage of Cells (Mean ± SEM) (n = 3 Donor Eyes) | ||
|---|---|---|---|---|
| SOX2+ GJA1− | SOX2+ GJA1+ | SOX2− GJA1+ | ||
| 1 | Central zone | 1.89 ± 0.84 | 7.27 ± 5.07 | 90.83 ± 5.9 |
| 2 | Germinative zone | 0 | 0 | 100 |
| 3 | Transitional zone | 0 | 0 | 100 |
| 4 | Equatorial zone | 0 | 0 | 100 |
| Spheres Formed/103 Cells (Spheres ≥ 60 μm) | |||
|---|---|---|---|
| Age (Years) | Central Zone (% of Spheres) | Germinative + Transitional Zone (% of Spheres) | Equatorial Zone (% of Spheres) |
| Spheres established from normal lens epithelium | |||
| 18 | 1.3 | 0.6 | 0.2 |
| 20 | 1.1 | 0 | 0 |
| 21 | 3.1 | 0.4 | 0 |
| 41 | 2.4 | 0 | 0 |
| 54 | 0.5 | 0 | 0 |
| Mean ± SD | 1.68 ± 1.04 | 0.2 ± 0.28 | 0.04 ± 0.08 |
| Spheres established from cataractous lens epithelium | |||
| 51 | 0.4 | 0 | 0 |
| 53 | 0.4 | 0 | 0 |
| 60 | 0.2 | 0 | 0 |
| Mean ± SD | 0.33 ± 0.11 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saranya, P.; Shekhar, M.; Haripriya, A.; Muthukkaruppan, V.; Gowri Priya, C. Towards the Identification and Characterization of Putative Adult Human Lens Epithelial Stem Cells. Cells 2023, 12, 2727. https://doi.org/10.3390/cells12232727
Saranya P, Shekhar M, Haripriya A, Muthukkaruppan V, Gowri Priya C. Towards the Identification and Characterization of Putative Adult Human Lens Epithelial Stem Cells. Cells. 2023; 12(23):2727. https://doi.org/10.3390/cells12232727
Chicago/Turabian StyleSaranya, Pandi, Madhu Shekhar, Aravind Haripriya, Veerappan Muthukkaruppan, and Chidambaranathan Gowri Priya. 2023. "Towards the Identification and Characterization of Putative Adult Human Lens Epithelial Stem Cells" Cells 12, no. 23: 2727. https://doi.org/10.3390/cells12232727
APA StyleSaranya, P., Shekhar, M., Haripriya, A., Muthukkaruppan, V., & Gowri Priya, C. (2023). Towards the Identification and Characterization of Putative Adult Human Lens Epithelial Stem Cells. Cells, 12(23), 2727. https://doi.org/10.3390/cells12232727