
Glucose Improves the Efficacy of Photobiomodulation in Changing ATP and ROS Levels in Mouse Fibroblast Cell Cultures


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Experimental Outline
- (1)
- Non-stressed (10% FBS) with glucose (~1 g/L of glucose);
- (2)
- Non-stressed (10% FBS) without glucose;
- (3)
- Stressed (1% FBS) with glucose (~1 g/L of glucose);
- (4)
- Stressed (1% FBS) without glucose.
2.2. Photobiomodulation Treatment
2.3. Cell Viability Test
2.4. Cell (Number) Proliferation and Protein Measurement
2.5. Reactive Oxygen Species (ROS) Assay
2.6. Adenosine Triphosphate (ATP) Assay
2.7. Statistics
3. Results
3.1. Non-Stressed (Normal) Conditions
3.1.1. Cell Viability
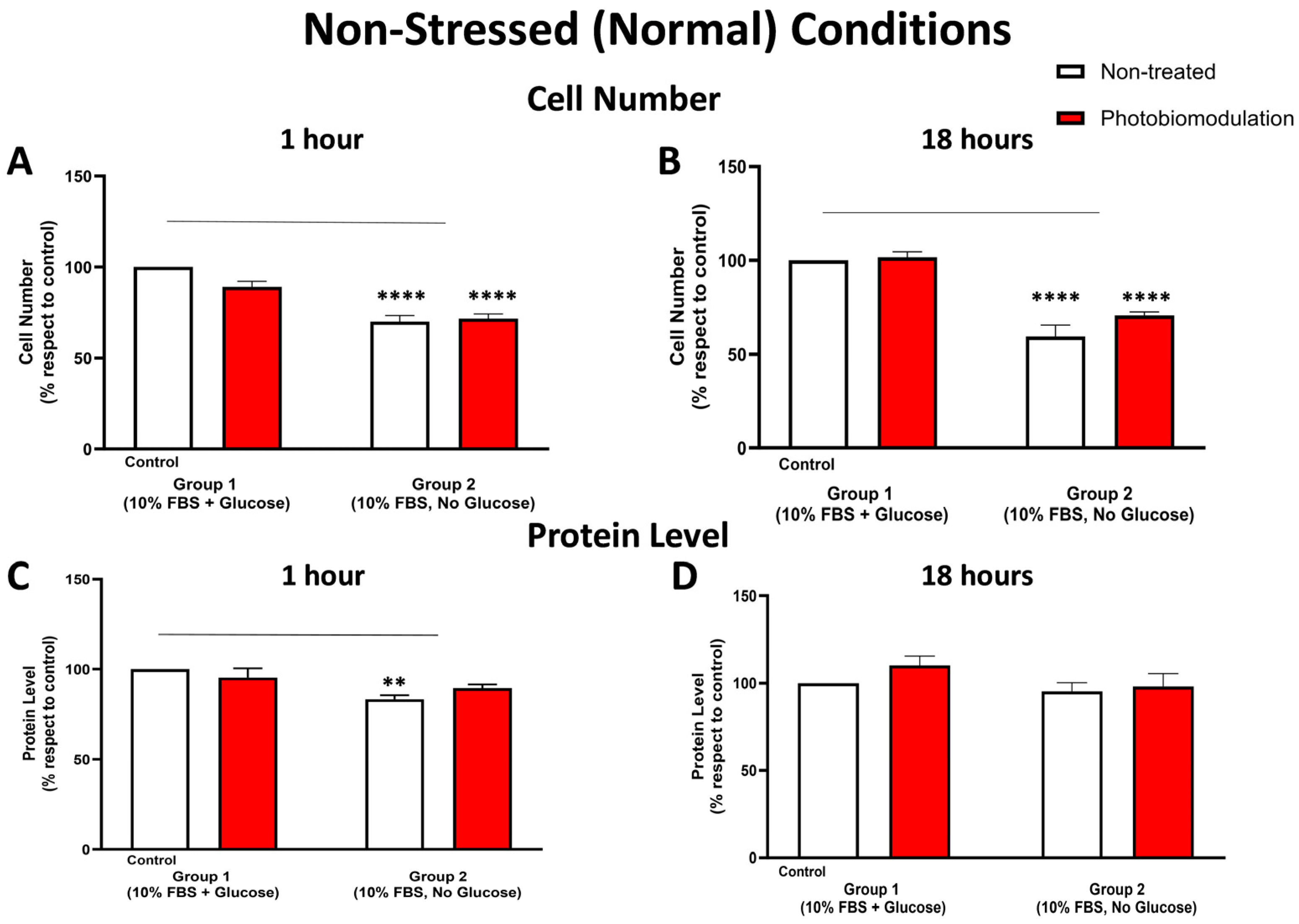
3.1.2. Cell Number (Proliferation) and Protein Content
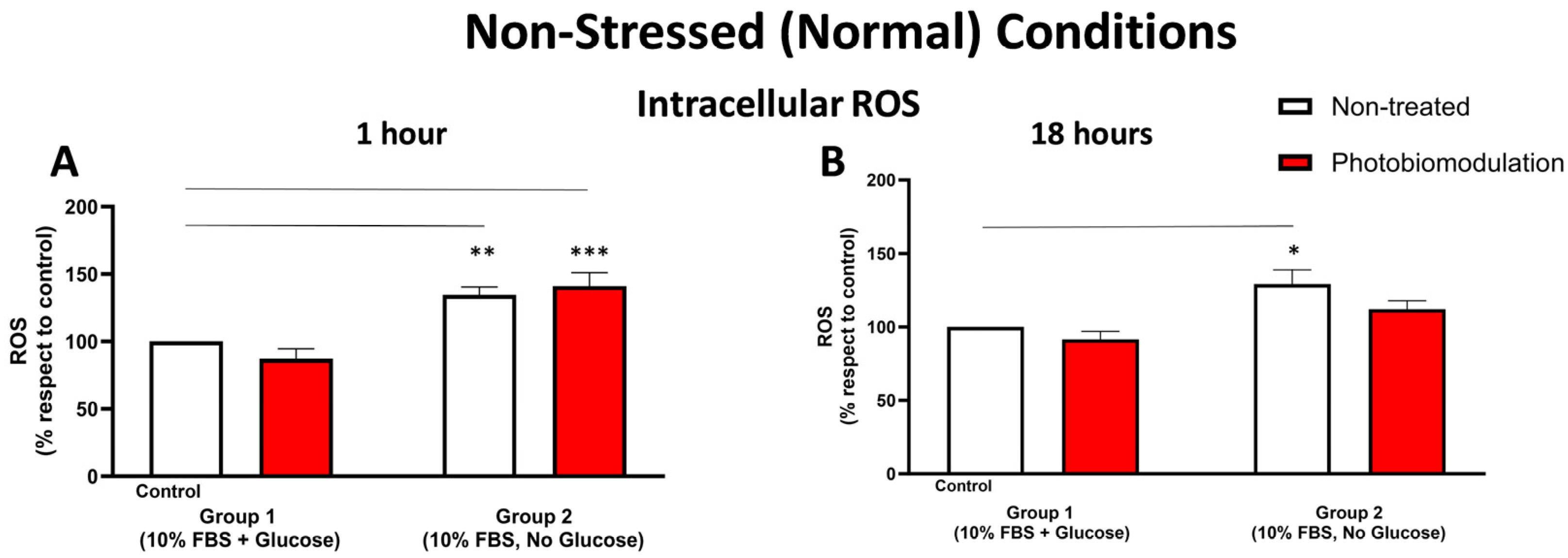
3.1.3. Cellular Stress (ROS Levels)
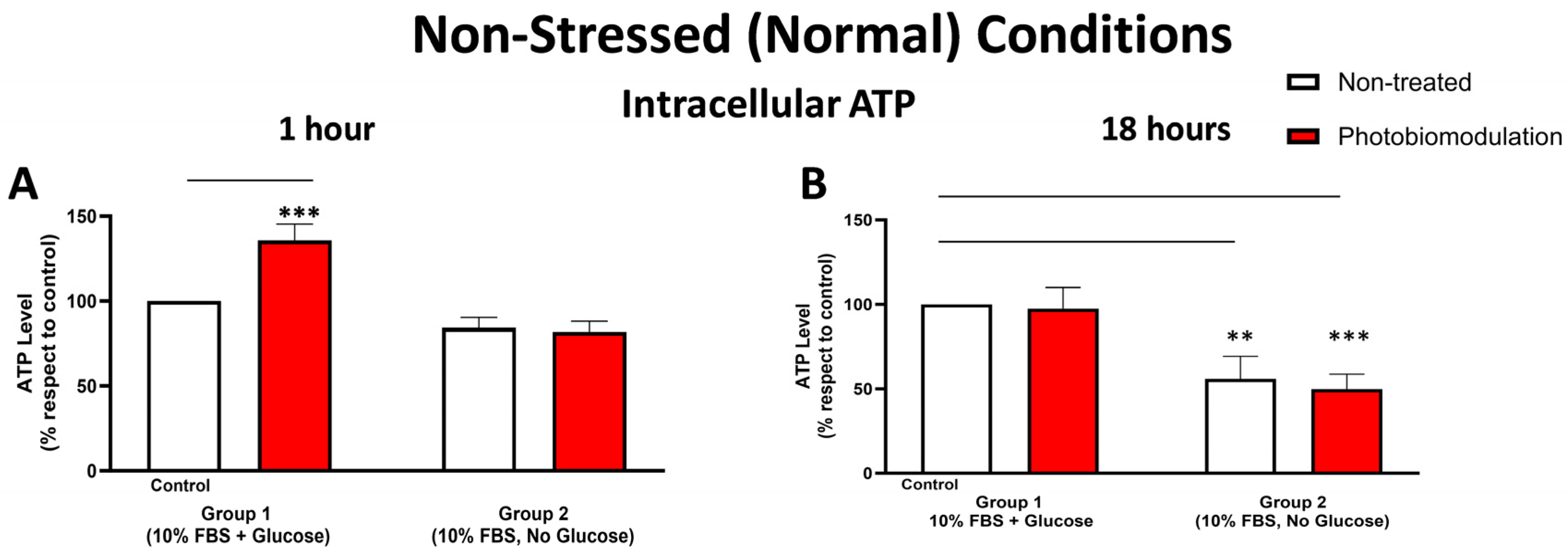
3.1.4. Metabolic (ATP) Activity
3.2. Stressed Conditions
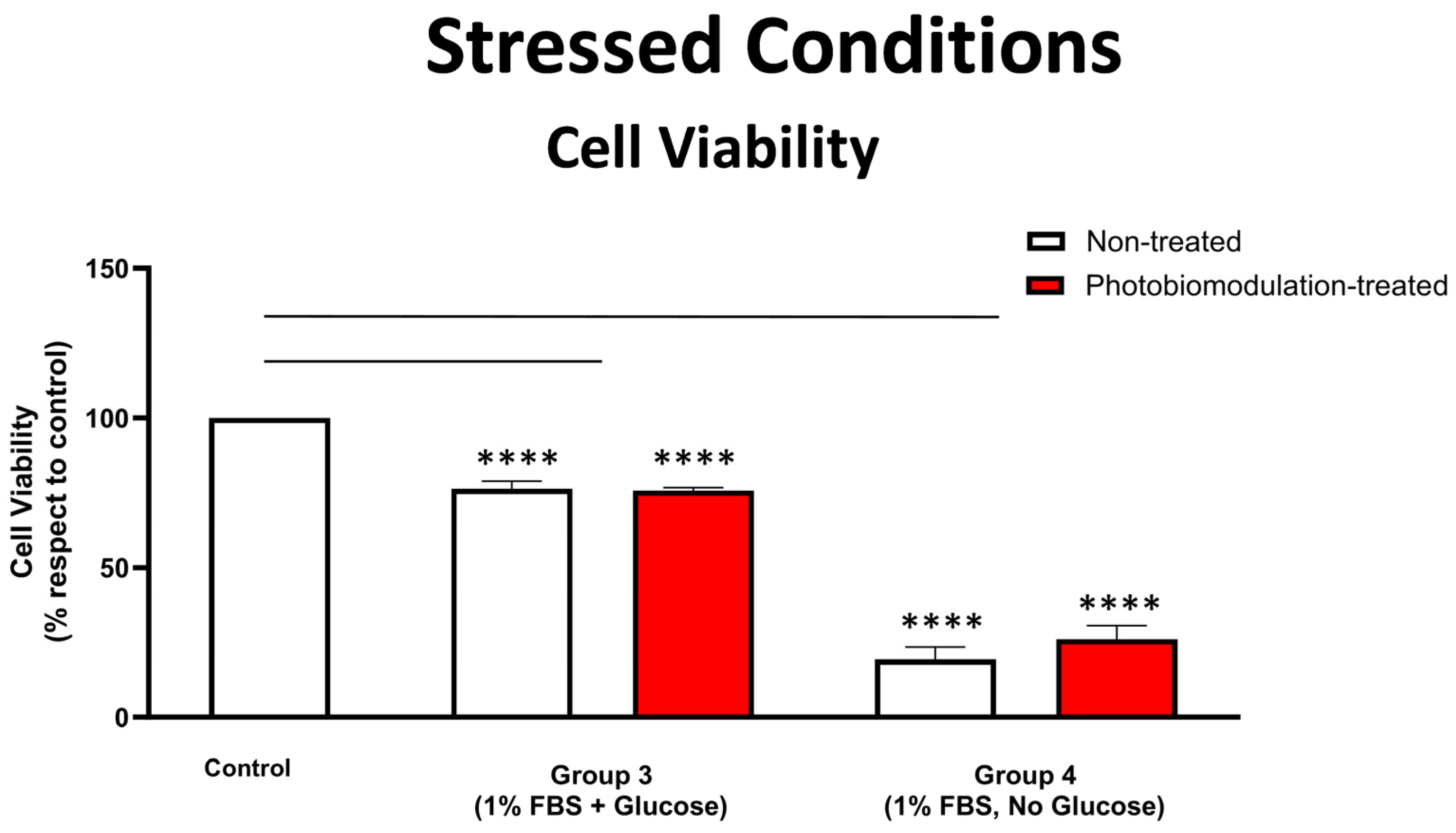
3.2.1. Cell Viability
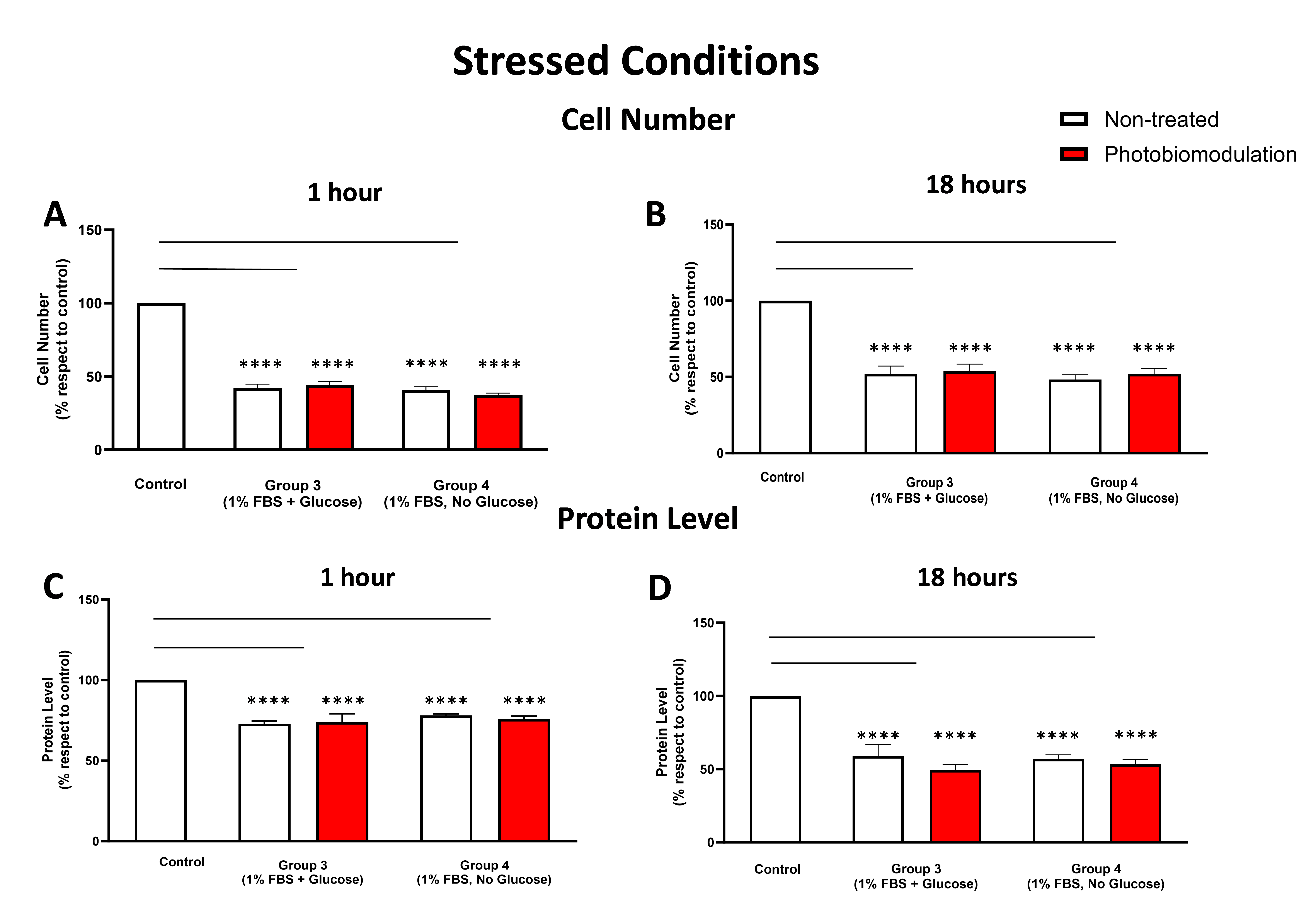
3.2.2. Cell Number (Proliferation) and Protein Content
3.2.3. Cellular Stress (ROS Levels)
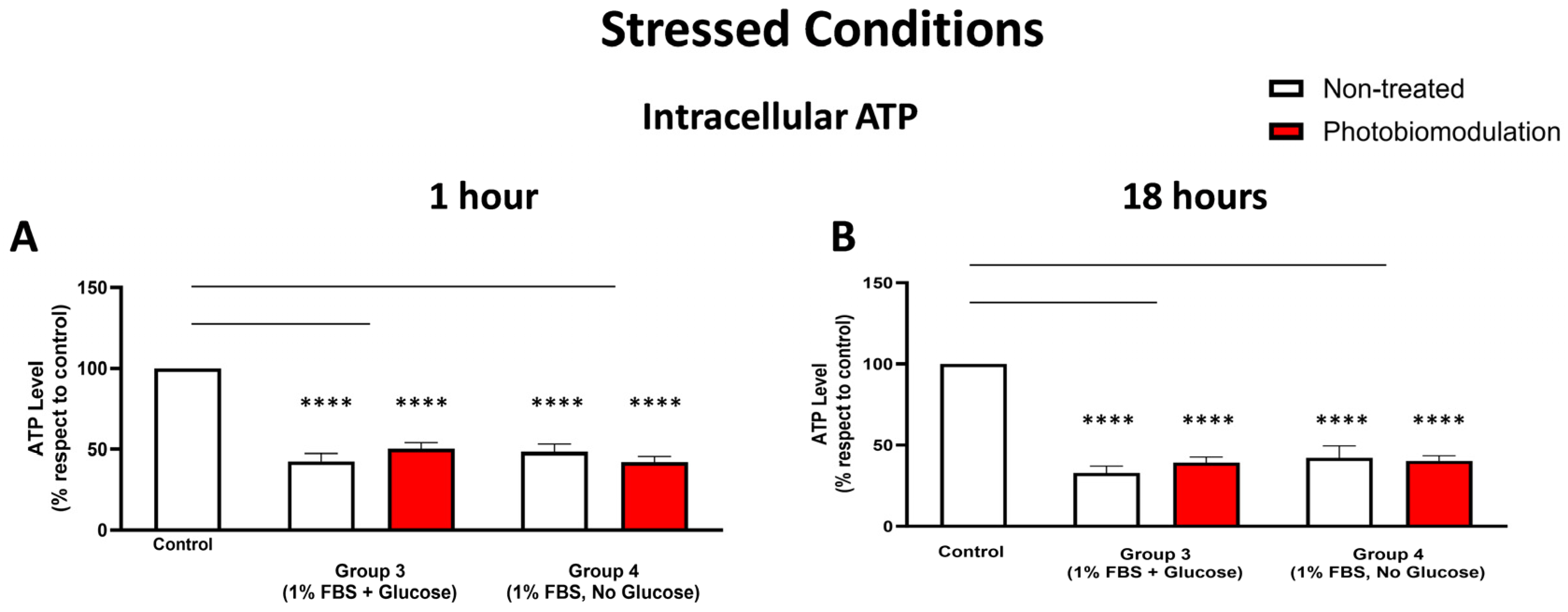
3.2.4. Metabolic (ATP) Activity
4. Discussion
4.1. Photobiomodulation Was Not Detrimental to Cell Survival
4.2. Photobiomodulation Did Not Enhance Most Measures of Cell Function and Survival
4.3. Photobiomodulation Was Effective Only with ATP and ROS Levels and When Glucose Was Available
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnstone, D.M.; Moro, C.; Stone, J.; Benabid, A.L.; Mitrofanis, J. Turning on Lights to Stop Neurodegeneration: The Potential of Near Infrared Light Therapy in Alzheimer’s and Parkinson’s Disease. Front. Neurosci. 2016, 9, 500. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, M.R. Shining light on the head: Photobiomodulation for brain disorders. BBA Clin. 2016, 6, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, C.; Liebert, A.; Pang, V.; Magistretti, P.; Mitrofanis, J. Lights on for Autism: Exploring Photobiomodulation as an Effective Therapeutic Option. Neurol. Int. 2022, 14, 884–893. [Google Scholar] [CrossRef] [PubMed]
- Valverde, A.; Mitrofanis, J. Photobiomodulation for Hypertension and Alzheimer’s Disease. J. Alzheimer’s Dis. 2022, 90, 1045–1055. [Google Scholar] [CrossRef]
- Bicknell, B.; Liebert, A.; Johnstone, D.; Kiat, H. Photobiomodulation of the microbiome: Implications for metabolic and inflammatory diseases. Lasers Med. Sci. 2019, 2, 317–327. [Google Scholar] [CrossRef]
- Liebert, A.; Krause, A.; Goonetilleke, N.; Bicknell, B.; Kiat, H. A Role for Photobiomodulation in the Prevention of Myocardial Ischemic Reperfusion Injury: A Systematic Review and Potential Molecular Mechanisms. Sci. Rep. 2017, 1, 42386. [Google Scholar] [CrossRef]
- Vanin, A.A.; Verhagen, E.; Barboza, S.D.; Costa, L.O.P.; Leal-Junior, E.C.P. Photobiomodulation therapy for the improvement of muscular performance and reduction of muscular fatigue associated with exercise in healthy people: A systematic review and meta-analysis. Lasers Med. Sci. 2018, 1, 181–214. [Google Scholar] [CrossRef]
- Barolet, D.; Christiaens, F.; Hamblin, M.R. Infrared and skin: Friend or foe. J. Photochem. Photobiol. B Biol. 2016, 155, 78–85. [Google Scholar] [CrossRef]
- Yu, W.; Chi, L.H.; Naim, J.O.; Lanzafame, R.J. Improvement of host response to sepsis by photobiomodulation. Lasers Surg. Med. 1997, 21, 262–268. [Google Scholar] [CrossRef]
- Passarella, S.; Karu, T. Absorption of monochromatic and narrow band radiation in the visible and near IR by both mitochondrial and non-mitochondrial photoacceptors results in photobiomodulation. J. Photochem. Photobiol. B Biol. 2014, 140, 344–358. [Google Scholar] [CrossRef]
- Pastore, D.; Greco, M.; Passarella, S. Specific helium-neon laser sensitivity of the purified cytochrome c oxidase. Int. J. Radiat. Biol. 2000, 76, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Amaroli, A.; Ravera, S.; Parker, S.; Panfoli, I.; Benedicenti, A.; Benedicenti, S. An 808-nm Diode Laser with a Flat-Top Handpiece Positively Photobiomodulates Mitochondria Activities. Photomed. Laser Surg. 2016, 34, 564–571. [Google Scholar] [CrossRef]
- Amaroli, A.; Pasquale, C.; Zekiy, A.; Utyuzh, A.; Benedicenti, S.; Signore, A.; Ravera, S. Photobiomodulation and Oxidative Stress: 980 nm Diode Laser Light Regulates Mitochondrial Activity and Reactive Oxygen Species Production. Oxid. Med. Cell. Longev. 2021, 2021, 6626286. [Google Scholar] [CrossRef] [PubMed]
- Ravera, S.; Ferrando, S.; Agas, D.; De Angelis, N.; Raffetto, M.; Sabbieti, M.G.; Signore, A.; Benedicenti, S.; Amaroli, A. 1064 nm Nd:YAG laser light affects transmembrane mitochondria respiratory chain complexes. J. Biophotonics 2019, 12, e201900101. [Google Scholar] [CrossRef] [PubMed]
- Salehpour, F.; Hamblin, M.R. Photobiomodulation for Parkinson’s Disease in Animal Models: A Systematic Review. Biomolecules 2020, 10, 610. [Google Scholar] [CrossRef]
- Hamblin, M.R.; Liebert, A. Photobiomodulation Therapy Mechanisms beyond Cytochrome c Oxidase. Photobiomodulation Photomed. Laser Surg. 2021, 40, 75–77. [Google Scholar] [CrossRef]
- Hamblin, M.R. Mechanisms and applications of the anti-inflammatory effects of photobiomodulation. AIMS Biophys. 2017, 3, 337–361. [Google Scholar] [CrossRef]
- Lima, P.L.; Pereira, C.V.; Nissanka, N.; Arguello, T.; Gavini, G.; da Costa Maranduba, C.M.; Diaz, F.; Moraes, C.T. Photobiomodulation enhancement of cell proliferation at 660 nm does not require cytochrome c oxidase. J. Photochem. Photobiol. B Biol. 2019, 194, 71–75. [Google Scholar] [CrossRef]
- Sommer, A.P.; Haddad, M.K.; Fecht, H.J. Light Effect on Water Viscosity: Implication for ATP Biosynthesis. Sci. Rep. 2015, 1, 12029. [Google Scholar] [CrossRef]
- Towle, H.C. Glucose as a regulator of eukaryotic gene transcription. Trends Endocrinol. Metab. 2005, 10, 489–494. [Google Scholar] [CrossRef]
- Nakrani, M.N.; Wineland, R.H.; Anjum, F. Physiology, Glucose Metabolism; StatPearls Publishing: Treasure Island, FL, USA, 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK560599/ (accessed on 17 July 2023).
- Kumari, A. (Ed.) Chapter 1—Glycolysis. In Sweet Biochemistry: Remembering Structures, Cycles, and Pathways by Mnemonics; Academic Press: Cambridge, MA, USA, 2018; pp. 1–5. Available online: https://www.sciencedirect.com/science/article/pii/B9780128144534000017 (accessed on 24 October 2017).
- Judge, A.; Dodd, M.S. Metabolism. Essays Biochem. 2020, 64, 607–647. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.; Cox, M. Lehninger Principles of Biochemistry, 5th ed.; W.H. Freeman: New York City, NY, USA, 2008; Available online: https://www.semanticscholar.org/paper/Lehninger-Principles-of-Biochemistry-(5th-edition)-Nelson-Cox/0b18428e9d89c9c52d46ff71b8101a5da3653ec0 (accessed on 15 June 2008).
- Gatenby, R.A.; Gillies, R.J. Why do cancers have high aerobic glycolysis? Nat. Rev. Cancer 2004, 11, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Preiss, J.; Walsh, D.A. The Comparative Biochemistry of Glycogen and Starch. In Biol Carbohydrates; Ginsburg, V., Robbins, P., Eds.; John Wiley Sons, Inc.: New York, NY, USA, 1981; Volume 1, Vii+320p, pp. 199–314. Available online: https://eurekamag.com/research/029/309/029309502.php (accessed on 1 January 1981).
- Magistretti, P.J.; Allaman, I. Lactate in the brain: From metabolic end-product to signalling molecule. Nat. Rev. Neurosci. 2018, 4, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Castro, K.M.R.; de Paiva Carvalho, R.L.; Junior, G.M.R.; Tavares, B.A.; Simionato, L.H.; Bortoluci, C.H.F.; Soto, C.A.T.; Ferraresi, C. Can photobiomodulation therapy (PBMT) control blood glucose levels and alter muscle glycogen synthesis? J. Photochem. Photobiol. B Biol. 2020, 207, 111877. [Google Scholar] [CrossRef]
- Powner, M.B.; Jeffery, G. Systemic glucose levels are modulated by specific wavelengths in the solar light spectrum that shift mitochondrial metabolism. PLoS ONE 2022, 17, e0276937. [Google Scholar] [CrossRef]
- Ray, R.S.; Agrawal, N.; Sharma, A.; Hans, R.K. Use of L-929 cell line for phototoxicity assessment. Toxicol. Vitr. 2008, 7, 1775–1781. [Google Scholar] [CrossRef]
- Liu, Y.; Song, X.D.; Liu, W.; Zhang, T.Y.; Zuo, J. Glucose deprivation induces mitochondrial dysfunction and oxidative stress in PC12 cell line. J. Cell. Mol. Med. 2003, 7, 49–56. [Google Scholar] [CrossRef]
- Marambio, P.; Toro, B.; Sanhueza, C.; Troncoso, R.; Parra, V.; Verdejo, H.; García, L.; Quiroga, C.; Munafo, D.; Díaz-Elizondo, J.; et al. Glucose deprivation causes oxidative stress and stimulates aggresome formation and autophagy in cultured cardiac myocytes. Biochim. Biophys. Acta Mol. Basis Dis. 2010, 6, 509–518. [Google Scholar] [CrossRef]
- Li, R.; Kato, H.; Taguchi, Y.; Deng, X.; Minagawa, E.; Nakata, T.; Umeda, M. Glucose Starvation-Caused Oxidative Stress Induces Inflammation and Autophagy in Human Gingival Fibroblasts. Antioxidants 2022, 11, 1907. [Google Scholar] [CrossRef]
- Fuchs, C.; Schenk, M.S.; Pham, L.; Cui, L.; Anderson, R.R.; Tam, J. Photobiomodulation Response from 660 nm is Different and More Durable than That from 980 nm. Lasers Surg. Med. 2021, 53, 1279–1293. [Google Scholar] [CrossRef]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free radicals: Properties, sources, targets, and their implication in various diseases. Indian J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS Generation and Antioxidant Defense Systems in Normal and Malignant Cells. Oxid. Med. Cell. Longev. 2019, 2019, 6175804. [Google Scholar] [CrossRef] [PubMed]
- Brondon, P.; Stadler, I.; Lanzafame, R.J. A study of the effects of phototherapy dose interval on photobiomodulation of cell cultures. Lasers Surg. Med. 2005, 36, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Pires-Oliveira, D.A.A.; Oliveira, R.F.; Machado, A.H.A.; Zângaro, R.A.; Pacheco-Soares, C. Laser Biomodulation on L 929 Cell Culture. Photomed. Laser Surg. 2010, 28, 167–171. [Google Scholar] [CrossRef]
- Luna, G.L.F.; de Andrade, A.L.M.; Brassolatti, P.; Bossini, P.S.; de Freitas Anibal, F.; Parizotto, N.A.; de Oliveira Leal, M. Biphasic Dose/Response of Photobiomodulation Therapy on Culture of Human Fibroblasts. Photobiomodulation, Photomedicine. Laser Surg. 2020, 38, 413–418. [Google Scholar] [CrossRef]
- Giannakopoulos, E.; Katopodi, A.; Rallis, M.; Politopoulos, K.; Alexandratou, E. The effects of low power laser light at 661 nm on wound healing in a scratch assay fibroblast model. Lasers Med. Sci. 2022, 1, 27. [Google Scholar] [CrossRef]
- Hurry, M.M.; Lefta, S.H.; Omran, D.G.; Ghaleb, R.A. Biphasic dose-response of low-level laser therapy on culture cells fibroblasts. AIP Conf. Proc. 2022, 2547, 30015. [Google Scholar] [CrossRef]
- Liang, H.L.; Whelan, H.T.; Eells, J.T.; Meng, H.; Buchmann, E.; Lerch-Gaggl, A.; Wong-Riley, M. Photobiomodulation partially rescues visual cortical neurons from cyanide-induced apoptosis. Neuroscience 2006, 2, 639–649. [Google Scholar] [CrossRef]
- Ying, R.; Liang, H.L.; Whelan, H.T.; Eells, J.T.; Wong-Riley, M.T.T. Pretreatment with near-infrared light via light-emitting diode provides added benefit against rotenone- and MPP+-induced neurotoxicity. Brain Res. 2008, 1243, 167–173. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Nagata, K.; Tedford, C.E.; Hamblin, M.R. Low-level laser therapy (810 nm) protects primary cortical neurons against excitotoxicity in vitro. J. Biophotonics 2014, 7, 656–664. [Google Scholar] [CrossRef]
- Chaudary, S.; Karner, L.; Weidinger, A.; Meixner, B.; Rieger, S.; Metzger, M.; Zipperle, J.; Dungel, P. In vitro effects of 635 nm photobiomodulation under hypoxia/reoxygenation culture conditions. J. Photochem. Photobiol. B Biol. 2020, 209, 111935. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, Y.Y.; Wang, Y.; Lyu, P.; Hamblin, M.R. Photobiomodulation of human adipose-derived stem cells using 810 nm and 980 nm lasers operates via different mechanisms of action. Biochim. Biophys. Acta Gen. Subj. 2017, 2, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, Y.Y.; Wang, Y.; Lyu, P.; Hamblin, M.R. Red (660 nm) or near-infrared (810 nm) photobiomodulation stimulates, while blue (415 nm), green (540 nm) light inhibits proliferation in human adipose-derived stem cells. Sci. Rep. 2017, 1, 7781. [Google Scholar] [CrossRef] [PubMed]
- Amaroli, A.; Ravera, S.; Baldini, F.; Benedicenti, S.; Panfoli, I.; Vergani, L. Photobiomodulation with 808-nm diode laser light promotes wound healing of human endothelial cells through increased reactive oxygen species production stimulating mitochondrial oxidative phosphorylation. Lasers Med. Sci. 2019, 3, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.L.; Whelan, H.T.; Eells, J.T.; Wong-Riley, M.T.T. Near-infrared light via light-emitting diode treatment is therapeutic against rotenone- and 1-methyl-4-phenylpyridinium ion-induced neurotoxicity. Neuroscience 2008, 4, 963–974. [Google Scholar] [CrossRef]
- Oyebode, O.A.; Houreld, N.N. Photobiomodulation at 830 nm Stimulates Migration, Survival and Proliferation of Fibroblast Cells. Diabetes Metab. Syndr. Obes. Targets Ther. 2022, 15, 2885–2900. [Google Scholar] [CrossRef]
- Weinrich, T.W.; Coyne, A.; Salt, T.E.; Hogg, C.; Jeffery, G. Improving mitochondrial function significantly reduces metabolic, visual, motor and cognitive decline in aged Drosophila melanogaster. Neurobiol. Aging 2017, 60, 34–43. [Google Scholar] [CrossRef]
- Mostafavinia, A.; Ahmadi, H.; Amini, A.; Roudafshani, Z.; Hamblin, M.R.; Chien, S.; Bayat, M. The effect of photobiomodulation therapy on antioxidants and oxidative stress profiles of adipose derived mesenchymal stem cells in diabetic rats. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2021, 262, 120157. [Google Scholar] [CrossRef]
- Nonarath, H.J.; Hall, A.E.; SenthilKumar, G.; Abroe, B.; Eells, J.T.; Liedhegner, E.S. 670 nm photobiomodulation modulates bioenergetics and oxidative stress, in rat Müller cells challenged with high glucose. PLoS ONE 2021, 16, e0260968. [Google Scholar] [CrossRef]
- Zheng, Q.; Zhang, J.; Zuo, X.; Sun, J.; Liang, Z.; Hu, X.; Wang, Z.; Li, K.; Song, J.; Ding, T.; et al. Photobiomodulation Promotes Neuronal Axon Regeneration after Oxidative Stress and Induces a Change in Polarization from M1 to M2 in Macrophages via Stimulation of CCL2 in Neurons: Relevance to Spinal Cord Injury. J. Mol. Neurosci. 2021, 6, 1290–1300. [Google Scholar] [CrossRef]
- Dong, T.; Zhang, Q.; Hamblin, M.R.; Wu, M.X. Low-level light in combination with metabolic modulators for effective therapy of injured brain. J. Cereb. Blood Flow Metab. 2015, 35, 1435–1444. [Google Scholar] [CrossRef] [PubMed]
- Hoh Kam, J.; Mitrofanis, J. Does photobiomodulation require glucose to work effectively? Neural Regen. Res. 2024, 5, 945–946. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoh Kam, J.; Mitrofanis, J. Glucose Improves the Efficacy of Photobiomodulation in Changing ATP and ROS Levels in Mouse Fibroblast Cell Cultures. Cells 2023, 12, 2533. https://doi.org/10.3390/cells12212533
Hoh Kam J, Mitrofanis J. Glucose Improves the Efficacy of Photobiomodulation in Changing ATP and ROS Levels in Mouse Fibroblast Cell Cultures. Cells. 2023; 12(21):2533. https://doi.org/10.3390/cells12212533
Chicago/Turabian StyleHoh Kam, Jaimie, and John Mitrofanis. 2023. "Glucose Improves the Efficacy of Photobiomodulation in Changing ATP and ROS Levels in Mouse Fibroblast Cell Cultures" Cells 12, no. 21: 2533. https://doi.org/10.3390/cells12212533
APA StyleHoh Kam, J., & Mitrofanis, J. (2023). Glucose Improves the Efficacy of Photobiomodulation in Changing ATP and ROS Levels in Mouse Fibroblast Cell Cultures. Cells, 12(21), 2533. https://doi.org/10.3390/cells12212533