1. Introduction
Human space exploration is expected to increase in the coming decades with a rise in both commercial and touristic spaceflight as well as deep-space interplanetary exploration missions. However, there are many health risks associated with long-term spaceflight. One of the most frequently reported issues by astronauts during their space missions are related to skin sensitivity (1.12/flight year, compared to 0.97/flight year for other notable medical events, such as upper respiratory symptoms [
1]) and include itches and dryness of the skin, as well as rashes and delayed wound healing [
1,
2,
3]. Small cutaneous injuries and delayed healing of wounds, with pus-forming wounds reported on wrists, fingers, and feet, were observed during spaceflight [
1]. Delayed wound healing is a threat for astronauts’ health as wounds disrupt the anatomy of the skin, thereby compromising its barrier function, which forms an active protection from environmental hazards [
4]. The skin heals wounds through a multiphase and multicomponent process. After wounding, a blood clot forms, which stops the bleeding. During the inflammatory phase, immune cells migrate to the wound site to clear any pathogens and debris. Cytokines and growth factors expressed by several cell types, including fibroblast, act as chemoattractants and stimulate the migration of different cell types [
5,
6,
7]. During the next phase, fibroblasts, keratinocytes, and endothelial cells migrate to the wound and start proliferating, which marks the next phase: the proliferating phase. The migration of these cells requires the continuous remodeling of several cytoskeletal components, whose function is to provide the cell with adhesion, contractility, and polarity [
8,
9]. Finally, during the remodeling phase, the wound will further contract and fibroblasts excrete an extracellular matrix (ECM) to remodel the tissue and form a scar. This final phase can last for many weeks after injury [
9,
10]. Interference in the wound healing process at any phase can lead to a delayed or defective repair and increases the risk of infection and health complications.
In space, astronauts are exposed to an environment different from the one on Earth. Long-term microgravity exposure deconditions the weight-bearing musculoskeletal system as well as the cardiovascular system. It induces upward-fluid-shifts, which may be linked to ocular and visual acuity problems that are clinically classified as spaceflight associated neuro-ocular syndrome (SANS) [
11,
12,
13,
14,
15]. Furthermore, higher levels of ionizing radiation increase the risks for developing cataracts, cardiovascular diseases, as well as the development of cancers later in life [
16,
17,
18,
19,
20,
21]. Finally, the confined and isolated environment of the spacecraft together with high workload increases the levels of psychological stress, and elevated stress hormone levels have been measured after both short- and long-term spaceflight [
22,
23,
24,
25,
26]. Long-term exposure to this spaceflight environment has shown to induce changes in astronauts’ skin (as reviewed in Radstake et al. [
27]), including a delayed epidermal proliferation, loss of elasticity, and degradation of dermal fibers [
28]. Besides, erythema and skin sensitivity in gravity-dependent areas have been reported post-flight in one astronaut who stayed in space for almost one year [
29]. Furthermore, thinning of the epidermis and decreased melanin concentration have been observed [
30]. On the contrary, improvement of hydration and barrier function as well as an unchanged skin density and thickness have also been reported [
31]. This indicated the variety in individual skin responses during spaceflight and highlights the need to further investigate skin responses to the spaceflight environment with larger sample sizes.
Unfortunately, access to astronaut samples is limited and in orbit experiments are costly and restricted due to logistical issues. To overcome these issues,
in vivo and
in vitro simulation models can be used to mimic certain aspects of the spaceflight environment and investigate their effects on different physiological systems. To simulate the effects of microgravity on cells, several ground-based facilities are available, such as clinostats, the RPM, and magnetic levitation [
32]. The RPM rotates with random velocity and direction around three axes, thereby averaging the total gravity vector experienced by the cells to values below 0.003
g [
33,
34]. To mimic certain aspects of radiation encountered in space, high-energy ion beams obtained at accelerator facilities can provide insights into the biological effect of exposure to cosmic radiation [
35]. Finally, biological effects related to chronic stress can be investigated by inducing elevated levels of glucocorticoids.
In vitro, this is often achieved by administrating soluble glucocorticoids to the cell culture medium.
As reviewed in Radstake et al. [
27], each of these spaceflight stressors alone can influence the proper functioning of cells and harm the integrity of the skin. Microgravity can affect the mechanosensitive structures of the cell, thereby inducing changes in the cells’ morphology and function. Gravitational unloading during spaceflight results in numerous cell-type-dependent alterations including alterations in cell proliferation, differentiation, expression of signaling molecules as well as gene expression [
36,
37]. Exposure to ionizing radiation can affect the function of fibroblasts by inducing DNA damage, apoptosis, and inflammation, and by altering gene expression, cell proliferation, and differentiation [
38,
39,
40,
41,
42,
43]. Finally, the effect of glucocorticoids on cells is regulated by the glucocorticoid receptor, which is a ligand-dependent transcription factor. Different isoforms of the glucocorticoid receptor differently regulate gene transcription, and therefore, the cellular response to glucocorticoids can vary largely among individuals, but also within different tissues at different phases of the cell cycle [
44,
45].
It is still poorly understood how these spaceflight stressors may interact and how such interactions may affect the skin. Furthermore, only a handful of studies have examined how the spaceflight environment affects the complex multi-phase process of cutaneous wound healing [
46,
47,
48,
49]. To address these issues, this work investigates the effect of combined exposure to simulated microgravity, ionizing radiation, and psychological stress on fibroblasts’ function related to the wound healing process. To this aim, we used an
in vitro wound healing assay of human primary dermal fibroblasts to study the expression of inflammatory cytokines and growth factors, relevant for the inflammatory phase of wound healing. Next, migration capacity and cytoskeletal reorganization, which are crucial for the proliferation phase, as well as expression of the dermal matrix proteins needed to remodel the skin tissue after wounding, were investigated.
2. Materials and Methods
2.1. Fibroblast Culture
Primary normal human dermal fibroblasts (NHDF) obtained from one donor (33-year-old Caucasian female) were purchased from PromoCell (C-12302) and cultured in Dulbecco’s Modified Eagle Medium containing GlutaMAXTM (DMEM, GIBCO, 10566016, Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS, GIBCO, 10500064, Fisher Scientific, Göteborg, Sweden) and 0.25% Penicillin-Streptomycin (Pen-Strep, Sigma-Aldrich, P4333, Merck KGaA, Darmstadt, Germany). Cells were incubated at 37 °C in a humidified atmosphere containing 5% CO2. Cells were passaged at 80–90% confluence using 0.05% Trypsin-EDTA (GIBCO, 25300062, Thermo Fisher Scientific, Waltham, MA, USA). All experiments were performed with asynchronized cells and at room temperature.
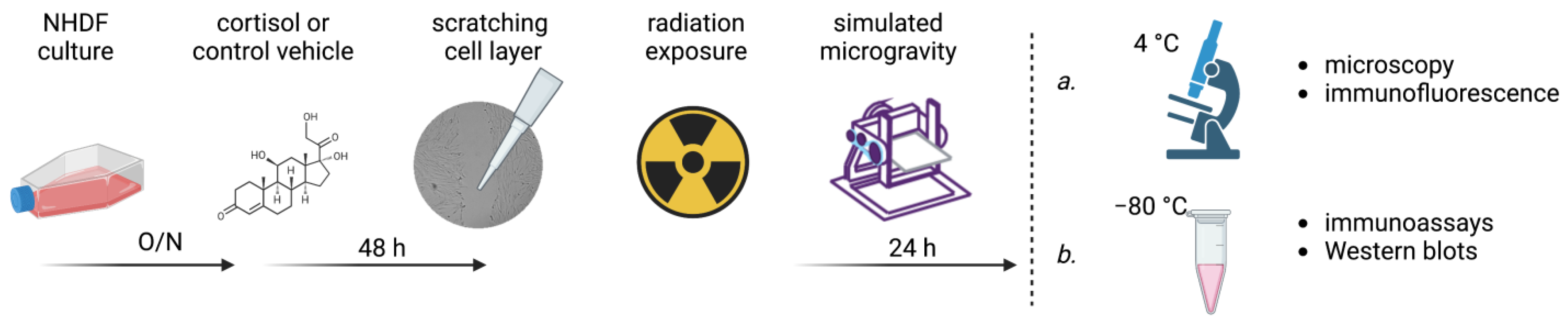
2.2. Experimental Procedures
Figure 1 shows a schematic representation of the experimental procedures. Cells were seeded inside SlideFlasks (Thermo Scientific Nunc Lab-Tek, 170920) or T12.5 flasks (Corning) at densities of ~10,000 cells/cm
2 using full serum DMEM and left to attach overnight. For each condition, a total of six replicates were used. Cells were then washed with phosphate-buffered saline (PBS) and incubated with DMEM containing hydrocortisone (HC, Sigma Aldrich, H0888, Merck KGaA, Darmstadt, Germany) at a concentration of 1 µmol/L or a control vehicle. This concentration was chosen as they are representative of circulating cortisol levels during sustained stress conditions and similar levels of cortisol have been measured in astronauts returning form space [
22,
25,
50,
51]. The hydrocortisone was first dissolved in 96% ethanol at a concentration of 1 mg/mL and further diluted in PBS to obtain a stock solution of 100 µmol/L hydrocortisone. The control vehicle consisted of 96% ethanol dissolved in PBS. Both solutions were then 1/100 diluted in the cell culture media. After 48 h of exposure to HC, cells were scratched using a bent 1 mL pipette tip and washed twice with PBS. Cell holders were then completely filled with CO
2-calibrated media containing either HC or the control vehicle, and airtight sealed using polymer integrated caps. Next, cells were irradiated with either X-rays, protons, carbon ions, or iron ions (see
Section 2.3 for an overview of different radiation facilities and parameters) at doses of 0.1, 0.5, and 1 Gy, to compare between relatively low space-relevant doses [
52] and a higher dose to test for dose-specific effects, or sham-irradiated (an overview of irradiation parameters is provided in
Table 1). After irradiation, cells were placed on an RPM or in a 1
g environment as controls for 24 h. Flasks that developed air bubbles while they were placed on the RPM were discarded. Afterwards, cells in SlideFlasks were rinsed with PBS and fixed using 10% Formalin solution (Sigma Aldrich, HT5014, Merck KGaA, Darmstadt, Germany).
Cells grown in T12.5 flasks were used for collection of cell lysates. In short, cells were dissociated using 0.25% Trypsin-EDTA (GIBCO, 25200072, Thermo Fisher Scientific, Waltham, MA, USA) and pellets were collected and washed using ice-cold PBS, after which they were re-suspended in ice-cold radioimmunopreciptation assay (RIPA) buffer (Pierce, 89901, Thermo Fisher Scientific, Waltham, MA, USA) containing Halt protease and phosphatase inhibitor cocktail (Thermo Scientific, 78440). Tissue Lyser II (Qiagen) at 30.00/s was used for 2 min, after which cells lysates were centrifuged at 14,000× g for 10 min at 4 °C to pellet down the cell debris and supernatant was collected. Cell lysates were stored at −80 °C.
2.3. Exposure to Ionizing Radiation
Fibroblasts were exposed to several radiation qualities including photons, protons, carbon ions, and iron ions at different radiation facilities in Europe. An overview of the radiation qualities, mean linear energy transfer (LET) values, and the microdosimetric quantities for the different radiation exposures can be found in
Table 1. The quantities listed in
Table 1 were obtained by means of computer simulations (see
Supplementary Materials) with PHITS [
53].
Cells were exposed to H250 X-rays at the Laboratory of Nuclear Calibration of the Belgian Nuclear Research Centre (SCK CEN) in Mol, Belgium. Flasks were placed on a Plexiglass plate in a horizontal position and irradiated with H250 X-rays from the top at 50 cm distance. The photon energy spectrum of the H250 X-rays can be found in the
Supplementary Materials. The exposures to carbon ions (90 MeV/n) and protons (150 MeV) were carried out at the Particle Therapy Research Center (PARTEC) facility in Groningen, The Netherlands. Samples were irradiated through the bottom of the culture vessel positioned vertically in the plateau of the Bragg curve. Irradiations were performed with a scanned beam with a homogeneous fluence. During proton irradiation, dose build-up was achieved using an 18 mm polycarbonate plate. Dose rates of approximately 0.5 Gy/min were used. Finally, fibroblasts were exposed to 1 GeV/n iron ions at the GSI-FAIR facility in Darmstadt, Germany. Samples were irradiated through the bottom of the culture vessel positioned in a vertical position with a scanned pencil beam with a homogeneous fluence in the plateau of the Bragg curve. For iron ions, a dose of 1 Gy corresponds to a fluence of 4 × 10
6 ions/cm
2.
2.4. Exposure to Simulated Microgravity
After irradiation, cells were placed on the RPM to expose them to simulated microgravity. Cell culture flasks were completely filled with medium and airtight sealed using caps filled with a polymer (SYLGARD 184 Silicon Elastomer, Dow, 01673921). Cells were exposed to simulated microgravity for 24 h. To minimize the effects of parasitic accelerations and shear stress acting upon the cells, moderate velocity of average 60 deg/s was chosen. In addition, cells were placed within 10 cm from the center of rotation.
2.5. Cytokine and Growth-Factor Synthesis
Multiplex immunoassays (LXSAHM-03, R&D Systems, Mineapolis, MN, USA) were used to investigate fibroblast expression levels of interleukin-6 (IL-6), interleukin-1 receptor antagonist (IL-1RA), and platelet-derived growth factor alpha (PDGF-α) in cell lysates following the manufacturer’s instructions. The Luminex MAGPIX system with xPONENT 4.3 software (Luminex Corporation, Austin, TX, USA. A DiaSorin Company) instrument was used for analyses of the multiplex immunoassays (using Belysa ® Immunoassay Curve Fitting Software (Merck KGaA, Darmstadt, Germany)). The ELISA assay (Human TGF beta 1 Elisa kit, ab108912, Abcam, Cambridge, UK) was further used for determining levels of transforming growth factor beta 1 (TGF-β1). Absorbance at 450 nm was measured using a 96 Plate Reader (Enzo Life Sciences, Inc., East Farmingdale, NY, USA) and additional background subtraction was done at 570 nm (analysis with Byonoy Software, Byonoy GmbH). Total protein quantification was done with a bicinchoninic acid assay (Sigma Aldrich, Merck KGaA, Darmstadt, Germany) and these data were used for normalization of the data.
2.6. In Vitro Scratch Wound Assay
Confluent cell monolayers were scratched after 48 h of cortisol incubation using a 1 mL bended pipette tip. Afterwards, the scratch was observed under a Leica microscope with a 5× objective and baseline images were captured at three different locations per scratch. To limit the effect of proliferation on wound closure, cells were incubated with medium containing 1% FBS after scratching. After fixation, cells were again imaged at the same location. Images were analyzed to determine open wound area using Matlab (R2021b, The Mathworks Inc., Natick, MA, USA). A high-throughput microscopy wound healing tool [
54] was used for generating a mask threshold. Masked images were then inspected and faulty masks were excluded from the data. Relative wound closure was calculated using the following equation:
where
t(0) is the pixel count in the wound area directly after scratching and
t(24) is the pixel count in the wound area 24 h after irradiation.
Finally, obtained values were then normalized to the 1g, 0 Gy controls without cortisol to obtain relative migration measures.
2.7. Cyotskeletal Remodeling
For each condition, three out of six fixed scratched samples were used for immunocytochemical visualization of actin stress fibers and vinculin focal adhesion complexes. Fixed cells were incubated in PBS containing 0.1% Triton-X 100 and 3% bovine serum albumin (BSA, A2153n Sigma-Aldrich, Merck KGaA, Darmstadt, Germany) for 5 min at room temperature, after which they were washed with Tris-buffered saline containing 0.1% Tween (TBS-T). For blocking, a solution of 5% goat serum in Tris-NaCl-blocking buffer (TNB) was used for one hour at room temperature. Primary antibody (mouse monoclonal anti-vinculin (7f9), Santa Cruz, sc-73614) at dilution of 1/500 in TNB was incubated overnight at 4 °C. Samples were washed with TBS and incubated with TNB containing secondary antibody (Alexa Fluor goat-anti-mouse 488, Invitrogen A11001, 1/500) and Phalloidin 594 TRITC (Invitrogen A12381) for actin stress fibers. Slides were mounted with mounting medium containing DAPI (Molecular Probes Prolong Diamond Antifade Mountant, p36962). A Nikon Ti Eclipse inverted widefield fluorescence microscope (Nikon Instruments) with a 60× objective and immersion oil and connected to a Prime BSI sCMOS camera was used to visualize the cellular components of isolated cells that had migrated into the open wound area. Z-stacks of 11 images were taken at 0.5 µm apart.
Image Analysis
Fiji (v1.53C,
https://fiji.sc) was used for image processing. All images were summed across the
z-axis.
Focal adhesions: Preprocessing steps for images of vinculin included background subtraction with rolling ball radius of 10 pixels. Afterwards, images were further processed using a median filter (radius of 6 pixels) to suppress somatic vinculin signal followed by a top-hat filter (using a disk with radius 6). Images were then thresholded following the Otsu algorithm. Finally, to determine the number of focal adhesions per images, spots larger than 75 pixels were counted per image. This threshold was determined to select mature focal adhesions that are associated with the cytoskeleton [
55].
Actin area and number of stress fibers: Actin images were thresholded following the Triangle algorithm, after which the total area per image was measured. For determination of the number of actin stress fibers, actin images were first background subtracted with a rolling ball with a radius of 10 pixels followed by a fast Fourier transformation (FFT) and low-frequency filtering, after which an inverse FFT was applied followed by Gaussian filter (σ = 1 pixel). Finally, images were thresholded following the Otsu algorithm. Per image, the total number of stress fibers was determined by identifying particles larger than 1000 pixels with circularity between 0 and 0.1.
Nuclei: Per image (224 × 224 micron), the total number of nuclei was determined by thresholding DAPI signal images following the Triangle algorithm and watershed to separate bordering nuclei.
The number of nuclei was used to determine the total actin area and actin stress fibers per cell as well as the amount of vinculin focal adhesion spots per cell. These values were then normalized to the 1g, 0 Gy controls without cortisol to obtain measures of relative cell dimension, relative number of focal adhesions, and relative number of stress fibers per cell. Per condition, on average 170 nuclei were imaged.
2.8. Extracellular Matrix Protein Expression
Western blot assays were performed to determine the expression of extracellular matrix (ECM) proteins of procollagen type I as well as fibronectin. Lysates were collected as described in
Section 2.2. Total protein quantification was done with bicinchoninic acid assay (Sigma Aldrich, BCA1, Merck KGaA, Darmstadt, Germany). MilliQ was used to dilute cell pellets and obtain a concentration of 0.33 µg/µL of protein per sample. A 4× Laemli buffer (1610747, BIO-RAD Laboratories) with β-mercaptoethanol (1/10) was added to the protein samples, followed by incubation at 95 °C for 10 min for samples used to measure expression of fibronectin. Per lane, 5 µg of protein samples were loaded onto a 4–15% Criterion TGX Stain-Free Precast Gel (5671085, BIO-RAD Laboratories) and transferred to a nitrocellulose membrane. Blocking was performed in 5% milk powder in TBS-T for 2 h at room temperature. Primary antibodies diluted in blocking buffer (rabbit polyclonal to collagen type I (1/1000, Abcam, ab34710) for collagen type I, and mouse monoclonal anti-fibronectin (1/1000, Sigma-Aldrich, F0916) for fibronectin) were incubated overnight at 4 °C. Additionally, primary antibodies for mouse monoclonal to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were added (1/10,000, Abcam, ab8425), which served as loading control. Membranes were then washed with TBS-T and incubated with secondary horseradish peroxidase (HRP)-conjugated antibodies diluted in blocking buffer (1/2000 for collagen type I, 1/1000 for fibronectin and 1/1000 for GAPDH). Enhanced chemiluminescence was used for the detection of HRP conjugates (1705061, BIO-RAD clarity kit used according to the manufacturer’s instructions). Blots were imaged using the Fusion FX (Vilber).
Protein bands for the target proteins were measured using the Bio1D (v15.06, Vilber Lourmat). Each band was normalized to the loading and internal control. Measures were then normalized to the 1g, 0 Gy controls without cortisol to obtain relative measures.
2.9. Statistical Analyses
General linear regression (assuming a Gaussian distribution) and data plotting was performed in R version 4.0.1 [
56] to test for the main and interaction effects of the independent variables stress, gravity, dose, and radiation quality. An overview of these factors can be found in
Table 2. For each experimental condition, the data were normalized to the 1
g, 0 Gy, control without cortisol to obtain relative values. The data were checked for outliers; data points 1.5 times the interquartile range above the upper quartile and below the lower quartile were removed from the data. Models were then tested for significant interaction effects of independent variables. If interaction effects were not significant, they were removed from the model. Akaike information criterion (AIC) values were considered as measure of the goodness-of-fit of the model. The residual errors were taken into account to guarantee unbiased estimates.
4. Discussion
In this paper, an in vitro simulation of the spaceflight environment has been used to investigate the effect on the fibroblasts’ wound healing capacity. The effect of simulated microgravity, ionizing radiation of several radiation qualities and at different doses, and stress hormones was investigated for each spaceflight stressor individually and in combination. Different functions of fibroblasts related to the wound healing process have been studied, covering the inflammation, proliferation, and remodeling phases. In the following sections, we will discuss how the different spaceflight stressors may interact and act upon the wound healing process.
4.1. Interaction of Simulated Microgravity and Cortisol during the Inflammatory Phase of Wound Healing
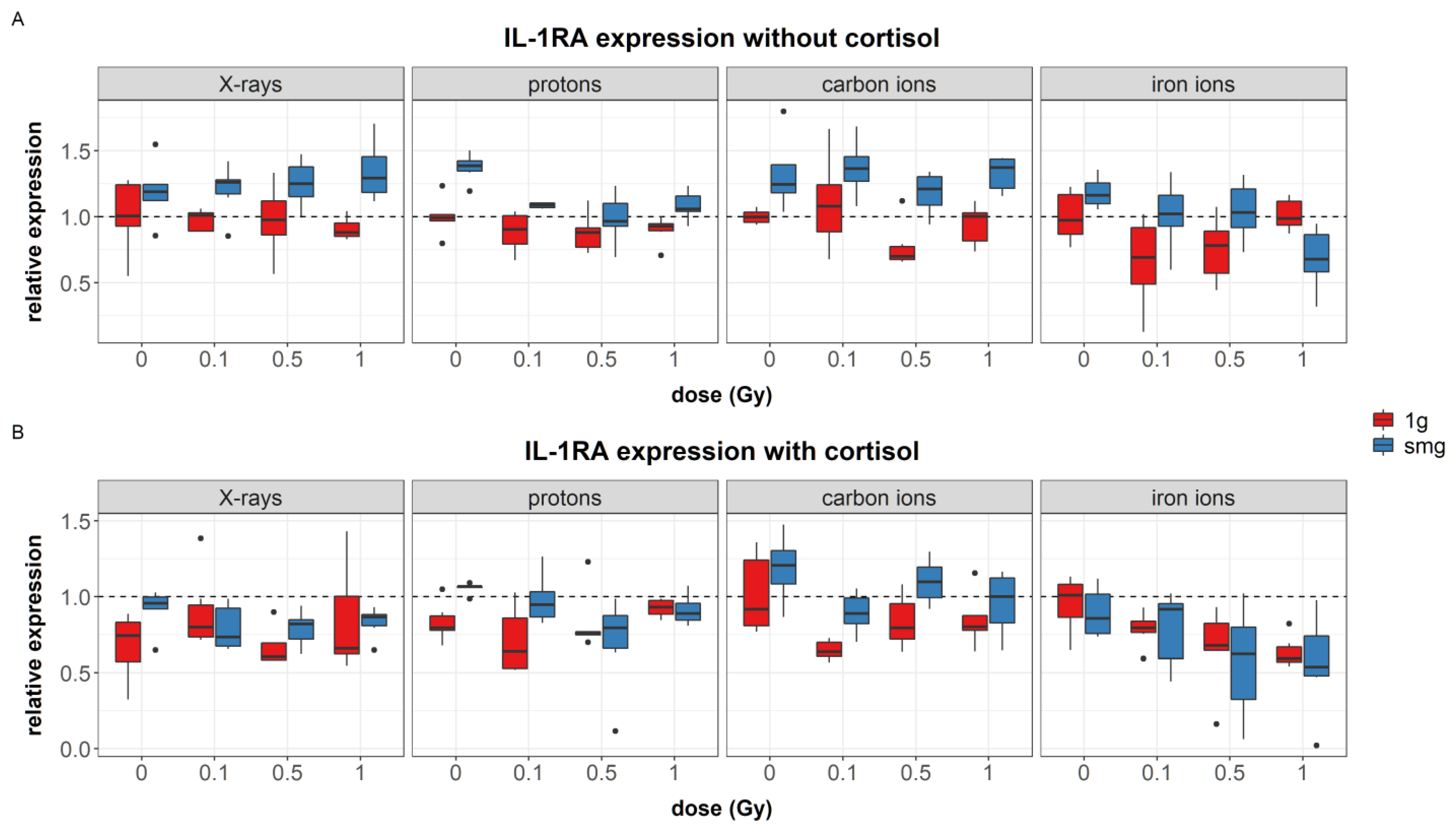
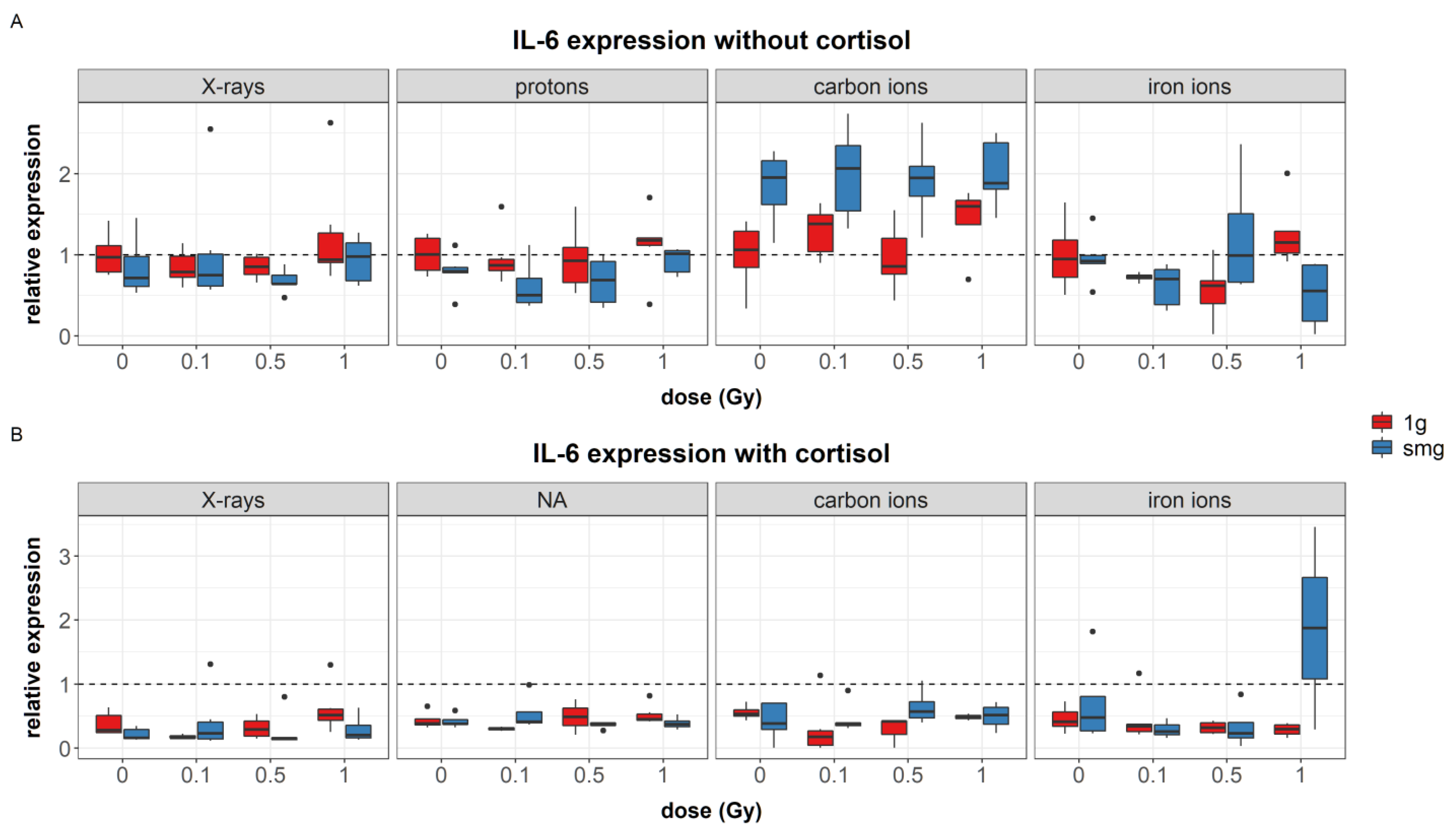
4.1.1. Simulated Microgravity Effects
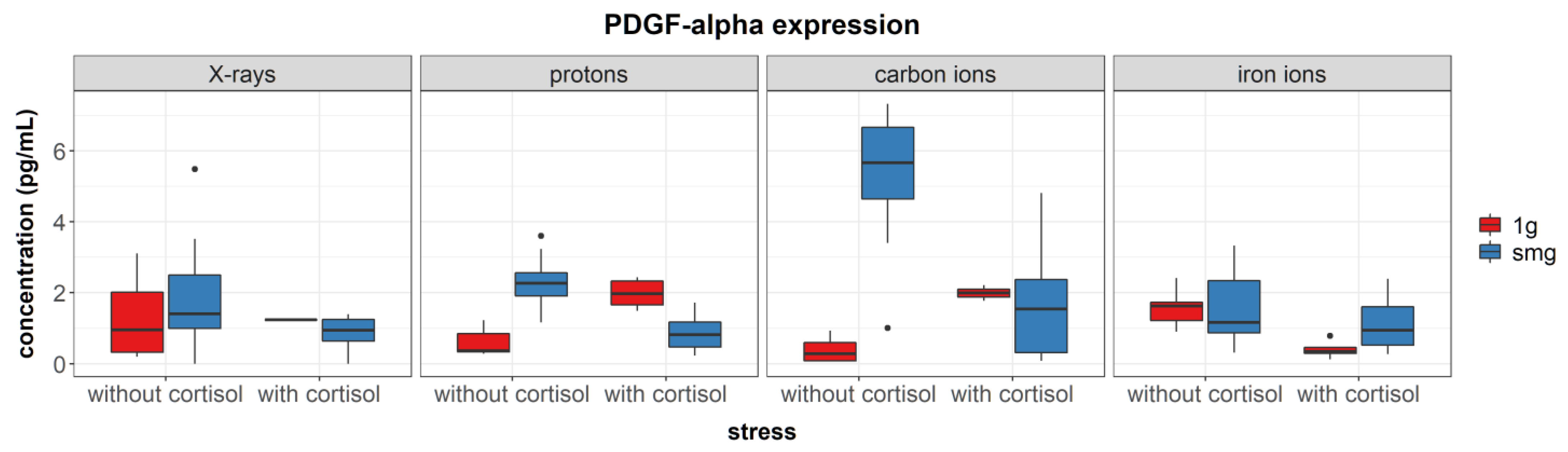
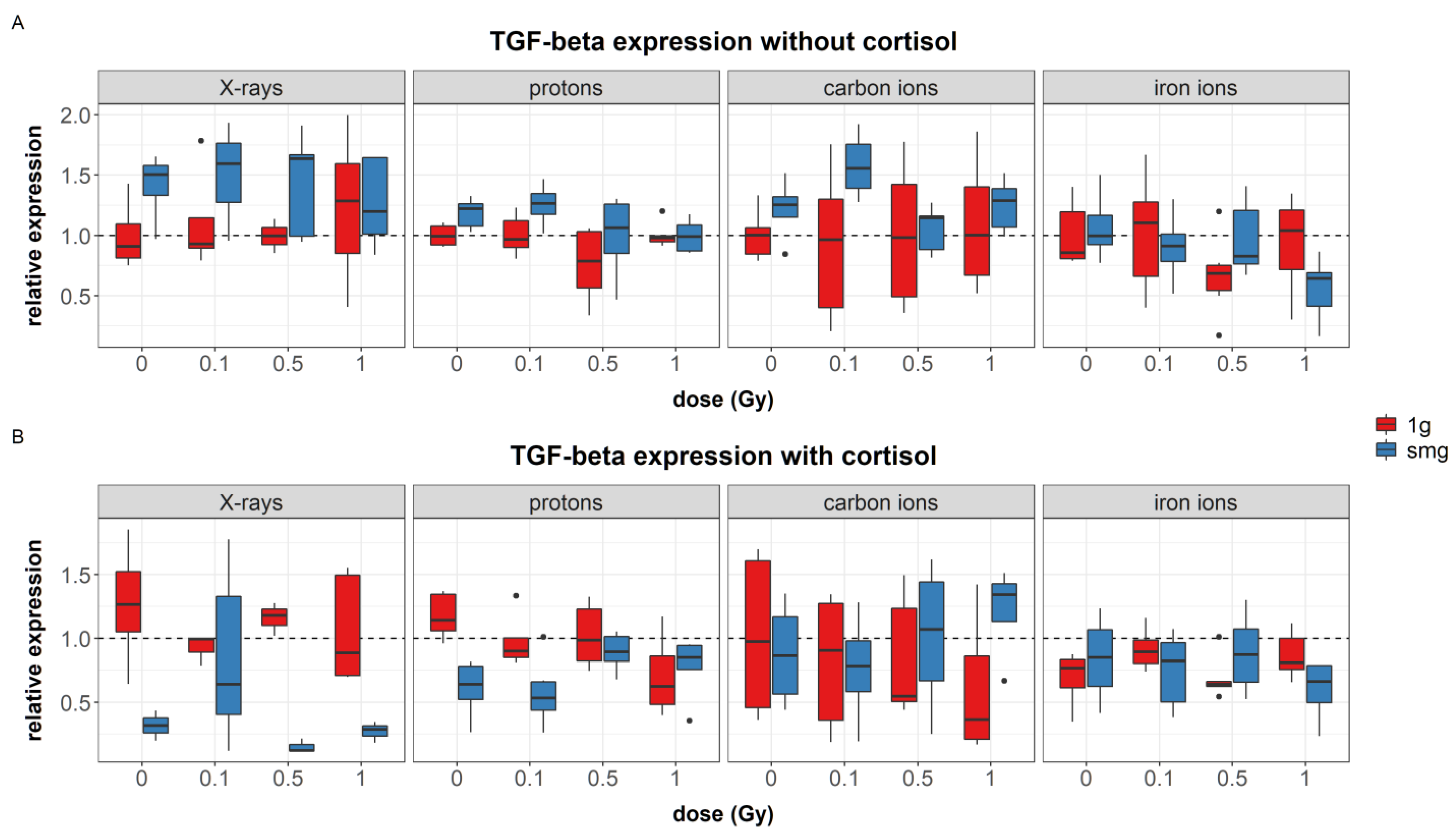
Simulated microgravity upregulated the expression of the growth factors PDGF-α (during experiments with carbon ions) and TGF-β. Furthermore, upregulation of IL-6 was seen during carbon ion experiments and IL-1RA seemed to be upregulated after exposure to simulated microgravity as well; although this main effect was not significant. Increased levels of TGF-β1 gene expression and IL-6 concentrations have been identified in endothelial cells after exposure to simulated microgravity; although no measurable effect was observed in TGF-β1 concentrations [
65]. Furthermore, increased excretion of IL-6 has been found in astronauts’ urine samples during the first day of spaceflight [
66].
PDGF-α works as a chemoattractant and enhances the proliferation of fibroblasts during later phases of wound healing. Furthermore, it stimulates the expression of ECM proteins and contraction of the wound [
61]. As PDGF-α and TGF-β show a synergistic relationship [
67], it is not surprising to also find increased levels of TGF-β in fibroblasts exposed to simulated microgravity. Although TGF-β1 during wound healing is important for fibroblast differentiation and stimulation of ECM protein expression, its inflammatory effect in the skin has been related to delayed wound healing in transgenic mice [
68].
IL-6 also plays an important role during the wound healing process. It works as a mitogen compound and IL-6-deficient mice show impaired wound healing [
58]. High levels of IL-6 can also be found in dermal and epidermal cells of psoriatic plaques [
69]. Injection of IL-6 results in erythema and an infiltration of lymphocytes in the dermis, which is indicative of inflammatory changes as the result of high levels of IL-6 [
70]. Additionally, IL-1RA regulates the wound healing process at several phases. Its absence in knockout mice is linked to delayed healing, as shown by delayed granulation tissue formation and neovascularization. Furthermore, reduced levels of type I collagen gene expression were also indicated in these mice [
57]. As IL-1RA competitively binds to the IL-1 receptor, the balance between IL-1RA and IL-1 is important for skin health. An increased IL-1RA/IL-1 ratio has been found in sun-exposed skin as well as in biopsies taken from skin sites of patients with inflammatory cutaneous disorders [
71].
Although all aforementioned cytokines and growth-factors are crucial for proper wound healing, overexpression can be linked to inflammatory skin conditions [
69,
70,
71]. Moreover, despite their stimulating function for fibroblast migration, a significantly delayed migration of fibroblasts exposed to simulated microgravity was observed in this study. This suggests that the increased expression of cytokines and growth factors that were observed in simulated microgravity either dampens fibroblast migration, or alternatively, the upregulation of these cytokines and growth factors is not sufficient to promote cell migration under simulated microgravity conditions.
4.1.2. Combined Effects
Besides the increase in cytokine and growth factor expression in fibroblasts exposed to simulated microgravity, cortisol decreased the expression of IL-6 and IL-1RA. Moreover, when cells were exposed to simulated microgravity in combination with cortisol, the upregulation of cytokine and growth factors, as observed in simulated microgravity only, was not found. Likewise, in some conditions, exposure to ionizing radiation also lowered the expression levels in cells exposed to simulated microgravity, as for instance seen after 1 Gy of iron ion exposure where IL-1RA expression in fibroblasts exposed to simulated microgravity was significantly lower as compared to the other groups in simulated microgravity. On the contrary, after exposure to 1 Gy of iron ions in fibroblasts that have also been exposed to a combination of simulated microgravity and cortisol, a significant increase in IL-6 expression was observed compared to the other groups. This indicates that, although simulated microgravity may upregulate the expression of certain cytokines and growth factors, when the effect of other spaceflight stressors is taken into account, the expression levels may alter. Furthermore, the effect of ionizing radiation may differ depending on whether fibroblasts have been exposed to a set of spaceflight stressors or one spaceflight stressor in isolation, and ultimately, this effect can furthermore depend on the radiation quality (LET and ion).
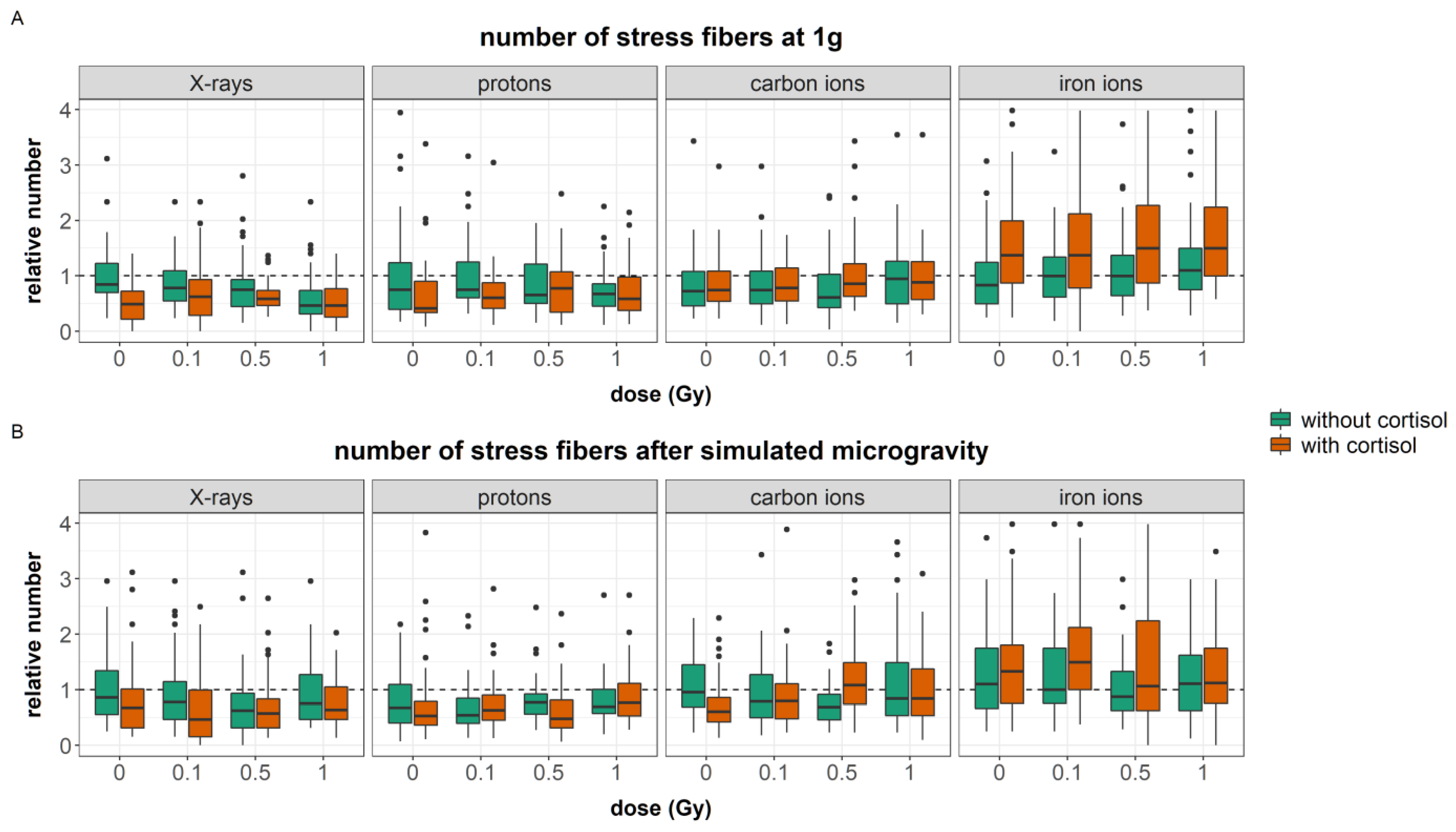
4.2. Fibroblast Migration and Cytoskeletal Remodeling Is Affected by Simulated Spaceflight Stressors
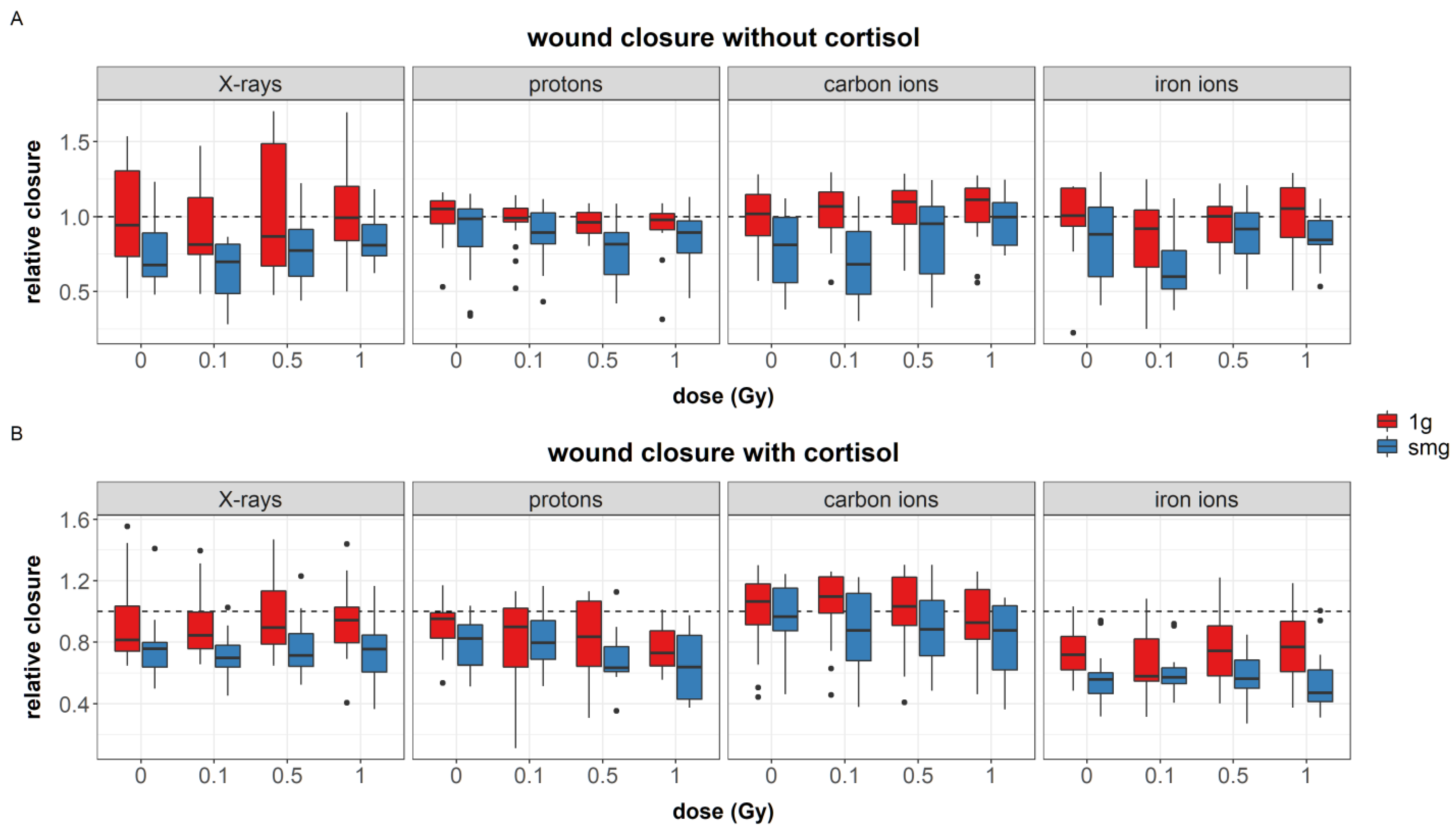
The effect of simulated microgravity on cell migration can be complex and even contradictory [
27]. In this paper, we have shown with a large dataset that simulated microgravity reduced the migration capacity of human dermal fibroblasts regardless of dose and cortisol exposure and for all experiments (
Figure 7). Simulated microgravity alone did not significantly alter cellular area or the number of focal adhesions or stress fibers. This is in contradiction with the loss of stress fibers observed in mouse osteoblasts and human epidermoid cancer cells during actual spaceflight [
72,
73] and in human umbilical vein endothelial cells and human mesenchymal stem cells during simulated microgravity conditions using the RPM or rotating wall vessel system [
74,
75,
76,
77,
78,
79]. Likewise, reduced motility of J-111 monocytes during spaceflight was linked to a disruption of actin fibers [
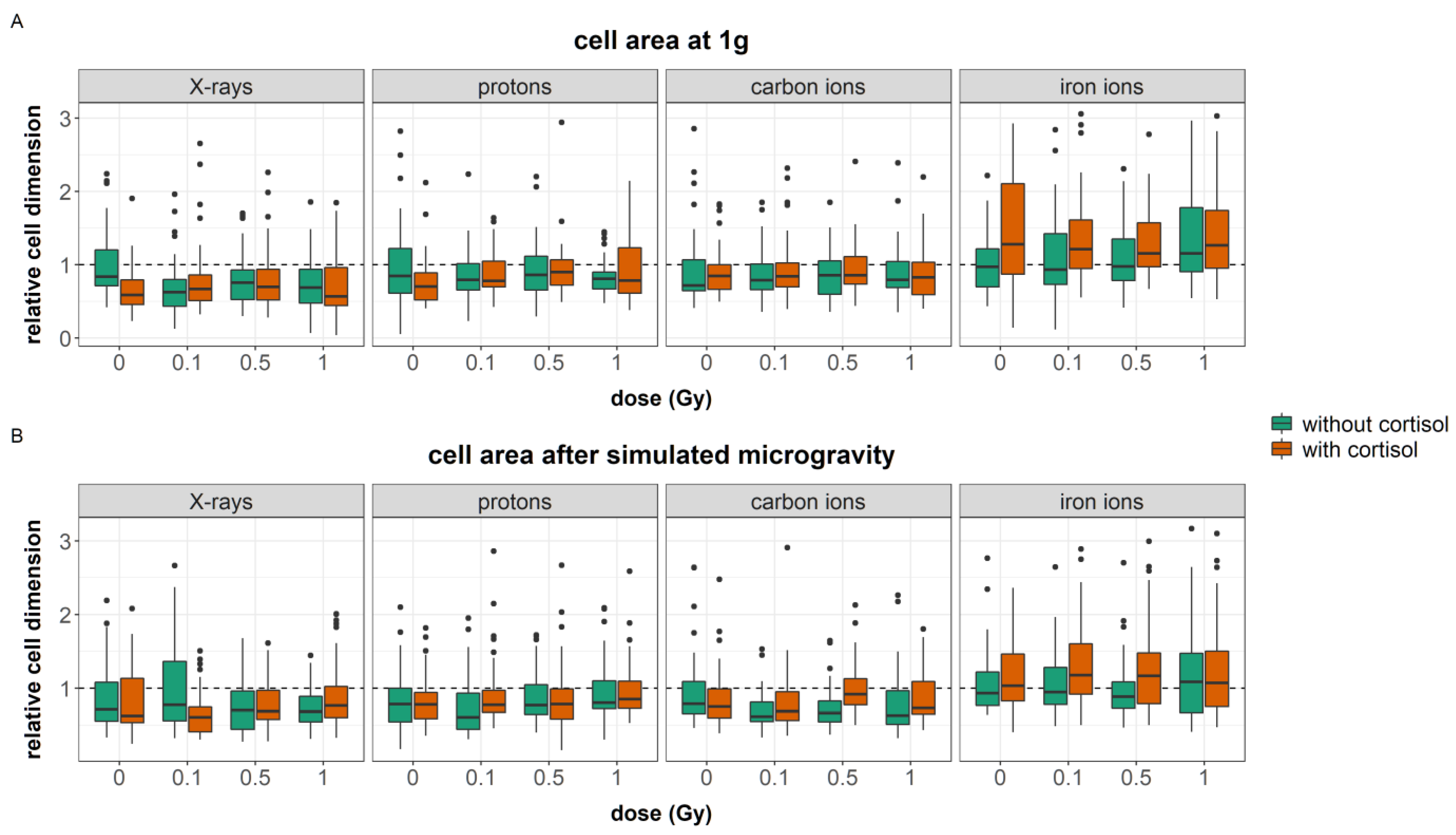
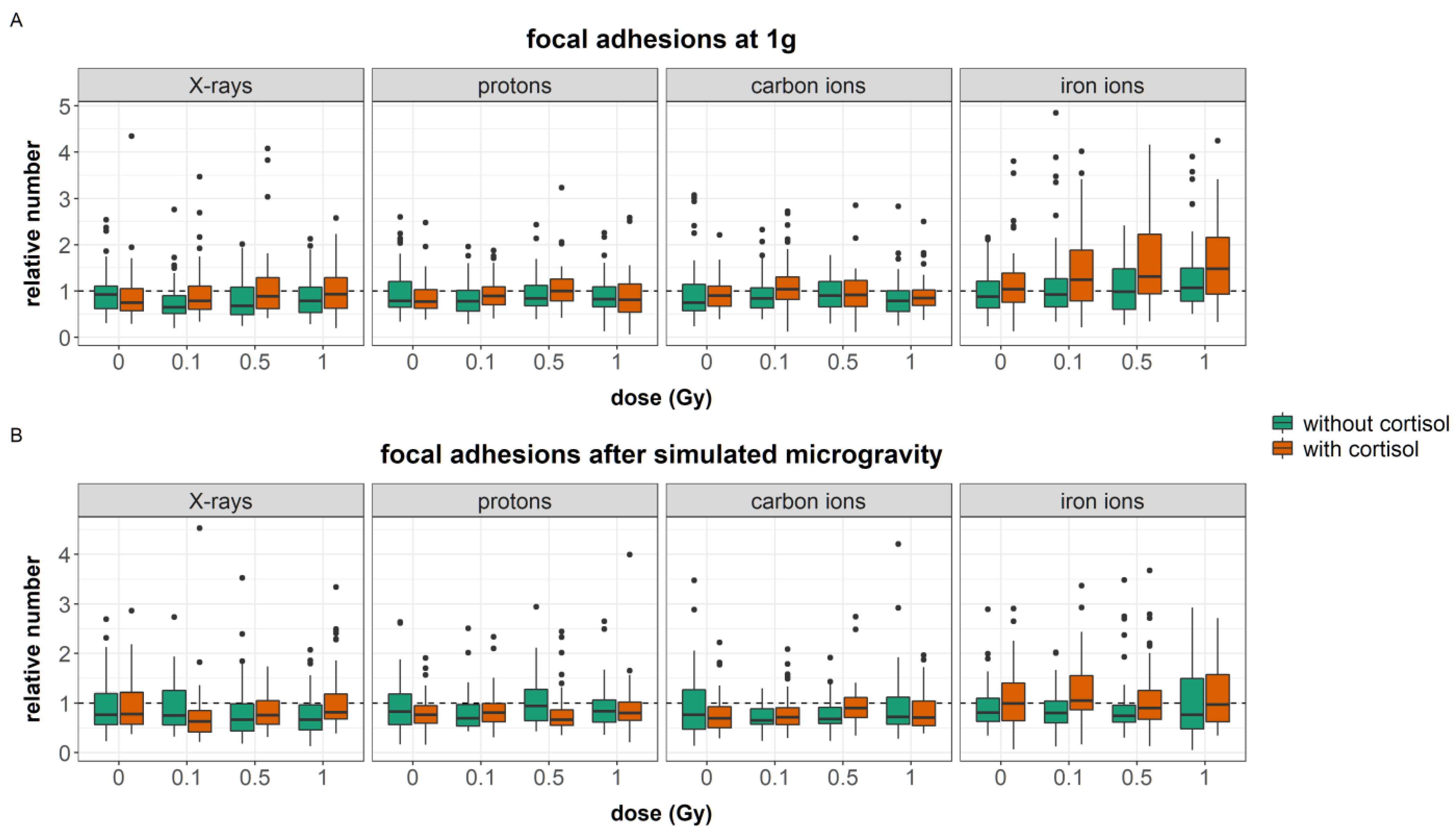
80]. Nevertheless, in our study, simulated microgravity interacted with cortisol and ionizing radiation at several endpoints. Cortisol exposure increased the number of focal adhesions. Furthermore, an increased cellular area was found in fibroblasts exposed to cortisol as compared to unexposed controls during carbon and iron ion experiments. Both cortisol-induced effects were not observed after exposure to a combination of cortisol and simulated microgravity. Furthermore, the effects of ionizing radiation on both the number of stress fibers and focal adhesions per cell were attenuated by simulated microgravity as well. This indicates that, although observations may show no main effect of simulated microgravity on cytoskeletal remodeling, the spaceflight stressor can interact with and influence the effect of other spaceflight stressors.
Similar to simulated microgravity, cortisol also reduced the migration capacity of fibroblasts, yet this effect was not always significant. This might be due to the dynamic aspect of fibroblast migration and the experimental design that was used in this study, where the observation of wound closure is only done at one specific time point. This notion is further supported by the data shown in Radstake et al. [
81], where measuring the migration of fibroblasts over a longer period of time shows an initial delay in migration, while at later time points, migration capacity and wound closure is recovered.
The cortisol-induced delayed migration was best observed during experiments with iron ions. During these experiments, fibroblasts had a larger cellular area, an increased number of stress fibers, and a reduced number of focal adhesions. The latter endpoint significantly increased with dose as a result of exposure to both cortisol and iron ions. Actin stress fibers are crucial for the contractility of cells needed for motility and proper migration [
62]. Cortisol has the ability to interact with and remodel the actin cytoskeleton, resulting in increased thickness and stability after exposure to glucocorticoids [
82]. Alterations in the number of actin stress fibers are linked to cellular stiffness, and a loss of actin stress fibers is linked to reduced cellular stiffness and migration capacity [
83]. Focal adhesions link actin stress fibers to the surrounding ECM. During migration, a remodeling of stress fibers and disassembly of focal adhesions is needed for the retraction of the rear part in migrating cells [
62]. However, increased cytoskeletal stiffening, as observed with increased number of stress fibers, leads to a strengthening of focal adhesions [
84].
In the results presented in this paper, the increased number of actin stress fibers and vinculin spots as a result of exposure to cortisol either with or without exposure to ionizing radiation as observed during iron ion experiments was linked to a reduced migration. This suggests that the increased number of stress fibers resulting from cortisol exposure, together with increased focal adhesion after exposure to cortisol in combination with iron ions as well as the increased area of the cells, are linked to reduced migratory capacity possibly due to the increased adhesion of cells to the substrate. This supports the notion of a “sweet-spot” of the number of actin fibers where dynamic remodeling of the actin cytoskeleton supports cellular motility. However, an increase in the number of fibers may fix cells to the substratum and limit their motility, especially when cellular dimensions are larger and an increased number of focal adhesions is present as well. The difference between the experiments suggests that the effect of cortisol on migration and number of actin stress fibers is not chronic, and timing differences may exist, as also observed in migration behavior.
Additionally, in this paper, a reduced expression of IL-6 was observed after cortisol exposure. IL-6 has been indicated to stimulate migration in smooth muscle cells by inducing cytoskeletal reorganization and induction of F-actin stress fibers [
85]. The reduced migration found after cortisol exposure, together with lower levels of IL-6, observed cytoskeletal differences between cortisol exposed cells, and unexposed controls further supports the notion of the effect of IL-6 on cytoskeletal rearrangements needed for migration.
Finally, exposure to 1 Gy of protons led to a reduced migration and lower expression of vinculin spots in fibroblasts regardless of exposure to other simulated spaceflight stressors. Ionizing radiation has been shown to be able to affect the migration capacity of cells, and both an increase and decrease can be observed after exposure to ionizing radiation. Moreover, ionizing radiation effects on cell migration are dependent on cell type, total dose, as well as timing (as reviewed in Verde et al. [
86]). In transformed fibroblasts, 1 Gy of X-ray exposure inhibited cell migration, which was explained by radiation-induced changes in cytoskeletal structure. However, in healthy cell lines, this effect was only observed at higher doses [
87]. Based on the results in this paper, ionizing radiation exposure leads to changes in the number of actin filaments and vinculin spots; although the effect depends on the radiation quality. Moreover, an effect of radiation quality on the cell dimension could be observed as well, where 1 Gy exposure to photons led to smaller cell bodies. Interestingly, exposure to 1 Gy of iron ions led to an increase in cellular dimension. However, this did not lead to observable changes in migration capacity for all groups. This suggests that, although ionizing radiation can induce changes in cytoskeletal architecture, which could lead to observable changes in cell migration, at doses relevant for space, the induced changes are reversible and not strong enough to affect the migration of fibroblasts, which was more strongly affected by other spaceflight stressors.
4.3. Simulated Spaceflight Stressors Interact and Affect the Expression of Dermal Matrix Proteins in Fibroblasts
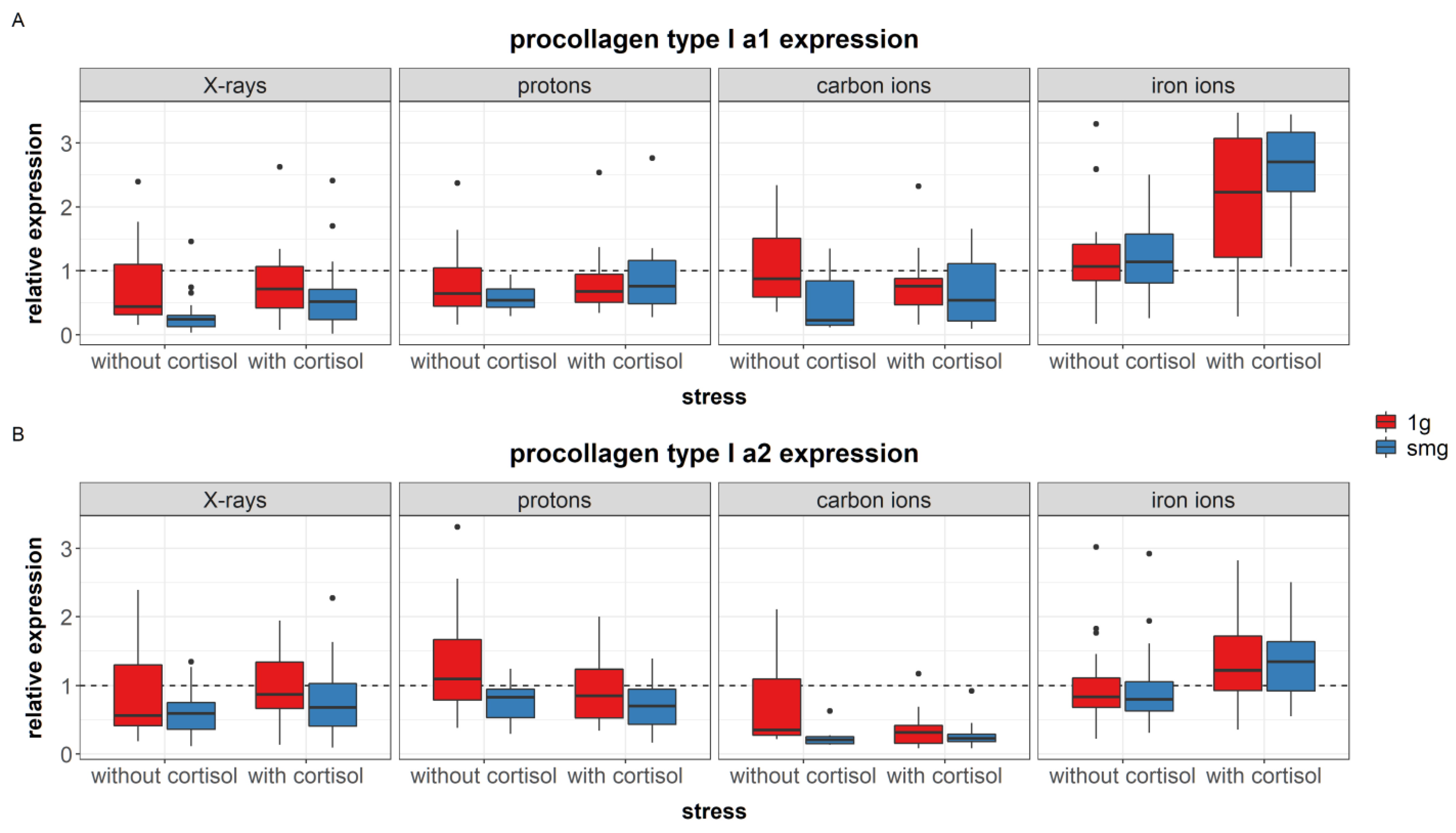
4.3.1. Simulated Microgravity Effects
Simulated microgravity downregulated the expression of fibronectin and procollagen type I in fibroblasts. This is in accordance with the literature where exposure to simulated microgravity reduced gene expression levels of fibronectin in fibroblasts after RPM exposure [
88], as well as type I collagen protein expression in osteoblasts (rotating wall vessel) [
89] and fibroblasts after RPM exposure [
48]. However, an upregulation of fibronectin can also be found in fibroblasts after simulated microgravity exposure using the RPM [
48]. On the contrary, during experiments with iron ions, an upregulation of procollagen type I α1 was found after simulated microgravity exposure. The 3D co-cultures of fibroblasts and keratinocytes have indicated an upregulation of type I procollagen α1 after exposure to simulated microgravity for 7 days with the use of a 3D clinostat [
90]. Furthermore, transcriptomic analyses of skin tissue from space-flown mice show increased expression levels of procollagen compared to ground controls [
91], and increased collagen content has also been found in astronauts after spaceflight [
30]. This indicates that type I procollagen is sensitive to simulated microgravity; although, its effect can differ depending on the complexity of the
in vitro and
in vivo model. Furthermore, contradictory results may be explained by slight differences in timing of exposures or sample collection after simulated microgravity exposure. More data at different time points would be needed to better understand the time-dependent response of fibroblasts procollagen type I expression under simulated microgravity. Furthermore, although expression levels of procollagens can be affected, it cannot be concluded from the results presented in this study how this affects collagen fiber formation and deposition in the ECM. Moreover, although excretion of procollagens can be increased as result of spaceflight, ECM degradation can still be observed, possibly due to excessive degradation of the newly formed procollagen [
91].
4.3.2. Combined Effects
In fibroblasts exposed to cortisol, fibronectin expression was significantly higher compared to controls. This finding is in agreement with the literature, as other studies have also found that treatment of fibroblasts cultures with cortisol induced upregulation of fibronectin [
92,
93,
94]. Moreover, when fibroblasts were exposed to simulated microgravity in combination with cortisol, a dose-dependent effect could be observed with a significant increase in fibronectin expression at higher doses of ionizing radiation. Besides the upregulation of fibronectin, in the case of iron ion experiments, procollagen type I was also upregulated in fibroblasts exposed to simulated microgravity and/or cortisol (although no dose-dependent effects could be established). High levels of fibronectin as well as collagen type I are linked to the formation of keloids and hypertrophic scars and are an indication of abnormal wound healing [
92,
95]. Expression of growth-factors TGF-β1 and PDGF-α induce the expression of fibronectin during wound healing [
60,
61,
96]. However, the dose-dependent upregulation of fibronectin in fibroblasts exposed to simulated microgravity and cortisol in these experiments was not linked to an increased expression of these growth factors and could, therefore, not be explained based on current data. More research is, therefore, needed to better understand these observations. Nevertheless, observation of the dose-dependent increase in fibronectin expression, which was most clearly observed in fibroblasts exposed to both simulated microgravity and cortisol, suggests that the complete simulated spaceflight environment could influence matrix deposition during wound healing, mostly when all three spaceflight stressors are taken into account. In turn, this could lead to increased matrix deposition and excessive scar formation as a result of exposure to the spaceflight environment.
4.4. Limitations and Future Directions
In this study, we have developed and tested an in vitro model of the spaceflight environment. After exposure of dermal fibroblasts to this simulated spaceflight environment, we observed findings such as altered expression of cytokines, cytoskeletal remodeling, and ECM protein expression, which are in line with findings from spaceflight studies. While the results in this paper help to improve our understanding on the interaction of different spaceflight stressors on the fibroblasts function during wound healing and provide a starting point for future experiments, some limitations remain, which should be addressed.
The chosen endpoints in this study represent some of the important functions of fibroblasts relevant for proper wound healing and skin integrity. However, the wound healing process consists of many components, and therefore, other parameters, which have not been considered in this study, can be studied in future experiments. The presented
in vitro model can be used to study other skin parameters, such as collagen type VII, a less abundant protein found in the skin, which, nevertheless, plays a crucial role in the skin’s integrity and wound healing [
97]. In addition, cytoskeletal structures of microtubuli and vimentin have previously been found to be altered during spaceflight [
98,
99,
100]. Other factors involved in the remodeling of the skin, such as matrix metalloproteinases, can also be considered [
101]. Therefore, it would be of interest to apply the described model of the simulated spaceflight environment to investigate other endpoints as well.
While the diverse role of skin fibroblasts during the wound healing process makes them a suitable
in vitro model for studying the effects of simulated spaceflight stressors on the wound healing process, this monoculture does not account for the complexity of the skin and the interaction between fibroblasts and other skin cells. The skin is a neuroendocrine system, which means that a complex interaction exists between the nervous system, endocrine system, and immune system. This complexity serves to maintain proper barrier function of the skin [
102]. As a result, fibroblasts’ function is regulated by a plethora of different cell types. For example, dysregulation of the immune system in microgravity might contribute to the increases in skin sensitivity as observed during spaceflight [
103,
104,
105]. Furthermore, generalization of the present results remains limited as fibroblasts were obtained from one donor. For these reasons, the present study does not account for inter-individual differences or skin complexity as found
in vivo. To explore these effects, the experiments could be repeated with cells obtained from multiple donors, and in more complex systems, such as organotypic skin cultures, skin-on-a-chip, and
in vivo models.
Another limitation of the applied methodology in this study is the use of the RPM to simulate the microgravity environment. While this method is a well-accepted method to study biological processes in cells that are dependent on the gravity vector, it comes, however, with constraints. Gravity artifacts may arise due to RPMs kinematic rotation, and hence, a moderate velocity and distance from the center of rotation should be considered [
106]. In addition, fluid motion inside the flasks due to the accelerations causes shear stress on cells attached to the vessel wall. Studies have shown that the amount of mechanical forces due to shear stress are higher on the RPM compared to clinostats, which differently affected cellular responses to the simulated microgravity environment [
107,
108]. While moderate velocity settings were chosen in this study, some effects of shear forces would remain and should be taken into account when interpreting these results. High shear stress influences fibroblasts arrangements and function related to wound healing and high shear stress has been related with increased migration speed in fibroblasts [
109]. Therefore, in future studies, to validate these results, the study should be repeated using real microgravity platforms.
With an eye on future interplanetary space missions, it is of great importance to develop effective countermeasures and protect astronauts against possible infections resulting from delayed wound healing. The development of wound dressings that promote healing and reduce risks of infections, but at the same time have a long shelf-life and take up little space, could hold promises to this aim. Of interest is the finding that treatment of wounds with platelet-rich plasma (PRP) in a wound healing model of the leech was shown to be successful in counteracting the delayed healing as result of simulated microgravity exposure [
46]. This model may be used to further test the effectiveness of PRP as a treatment to restore wound healing in simulated microgravity combined with radiation and cortisol exposure as well. In addition, it would be of interest to investigate if and how the skin-related spaceflight effects return back to normal once astronauts are back on Earth, or at partial gravity of the Moon and Mars. For these aims, the current
in vitro model could provide a methodology to study the effect of exposure to changing gravity fields on the
in vitro wound healing process. Finally, with the increase in both commercial and touristic spaceflight, efforts should be made to reduce the risks of obtaining skin injuries as a result of, for instance, friction induced by spacesuits.
5. Conclusions
To the best of our knowledge, this study is the first to combine simulated microgravity, ionizing radiation, and cortisol to simulate the spaceflight environment in vitro. Using this model, we have investigated how wound healing capacities of dermal fibroblasts were affected after exposure to this set of spaceflight stressors. It can be concluded from our study that the simulated spaceflight environment can affect fibroblast wound healing capacity at any phase during the wound healing process. Moreover, as shown by the interaction effect between simulated microgravity and cortisol, the effects of exposure to one single spaceflight stressor can be altered when other spaceflight stressors are considered as well. Finally, some spaceflight stressors, such as ionizing radiation, only showed effects when fibroblasts were exposed to simulated microgravity and cortisol as well, while the response could depend on the radiation quality.
The interaction of the different spaceflight stressors highlights the complexity that needs to be taken into account when studying the effect of spaceflight on certain biological processes. Furthermore, the wound healing process consists of a complex and delicate interaction between different cellular components and phases. Fibroblasts play an important role during the wound healing process and their sensitivity to exposure to simulated spaceflight stressors makes this important function of the skin especially vulnerable under spaceflight conditions. Countermeasures should be developed to reduce the risk of delayed and impaired healing, which challenges the barrier function of the skin and increases the risk of infections and health complications.


 ,
, 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}