
Conversion of Unmodified Stem Cells to Pacemaker Cells by Overexpression of Key Developmental Genes


Abstract
1. Introduction
2. Materials and Methods
2.1. Adipose-Derived Stem Cells
2.2. Lentivector Preparation
2.3. Lentiviral Production
2.4. Cell Culture
2.5. Immunohistochemistry (IHC)
2.6. Cell Sorting by Size
2.7. qRT-PCR
2.8. Patch Clamp Electrophysiology
3. Results
3.1. Single Factors TBX3 and TBX5 and Their Combinations with SHOX2 and HCN2 Quickly Induce ASC Differentiation
3.2. SHOX2-HCN2-TBX5 Is the Most Efficient Combination
3.3. SHOX2-HCN2-TBX5 Converted Cells Express Pacemaker Factors CX30.2 and HCN4
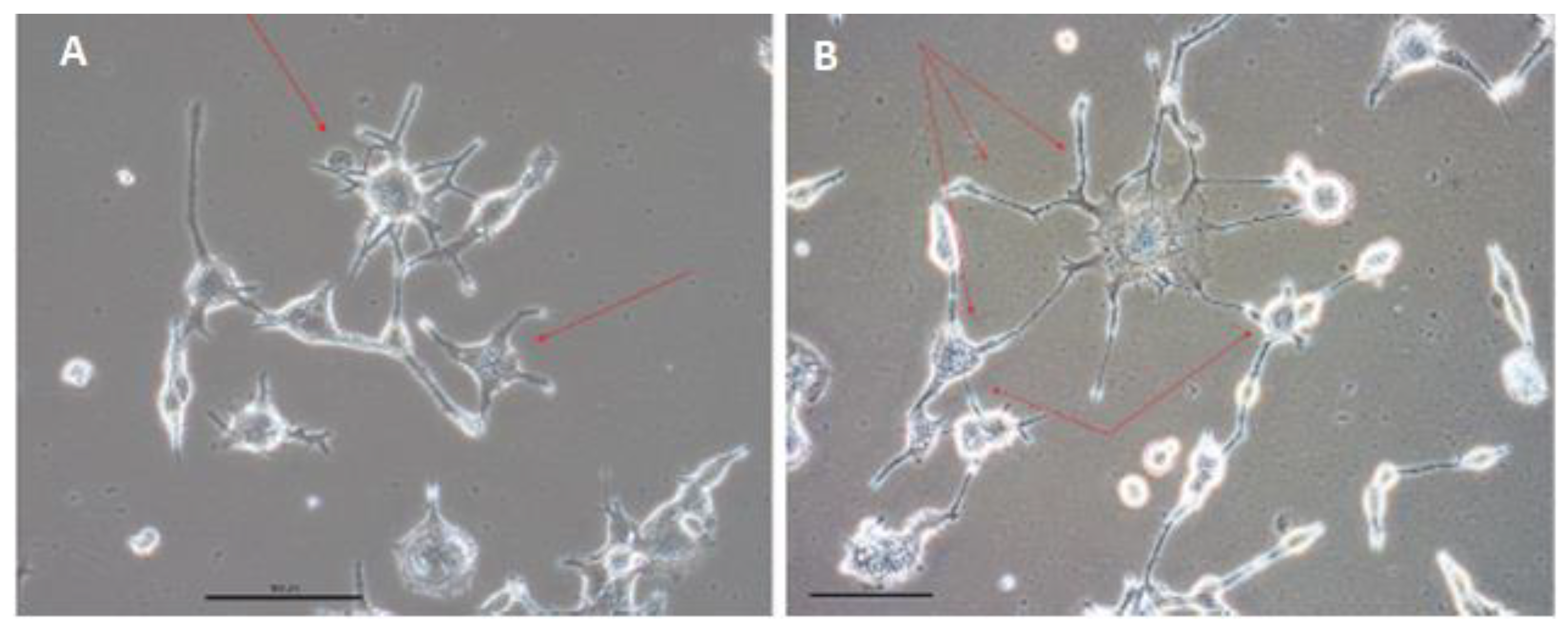
3.4. Two Types of Converted Cells Have Pacemaker-Like Characteristics
4. Discussion
4.1. Unmodified ADRCs Containing Multipotent ASCs May Be Used as Starting Material for Forward Programming of Pacemaker-Like Cells
4.2. Cell Identities
4.3. ASCs as the Cells of Choice for Pacemaker Programming
4.4. Choice of Gene Delivery Vehicle
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Glossary
References
- Christoffels, V.M.; Moorman, A.F. Development of the cardiac conduction system: Why are some regions of the heart more arrhythmogenic than others? Circ. Arrhythm. Electrophysiol. 2009, 2, 195–207. [Google Scholar] [CrossRef]
- Dobrzynski, H.; Boyet, M.R.; Anderson, R.H. New insights into pacemaker activity: Promoting understanding of sick sinus syndrome. Circulation 2007, 115, 1921–1932. [Google Scholar] [CrossRef]
- De Ponti, R.; Marazzato, J.; Bagliani, G.; Leonelli, F.M.; Padeletti, L. Sick sinus syndrome. Card. Electrophysiol. Clin. 2018, 10, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Madhavan, M.; Mulpuru, S.K.; McLeod, C.J.; Cha, Y.M.; Friedman, P.A. Advances and future directions in cardiac pacemakers: Part 2 of a 2-part series. J. Am. Coll. Cardiol. 2017, 69, 211–235. [Google Scholar] [CrossRef] [PubMed]
- Mulpuru, S.K.; Madhavan, M.; McLeod, C.J.; Cha, Y.M.; Friedman, P.A. Cardiac pacemakers: Function, troubleshooting, and management: Part 1 of a 2-part series. J. Am. Coll. Cardiol. 2017, 69, 189–210. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, E.; Goldhaber, J.I.; Marbán, E. Next-generation pacemakers: From small devices to biological pacemakers. Nat. Rev. Cardiol. 2018, 15, 139–150. [Google Scholar] [CrossRef]
- Husse, B.; Franz, W.M. Generation of cardiac pacemaker cells by programming and differentiation. Biochim. Biophys. Acta 2016, 1863, 1948–1952. [Google Scholar] [CrossRef]
- Arnold, S.J.; Robertson, E.J. Making a commitment: Cell lineage allocation and axis patterning in the early mouse embryo. Nat. Rev. Mol. Cell Biol. 2009, 10, 91–103. [Google Scholar] [CrossRef]
- Burridge, P.W.; Keller, G.; Gold, J.D.; Wu, J.C. Production of de novo cardiomyocytes: Human pluripotent stem cell differentiation and direct reprogramming. Cell Stem Cell 2012, 10, 16–28. [Google Scholar] [CrossRef]
- Espinoza-Lewis, R.A.; Yu, L.; He, F.; Liu, H.; Tang, R.; Shi, J.; Sun, X.; Martin, J.F.; Wang, D.; Yang, J.; et al. Shox2 is essential for the differentiation of cardiac pacemaker cells by repressing Nkx2-5. Dev. Biol. 2009, 327, 376–385. [Google Scholar] [CrossRef]
- Greulich, F.; Rudat, C.; Kispert, A. Mechanisms of T-box gene function in the developing heart. Cardiovasc. Res. 2011, 91, 212–222. [Google Scholar] [CrossRef]
- Munshi, N.V. Gene regulatory networks in cardiac conduction system development. Circ. Res. 2012, 110, 1525–1537. [Google Scholar] [CrossRef]
- Mori, A.D.; Zhu, Y.; Vahora, I.; Nieman, B.; Koshiba-Takeuchi, K.; Davidson, L.; Pizard, A.; Seidman, J.G.; Seidman, C.E.; Chen, X.J.; et al. Tbx5-dependent rheostatic control of cardiac gene expression and morphogenesis. Dev. Biol. 2006, 297, 566–586. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Evans, S.M.; Sun, Y. Development of the cardiac pacemaker. Cell. Mol. Life Sci. 2017, 74, 1247–1259. [Google Scholar] [CrossRef]
- Pezhouman, A.; Engel, J.L.; Nguyen, N.B.; Skelton, R.J.P.; Gilmore, W.B.; Qiao, R.; Sahoo, D.; Zhao, P.; Elliott, D.A.; Ardehali, R. Isolation and characterization of human embryonic stem cell-derived heart field-specific cardiomyocytes unravels new insights into their transcriptional and electrophysiological profiles. Cardiovasc. Res. 2022, 118, 828–843. [Google Scholar] [CrossRef]
- Wiese, C.; Grieskamp, T.; Airik, R.; Mommersteeg, M.T.M.; Gardiwal, A.; de Geier-de Vries, C.; Schuster-Gossler, K.; Moorman, A.F.M.; Kispert, A.; Christoffels, V.M. Formation of the sinus node head and differentiation of sinus node myocardium are independently regulated by Tbx18 and Tbx3. Circ. Res. 2009, 104, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Plotnikov, A.N.; Shlapakova, I.; Szabolcs, M.J.; Danilo, P., Jr.; Lorell, B.H.; Potapova, I.A.; Lu, Z.; Rosen, A.B.; Mathias, R.T.; Brink, P.R.; et al. Xenografted adult human mesenchymal stem cells provide a platform for sustained biological pacemaker function in canine heart. Circulation 2007, 116, 706–713. [Google Scholar] [CrossRef] [PubMed]
- Bakker, M.L.; Boink, G.J.; Boukens, B.J.; Verkerk, A.O.; van den Boogaard, M.; den Haan, A.D.; Hoogaars, W.M.; Buermans, H.P.; de Bakker, J.M.; Seppen, J.; et al. T-box transcription factor TBX3 reprograms mature cardiac myocytes into pacemaker-like cells. Cardiovasc. Res. 2012, 94, 439–449. [Google Scholar] [CrossRef]
- Jun, C.; Zhihui, Z.; Lu, W.; Yaoming, N.; Lei, W.; Yao, Q.; Zhiyuan, S. Canine bone marrow mesenchymal stromal cells with lentiviral mHCN4 gene transfer create cardiac pacemakers. Cytotherapy 2012, 14, 529–539. [Google Scholar] [CrossRef]
- Boink, G.J.; Duan, L.; Nearing, B.D.; Shlapakova, I.N.; Sosunov, E.A.; Anyukhovsky, E.P.; Bobkov, E.; Kryukova, Y.; Ozgen, N.; Danilo, P., Jr.; et al. HCN2/SkM1 gene transfer into canine left bundle branch induces stable, autonomically responsive biological pacing at physiological heart rates. J. Am. Coll. Cardiol. 2013, 61, 1192–1201. [Google Scholar] [CrossRef]
- Kapoor, N.; Liang, W.; Marban, E.; Cho, H.C. Transcription factor-driven conversion of quiescent cardiomyocytes to pacemaker cells. Nat. Biotechnol. 2013, 31, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.J.; Husse, B.; Rimmbach, C.; Krebs, S.; Stieber, J.; Steinhoff, G.; Dendorfer, A.; Franz, W.M.; David, R. Programming and isolation of highly pure physiologically and pharmacologically functional sinus-nodal bodies from pluripotent stem cells. Stem Cell Rep. 2014, 2, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Ionta, V.; Liang, W.; Kim, E.H.; Rafie, R.; Giacomello, A.; Marban, E.; Cho, H.C. SHOX2 overexpression favors differentiation of embryonic stem cells into cardiac pacemaker cells, improving biological pacing ability. Stem Cell Rep. 2015, 4, 129–142. [Google Scholar] [CrossRef]
- Saito, Y.; Nakamura, K.; Yoshida, M.; Sugiyama, H.; Ohe, T.; Kurokawa, J.; Furukawa, T.; Takano, M.; Nagase, S.; Morita, H.; et al. Enhancement of spontaneous activity by HCN4 overexpression in mouse embryonic stem cell-derived cardiomyocytes—A possible biological pacemaker. PLoS ONE 2015, 10, e0138193. [Google Scholar] [CrossRef]
- Saito, Y.; Nakamura, K.; Yoshida, M.; Sugiyama, H.; Akagi, S.; Miyoshi, T.; Morita, H.; Ito, H. Enhancement of pacing function by HCN4 overexpression in human pluripotent stem cell-derived cardiomyocytes. Stem Cell Res. Ther. 2022, 13, 141. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Yang, P.; Luo, S.; Zhang, Z.; Li, H.; Zhu, P.; Song, Z. Shox2 influences mesenchymal stem cell fate in a co-culture model in vitro. Mol. Med. Rep. 2016, 14, 637–642. [Google Scholar] [CrossRef][Green Version]
- Yang, M.; Zhang, G.G.; Wang, T.; Wang, X.; Tang, Y.H.; Huang, H.; Barajas-Martinez, H.; Hu, D.; Huang, C.X. TBX18 gene induces adipose-derived stem cells to differentiate into pacemaker-like cells in the myocardial microenvironment. Int. J. Mol. Med. 2016, 38, 1403–1410. [Google Scholar] [CrossRef]
- Yang, M.; Zhao, Q.; Zhao, H.; Yang, A.; Wang, F.; Wang, X.; Tang, Y.; Huang, C. Adipose-derived stem cells overexpressing SK4 calcium-activated potassium channel generate biological pacemakers. Int. J. Mol. Med. 2019, 44, 2103–2112. [Google Scholar] [CrossRef]
- Chan, P.K.W.; Geng, L.; Gao, Y.; Keung, W.; Li, R.A. AAV-mediated conversion of human pluripotent stem cell-derived pacemaker. Biochem. Biophys. Res. Commun. 2017, 494, 346–351. [Google Scholar] [CrossRef]
- Li, Y.; Yang, M.; Zhang, G.; Li, L.; Ye, B.; Huang, C.; Tang, Y. Transcription factor TBX18 promotes adult rat bone mesenchymal stem cell differentiation to biological pacemaker cells. Int. J. Mol. Med. 2018, 41, 845–851. [Google Scholar] [CrossRef]
- Sun, A.J.; Qiao, L.; Huang, C.; Zhang, X.; Li, Y.Q.; Yang, X.Q. Comparison of mouse brown and white adipose-derived stem cell differentiation into pacemaker-like cells induced by TBX18 transduction. Mol. Med. Rep. 2018, 17, 7055–7064. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Yang, Y.J.; Lin, Y.Z.; Peng, S.; Lin, S.; Song, Z.Y. Transcription factor Tbx18 induces the differentiation of c-kit+ canine mesenchymal stem cells (cMSCs) into SAN-like pacemaker cells in a co-culture model in vitro. Am. J. Transl. Res. 2018, 10, 2511–2528. [Google Scholar]
- Darche, F.F.; Rivinius, R.; Köllensperger, E.; Leimer, U.; Germann, G.; Seckinger, A.; Hose, D.; Schröter, J.; Bruehl, C.; Draguhn, A.; et al. Pacemaker cell characteristics of differentiated and HCN4-transduced human mesenchymal stem cells. Life Sci. 2019, 232, 116620. [Google Scholar] [CrossRef] [PubMed]
- Gorabi, A.M.; Hajighasemi, S.; Tafti, H.A.; Atashi, A.; Soleimani, M.; Aghdami, N.; Saeid, A.K.; Khori, V.; Panahi, Y.; Sahebkar, A. TBX18 transcription factor overexpression in human-induced pluripotent stem cells increases their differentiation into pacemaker-like cells. J. Cell. Physiol. 2019, 234, 1534–1546. [Google Scholar] [CrossRef] [PubMed]
- Gorabi, A.M.; Hajighasemi, S.; Khori, V.; Soleimani, M.; Rajaei, M.; Rabbani, S.; Atashi, A.; Ghiaseddin, A.; Saeid, A.K.; Tafti, H.A.; et al. Functional biological pacemaker generation by T-Box18 protein expression via stem cell and viral delivery approaches in a murine model of complete heart block. Pharmacol. Res. 2019, 141, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, N.; Liu, L.; Zhang, H.; Xue, X.; Shao, X.; Zhang, Y.; Lang, X. Genetically modified porcine mesenchymal stem cells by lentiviral Tbx18 create a biological pacemaker. Stem Cells Int. 2019, 2019, 3621314. [Google Scholar] [CrossRef]
- Végh, A.M.D.; den Haan, A.D.; Cócera Ortega, L.; Verkerk, A.O.; Sluijter, J.P.G.; Bakker, D.; van Amersfoorth, S.; van Veen, T.A.B.; Klerk, M.; Seppen, J.; et al. Cardiomyocyte progenitor cells as a functional gene delivery vehicle for long-term biological pacing. Molecules 2019, 24, 181. [Google Scholar] [CrossRef]
- Végh, A.M.D.; Verkerk, A.O.; Cócera Ortega, L.; Wang, J.; Geerts, D.; Klerk, M.; Lodder, K.; Nobel, R.; Tijsen, A.J.; Devalla, H.D.; et al. Toward biological pacing by cellular delivery of Hcn2/SkM1. Front. Physiol. 2021, 11, 588679. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, C. A new combination of transcription factors increases the harvesting efficiency of pacemaker-like cells. Mol. Med. Rep. 2019, 19, 3584–3592. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, F.; Zhang, W.; Yang, M.; Tang, Y.; Wang, X.; Zhao, Q.; Huang, C. Overexpression of TBX3 in human induced pluripotent stem cells (hiPSCs) increases their differentiation into cardiac pacemaker-like cells. Biomed. Pharmacother. 2020, 130, 110612. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, F.; Tang, Y.; Wang, X.; Wang, T.; Zhao, Q.; Huang, C. HCN2 and TBX3 reprogram human-induced pluripotent stem cells-derived cardiomyocytes into pacemaker-like cells. DNA Cell Biol. 2020, 39, 289–298. [Google Scholar] [CrossRef]
- Raghunathan, S.; Islas, J.F.; Mistretta, B.; Iyer, D.; Shi, L.; Gunarathne, P.H.; Ko, G.; Schwartz, R.J.; McConnell, B.K. Conversion of human cardiac progenitor cells into cardiac pacemaker-like cells. J. Mol. Cell. Cardiol. 2020, 138, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhao, H.; Yin, L.; Tang, Y.; Wang, X.; Zhao, Q.; Wang, T.; Huang, C. Transcription factor TBX18 reprograms vascular smooth muscle cells of ascending aorta to pacemaker-like cells. DNA Cell Biol. 2019, 38, 1470–1479. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, H.; Quan, D.; Tang, Y.; Wang, X.; Huang, C. Tbx18 promoted the conversion of human-induced pluripotent stem cell-derived cardiomyocytes into sinoatrial node-like pacemaker cells. Cell Biol. Int. 2022, 46, 403–414. [Google Scholar] [CrossRef]
- Ieda, M.; Fu, J.D.; Delgado-Olguin, P.; Vedantham, V.; Hayashi, Y.; Bruneau, B.G.; Srivastava, D. Direct reprogramming of fibroblasts into functional cardiomyocytes by defined factors. Cell 2010, 142, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Protze, S.; Khattak, S.; Poulet, C.; Lindemann, D.; Tanaka, E.M.; Ravens, U. A new approach to transcription factor screening for reprogramming of fibroblasts to cardiomyocyte-like cells. J. Mol. Cell. Cardiol. 2012, 53, 323–332. [Google Scholar] [CrossRef]
- Song, K.; Nam, Y.J.; Luo, X.; Qi, X.; Tan, W.; Huang, G.N.; Acharya, A.; Smith, C.L.; Tallquist, M.D.; Neilson, E.G.; et al. Heart repair by reprogramming non-myocytes with cardiac transcription factors. Nature 2012, 485, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Alt, E.U.; Schmitz, C.; Bai, X. Perspective: Why and how ubiquitously distributed, vascular-associated, pluripotent stem cells in the adult body (vaPS Cells) are the next generation of medicine. Cells 2021, 10, 2303. [Google Scholar] [CrossRef]
- Alt, E.U.; Winnier, G.; Haenel, A.; Rothoerl, R.; Solakoglu, O.; Alt, C.; Schmitz, C. Towards a comprehensive understanding of UA-ADRCs (uncultured, autologous, fresh, unmodified, adipose derived regenerative cells, isolated at point of care) in regenerative medicine. Cells 2020, 9, 1097. [Google Scholar] [CrossRef]
- Baxter, M.A.; Wynn, R.F.; Jowitt, S.N.; Wraith, J.E.; Fairbairn, L.J.; Bellantuono, I. Study of telomere length reveals rapid aging of human marrow stromal cells following in vitro expansion. Stem Cells 2004, 22, 675–682. [Google Scholar] [CrossRef]
- Beane, O.S.; Fonseca, V.C.; Cooper, L.L.; Koren, G.; Darling, E.M. Impact of aging on the regenerative properties of bone marrow-, muscle-, and adipose-derived mesenchymal stem/stromal cells. PLoS ONE 2014, 9, e115963. [Google Scholar] [CrossRef] [PubMed]
- Bruno, I.; Martinez, R.; Sanchez, A.; Friddle, C.; McClure, S.R. Characterization of nucleated cells from equine adipose tissue and bone marrow aspirate processed for point-of-care use. J. Equine Vet. Sci. 2014, 34, 1118–1127. [Google Scholar] [CrossRef]
- Yu, H.; Lu, K.; Zhu, J.; Wang, J. Stem cell therapy for ischemic heart diseases. Br. Med. Bull. 2017, 121, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Winnier, G.E.; Valenzuela, N.; Peters-Hall, J.; Kellner, J.; Alt, C.; Alt, E.U. Isolation of adipose tissue derived regenerative cells from human subcutaneous tissue with or without the use of an enzymatic reagent. PLoS ONE 2019, 14, e0221457. [Google Scholar] [CrossRef]
- Stalmann, U.S.A.; Banjanin, B.; Snoeren, I.A.M.; Nagai, J.S.; Leimkühler, N.B.; Li, R.; Benabid, A.; Pritchard, J.E.; Malyaran, H.; Neuss, S.; et al. Single-cell analysis of cultured bone marrow stromal cells reveals high similarity to fibroblasts in situ. Exp. Hematol. 2022, 110, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Reiser, J.; Lai, Z.; Zhang, X.Y.; Brady, R.O. Development of multigene and regulated lentivirus vectors. J. Virol. 2000, 74, 10589–10599. [Google Scholar] [CrossRef]
- Islas, J.F.; Liu, Y.; Weng, K.C.; Robertson, M.J.; Zhang, S.; Prejusa, A.; Harger, J.; Tikhomirova, D.; Chopra, M.; Iyer, D.; et al. Transcription factors ETS2 and MESP1 transdifferentiate human dermal fibroblasts into cardiac progenitors. Proc. Natl. Acad. Sci. USA 2012, 109, 13016–13021. [Google Scholar] [CrossRef]
- White, S.M.; Claycomb, W.C. Embryonic stem cells form an organized, functional cardiac conduction system in vitro. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H670–H679. [Google Scholar] [CrossRef] [PubMed]
- Verheijck, E.E.; Wessels, A.; van Ginneken, A.C.; Bourier, J.; Markman, M.W.; Vermeulen, J.L.; de Bakker, J.M.; Lamers, W.H.; Opthof, T.; Bouman, L.N. Distribution of atrial and nodal cells within the rabbit sinoatrial node: Models of sinoatrial transition. Circulation 1998, 97, 1623–1631. [Google Scholar] [CrossRef]
- Hashem, S.I.; Claycomb, W.C. Genetic isolation of stem cell-derived pacemaker-nodal cardiac myocytes. Mol. Cell. Biochem. 2013, 383, 161–171. [Google Scholar] [CrossRef] [PubMed]
- King, N.M.; Perrin, J. Ethical issues in stem cell research and therapy. Stem Cell Res. Ther. 2014, 5, 85. [Google Scholar] [CrossRef]
- Fujikawa, T.; Oh, S.H.; Pi, L.; Hatch, H.M.; Shupe, T.; Petersen, B.E. Teratoma formation leads to failure of treatment for type I diabetes using embryonic stem cell-derived insulin-producing cells. Am. J. Pathol. 2005, 166, 1781–1791. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, U.; Benvenisty, N. The tumorigenicity of human embryonic and induced pluripotent stem cells. Nat. Rev. Cancer. 2011, 11, 268–277. [Google Scholar] [CrossRef]
- DiFrancesco, D.; Ferroni, A.; Mazzanti, M.; Tromba, C. Properties of the hyperpolarizing-activated current (if) in cells isolated from the rabbit sino-atrial node. J. Physiol. 1986, 377, 61–88. [Google Scholar] [CrossRef]
- Wu, J.; Schuessler, R.B.; Rodefeld, M.D.; Saffitz, J.E.; Boineau, J.P. Morphological and membrane characteristics of spider and spindle cells isolated from rabbit sinus node. Am. J. Physiol. Heart Circ. Physiol. 2001, 80, H1232–H1240. [Google Scholar] [CrossRef]
- Schäffler, A.; Büchler, C. Concise review: Adipose tissue-derived stromal cells--basic and clinical implications for novel cell-based therapies. Stem Cells. 2007, 25, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Gentile, P.; Orlandi, A.; Scioli, M.G.; Di Pasquali, C.; Bocchini, I.; Cervelli, V. Concise review: Adipose-derived stromal vascular fraction cells and platelet-rich plasma: Basic and clinical implications for tissue engineering therapies in regenerative surgery. Stem Cells Transl. Med. 2012, 1, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Warren, L.; Lin, C. mRNA-based genetic reprogramming. Mol. Ther. 2019, 27, 729–734. [Google Scholar] [CrossRef]
- Steinle, H.; Weber, M.; Behring, A.; Mau-Holzmann, U.; von Ohle, C.; Popov, A.F.; Schlensak, C.; Wendel, H.P.; Avci-Adali, M. Reprogramming of urine-derived renal epithelial cells into iPSCs using srRNA and consecutive differentiation into beating cardiomyocytes. Mol. Ther. Nucleic Acids. 2017, 17, 907–921. [Google Scholar] [CrossRef]
- Wesselhoeft, R.A.; Kowalski, P.S.; Parker-Hale, F.C.; Huang, Y.; Bisaria, N.; Anderson, D.G. RNA circularization diminishes immunogenicity and can extend translation duration in vivo. Mol. Cell. 2019, 74, 508–520. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group Name | Description of Treatments |
|---|---|
| SHOX2 | ASCs transduced with SHOX2 |
| TBX3 | ASCs transduced with TBX3 |
| TBX5 | ASCs transduced with TBX5 |
| TBX18 | ASCs transduced with TBX18 |
| HCN2 | ASCs transduced with HCN2 |
| SH | ASCs transduced with double combination of SHOX2 and HCN2 |
| SHT3 | ASCs transduced with triple combination of SHOX2, TBX3 and HCN2 |
| SHT5 | ASCs transduced with triple combination of SHOX2, TBX5 and HCN2 |
| SHT18 | ASCs transduced with triple combination of SHOX2, TBX18 and HCN2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karimi, T.; Pan, Z.; Potaman, V.N.; Alt, E.U. Conversion of Unmodified Stem Cells to Pacemaker Cells by Overexpression of Key Developmental Genes. Cells 2023, 12, 1381. https://doi.org/10.3390/cells12101381
Karimi T, Pan Z, Potaman VN, Alt EU. Conversion of Unmodified Stem Cells to Pacemaker Cells by Overexpression of Key Developmental Genes. Cells. 2023; 12(10):1381. https://doi.org/10.3390/cells12101381
Chicago/Turabian StyleKarimi, Tahereh, Zhizhong Pan, Vladimir N. Potaman, and Eckhard U. Alt. 2023. "Conversion of Unmodified Stem Cells to Pacemaker Cells by Overexpression of Key Developmental Genes" Cells 12, no. 10: 1381. https://doi.org/10.3390/cells12101381
APA StyleKarimi, T., Pan, Z., Potaman, V. N., & Alt, E. U. (2023). Conversion of Unmodified Stem Cells to Pacemaker Cells by Overexpression of Key Developmental Genes. Cells, 12(10), 1381. https://doi.org/10.3390/cells12101381