Abstract
In filamentous fungal Aspergillus species, growth, development, and secondary metabolism are genetically programmed biological processes, which require precise coordination of diverse signaling elements, transcription factors (TFs), upstream and downstream regulators, and biosynthetic genes. For the last few decades, regulatory roles of these controllers in asexual/sexual development and primary/secondary metabolism of Aspergillus species have been extensively studied. Among a wide spectrum of regulators, a handful of global regulators govern upstream regulation of development and metabolism by directly and/or indirectly affecting the expression of various genes including TFs. In this review, with the model fungus Aspergillus nidulans as the central figure, we summarize the most well-studied main upstream regulators and their regulatory roles. Specifically, we present key functions of heterotrimeric G proteins and G protein-coupled receptors in signal transduction), the velvet family proteins governing development and metabolism, LaeA as a global regulator of secondary metabolism, and NsdD, a key GATA-type TF, affecting development and secondary metabolism and provide a snapshot of overall upstream regulatory processes underlying growth, development, and metabolism in Aspergillus fungi.
1. Introduction
Fungi are of crucial importance in human lives and natural environments, largely because of their various roles in medicine production, environmental recycling, food process, and agriculture. Due to their strong influence on humankind, researchers initiated an investigation on metabolic mutants of the filamentous fungus Neurospora crassa in 1941, which established the basis of genetics: the one gene, one protein hypothesis. Since then, fungi have served as model organisms for eukaryotic genetic studies advancing the knowledge of modern genetics [1]. Among multitudinous fungi, the genus Aspergillus is comprised of the most common and ubiquitous fungi, with more than 340 species being identified [2]. In addition to the prevalence, their relevance to the various areas in human health (Aspergillus fumigatus), industry (Aspergillus niger and Aspergillus oryzae), agriculture (Aspergillus flavus), and genetic research (Aspergillus nidulans) has drawn much attention from researchers leading to extensive genetic studies on Aspergillus species to understand the regulatory mechanisms of fungal growth, development, and metabolism.
Aspergillus fungi may undergo three different life cycles: asexual, sexual, and parasexual cycles [1]. Approximately only one-third of the identified Aspergillus species are known to have sexual development under appropriate conditions, and they primarily reproduce through asexual sporulation (conidiation) [3]. These developmental processes result in the formation of specialized structures, which generate and bear sexual (ascospores) or asexual (conidia) spores. In some Aspergillus species, it has been reported that developmental phases are tightly coordinated with the production of certain secondary metabolites including mycotoxins [4,5,6]. Aspergillus development is complex, precisely timed, and genetically programmed events involving specialized cellular differentiation, temporal and spatial regulation of gene expression, and intra- and inter-cellular communications, involving coordination of various TFs and regulators. Within this complex system, some regulators function at the upstream level bringing huge impacts on Aspergillus biology by controlling the expression of diverse downstream genes, which eventually leads to the alteration of developmental and metabolic processes. Not only TFs, but also any genes or molecules affecting gene expression are considered as upstream regulators [7].
A large array of upstream regulators affecting conidiation, sexual development, and secondary metabolism in Aspergillus species have been identified and functionally studied [8,9,10,11,12,13,14]. In this review, we summarize a few different types of upstream regulators, known for their roles in main upstream regulation of Aspergillus biology (Figure 1): G proteins and G protein-coupled receptors (signal transduction), the velvet family proteins (development and metabolism), LaeA (secondary metabolism), and NsdD (development and secondary metabolism).
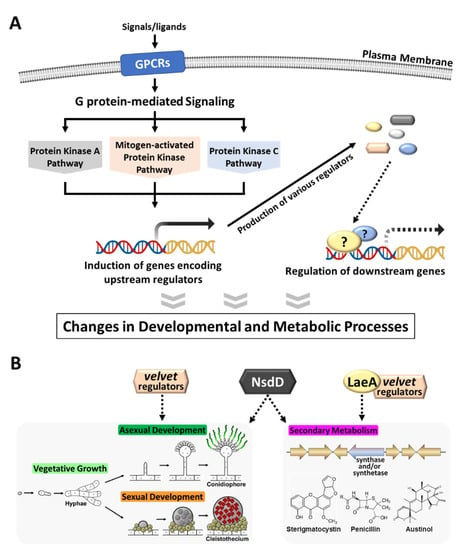
Figure 1.
A schematic overview of upstream regulation in Aspergillus species. (A) The heterotrimeric G protein-mediated signaling pathway, initiated by binding of ligands or signals to the corresponding G-protein coupled receptors (GPCRs), relays external environmental signals inside the cell and initiates downstream signaling pathway(s): protein kinase A (PKA), mitogen-activated protein kinase (MAPK), and protein kinase C (PKC). As a result, the expression of genes encoding upstream regulators changed either up-regulated or down-regulated, which in turn affects expression levels of various downstream genes. Fungi efficiently control their growth, development, and metabolism through these signal transduction pathways. (B) Three different types of well-studied upstream regulations occurred by the velvet family regulators (developmental regulators), LaeA (a global regulator of secondary metabolism), and NsdD (a key regulator of asexual/sexual development). Solid line represents an activating role and dotted line represents a regulating role (either activating or repressing).
2. Heterotrimeric G Protein Signaling Governs Development and Metabolism
Sensing the external environment and adapting to surroundings are crucial for fungi to coordinate their growth and development accordingly. Fungi are responsive to environmental clues through the heterotrimeric G protein signaling pathway. G protein-coupled receptors (GPCRs) relay signals from the extracellular environment inside the cell by existing across a cell membrane. These receptors react to a large spectrum of external signals including light, hormones, neurotransmitters, cytokines, growth factors, cell adhesion molecules, and nutrients, such as sugars, amino acids, and nitrogen sources [15]. Binding to extracellular molecules causes a conformational change in GPCRs and this alteration then activates the interaction between the GPCR and G protein, which bound to C-terminus of the GPCR in cytosol. When the G protein becomes active by exchanging GDP with GTP, it can trigger the production of thousands of second messenger molecules, such as cyclic AMP (cAMP), diacylglycerol (DAG), and inositol 1, 4, 5-triphosphate (IP3), which initiate and coordinate further intracellular signaling pathways [16]. In fungi, G protein-mediated signaling pathways include the cAMP-dependent protein kinase (PKA) and the mitogen-activated protein kinase (MAPK) pathways. Through these sequences of events, fungi regulate their growth, development, morphogenesis, mating, metabolism, virulence, and mycotoxin biosynthesis according to the environments where they are situated [17,18].
2.1. G Protein-Coupled Receptors (GPCRs)
G protein-coupled receptors (GPCRs) are transmembrane proteins and the largest class of cell surface receptors in fungi. GPCRs are plasma-membrane-localized proteins that communicate changes in the environment to intracellular heterotrimeric G proteins [16]. GPCRs contain seven transmembrane (7-TM) helices connected by intracellular and extracellular loops, with an extracellular amino-terminus (N-terminus) and the carboxyl-terminus (C-terminus) extending into the cytoplasm. Although the majority of GPCRs consist of 7-TM helices, some GPCRs are containing 5-TM or 6-TM helices, such as GprB, GprG, GprN, and NopA. According to the previous studies, Aspergillus GPCRs are classified into ten different classes. Eighteen GPCRs named GprA to GprS and NopA belong to Class I to IX, and Class X GPCRs are Pth11-like receptors, which promote fungal-plant pathogenic interactions in A. nidulans (Table 1). GprN was specifically identified in A. nidulans, and GprR and GprS were exclusively identified in A. flavus, which contain the regulator of G protein signaling (RGS) domain and PQ-loop repeat domain, respectively [19]. Despite their prevalence and fundamental roles in fungi, only a few GPCRs have been identified and functionally characterized. In recent years, the rapid development and evolution of Next-Generation Sequencing technology have boosted GPCR studies by enabling accessible large-scale whole genome sequencing, which led to figuring out putative GPCRs in genome based on structural similarities and putative activating ligands. These studies have revealed that the A. nidulans genome encodes 86 putative GPCRs, which can be divided into 16 GPCRs in 9 categories (Class I to IX) and 70 class X Pth11-like receptors [20,21,22,23]. The major producer of aflatoxins A. flavus’ genome encodes fifteen putative GPCRs in nine categories (Class I to IX), and Class X GPCRs still remain to be identified (reviewed in [19]). In the opportunistic human pathogen A. fumigatus, the genome encodes fifteen putative classical GPCRs (Class I to IX), yet only five of them (GprC, GprD, GprK, GprM, and GprJ) have been characterized [24,25,26]. Functional studies on GPCRs have unveiled that they play significant roles in overall fungal biology relating to nutrient sensing, fungal development, pheromone response, fruiting body formation, mycotoxin production, and pathogenesis. According to functional characteristics, GPCRs can be categorized into 10 groups: pheromone (classes I and II), carbon (III), nitrogen (IV), cAMP receptor-like (V), RGS (Regulator of G protein signaling, VI), MG00532-like (VII), mPR-like (VIII), microbial opsin (IX), and Pth11-like (X) receptors.
The pheromone receptors were first identified in Saccharomyces cerevisiae. Two different pheromone receptors, Ste2p (α-factor receptor) and Ste3p (a-factor receptor), presented in the cell membranes of opposite haploid mating types (MATα and MATa). When yeast cells are exposed to the pheromone secreted by the opposite mating type, their pheromone receptors are activated and initiate G protein-mediated signaling pathway leading to the eventual fusion with the mating partner [27]. Carbon-sensing receptors regulate the response to carbon sources in fungi. In the filamentous fungus N. crassa, GPR-4 (G-protein coupled receptor 4) physically interacts with the Gα (GNA-1) to regulate carbon source-dependent growth and development. The gpr-4 null mutants displayed less mass accumulation compared to the WT in carbon-limited conditions and no transient increase in cAMP levels upon a nutrient shift from carbon-limited to glucose-rich media, which was normally observed in WT [28]. Nitrogen-sensing receptors act in a very similar way to carbon-sensing ones. In Schizosaccharomyces pombe, the Stm1 receptor, coupling with the Gα2 protein, is required for proper recognition of nitrogen starvation signals. Overexpression of Stm1 led to the inhibition of vegetative growth and the decrease in intracellular cAMP levels even under nutrient-rich conditions [29]. The cAMP receptors (cARs) were firstly identified in Dictyostelium discoideum and then the sequences of cARs were used to predict cAMP receptor-like GPCRs (Crls) in D. discoideum and likely fungal cAMP receptor genes. The cARs are known to play significant roles during divergent developmental stages and in distinct subsets of developing cells within multicellular aggregates by interacting with secreted cAMP [30,31,32]. The N. crassa GPR-1, distantly related to the four cAMP receptors (cAR1 to cAR4) and three cAMP receptor-like GPCRs (CrlA to CrlC), was the first cAMP receptor-like GPCR characterized in ascomycete fungi. In N. crassa, GPR-1 is localized in female reproductive structures and regulates female sexual development [33]. The GPCR containing RGS domain was firstly discovered in Arabidopsis thaliana. The A. thaliana RGS, AtRGS1 protein negatively regulates the Gpa1 Gα subunit affecting cellular proliferation. Canonical GPCRs cause the conformational change of G protein triggering the GDP-GTP exchange, but instead, AtRGS1 interacts with the active Gα subunit resulting in hydrolysis of GTP, which in turn deactivates the G protein [34]. This type of GPCR has been found in several species of filamentous fungi. In Aspergillus species, GprK containing both 7-TM and RGS domains is similar to AtRGS1 and involved in germination, development, and stress response [25]. The MG00532 group was represented by a protein with weak homology to rat growth hormone-releasing factor. The mPR-like class of GPCR includes proteins related to the human membrane progesterone receptors (mPRs), which mediate an array of rapid, cell surface-initiated progesterone actions in the reproductive system involving activation of intracellular signaling pathways [35,36]. The microbial opsins are a class of retinal-binding proteins with seven membrane-spanning domains that form rhodopsins by interacting with the retinal and function as light-responsive ion pumps or sensory receptors. The NOP-1 protein of N. crassa, closely related to archaeal opsins, was the first opsin characterized in filamentous fungi and known to bind all-trans retinal by using a Schiff base linkage and play a role in N. crassa photobiology [37]. The PTH11 protein was first discovered in the plant pathogenic fungus Magnaporthe oryzae and identified as an activator of appressorium differentiation in response to inductive surfaces. The aberration of the pth11 gene in M. oryzae led to the defect in pathogenicity [22]. Although a large number of Pth11-like GPCRs have been predicted, in Aspergillus, their exact functions remain heavily unknown [19,38].
In the presence of extracellular signals, corresponding GPCRs recognize molecules and relay the signal inside the cell. The recognition of external cues by GPCRs provokes the conformational change of G protein, which in turn initiates the G protein signaling pathways including the cAMP-dependent protein kinase (PKA) and the mitogen-activated protein kinase (MAPK) pathways.

Table 1.
G protein-coupled receptors in Aspergillus species.
Table 1.
G protein-coupled receptors in Aspergillus species.
| Gene | Class | TM No. | Conserved Domains (Note) | Ref |
|---|---|---|---|---|
| gprA | I | 7-TM | Ste2; alpha-pheromone receptor (S. cerevisiae pheromone receptor) | [39] |
| gprB | II | 5-TM | Ste3; a-pheromone receptor (S. cerevisiae pheromone receptor) | [39] |
| gprC | III | 7-TM | Git3; Gpa2 C-term; Family 1-like (S. pombe glucose receptor) | [40] |
| gprD | III | 7-TM | Git3; Gpa2 C-term (S. pombe glucose receptor) | [40] |
| gprE | III | 7-TM | Git3; Gpa2 C-term (S. pombe glucose receptor) | [40] |
| gprF | IV | 7-TM | PQ-loop repeat (S. pombe nitrogen sensor) | [40] |
| gprG | IV | 5-TM | PQ-loop repeat (S. pombe nitrogen sensor) | [40] |
| gprJ | IV | 7-TM | PQ-loop repeat (S. pombe nitrogen sensor) | [38] |
| gprS | IV | 7-TM | PQ loop repeat (S. pombe nitrogen sensor) | [19] |
| gprH | V | 7-TM | Secretin-like; cAR (D. discoideum); Family 2-like (signal through cAMP pathways) | [40] |
| gprI | V | 7-TM | cAR (D. discoideum); Family 2-like (signal through cAMP pathways) | [40] |
| gprK | VI | 7-TM | RGS (regulator of G protein signaling) | [38] |
| gprR | VI | 7-TM | RGS (regulator of G protein signaling) | [19] |
| gprM | VII | 7-TM | MG00532-like | [38] |
| gprN | VII | 6-TM | MG00532-like | [38] |
| gprO | VIII | 7-TM | mPR-like; Hemolysin-III related (broad range of ligands) | [38] |
| gprP | VIII | 7-TM | mPR-like; Hemolysin-III related (broad range of ligands) | [38] |
| gprQ | VIII | 5-TM | mPR-like; Hemolysin-III related (broad range of ligands) | [38] |
| nopA | IX | 6-TM | Bacterial rhodopsin-like (photoreactive) | [38] |
| pth11-like | X | 7-TM | Pth11-related group | [38] |
2.2. G Protein-Mediated Signaling Pathway
Heterotrimeric guanine nucleotide-binding proteins (G proteins), consisting of alpha, beta, and gamma subunits, have been characterized in diverse eukaryotic organisms demonstrating their pivotal roles in major signal transduction pathways during the responses of cells to extracellular stimuli. The G proteins, present in all eukaryotic cells, control metabolic and developmental pathways (reviewed in [41]). In filamentous fungi, the first G proteins, particularly α subunits, were discovered in N. crassa in the early 1990s. Thereafter, the G proteins FadA (α subunit), SfaD (β subunit), and GpgA (γ subunit) were identified in A. nidulans [42,43,44]. The inactive G protein heterotrimer is composed of α, β, and γ subunits, whereas the α and γ subunits are associated with the plasma membrane. Upon binding of specific ligands, GPCRs experience a conformational change, then physically interact with heterotrimeric G proteins. This physical interaction results in an exchange of GTP for GDP on the Gα subunit, which in turn leads to the dissociation of the heterotrimer into the GTP-Gα subunit and the Gβγ heterodimer. Once dissociated, the GTP-Gα and the Gβγ become active so that GTP-Gα, Gβγ, or both moieties can relay and amplify signals by regulating activities of downstream effector proteins in divergent signal transduction pathways. Then, RGS proteins interact with an activated GTP-Gα subunit and increase its intrinsic GTPase activity. The GTP hydrolysis enables GDP-Gα subunit to reassociate with Gβγ heterodimer and plasma membrane, becoming into the inactive form of heterotrimeric G proteins again. In fungi, G protein-mediated signaling pathway is transmitted through one or more of the following pathways: (1) cAMP-dependent protein kinase (PKA), (2) mitogen-activated protein kinase (MAPK), and (3) Ca2+- and DAG-dependent protein kinase C (PKC) (Figure 2) [45,46].
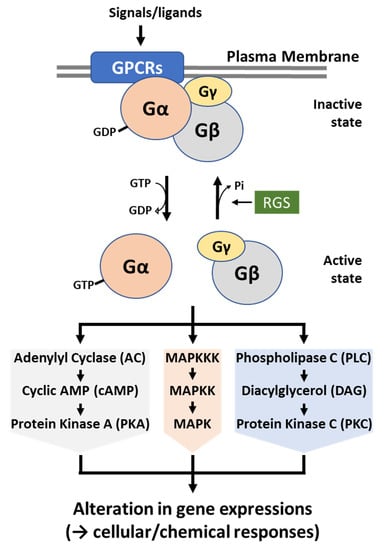
Figure 2.
A schematic diagram of heterotrimeric G protein-mediated signaling pathway. Upon the recognition of external ligands, a G protein-coupled receptor (GPCR) is sensitized and interacts with a nearby G protein. Then, G proteins become active by the interaction with GTP and initiate downstream signaling pathway(s): PKA, MAPK, and PKC. This signal transduction results in the differential expression of genes involved in growth, development, morphogenesis, mating, metabolism, virulence, and mycotoxin biosynthesis. The activated G proteins are then negatively controlled by regulators of G protein signaling (RGS) and become inactive again.
The Aspergillus species possess three distinct groups of Gα proteins. Each group of Gα was assigned according to the amino acid sequence similarity with the N. crassa Gα proteins; Gna-1 (group I), Gna-2 (group II), and Gna-3 (group III). The group I Gα proteins possess a consensus sequence for myristoylation (MGXXXS) at the N-terminus and a site for ADP-ribosylation by pertussis toxin (CAAX) at the C-terminus. Most well-characterized filamentous fungi are known to possess a single group I Gα protein and its function has been well elucidated. The group III Gα proteins are also highly conserved and possess a myristoylation at the N-terminus. They are known to positively influence cAMP levels. However, the functions of group II Gα proteins are not as obvious as group I and III Gα proteins (reviewed in [20]).
In A. nidulans, the first characterized group I Gα subunit was FadA, showing 93% identity of AA sequence to N. crassa Gna-1 and, thereafter, GanA (group II) and GanB (group III) had been identified. FadA (fluffy autolytic dominant) was initially investigated for the fluffy autolytic phenotype, which was attributed to an uncontrolled vegetative growth followed by autolysis [42]. The dominant activating mutations on FadA resulted in the expression of the fluffy autolytic phenotype and the inhibition of mycotoxin production, especially sterigmatocystin (ST), while dominant interfering FadA mutants displayed reduced vegetative growth, enhanced asexual sporulation, and precocious ST production [42,47]. Constitutively active dominant FadA mutants were presumed to maintain a longer period of the activated state of FadA-GTP due to the decreased intrinsic GTPase activity. Taken together, these results indicated that activated GTP-FadA (Gα) mediates signaling via cAMP-dependent PKA that promotes vegetative growth, which in turn suppresses asexual sporulation, sexual development, and mycotoxin production in Aspergillus.
The role of the group II Gα subunit, GanA has been well studied in A. fumigatus. GanA shares 46.3% and 44.3% identity with GpaA (group I, the homolog of FadA) and GpaB (group III, the homolog of GanB) in A. fumigatus, respectively. The mRNA level of ganA was highly expressed at both early (6 h) and later (48 h) time of asexual development and the deletion of the ganA gene resulted in faster germination, but decreased radial growth compared to those of WT on solid media; however, it did not show any significant impact on asexual development, unlike other Gα proteins. In addition, the ΔganA strain displayed reduced mRNA level of gliotoxin biosynthesis transcription factor gliZ and decreased GT production compared to those of WT as well. Interestingly, the ganA null mutant exhibited the highest activity of PKA in conidia and PKC in mycelia among Gα mutants [48]. These results demonstrated that GanA plays important roles in vegetative growth, asexual development, and mycotoxin production through PKA or PKC signaling pathway in a similar, but slightly different way from other groups of Gα proteins.
The group III Gα protein (GanB) is functionally well characterized in A. nidulans by Chang et al. [49] and Lafon et al. [50]. They revealed that GanB positively regulates conidial germination but inhibits asexual sporulation by mediating a rapid and transient increase in cAMP levels in response to the presence of extracellular glucose during the early phase of germination. Moreover, Lafon et al. [50] elucidated that GanB (Gα) and SfaD::GpgA (Gβγ) form a heterotrimeric complex. Collectively, G protein α subunits in Aspergillus mediate signaling that promotes vegetative growth and stress responses, which in turn inhibit fungal development and mycotoxin production (Figure 3).
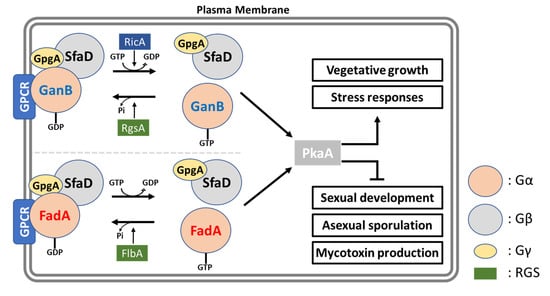
Figure 3.
Heterotrimeric G protein-mediated signaling pathway in A. nidulans. In A. nidulans, two Gα subunits (FadA and GanB) regulate vegetative growth, development, and mycotoxin production via PKA pathway when they are in the activated state. The regulators of G protein signaling (RGS) FlbA and RgsA turn activated FadA and GanB back into the inactive state, respectively. RicA is a GDP/GTP exchange factor involved in the activation of G protein subunits.
Most filamentous fungi are predicted to have a highly conserved single Gβ subunit (from 66 to 92% identical with N. crassa Gnb-1) and Gγ subunit (from 39 to 92% identical with N. crassa Gng-1) [20]. Previous studies on the Gβ and Gγ mutations have shown that mutational inactivation of genes encoding these proteins affected vegetative growth, conidiation, and sexual development in filamentous fungi [51,52]. Particularly, Krystofova and Borkovich [33] demonstrated that the gng-1 and gnb-1 loss-of-function mutations displayed similar phenotypes, such as female sterility, defective conidiation, low levels of intracellular cAMP, and a severe reduction in Gα protein levels in N. crassa. In addition, they proposed that Gng-1 (Gγ) physically interacts with Gnb-1 (Gβ) and forms the Gnb-1:Gng-1 (Gβγ) heterodimer during signaling pathways. In the genus Aspergillus, the Gβ subunit SfaD and Gγ subunit GpgA were identified and well characterized. Rosén et al. [43] isolated SfaD composed of 352 AA that shares 86% identity with N. crassa Gnb-1 and revealed that SfaD has a conserved Trp-Asp sequence, which is known as a WD40 domain. They revealed that SfaD plays crucial roles in vegetative growth, conidial sporulation, sexual development, and ST production in A. nidulans. Moreover, Seo et al. [44] identified the Gγ subunit GpgA, which consists of 90 AA that shows 65% identity with N. crassa Gng-1. The gpgA loss-of-function mutation exhibited reduced vegetative growth, delayed conidiation, and no sexual fruiting body formation, similar to ΔsfaD mutants. Later, Lafon et al. [50] revealed that the SfaD:GpgA (Gβγ) heterodimer is crucial for the proper activation of GanB (Gα), while GanB plays a primary role in the PKA signaling pathway in response to glucose.
Timely modulation of G protein-mediated signaling pathways is the key for fungi in sensing and responding to internal/external signals and various stress conditions. Upon the recognition of extracellular signals, cells need to activate G proteins immediately, so they can translate diverse incoming signals into corresponding cellular responses opportunely. However, the prolonged activated state of GTP-Gα can cause various defects in fungal development and metabolism. Thus, the neutralization of activated GTP-Gα into the inactive form on time is as significant as the activation. These tight upstream regulations play crucial roles in vegetative growth, development, mycotoxin production, and virulence in fungi. There are three different types of regulators present in Aspergillus species: phosducin-like proteins (PhLPs), regulators of G protein signaling (RGSs), and a GDP/GTP exchange factor (RicA).
Phosducin-like proteins are a group of evolutionarily conserved positive regulators of Gβγ heterodimer function. PhLPs act as molecular chaperones during Gβγ assembly by stabilizing the nascent Gβ subunit until it associates with the Gγ protein [53,54]. In A. nidulans, three potential PhLPs (PhnA, PhnB, and PhnC) were identified based on the AA sequence similarity with Bdm-1, which is a known fungal Gβγ activator in the chestnut blight fungus Cryphonectria parasitica [55]. Among them, the function of PhnA was firstly investigated by Seo and Yu [56] due to its highest similarity with Bdm-1. Seo and Yu [56] revealed that PhnA is required for proper SfaD functionality, sexual reproduction, and mycotoxin biosynthesis showing consistent results with the roles of SfaD::GpgA heterodimer.
RGSs are a group of proteins containing a conserved ∼130 AA RGS box, which physically interact with an activated GTP-Gα and accelerate the intrinsic GTPase activity of the Gα subunit, resulting in the attenuation of G protein-mediated signaling pathways [46,57]. In Aspergillus, several RGSs have been identified including FlbA, RgsA, RgsB, RgsC, GprK, and Rax1. Among them, FlbA and RgsA are the most well-characterized RGSs (Figure 3). The FlbA consists of 719 amino acids containing 1 RGS box and 2 DEP (Dishevelled, EGL-10, and Pleckstrin) domains. The DEP is a globular protein domain of ~80 AA commonly found in proteins involved in G-protein signaling, however, the repeated pattern of DEP is only observed in fungi [58]. Along with the GTPase-activating RGS domain, the DEP domain may play a role in guiding RGS proteins to the Golgi and plasma membranes [59]. The flbA loss-of-function exhibited the fluffy-autolytic phenotype, which was observed in activating dominant FadA (Gα) mutants. In addition, the fadA deletion mutants did not display the fluffy-autolytic phenotype caused by ΔflbA and restored asexual development and mycotoxin production. The primary role of FlbA is to attenuate G protein-mediated signaling by deactivating GTP-FadA (group I Gα) protein, whereas FadA, SfaD, and GpgA constitute the major G protein heterotrimer modulating growth, development, and secondary metabolism in A. nidulans [42,47]. The RgsA consists of 362 AA containing one RGS box in the N-terminal region. Unlike FlbA regulating group I Gα subunit, RgsA negatively regulates GanB (group III Gα) signaling, which promotes stress responses via the PKA pathway resulting in the inhibition of asexual development [58].
The GDP/GTP exchange factor RicA is recently discovered compared to other regulators of G proteins in Aspergillus. The ricA deletion mutants displayed severely reduced colony growth, and a total absence of asexual sporulation and sexual development in A. nidulans [60]. Kwon et al. [60] introduced the A. fumigatus ricA gene (AfricA) into A. nidulans ΔricA and found that the overexpression of AfricA in the ΔAnricA mutant partially restored colony growth and asexual development. In addition, they revealed that the removal of only rgsA, not sfgA, flbA, rgsB, or rgsC, restored vegetative growth and conidiation in ΔAnricA and that RicA can physically interact with GanB (Gα) in vitro in yeast. These results led Kwon et al. [60] to conclude that RicA primarily activates the GanB initiating PKA signaling cascade in A. nidulans.
3. The velvet Regulators and the Global Regulator of Secondary Metabolism LaeA
In filamentous fungi, fungal development, and secondary metabolism are intimately associated via the activities of the fungal-specific velvet family regulatory proteins and the global regulator of secondary metabolism LaeA. The velvet regulators form various complexes playing divergent roles in fungal development. The VosA-VelB heterodimer inhibits conidial germination, but positively regulates trehalose synthesis and β-glucan biogenesis. Moreover, VeA bridges VelB and LaeA to form the VelB-VeA-LaeA (velvet) heterotrimeric complex in the absence of light and this velvet complex controls not only secondary metabolism, but also the formation of Hülle cells, which nurse the nascent sexual fruiting bodies. This section summarizes the functions of velvet proteins and LaeA in Aspergillus.
3.1. The velvet Family Regulators
The first study of the velvet family regulators was conducted on the velvetA1 mutant (renamed as veA1 afterward). The colony of this mutant showed a flat and velvety appearance regardless of the presence or absence of light [61]. The functions of the veA gene have been extensively characterized in Aspergillus species. The veA gene is necessary for proper sexual development and secondary metabolism in Aspergillus [62,63,64,65,66,67,68]. In 2007, a major advancement in the velvet family study was made by Ni and Yu [69]. They demonstrated that the novel regulator VosA (viability of spores) governs sporogenesis and trehalose biogenesis, which in turn determines the viability of spores in A. nidulans. In addition, they discovered two other proteins that are similar to VosA; they named them VelB (velvet-like protein) and VelC. These four proteins were designated as the velvet family of regulators [69]. The velvet family proteins are highly conserved in Aspergilli and they all share a fungi-specific and highly conserved velvet domain, which consists of approximately 170–300 AA sequences with three conserved motifs [9].
3.2. The velvet Regulators in A. nidulans
VeA was the first member of the velvet family regulators identified in the 1960s [70]. VeA is composed of 573 amino acids containing the velvet domain in the N-terminal region (Figure 4). In addition, in the N-terminal region of this regulator, a potential nuclear localization signal (NLS) and nuclear export signal (NES) domains were found, suggesting roles in the nuclear localization of VeA [71]. Moreover, VeA contains a putative PEST (proline (P)-, glutamic acid (E)-, serine (S)-, and threonine (T)-rich) sequence in the C-terminal region [62], which is commonly found in rapidly degraded proteins [72]. VeA is a key light-dependent developmental regulator that positively regulates sexual development, which in turn suppresses asexual sporulation in A. nidulans [73,74]. The veA loss-of-function mutations resulted in the complete absence of sexual fruiting body formation, even under sexual development-promoting conditions, whereas the overexpression of veA enhanced the production of cleistothecia but inhibited asexual sporulation [62]. Furthermore, VeA is known to play crucial roles in secondary metabolism. VeA acts as an activator on sterigmatocystin production but inhibits penicillin biosynthesis [63,75]. Underlying the significant roles of VeA in fungal development and secondary metabolism, the nuclear localization of VeA is a vital factor. The VeA protein is constitutively expressed during the life cycle of A. nidulans but is mostly found in the cytoplasm under the presence of light [62,71,76]. On the other hand, in the dark, VeA enters the nucleus, forms VelB-VeA-LaeA heterotrimeric complex, and controls sexual development and mycotoxin production [71,77].
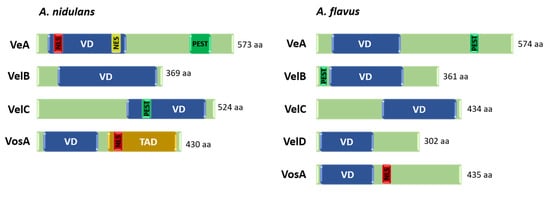
Figure 4.
Domain architecture of the velvet regulators in A. nidulans and A. flavus. VD: velvet domain, NLS: Nuclear localization signal, NES: nuclear export signal, PEST: Proline (P)-, glutamic acid (E)-, serine (S)-, and threonine (T)-rich sequence, TAD: transcription activation domain, aa: amino acids.
VosA consists of 430 AA containing the velvet, NLS, and potential TAD (transcription activation) domains, indicating that it may function as a transcription factor. VosA protein is expressed during vegetative growth and the early stage of asexual and sexual development, however, primarily localized in the nucleus of mature conidia. Interestingly, the expression of vosA is regulated by AbaA. In phialides, AbaA binds to the promoter region of vosA and induces the accumulation of vosA mRNA in conidia during the late phase of asexual development [67,69]. VosA is a key regulator of conidiation and sexual development. The vosA null mutants produced asexual developmental structures (conidiophores) in the liquid submerged culture, where the wild type solely undergoes vegetative growth and produced fewer numbers of cleistothecia compared to the WT. In addition, the deletion of vosA resulted in the accumulation of high mRNA levels of the brlA gene, which is a key initiative factor of conidiation, indicating VosA is a key negative regulator of brlA [69]. Moreover, VosA controls various biological processes including conidia wall integrity, spore viability, conidial germination, and focal trehalose biogenesis [78,79].
VelB is a 369-AA protein containing the velvet domain covering the entire protein. The velB gene is mostly expressed during the life cycle, but particularly high levels of velB mRNA are observed during vegetative growth and in the late phases of asexual and sexual development. Similarly, VelB protein is detectable during entire vegetative growth and in early developmental stages. VelB has divergent functions regulating vegetative growth, development, and secondary metabolism. VelB negatively regulates conidial germination but acts as an activator of asexual development. The velB deletion mutants showed increased conidial germination rates yet exhibited a reduced conidia production and decreased expression levels of asexual development-related genes, such as brlA and abaA. In addition, the deletion of velB led to the enhanced production of brown pigments [67,77].
VelC is composed of 524 AA containing the velvet and PEST domains in the C-terminal region. Unlike other velvet family members, the mRNA of velC specifically accumulates during the early phase of sexual development. The aberration of the velC gene led to the slightly enhanced conidia production and increased the mRNA levels of all three central regulatory genes of conidiation, brlA, abaA, and wetA, regardless of the presence or absence of light. In addition, the deletion of velC resulted in a decreased production of sexual fruiting bodies, while overexpression of this gene led to increased production of cleistothecia but a decreased number of conidia. These suggest that VelC plays a role in the activation of sexual development [9].
Individual velvet protein plays multifunctional roles in Aspergillus development and metabolism; however, the most significant trait of velvet proteins is that they interact with partner proteins including themselves, and form complexes in multiple combinations, which govern the various processes of fungal biology. The formation of velvet protein complexes occurs in a cell type- and/or timing-specific manners. Among all different velvet protein complexes, the three most extensively studied complexes, VosA-VelB, VelB-VeA, and VelB-VeA-LaeA, are discussed here (Figure 5). During germination, the VosA-VelB heterodimer inhibits conidial germination rates. Moreover, the VosA-VelB heterodimer controls spore viability, trehalose biogenesis, β-glucan synthesis, and tolerance of conidia to various stresses, such as heat and oxidative stress. The VelB-VeA complex is a key participant in sexual development. Although the molecular mechanism of the VelB-VeA heterodimer formation has not been clearly identified yet, the VelB-VeA complex plays an important role in sexual development as an activator. Furthermore, as VeA bridges between the VelB-VeA heterodimer and LaeA, the VelB-VeA complex interacts with LaeA and forms the VelB-VeA-LaeA heterotrimeric complex in the nucleus. This VelB-VeA-LaeA complex regulates sterigmatocystin production and sexual development in the dark. In addition, it controls the expression of secondary metabolism-related genes at transcriptional or epigenetic levels [77,80,81].
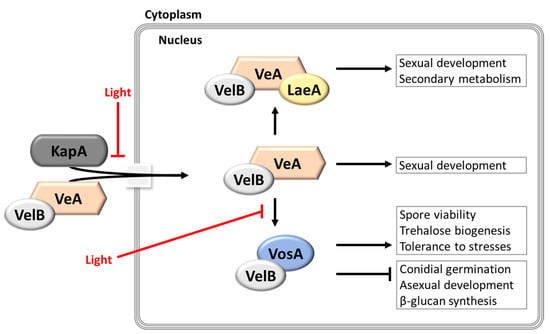
Figure 5.
The functions of velvet family proteins in Aspergillus species. This model is adopted and modified from [82]. VeA enters the nucleus together with VelB and the importin-α KapA in the dark. In the nucleus, velvet family proteins and LaeA can form three different complexes depending on the presence or absence of light: VelB-VosA, VelB-VeA, and VelB-VeA-LaeA. These complexes regulate different biological processes.
3.3. The velvet Regulators in Other Aspergillus Species
Most Aspergillus species have four velvet family regulators (VeA, VelB, VelC, and VosA), but a recent study newly identified the fifth member of the velvet family, VelD, in A. flavus [83]. Although the velvet family proteins are highly conserved in Aspergilli, their functions might have been divergent depending on the species. In the plant pathogenic and mycotoxigenic fungus A. flavus, the VeA protein was first identified among the velvet regulators and revealed to regulate asexual and sexual development and mycotoxin production including the most carcinogenic mycotoxin aflatoxin [84]. The deletion of the veA gene decreased the production of conidia and completely blocked sclerotia formation. The veA null mutant was also unable to produce aflatoxins and aflatrem. Moreover, VeA regulates the mRNA expression of genes associated with various secondary metabolite production, such as aflatoxin, aflatrem, and asparasone [84,85]. Similar to A. nidulans VeA, A. flavus VeA interacts with VelB and LaeA to form the VelB-VeA-LaeA complex regulating sclerotia formation and aflatoxin production [68]. Of note, the deletion of veA or velB, but not laeA, resulted in the impaired conidiation, implying the positive regulation of VeA-VelB on asexual development in A. flavus [68]. The VosA-VelB heterodimer is required for proper trehalose biosynthesis and tolerance of conidia to various stresses. The newly identified VelD plays a role in aflatoxin production as the ΔvelD mutant showed no aflatoxin production [83]. In the opportunistic human pathogenic fungus A. fumigatus, velvet family regulators except VelC are required for proper asexual development. The veA, velB, or vosA null mutants exhibited asexual development even in the liquid submerged culture, where only vegetative growth occurs for the WT and the accumulation of high brlA mRNA levels, indicating a repressive role of these velvet regulators in conidiation. In addition, VeA positively regulates the production of gliotoxin, which is known to inhibit the human immune response. Unlike A. nidulans and A. flavus, the roles of the VelB-VeA-LaeA complex in A. fumigatus are not clear yet; however, the cross-species complementation analysis suggests that the VelB-VeA-LaeA complex of A. fumigatus plays a similar role with those of A. nidulans in that the introduction of the A. nidulans veA gene into the A. fumigatus ΔveA restored the normal phenotypes in A. fumigatus. The VosA-VelB complex is necessary for spore viability, trehalose biosynthesis, and tolerance of conidia to UV and oxidative stresses [67].
3.4. LaeA, a Global Regulator of Secondary Metabolism
Secondary metabolism is inseparable from fungal growth and development. Secondary metabolites have received much attention due to their broad spectrum of pharmaceutical and/or toxic properties: antibiotic, antiviral, antitumor, and immunosuppressive activities, and phytotoxic and mycotoxic activities [4,86,87]. Two decades ago, Butchko et al. [88] performed mutagenesis screening on 23 mutants that exhibited loss of sterigmatocystin (ST) production but normal asexual development in A. nidulans to reveal genes that are specific for the regulation of secondary metabolism. Thereafter, Bok and Keller investigated one of these mutants and identified a novel nuclear protein, LaeA, as a global regulator of secondary metabolism in Aspergillus (Figure 6) [87].
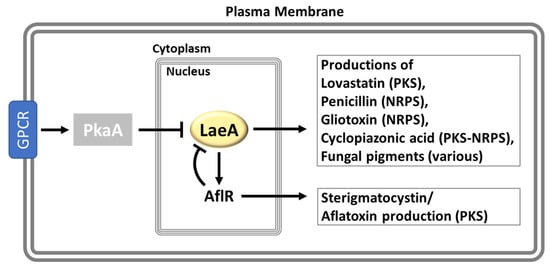
Figure 6.
The functions of LaeA in Aspergillus. See the “LaeA, a Global Regulator of Secondary Metabolism” section.
In A. nidulans, LaeA is required not only for the biosynthesis of a large array of secondary metabolites (SM) but also for the proper expression of corresponding SM biosynthetic gene clusters. The deletion of laeA inhibited the production of ST, the β-lactam antibiotics penicillin (PN), the anti-hypercholesterolaemic agent lovastatin (LOV), and the biosynthesis of mycelial pigments, which is a visually noticeable phenotype of ΔlaeA. The laeA null mutant exhibited a near absent mRNA expression of the aflR and stcU genes encoding a transcription factor and a biosynthetic enzyme required for ST production. Furthermore, the transcriptional profiling analysis of 26 genes consisting of the entire ST biosynthetic gene cluster elucidated that the transcriptional regulation of LaeA is ST cluster-specific, as the genes adjacent to the ST cluster are not affected. To understand the effect of LaeA in the production of LOV, Bok and Keller introduced the partial LOV cluster of A. terreus into the A. nidulans ΔlaeA, producing the LOV intermediate monocolin J (MONJ). The ΔlaeA/LOV+ strain displayed reduced mRNA levels of both lovE (encoding a LOV-specific Zn2Cys6 transcription factor) and lovC (a LOV biosynthetic gene), and diminished MONJ production as well. Overexpression of laeA elevated the expression levels of genes required for PN, and LOV biosynthesis (ipnA, lovE, and lovC) and the production of corresponding secondary metabolites, while, interestingly, ST production was unaffected [87].
A. flavus LaeA exhibits mostly similar functions consistent with A. nidulans LaeA yet also plays distinct roles in growth and sexual development. The ΔlaeA mutant lost the ability to produce many secondary metabolites including aflatoxin B1 and B2, cyclopiazonic acid, kojic acid (on YES media), and oryzachlorin (on DG18 media). On the other hand, overexpression of laeA led to the enhancement of some secondary metabolite productions, which are not typically observed in the WT. The sclerotia-specific metabolites Paspaline/paspalinine, aflatrem, and aflavinines were produced exclusively in the OE::laeA. This phenomenon is highly correlated with the increased sclerotia formation in the OE::laeA strain. Along with the effect in sclerotia production, LaeA affects seed colonization and lipase activity, closely related to the pathogenicity of A. flavus [89].
The opportunistic human pathogen, A. fumigatus, has been extensively studied due to its notorious virulence in humans constituting the majority of invasive aspergillosis in immunocompromised individuals [90]. Secondary metabolites including toxins and melanins have been recognized as virulence factors of invasive aspergillosis. Deletion of laeA suppressed not only the production of multiple secondary metabolites including the immunotoxin gliotoxin but also the expression of 13 SM biosynthetic gene clusters including A. fumigatus-specific mycotoxin clusters. The transcriptomic profiling analyses of WT, ΔlaeA, and C’laeA strains revealed that LaeA positively controls the expression of up to 40% of major classes of SM biosynthetic genes such as nonribosomal peptide synthetases, polyketide synthases, and P450 monooxygenases [91]. Regarding the effect of LaeA on virulence, two A. fumigatus ΔlaeA strains exhibited decreased virulence in the mouse pulmonary model, which is attributed to the reduced killing of neutrophil cells. These suggest a strong correlation between LaeA-mediated toxin production and invasive aspergillosis development by A. fumigatus [92,93].
4. NsdD, a Key Regulator of Conidiation and Sexual Development
The general life cycle of Aspergillus begins with vegetative growth. Spores start to form small germ tubes (germlings) and these tubes elongate in a highly-polarized manner resulting in hyphal growth. Under certain favorable conditions, Aspergilli initiate asexual or sexual reproductive processes. Aspergillus species primarily reproduce through asexual sporulation (conidiation), while few of them can reproduce via sexual development.
Asexual development (conidiation) in Aspergilli takes place via orchestrated gene expression of multiple positive and negative regulators. In order to initiate conidiation, upstream activators induce the activation of brlA, which encodes C2H2 zinc finger TF [94,95], initiating the development of conidiophore and activating the expression of abaA. Then, the AbaA and WetA play crucial roles in conidiophore maturation during the middle and late stages of conidiation, respectively. This central regulatory pathway (brlA → abaA → wetA) acts in concert with other genes to control conidiation-specific gene expression, and determine the order of gene activation during development and spore maturation [96]. During this asexual stage in the lifecycle, Aspergillus species produce multicellular reproductive organs, termed conidiophores, each of which produces multiple chains of non-motile conidia.
Previous studies revealed that the evolutionarily conserved GATA-type transcription factor (TF) NsdD acts as a key negative regulator of asexual development by downregulating the expression of brlA in Aspergillus (Figure 7) [14,97]. The NsdD directly binds to the promoter regions of the brlA gene and represses brlA expression in concert with another repressor VosA. The deletion of nsdD resulted in accelerated and precocious activation of conidiation; the mutant even produced asexual developmental organs under liquid submerged cultures where conidiation never takes place in the WT and exhibited the increased production of conidia on solid media. NsdD also plays a significant role in conidiophore morphogenesis. Deletion of nsdD resulted in abnormal hyphal branching during vegetative growth (data not shown). The nsdD null mutant in A. flavus displayed the formation of approximately 10 times shorter and 4 times smaller conidiophores that resemble closer to those of A. nidulans WT [14,98]. Moreover, NsdD regulates mycotoxin production including sterigmatocystin (ST) and aflatoxin (AF) in A. nidulans and A. flavus, respectively. Furthermore, previous studies have found that NsdD is required for proper sexual development. The deletion of nsdD resulted in no fruiting body formation, even under the sexual development promoting conditions. In contrast, overexpression of nsdD led to the increased formation of fruiting bodies and displayed resistance to certain inhibitory effects on sexual fruiting in Aspergillus [98,99,100]. Although the pleiotropic characteristics of NsdD regarding the development and metabolism of the genus Aspergillus have been well studied during the last two decades, the regulatory mechanism underlying how the single GATA-type TF NsdD governs all distinct aspects of fungal biology remains to be investigated.
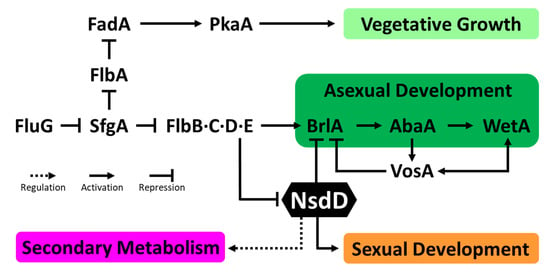
Figure 7.
A genetic model for growth, developmental, and metabolic control in Aspergillus. An arrow with a solid line indicates a positive regulation (activation) in a relationship, an arrow with a dotted line indicates an unspecified regulation (can be activating or repressing), and a blunt-ended line indicates a repressive role in the relationship.
5. Conclusions
In filamentous fungi Aspergillus species, biological and molecular processes are orchestrated by various temporal and spatial combinations of signal transduction mediators, transcription factors, regulators, and genes. In this review, among a large array of developmental and metabolic regulators identified in aspergilli, we focused on the most important upstream regulators that are known for their global impacts on Aspergillus biology, then summarized their specific roles on growth, development, and secondary metabolism. Due to their significances in fungal development and metabolism, a variety of studies have been conducted; thus, researchers now have a better understanding on their biological roles, but the detailed mechanisms of how individual regulators exert such diverse effects on different aspects of Aspergillus biology have yet to be unveiled. Genome-wide gene regulatory network analysis will be necessary to identify genes that are under the direct or indirect control of theses regulators, and to understand the relationship between direct and indirect target genes. The network study will provide valuable information upon which to understand the underlying regulatory mechanisms of main upstream regulators in governing development and metabolism of Aspergillus species.
Author Contributions
Conceptualization, H.M., K.-H.H. and J.-H.Y.; writing—original draft preparation, H.M.; writing—review and editing, H.M., K.-H.H. and J.-H.Y.; funding acquisition, K.-H.H. and J.-H.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Institute of Food and Agriculture and the National Institute of Agricultural Sciences (PJ015298) to J.-H.Y., United States Department of Agriculture and Hatch project (7000326) to J.-H.Y., and Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2017R1D1A3B06035312) to H.-K.H. H.M. was in part supported by the William H. Peterson Fellowship from the Department of Bacteriology at UW-Madison and the Robert H. and Carol L. Deibel Distinguished Graduate Fellowship from the Food Research Institute at UW-Madison.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Casselton, L.; Zolan, M. The art and design of genetic screens: Filamentous fungi. Nat. Rev. Genet. 2002, 3, 683–697. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J. An overview of the genus Aspergillus. In Aspergillus Molecular Biology and Genomics; Caister Academic Press: Norfolk, UK, 2010; pp. 1–17. [Google Scholar]
- Geiser, D.M. Sexual structures in Aspergillus: Morphology, importance and genomics. Med. Mycol. 2009, 47, S21–S26. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar] [CrossRef]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Keller, N. Regulation of secondary metabolism in filamentous fungi. Annu. Rev. Phytopathol. 2005, 43, 437–458. [Google Scholar] [CrossRef] [PubMed]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Wu, M.Y.; Mead, M.E.; Lee, M.K.; Neuhaus, G.F.; Adpressa, D.A.; Martien, J.I.; Son, Y.E.; Moon, H.; Amador-Noguez, D.; Han, K.H.; et al. Transcriptomic, Protein-DNA Interaction, and Metabolomic Studies of VosA, VelB, and WetA in Aspergillus nidulans Asexual Spores. mBio 2021, 12, e03128-20. [Google Scholar] [CrossRef]
- Park, H.-S.; Yu, J.-H. Velvet regulators in Aspergillus spp. Microbiol. Biotechnol. Lett. 2016, 44, 409–419. [Google Scholar] [CrossRef]
- Cho, H.J.; Son, S.H.; Chen, W.; Son, Y.E.; Lee, I.; Yu, J.H.; Park, H.S. Regulation of Conidiogenesis in Aspergillus flavus. Cells 2022, 11, 2796. [Google Scholar] [CrossRef]
- Ojeda-López, M.; Chen, W.; Eagle, C.E.; Gutiérrez, G.; Jia, W.L.; Swilaiman, S.S.; Huang, Z.; Park, H.S.; Yu, J.H.; Cánovas, D.; et al. Evolution of asexual and sexual reproduction in the aspergilli. Stud. Mycol. 2018, 91, 37–59. [Google Scholar] [CrossRef]
- Park, H.-S.; Lee, M.-K.; Han, K.-H.; Kim, M.-J.; Yu, J.-H. Developmental decisions in Aspergillus nidulans. In Biology of the Fungal Cell; Springer: Berlin/Heidelberg, Germany, 2019; pp. 63–80. [Google Scholar]
- Etxebeste, O.; Otamendi, A.; Garzia, A.; Espeso, E.A.; Cortese, M.S. Rewiring of transcriptional networks as a major event leading to the diversity of asexual multicellularity in fungi. Crit. Rev. Microbiol. 2019, 45, 548–563. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Kwon, N.J.; Lee, I.S.; Jung, S.; Kim, S.C.; Yu, J.H. Negative regulation and developmental competence in Aspergillus. Sci. Rep. 2016, 6, 28874. [Google Scholar] [CrossRef] [PubMed]
- Maller, J.L. Signal transduction. Fishing at the cell surface. Science 2003, 300, 594–595. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Hsueh, Y.P.; Heitman, J. Magnificent seven: Roles of G protein-coupled receptors in extracellular sensing in fungi. FEMS Microbiol. Rev. 2008, 32, 1010–1032. [Google Scholar] [CrossRef]
- Lengeler, K.B.; Davidson, R.C.; D’Souza, C.; Harashima, T.; Shen, W.C.; Wang, P.; Pan, X.; Waugh, M.; Heitman, J. Signal transduction cascades regulating fungal development and virulence. Microbiol. Mol. Biol. Rev. 2000, 64, 746–785. [Google Scholar] [CrossRef]
- Rispail, N.; Soanes, D.M.; Ant, C.; Czajkowski, R.; Grünler, A.; Huguet, R.; Perez-Nadales, E.; Poli, A.; Sartorel, E.; Valiante, V.; et al. Comparative genomics of MAP kinase and calcium-calcineurin signalling components in plant and human pathogenic fungi. Fungal Genet. Biol. 2009, 46, 287–298. [Google Scholar] [CrossRef]
- Affeldt, K.J.; Carrig, J.; Amare, M.; Keller, N.P. Global survey of canonical Aspergillus flavus G protein-coupled receptors. mBio 2014, 5, e01501–e01514. [Google Scholar] [CrossRef]
- Li, L.; Wright, S.J.; Krystofova, S.; Park, G.; Borkovich, K.A. Heterotrimeric G protein signaling in filamentous fungi. Annu. Rev. Microbiol. 2007, 61, 423–452. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.A.; Schrevens, S.; van Dijck, P.; Goldman, G.H. Fungal G-protein-coupled receptors: Mediators of pathogenesis and targets for disease control. Nat. Microbiol. 2018, 3, 402–414. [Google Scholar] [CrossRef] [PubMed]
- DeZwaan, T.M.; Carroll, A.M.; Valent, B.; Sweigard, J.A. Magnaporthe grisea pth11p is a novel plasma membrane protein that mediates appressorium differentiation in response to inductive substrate cues. Plant Cell 1999, 11, 2013–2030. [Google Scholar] [CrossRef]
- Dilks, T.; Halsey, K.; De Vos, R.P.; Hammond-Kosack, K.E.; Brown, N.A. Non-canonical fungal G-protein coupled receptors promote Fusarium head blight on wheat. PLoS Pathog. 2019, 15, e1007666. [Google Scholar] [CrossRef] [PubMed]
- Gehrke, A.; Heinekamp, T.; Jacobsen, I.D.; Brakhage, A.A. Heptahelical receptors GprC and GprD of Aspergillus fumigatus Are essential regulators of colony growth, hyphal morphogenesis, and virulence. Appl. Environ. Microbiol. 2010, 76, 3989–3998. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.G.; Kim, S.S.; Yu, J.H.; Shin, K.S. Characterization of gprK Encoding a Putative Hybrid G-Protein-Coupled Receptor in Aspergillus fumigatus. PLoS ONE 2016, 11, e0161312. [Google Scholar] [CrossRef] [PubMed]
- Filho, A.; Brancini, G.T.P.; de Castro, P.A.; Valero, C.; Ferreira Filho, J.A.; Silva, L.P.; Rocha, M.C.; Malavazi, I.; Pontes, J.G.M.; Fill, T.; et al. Aspergillus fumigatus G-Protein Coupled Receptors GprM and GprJ Are Important for the Regulation of the Cell Wall Integrity Pathway, Secondary Metabolite Production, and Virulence. mBio 2020, 11, e02458-20. [Google Scholar] [CrossRef] [PubMed]
- Bardwell, L. A walk-through of the yeast mating pheromone response pathway. Peptides 2005, 26, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Borkovich, K.A. GPR-4 is a predicted G-protein-coupled receptor required for carbon source-dependent asexual growth and development in Neurospora crassa. Eukaryot Cell 2006, 5, 1287–1300. [Google Scholar] [CrossRef]
- Chung, K.S.; Won, M.; Lee, S.B.; Jang, Y.J.; Hoe, K.L.; Kim, D.U.; Lee, J.W.; Kim, K.W.; Yoo, H.S. Isolation of a novel gene from Schizosaccharomyces pombe: stm1+ encoding a seven-transmembrane loop protein that may couple with the heterotrimeric Galpha 2 protein, Gpa2. J. Biol. Chem. 2001, 276, 40190–40201. [Google Scholar] [CrossRef]
- Johnson, R.L.; Gundersen, R.; Hereld, D.; Pitt, G.S.; Tugendreich, S.; Saxe, C.L.; Kimmel, A.R.; Devreotes, P.N. G-protein-linked signaling pathways mediate development in Dictyostelium. Cold Spring Harb. Symp. Quant. Biol. 1992, 57, 169–176. [Google Scholar] [CrossRef]
- Insall, R.H.; Soede, R.D.; Schaap, P.; Devreotes, P.N. Two cAMP receptors activate common signaling pathways in Dictyostelium. Mol. Biol. Cell 1994, 5, 703–711. [Google Scholar] [CrossRef]
- Swaney, K.F.; Huang, C.H.; Devreotes, P.N. Eukaryotic chemotaxis: A network of signaling pathways controls motility, directional sensing, and polarity. Annu. Rev. Biophys. 2010, 39, 265–289. [Google Scholar] [CrossRef]
- Krystofova, S.; Borkovich, K.A. The predicted G-protein-coupled receptor GPR-1 is required for female sexual development in the multicellular fungus Neurospora crassa. Eukaryot Cell 2006, 5, 1503–1516. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.G.; Willard, F.S.; Huang, J.; Liang, J.; Chasse, S.A.; Jones, A.M.; Siderovski, D.P. A seven-transmembrane RGS protein that modulates plant cell proliferation. Science 2003, 301, 1728–1731. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Bond, J.; Thomas, P. Identification, classification, and partial characterization of genes in humans and other vertebrates homologous to a fish membrane progestin receptor. Proc. Natl. Acad. Sci. USA 2003, 100, 2237–2242. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P. Characteristics of membrane progestin receptor alpha (mPRalpha) and progesterone membrane receptor component 1 (PGMRC1) and their roles in mediating rapid progestin actions. Front. Neuroendocrinol. 2008, 29, 292–312. [Google Scholar] [CrossRef] [PubMed]
- Bieszke, J.A.; Spudich, E.N.; Scott, K.L.; Borkovich, K.A.; Spudich, J.L. A eukaryotic protein, NOP-1, binds retinal to form an archaeal rhodopsin-like photochemically reactive pigment. Biochemistry 1999, 38, 14138–14145. [Google Scholar] [CrossRef]
- Lafon, A.; Han, K.H.; Seo, J.A.; Yu, J.H.; d’Enfert, C. G-protein and cAMP-mediated signaling in aspergilli: A genomic perspective. Fungal Genet. Biol. 2006, 43, 490–502. [Google Scholar] [CrossRef]
- Seo, J.A.; Han, K.H.; Yu, J.H. The gprA and gprB genes encode putative G protein-coupled receptors required for self-fertilization in Aspergillus nidulans. Mol. Microbiol. 2004, 53, 1611–1623. [Google Scholar] [CrossRef]
- Han, K.H.; Seo, J.A.; Yu, J.H. A putative G protein-coupled receptor negatively controls sexual development in Aspergillus nidulans. Mol. Microbiol. 2004, 51, 1333–1345. [Google Scholar] [CrossRef]
- Simon, M.I.; Strathmann, M.P.; Gautam, N. Diversity of G proteins in signal transduction. Science 1991, 252, 802–808. [Google Scholar] [CrossRef]
- Yu, J.H.; Wieser, J.; Adams, T.H. The Aspergillus FlbA RGS domain protein antagonizes G protein signaling to block proliferation and allow development. EMBO J. 1996, 15, 5184–5190. [Google Scholar] [CrossRef]
- Rosén, S.; Yu, J.H.; Adams, T.H. The Aspergillus nidulans sfaD gene encodes a G protein beta subunit that is required for normal growth and repression of sporulation. EMBO J. 1999, 18, 5592–5600. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.A.; Han, K.H.; Yu, J.H. Multiple roles of a heterotrimeric G-protein gamma-subunit in governing growth and development of Aspergillus nidulans. Genetics 2005, 171, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Neves, S.R.; Ram, P.T.; Iyengar, R. G protein pathways. Science 2002, 296, 1636–1639. [Google Scholar] [CrossRef] [PubMed]
- McCudden, C.R.; Hains, M.D.; Kimple, R.J.; Siderovski, D.P.; Willard, F.S. G-protein signaling: Back to the future. Cell. Mol. Life Sci. 2005, 62, 551–577. [Google Scholar] [CrossRef]
- Hicks, J.K.; Yu, J.H.; Keller, N.P.; Adams, T.H. Aspergillus sporulation and mycotoxin production both require inactivation of the FadA G alpha protein-dependent signaling pathway. EMBO J. 1997, 16, 4916–4923. [Google Scholar] [CrossRef]
- Choi, Y.H.; Lee, N.Y.; Kim, S.S.; Park, H.S.; Shin, K.S. Comparative Characterization of G Protein α Subunits in Aspergillus fumigatus. Pathogens 2020, 9, 272. [Google Scholar] [CrossRef]
- Chang, M.H.; Chae, K.S.; Han, D.M.; Jahng, K.Y. The GanB Galpha-protein negatively regulates asexual sporulation and plays a positive role in conidial germination in Aspergillus nidulans. Genetics 2004, 167, 1305–1315. [Google Scholar] [CrossRef]
- Lafon, A.; Seo, J.A.; Han, K.H.; Yu, J.H.; d’Enfert, C. The heterotrimeric G-protein GanB(alpha)-SfaD(beta)-GpgA(gamma) is a carbon source sensor involved in early cAMP-dependent germination in Aspergillus nidulans. Genetics 2005, 171, 71–80. [Google Scholar] [CrossRef]
- Kasahara, S.; Nuss, D.L. Targeted disruption of a fungal G-protein beta subunit gene results in increased vegetative growth but reduced virulence. Mol. Plant Microbe Interact. 1997, 10, 984–993. [Google Scholar] [CrossRef]
- Yang, Q.; Poole, S.I.; Borkovich, K.A. A G-protein beta subunit required for sexual and vegetative development and maintenance of normal G alpha protein levels in Neurospora crassa. Eukaryot. Cell 2002, 1, 378–390. [Google Scholar] [CrossRef]
- Lukov, G.L.; Hu, T.; McLaughlin, J.N.; Hamm, H.E.; Willardson, B.M. Phosducin-like protein acts as a molecular chaperone for G protein betagamma dimer assembly. EMBO J. 2005, 24, 1965–1975. [Google Scholar] [CrossRef] [PubMed]
- Lukov, G.L.; Baker, C.M.; Ludtke, P.J.; Hu, T.; Carter, M.D.; Hackett, R.A.; Thulin, C.D.; Willardson, B.M. Mechanism of assembly of G protein betagamma subunits by protein kinase CK2-phosphorylated phosducin-like protein and the cytosolic chaperonin complex. J. Biol. Chem. 2006, 281, 22261–22274. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, S.; Wang, P.; Nuss, D.L. Identification of bdm-1, a gene involved in G protein beta-subunit function and alpha-subunit accumulation. Proc. Natl. Acad. Sci. USA 2000, 97, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.A.; Yu, J.H. The phosducin-like protein PhnA is required for Gbetagamma-mediated signaling for vegetative growth, developmental control, and toxin biosynthesis in Aspergillus nidulans. Eukaryot. Cell 2006, 5, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Ross, E.M.; Wilkie, T.M. GTPase-activating proteins for heterotrimeric G proteins: Regulators of G protein signaling (RGS) and RGS-like proteins. Annu. Rev. Biochem. 2000, 69, 795–827. [Google Scholar] [CrossRef]
- Han, K.H.; Seo, J.A.; Yu, J.H. Regulators of G-protein signalling in Aspergillus nidulans: RgsA downregulates stress response and stimulates asexual sporulation through attenuation of GanB (Galpha) signalling. Mol. Microbiol. 2004, 53, 529–540. [Google Scholar] [CrossRef]
- Burchett, S.A. Regulators of G protein signaling: A bestiary of modular protein binding domains. J. Neurochem. 2000, 75, 1335–1351. [Google Scholar] [CrossRef]
- Kwon, N.J.; Park, H.S.; Jung, S.; Kim, S.C.; Yu, J.H. The putative guanine nucleotide exchange factor RicA mediates upstream signaling for growth and development in Aspergillus. Eukaryot. Cell 2012, 11, 1399–1412. [Google Scholar] [CrossRef]
- Mooney, J.L.; Hassett, D.E.; Yager, L.N. Genetic analysis of suppressors of the veA1 mutation in Aspergillus nidulans. Genetics 1990, 126, 869–874. [Google Scholar] [CrossRef]
- Kim, H.; Han, K.; Kim, K.; Han, D.; Jahng, K.; Chae, K. The veA gene activates sexual development in Aspergillus nidulans. Fungal Genet. Biol. 2002, 37, 72–80. [Google Scholar] [CrossRef]
- Kato, N.; Brooks, W.; Calvo, A.M. The expression of sterigmatocystin and penicillin genes in Aspergillus nidulans is controlled by veA, a gene required for sexual development. Eukaryot. Cell 2003, 2, 1178–1186. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.M.; Bok, J.; Brooks, W.; Keller, N.P. veA is required for toxin and sclerotial production in Aspergillus parasiticus. Appl. Environ. Microbiol. 2004, 70, 4733–4739. [Google Scholar] [CrossRef] [PubMed]
- Amaike, S.; Keller, N.P. Distinct roles for VeA and LaeA in development and pathogenesis of Aspergillus flavus. Eukaryot. Cell 2009, 8, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Dhingra, S.; Andes, D.; Calvo, A.M. VeA regulates conidiation, gliotoxin production, and protease activity in the opportunistic human pathogen Aspergillus fumigatus. Eukaryot. Cell 2012, 11, 1531–1543. [Google Scholar] [CrossRef]
- Park, H.S.; Bayram, O.; Braus, G.H.; Kim, S.C.; Yu, J.H. Characterization of the velvet regulators in Aspergillus fumigatus. Mol. Microbiol. 2012, 86, 937–953. [Google Scholar] [CrossRef]
- Chang, P.K.; Scharfenstein, L.L.; Li, P.; Ehrlich, K.C. Aspergillus flavus VelB acts distinctly from VeA in conidiation and may coordinate with FluG to modulate sclerotial production. Fungal Genet. Biol. 2013, 58–59, 71–79. [Google Scholar] [CrossRef]
- Ni, M.; Yu, J.H. A novel regulator couples sporogenesis and trehalose biogenesis in Aspergillus nidulans. PLoS ONE 2007, 2, e970. [Google Scholar] [CrossRef]
- Käfer, E. Origins of translocations in Aspergillus nidulans. Genetics 1965, 52, 217–232. [Google Scholar] [CrossRef]
- Stinnett, S.M.; Espeso, E.A.; Cobeño, L.; Araújo-Bazán, L.; Calvo, A.M. Aspergillus nidulans VeA subcellular localization is dependent on the importin alpha carrier and on light. Mol. Microbiol. 2007, 63, 242–255. [Google Scholar] [CrossRef]
- Rogers, S.; Wells, R.; Rechsteiner, M. Amino acid sequences common to rapidly degraded proteins: The PEST hypothesis. Science 1986, 234, 364–368. [Google Scholar] [CrossRef]
- Timberlake, W.E. Molecular genetics of Aspergillus development. Annu. Rev. Genet. 1990, 24, 5–36. [Google Scholar] [CrossRef] [PubMed]
- Yager, L.N. Early developmental events during asexual and sexual sporulation in Aspergillus nidulans. Biotechnology 1992, 23, 19–41. [Google Scholar] [PubMed]
- Spröte, P.; Brakhage, A.A. The light-dependent regulator velvet A of Aspergillus nidulans acts as a repressor of the penicillin biosynthesis. Arch. Microbiol. 2007, 188, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Sarikaya-Bayram, Ö.; Palmer, J.M.; Keller, N.; Braus, G.H.; Bayram, Ö. One Juliet and four Romeos: VeA and its methyltransferases. Front. Microbiol. 2015, 6, 1. [Google Scholar] [CrossRef]
- Bayram, O.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.J.; Keller, N.P.; Yu, J.H.; et al. VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef]
- Ahmed, Y.L.; Gerke, J.; Park, H.S.; Bayram, Ö.; Neumann, P.; Ni, M.; Dickmanns, A.; Kim, S.C.; Yu, J.H.; Braus, G.H.; et al. The velvet family of fungal regulators contains a DNA-binding domain structurally similar to NF-κB. PLoS Biol. 2013, 11, e1001750. [Google Scholar] [CrossRef]
- Park, H.S.; Man Yu, Y.; Lee, M.K.; Jae Maeng, P.; Chang Kim, S.; Yu, J.H. Velvet-mediated repression of β-glucan synthesis in Aspergillus nidulans spores. Sci. Rep. 2015, 5, 10199. [Google Scholar] [CrossRef]
- Atoui, A.; Kastner, C.; Larey, C.M.; Thokala, R.; Etxebeste, O.; Espeso, E.A.; Fischer, R.; Calvo, A.M. Cross-talk between light and glucose regulation controls toxin production and morphogenesis in Aspergillus nidulans. Fungal Genet. Biol. 2010, 47, 962–972. [Google Scholar] [CrossRef]
- Reyes-Dominguez, Y.; Bok, J.W.; Berger, H.; Shwab, E.K.; Basheer, A.; Gallmetzer, A.; Scazzocchio, C.; Keller, N.; Strauss, J. Heterochromatic marks are associated with the repression of secondary metabolism clusters in Aspergillus nidulans. Mol. Microbiol. 2010, 76, 1376–1386. [Google Scholar] [CrossRef]
- Sarikaya Bayram, O.; Bayram, O.; Valerius, O.; Park, H.S.; Irniger, S.; Gerke, J.; Ni, M.; Han, K.H.; Yu, J.H.; Braus, G.H. LaeA control of velvet family regulatory proteins for light-dependent development and fungal cell-type specificity. PLoS Genet. 2010, 6, e1001226. [Google Scholar] [CrossRef]
- Eom, T.J.; Moon, H.; Yu, J.H.; Park, H.S. Characterization of the velvet regulators in Aspergillus flavus. J. Microbiol. 2018, 56, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Duran, R.M.; Cary, J.W.; Calvo, A.M. Production of cyclopiazonic acid, aflatrem, and aflatoxin by Aspergillus flavus is regulated by veA, a gene necessary for sclerotial formation. Appl. Microbiol. Biotechnol. 2007, 73, 1158–1168. [Google Scholar] [CrossRef] [PubMed]
- Cary, J.W.; Harris-Coward, P.Y.; Ehrlich, K.C.; Di Mavungu, J.D.; Malysheva, S.V.; De Saeger, S.; Dowd, P.F.; Shantappa, S.; Martens, S.L.; Calvo, A.M. Functional characterization of a veA-dependent polyketide synthase gene in Aspergillus flavus necessary for the synthesis of asparasone, a sclerotium-specific pigment. Fungal Genet. Biol. 2014, 64, 25–35. [Google Scholar] [CrossRef]
- Demain, A.L.; Fang, A. The natural functions of secondary metabolites. Adv. Biochem. Eng. Biotechnol. 2000, 69, 1–39. [Google Scholar] [CrossRef]
- Bok, J.W.; Keller, N.P. LaeA, a regulator of secondary metabolism in Aspergillus spp. Eukaryot. Cell 2004, 3, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Butchko, R.A.; Adams, T.H.; Keller, N.P. Aspergillus nidulans mutants defective in stc gene cluster regulation. Genetics 1999, 153, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Kale, S.P.; Milde, L.; Trapp, M.K.; Frisvad, J.C.; Keller, N.P.; Bok, J.W. Requirement of LaeA for secondary metabolism and sclerotial production in Aspergillus flavus. Fungal Genet. Biol. 2008, 45, 1422–1429. [Google Scholar] [CrossRef]
- Latgé, J.P.; Chamilos, G. Aspergillus fumigatus and Aspergillosis in 2019. Clin. Microbiol. Rev. 2019, 33, e00140-18. [Google Scholar] [CrossRef]
- Perrin, R.M.; Fedorova, N.D.; Bok, J.W.; Cramer, R.A.; Wortman, J.R.; Kim, H.S.; Nierman, W.C.; Keller, N.P. Transcriptional regulation of chemical diversity in Aspergillus fumigatus by LaeA. PLoS Pathog. 2007, 3, e50. [Google Scholar] [CrossRef]
- Bok, J.W.; Balajee, S.A.; Marr, K.A.; Andes, D.; Nielsen, K.F.; Frisvad, J.C.; Keller, N.P. LaeA, a regulator of morphogenetic fungal virulence factors. Eukaryot. Cell 2005, 4, 1574–1582. [Google Scholar] [CrossRef]
- Sugui, J.A.; Pardo, J.; Chang, Y.C.; Müllbacher, A.; Zarember, K.A.; Galvez, E.M.; Brinster, L.; Zerfas, P.; Gallin, J.I.; Simon, M.M.; et al. Role of laeA in the Regulation of alb1, gliP, Conidial Morphology, and Virulence in Aspergillus fumigatus. Eukaryot. Cell 2007, 6, 1552–1561. [Google Scholar] [CrossRef] [PubMed]
- Adams, T.H.; Boylan, M.T.; Timberlake, W.E. brlA is necessary and sufficient to direct conidiophore development in Aspergillus nidulans. Cell 1988, 54, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Timberlake, W.E. Identification of Aspergillus brlA response elements (BREs) by genetic selection in yeast. Genetics 1993, 133, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Yu, J.H. Genetic control of asexual sporulation in filamentous fungi. Curr. Opin. Microbiol. 2012, 15, 669–677. [Google Scholar] [CrossRef]
- Lee, M.K.; Kwon, N.J.; Choi, J.M.; Lee, I.S.; Jung, S.; Yu, J.H. NsdD is a key repressor of asexual development in Aspergillus nidulans. Genetics 2014, 197, 159–173. [Google Scholar] [CrossRef]
- Cary, J.W.; Harris-Coward, P.Y.; Ehrlich, K.C.; Mack, B.M.; Kale, S.P.; Larey, C.; Calvo, A.M. NsdC and NsdD affect Aspergillus flavus morphogenesis and aflatoxin production. Eukaryot. Cell 2012, 11, 1104–1111. [Google Scholar] [CrossRef]
- Han, K.H.; Han, K.Y.; Yu, J.H.; Chae, K.S.; Jahng, K.Y.; Han, D.M. The nsdD gene encodes a putative GATA-type transcription factor necessary for sexual development of Aspergillus nidulans. Mol. Microbiol. 2001, 41, 299–309. [Google Scholar] [CrossRef]
- Szewczyk, E.; Krappmann, S. Conserved regulators of mating are essential for Aspergillus fumigatus cleistothecium formation. Eukaryot. Cell 2010, 9, 774–783. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).