
Transcriptomic Analysis Reveals JAK2/MPL-Independent Effects of Calreticulin Mutations in a C. elegans Model

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. C. Elegans Strains and Maintenance
2.2. Characterization of the Larval Development of C. Elegans Strains
2.3. RNA Extraction
2.4. Whole-Genome Transcriptomic Analysis
2.5. Quantitative Real-Time PCR (qPCR)
2.6. Statistics
3. Results
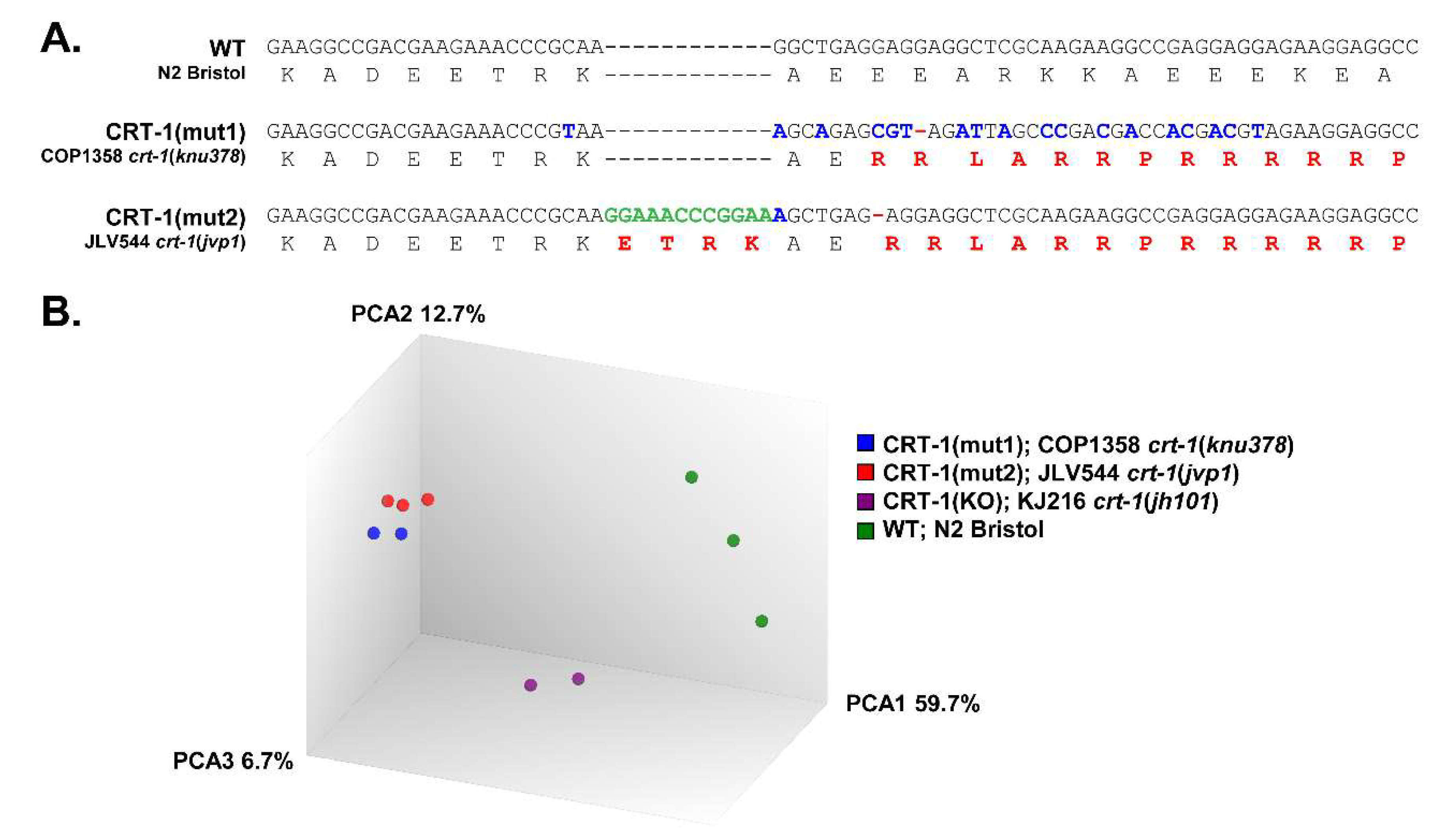
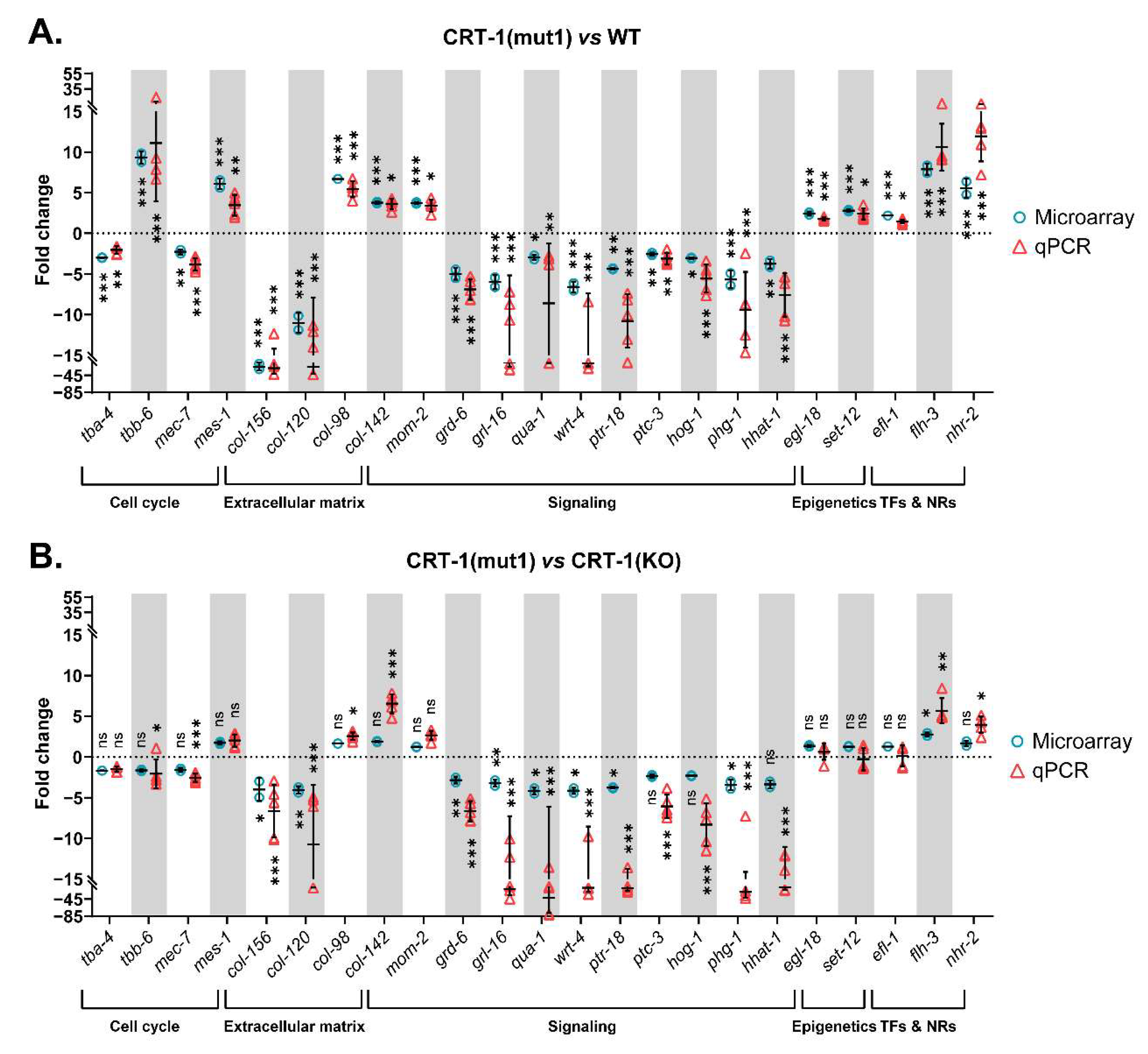
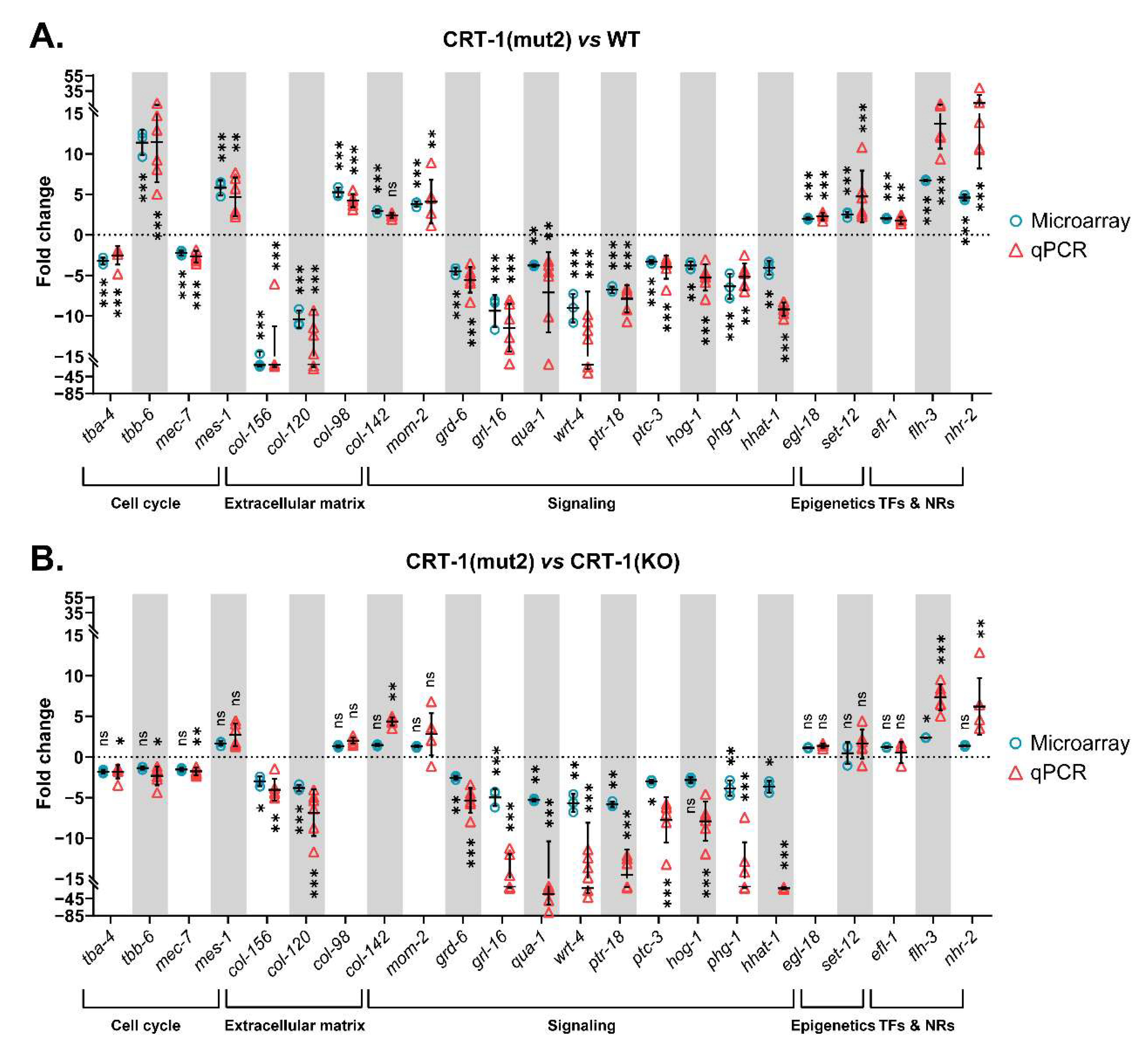
3.1. Type 1 and Type 2-like Calreticulin Mutants Behave Similarly to Each Other and Differently to the Wild-Type and Knockout Strains and Show a Transcriptional Alteration of Genes Participating in Processes That Have a Role in Cancer and MPNs
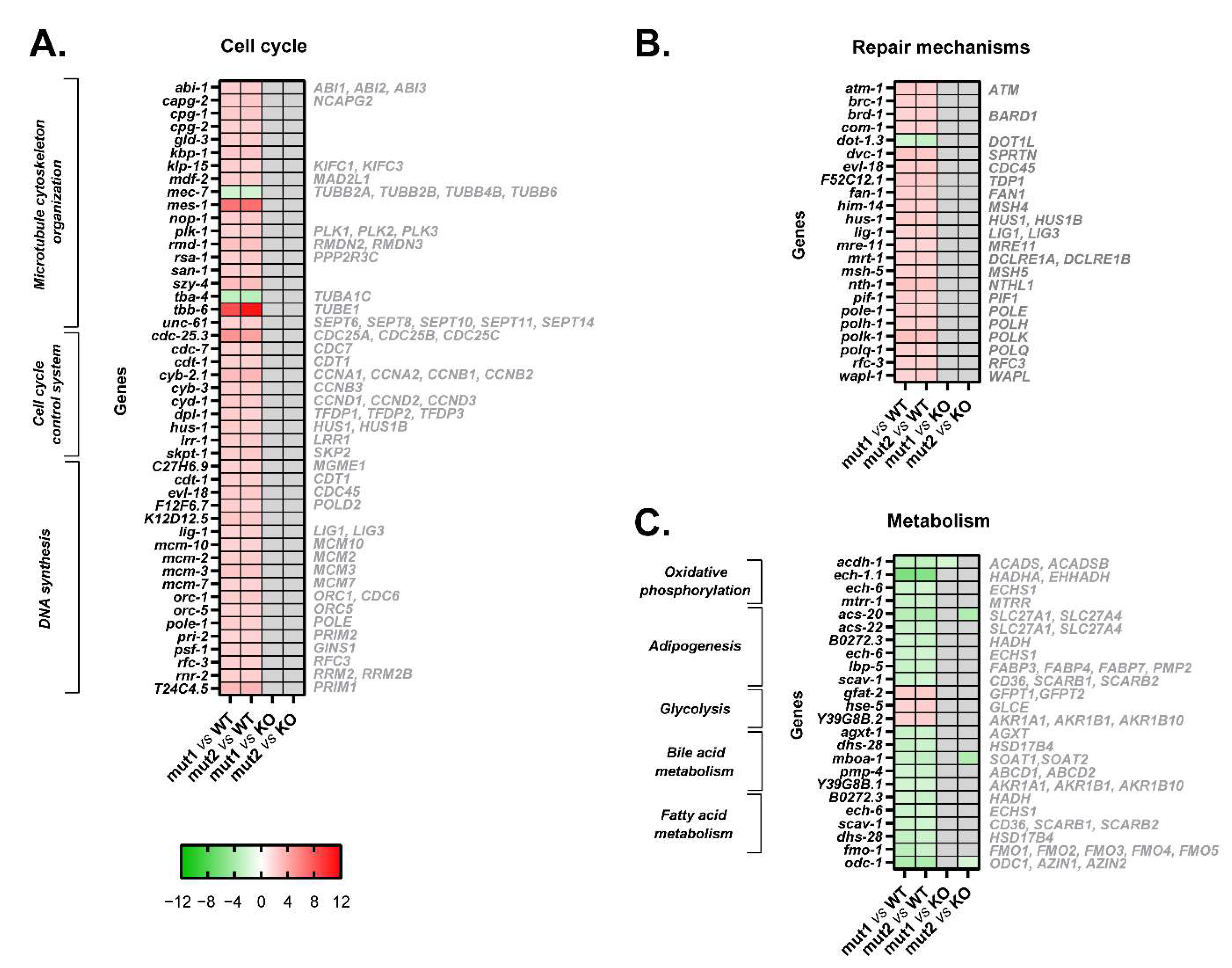
3.2. Mutant Calreticulin Induces an Aberrant Expression of Cell Cycle Players
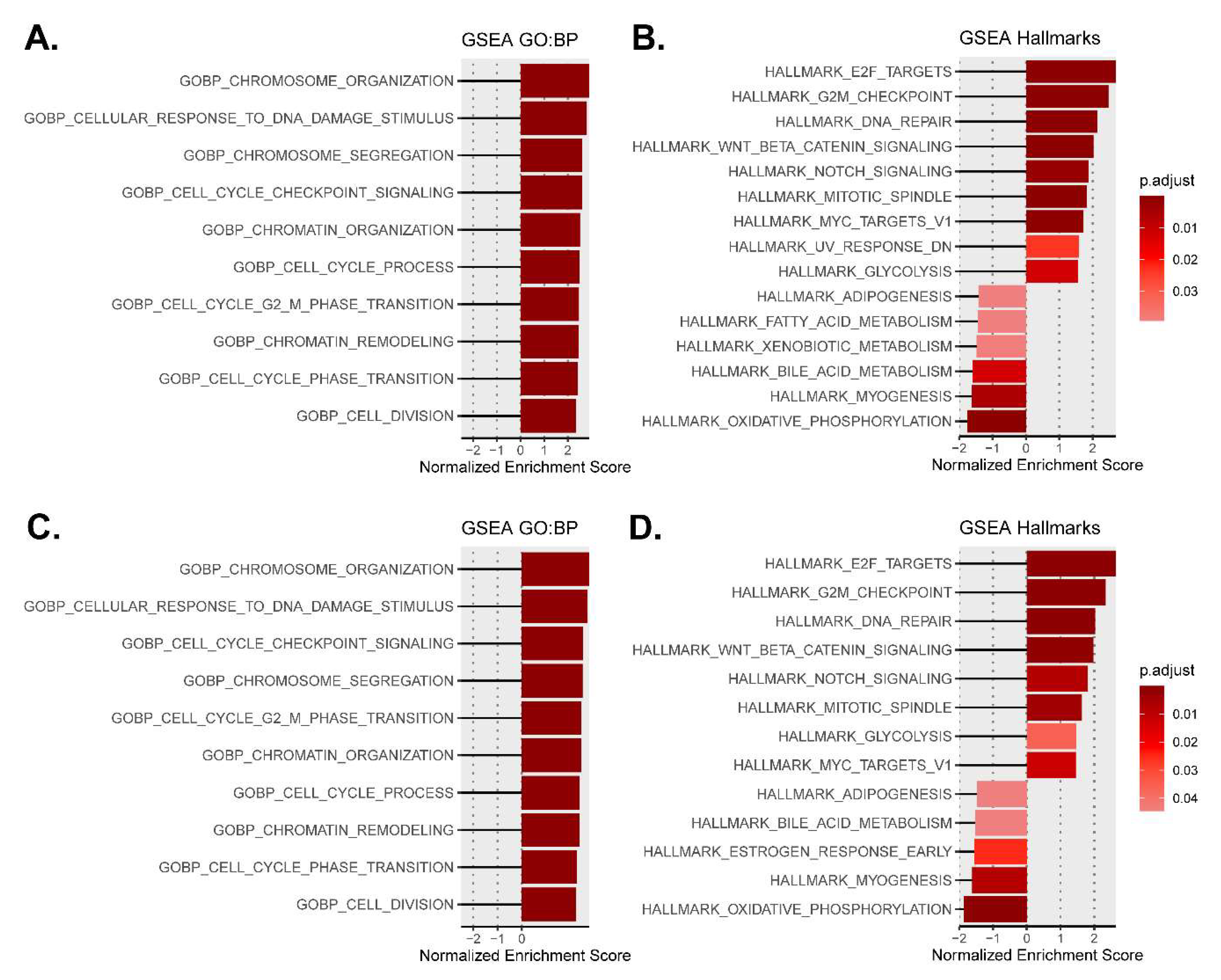
3.3. Mutant Calreticulin Induces the Activation of Repair Mechanisms
3.4. Mutant Calreticulin Dysregulates Cellular Metabolism
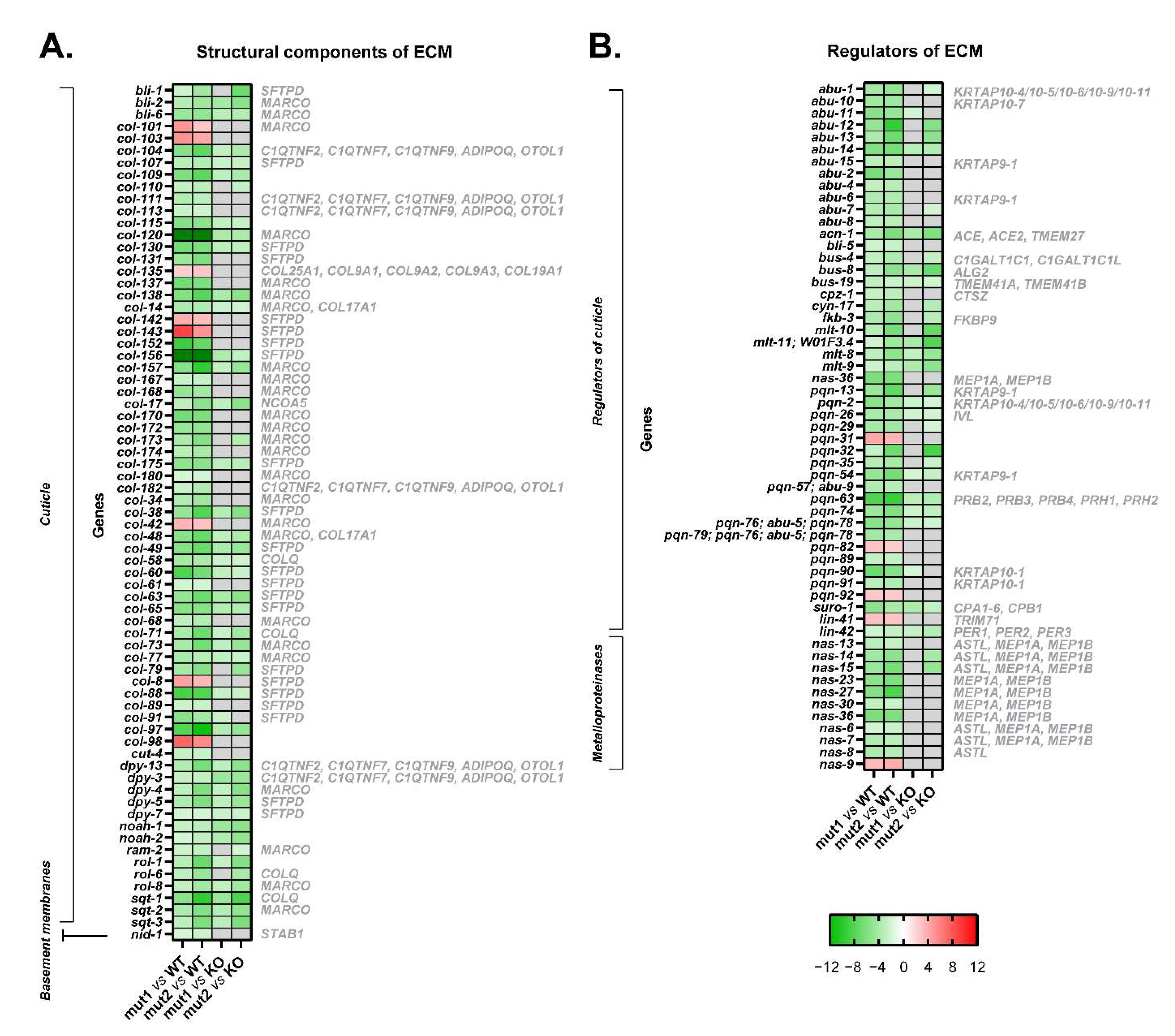
3.5. The Introduction of Both Type 1 and Type 2-like Mutations in Calreticulin Seems to Disrupt the Composition of the Extracellular Matrix
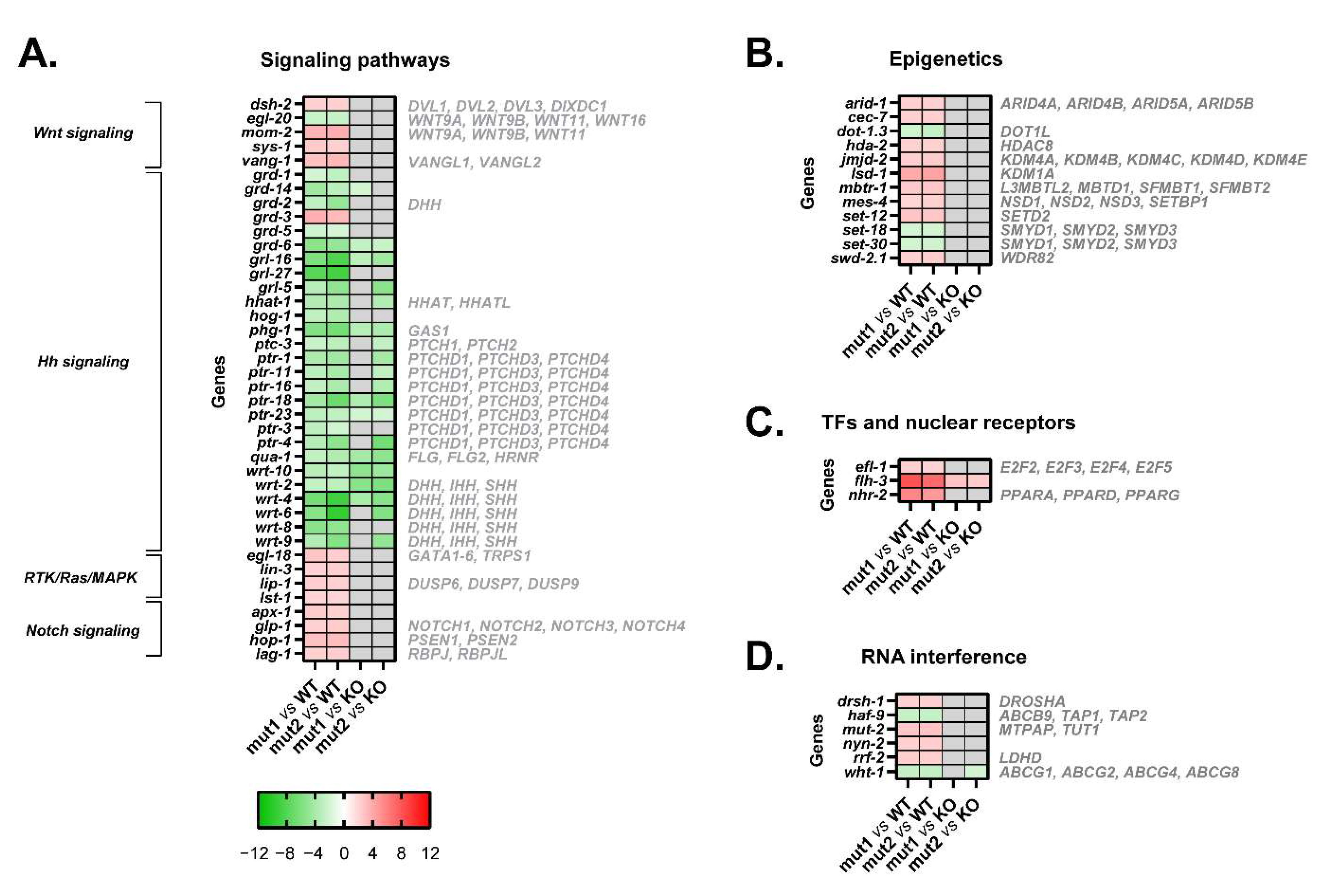
3.6. Mutant Calreticulin Worms Show a Different Expression Pattern of Genes Whose Products Participate in Signal Transduction Pathways
3.7. Mutant Calreticulin also Seems to Disrupt Epigenetic Mechanisms
3.8. Transcription Factors Could Shed Some Light on the Aberrant Mechanisms by Which Mutant Calreticulin Alters the Transcriptome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guijarro-Hernández, A.; Vizmanos, J.L. A Broad Overview of Signaling in Ph-Negative Classic Myeloproliferative Neoplasms. Cancers 2021, 13, 984. [Google Scholar] [CrossRef] [PubMed]
- Michalak, M.; Corbett, E.F.; Mesaeli, N.; Nakamura, K.; Opas, M. Calreticulin: One protein, one gene, many functions. Biochem. J. 1999, 344, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.A.; Groenendyk, J.; Michalak, M. Calreticulin signaling in health and disease. Int. J. Biochem. Cell Biol. 2012, 44, 842–846. [Google Scholar] [CrossRef]
- Araki, M.; Komatsu, N. The role of calreticulin mutations in myeloproliferative neoplasms. Int. J. Hematol. 2020, 111, 200–205. [Google Scholar] [CrossRef]
- Corsi, A.K.; Wightman, B.; Chalfie, M. A transparent window into biology: A primer on Caenorhabditis elegans. Genetics 2015, 200, 387–407. [Google Scholar] [CrossRef]
- Smith, M.J. A C. elegans gene encodes a protein homologous to mammalian calreticulin. DNA Seq. 1992, 2, 235–240. [Google Scholar] [CrossRef]
- Park, B.J.; Lee, D.G.; Yu, J.R.; Jung, S.K.; Choi, K.; Lee, J.; Lee, J.; Kim, Y.S.; Lee, J.I.; Kwon, J.; et al. Calreticulin, a calcium-binding molecular chaperone, is required for stress response and fertility in Caenorhabditis elegans. Mol. Biol. Cell 2001, 12, 2835–2845. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Singaravelu, G.; Park, B.J.; Ahnn, J. Differential requirement of unfolded protein response pathway for calreticulin expression in Caenorhabditis elegans. J. Mol. Biol. 2007, 372, 331–340. [Google Scholar] [CrossRef]
- Zahreddine, H.; Zhang, H.; Diogon, M.; Nagamatsu, Y.; Labouesse, M. CRT-1/calreticulin and the E3 ligase EEL-1/HUWE1 control hemidesmosome maturation in C. elegans development. Curr. Biol. 2010, 20, 322–327. [Google Scholar] [CrossRef]
- Wang, W.A.; Liu, W.X.; Durnaoglu, S.; Lee, S.K.; Lian, J.; Lehner, R.; Ahnn, J.; Agellon, L.B.; Michalak, M. Loss of Calreticulin Uncovers a Critical Role for Calcium in Regulating Cellular Lipid Homeostasis. Sci. Rep. 2017, 7, 5941. [Google Scholar] [CrossRef]
- Kim, W.; Underwood, R.S.; Greenwald, I.; Shaye, D.D. OrthoList 2: A new comparative genomic analysis of human and Caenorhabditis elegans genes. Genetics 2018, 210, 445–461. [Google Scholar] [CrossRef] [PubMed]
- Eder-Azanza, L.; Navarro, D.; Aranaz, P.; Novo, F.J.; Cross, N.C.; Vizmanos, J.L. Bioinformatic analyses of CALR mutations in myeloproliferative neoplasms support a role in signaling. Leukemia 2014, 28, 2106–2109. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Dolgalev, I. msigdbr: MSigDB Gene Sets for Multiple Organisms in a Tidy Data Format. R Package Version 7.5.1. 2022. Available online: https://igordot.github.io/msigdbr/ (accessed on 8 April 2022).
- Hanahan, D. Hallmarks of cancer: New dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Berkowitz, L.A.; Strome, S. MES-1, a protein required for unequal divisions of the germline in early C. elegans embryos, resembles receptor tyrosine kinases and is localized to the boundary between the germline and gut cells. Development 2000, 127, 4419–4431. [Google Scholar] [CrossRef] [PubMed]
- Ow, M.C.; Martinez, N.J.; Olsen, P.H.; Silverman, H.S.; Barrasa, M.I.; Conradt, B.; Walhout, A.J.; Ambros, V. The FLYWCH transcription factors FLH-1, FLH-2, and FLH-3 repress embryonic expression of microRNA genes in C. elegans. Genes Dev. 2008, 22, 2520–2534. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Zhang, M.; Li, Y.; Liu, Y.; Cui, Q.; Wang, N. PPARgene: A database of experimentally verified and computationally predicted PPAR target genes. PPAR Res. 2016, 2016, 6042162. [Google Scholar] [CrossRef]
- Delaval, B.; Lelièvre, H.; Birnbaum, D. Myeloproliferative disorders: The centrosome connection. Leukemia 2005, 19, 739–1744. [Google Scholar] [CrossRef]
- Lee, J.Y.; Hong, W.J.; Majeti, R.; Stearns, T. Centrosome-kinase fusions promote oncogenic signaling and disrupt centrosome function in myeloproliferative neoplasms. PLoS ONE 2014, 9, e92641. [Google Scholar] [CrossRef] [PubMed]
- Vincelette, N.D.; Moon, J.; Kuykendall, A.T.; Zhang, L.; Komrokji, R.S.; Murphy, D.; Cleveland, J.L.; Yun, S. C-MYC augments the proliferation and survival of hematopoietic stem cells and multipotent progenitors to drive myeloproliferative neoplasms. Blood 2021, 138 (Suppl. 1), 28. [Google Scholar] [CrossRef]
- Zini, R.; Guglielmelli, P.; Pietra, D.; Rumi, E.; Rossi, C.; Rontauroli, S.; Genovese, E.; Fanelli, T.; Calabresi, L.; Bianchi, E.; et al. CALR mutational status identifies different disease subtypes of essential thrombocythemia showing distinct expression profiles. Blood Cancer J. 2017, 7, 638. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, M.; Bornemann, A.; Schubert, C.; Gezer, D.; Kricheldorf, K.; Isfort, S.; Brümmendorf, T.H.; Schemionek, M.; Chatain, N.; Skorski, T.; et al. Transcriptional alteration of DNA repair genes in Philadelphia chromosome negative myeloproliferative neoplasms. Ann. Hematol. 2019, 98, 2703–2709. [Google Scholar] [CrossRef] [PubMed]
- Rao, T.N.; Hansen, N.; Hilfiker, J.; Rai, S.; Majewska, J.M.; Leković, D.; Gezer, D.; Andina, N.; Galli, S.; Cassel, T.; et al. JAK2-mutant hematopoietic cells display metabolic alterations that can be targeted to treat myeloproliferative neoplasms. Blood 2019, 134, 1832–1846. [Google Scholar] [CrossRef] [PubMed]
- Cacemiro, M.C.; Cominal, J.G.; Almeida, F.C.; Oliveira, M.C.; Sorgi, C.A.; Figueiredo-Pontes, L.L.; Faccioli, L.H.; Gardinassi, L.G.; Castro, F.A. Metabolic profile of bone marrow plasma in myeloproliferative neoplasms. Hematol. Transfus. Cell Ther. 2021, 43 (Suppl. 1), S137. [Google Scholar] [CrossRef]
- Leoncini, G.; Maresca, M.; Balestrero, F.; Benatti, U.; Armani, U.; Piana, A. Platelet membrane fatty acids in thrombocytosis due to myeloproliferative disorders. Cell Biochem. Funct. 1984, 2, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Tibes, R.; Mesa, R.A. Targeting hedgehog signaling in myelofibrosis and other hematologic malignancies. J. Hematol. Oncol. 2014, 7, 18. [Google Scholar] [CrossRef][Green Version]
- Klein, C.; Zwick, A.; Kissel, S.; Forster, C.U.; Pfeifer, D.; Follo, M.; Illert, A.L.; Decker, S.; Benkler, T.; Pahl, H.; et al. Ptch2 loss drives myeloproliferation and myeloproliferative neoplasm progression. J. Exp. Med. 2016, 213, 273–290. [Google Scholar] [CrossRef]
- Kollmann, K.; Warsch, W.; Gonzalez-Arias, C.; Nice, F.L.; Avezov, E.; Milburn, J.; Li, J.; Dimitropoulou, D.; Biddie, S.; Wang, M.; et al. A novel signaling screen demonstrates that CALR mutations activate essential MAPK signaling and facilitate megakaryocyte differentiation. Leukemia 2017, 31, 934–944. [Google Scholar] [CrossRef]
- Chung, E.; Hsu, C.L.; Kondo, M. Constitutive MAP kinase activation in hematopoietic stem cells induces a myeloproliferative disorder. PLoS ONE 2011, 6, e28350. [Google Scholar] [CrossRef]
- Licht, J.D. SETD2: A complex role in blood malignancy. Blood 2017, 130, 2576–2578. [Google Scholar] [CrossRef]
- Behnert, A.; Lee, A.G.; Young, E.P.; Breese, M.R.; Leung, S.G.; Behroozfard, I.; Maruffi, M.; Sweet-Cordero, E.A.; Dvorak, C.C.; Chu, J.; et al. NUP98-NSD1 driven MDS/MPN in childhood masquerading as JMML. J. Pediatr. Hematol. Oncol. 2021, 43, e808–e811. [Google Scholar] [CrossRef] [PubMed]
- Fatica, A.; Fazi, F. MicroRNA-regulated pathways in hematological malignancies: How to avoid cells playing out of tune. Int. J. Mol. Sci. 2013, 14, 20930–20953. [Google Scholar] [CrossRef] [PubMed]
- Skov, V.; Thomassen, M.; Kjær, L.; Riley, C.; Larsen, T.S.; Bjerrum, O.W.; Kruse, T.A.; Hasselbalch, H.C. Whole blood transcriptional profiling reveals highly deregulated atherosclerosis genes in myeloproliferative cancer. Blood 2018, 132 (Suppl. 1), 3071. [Google Scholar] [CrossRef]
- Lambert, J.; Saliba, J.; Calderon, C.; Sii-Felice, K.; Salma, M.; Edmond, V.; Alvarez, J.C.; Delord, M.; Marty, C.; Plo, I.; et al. PPARγ agonists promote the resolution of myelofibrosis in preclinical models. J. Clin. Investig. 2021, 131, e136713. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Bates, T.M.; Lehmann, G.M.; Simpson-Haidaris, P.J.; Bernstein, S.H.; Sime, P.J.; Phipps, R.P. Role of peroxisome proliferator-activated receptor gamma and its ligands in the treatment of hematological malignancies. PPAR Res. 2008, 2008, 834612. [Google Scholar] [CrossRef]
- Owusu, B.Y.; Zimmerman, K.A.; Murphy-Ullrich, J.E. The role of the endoplasmic reticulum protein calreticulin in mediating TGF-β-stimulated extracellular matrix production in fibrotic disease. J. Cell Commun. Signal. 2018, 12, 289–299. [Google Scholar] [CrossRef]
- Prins, D.; Green, A.R. Mutant CALR functions: Gains and losses. Blood 2020, 136, 6–7. [Google Scholar] [CrossRef]
- Lim, K.-H.; Chang, Y.-C.; Chiang, Y.-H.; Lin, H.C.; Chang, C.-Y.; Lin, C.-S.; Huang, L.; Wang, W.-T.; Chen, C.G.-S.; Chou, W.-C.; et al. Expression of CALR mutants causes mpl-dependent thrombocytosis in zebrafish. Blood Cancer J. 2016, 6, e481. [Google Scholar] [CrossRef]
- Marty, C.; Pecquet, C.; Nivarthi, H.; El-Khoury, M.; Chachoua, I.; Tulliez, M.; Villeval, J.L.; Raslova, H.; Kralovics, R.; Constantinescu, S.; et al. Calreticulin mutants in mice induce an MPL-dependent thrombocytosis with frequent progression to myelofibrosis. Blood 2016, 127, 1317–1324. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guijarro-Hernández, A.; Eder-Azanza, L.; Hurtado, C.; Navarro-Herrera, D.; Ezcurra, B.; Novo, F.J.; Cabello, J.; Vizmanos, J.L. Transcriptomic Analysis Reveals JAK2/MPL-Independent Effects of Calreticulin Mutations in a C. elegans Model. Cells 2023, 12, 186. https://doi.org/10.3390/cells12010186
Guijarro-Hernández A, Eder-Azanza L, Hurtado C, Navarro-Herrera D, Ezcurra B, Novo FJ, Cabello J, Vizmanos JL. Transcriptomic Analysis Reveals JAK2/MPL-Independent Effects of Calreticulin Mutations in a C. elegans Model. Cells. 2023; 12(1):186. https://doi.org/10.3390/cells12010186
Chicago/Turabian StyleGuijarro-Hernández, Ana, Laura Eder-Azanza, Cristina Hurtado, David Navarro-Herrera, Begoña Ezcurra, Francisco Javier Novo, Juan Cabello, and José Luis Vizmanos. 2023. "Transcriptomic Analysis Reveals JAK2/MPL-Independent Effects of Calreticulin Mutations in a C. elegans Model" Cells 12, no. 1: 186. https://doi.org/10.3390/cells12010186
APA StyleGuijarro-Hernández, A., Eder-Azanza, L., Hurtado, C., Navarro-Herrera, D., Ezcurra, B., Novo, F. J., Cabello, J., & Vizmanos, J. L. (2023). Transcriptomic Analysis Reveals JAK2/MPL-Independent Effects of Calreticulin Mutations in a C. elegans Model. Cells, 12(1), 186. https://doi.org/10.3390/cells12010186