
Essential Amino Acid Intake Is Required for Sustaining Serum Insulin-like Growth Factor-I Levels but Is Not Necessarily Needed for Body Growth

 and
and 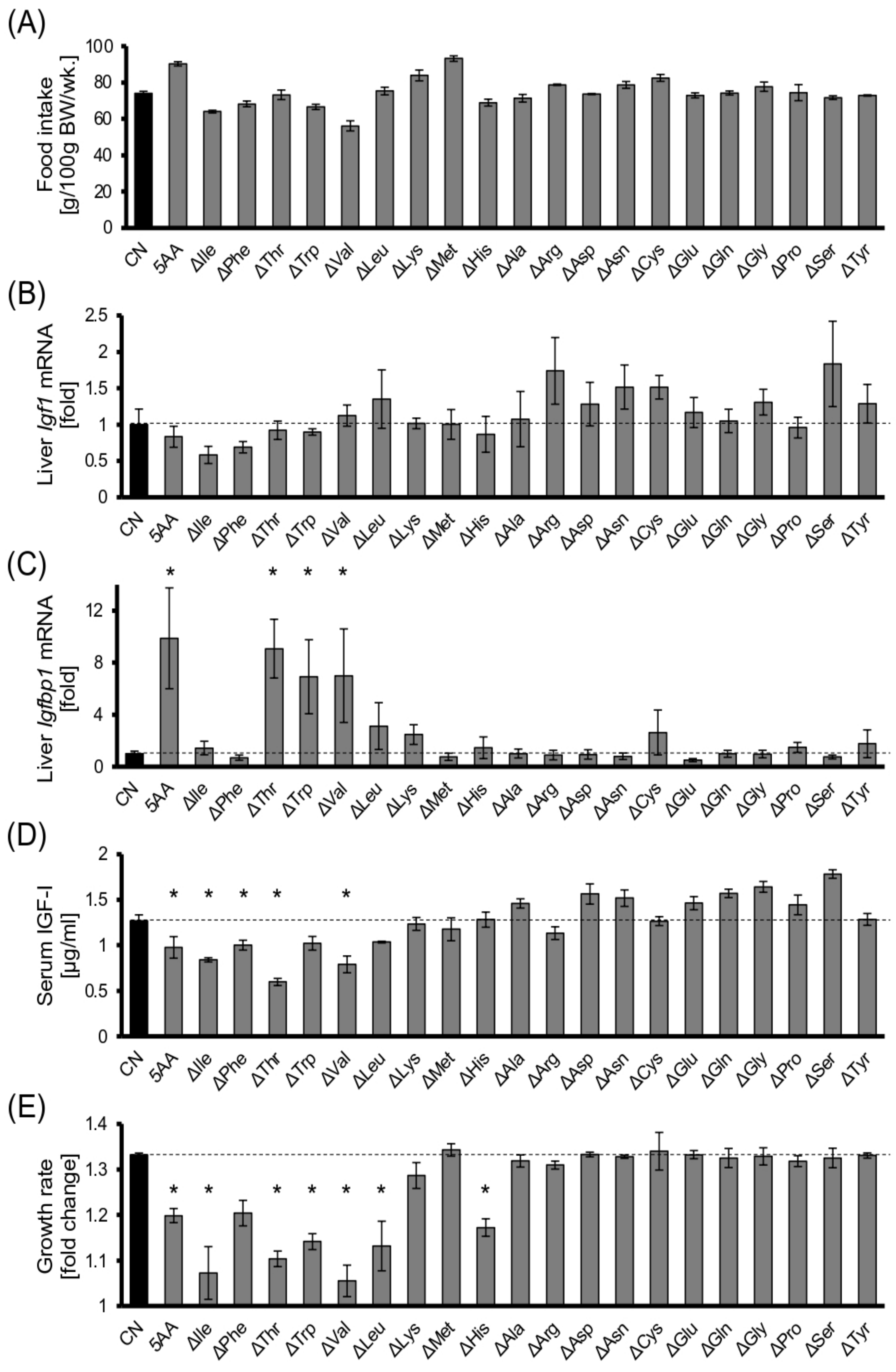{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Blood Parameters
2.4. Cell Culture and Cell Experiments
2.5. RNA Extraction and Real-Time Quantitative PCR (qPCR)
2.6. Immunoblotting
2.7. Statistical Analysis
3. Results
3.1. Dietary EAA Restriction Causes Serum IGF-I Reduction and Growth Retardation in Young Rats
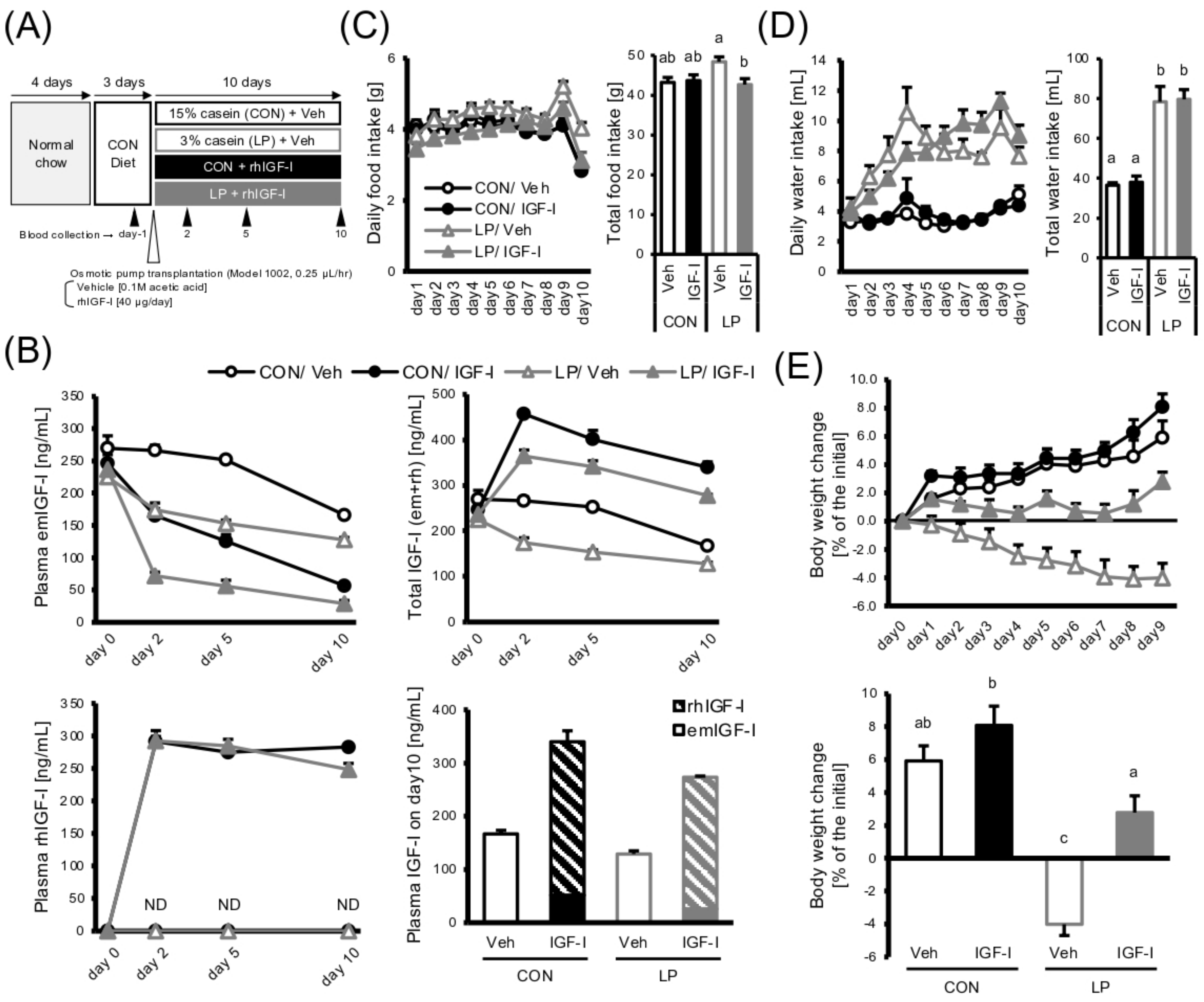
3.2. IGF-I Infusion Restores the Growth Retardation under Protein Malnutrition
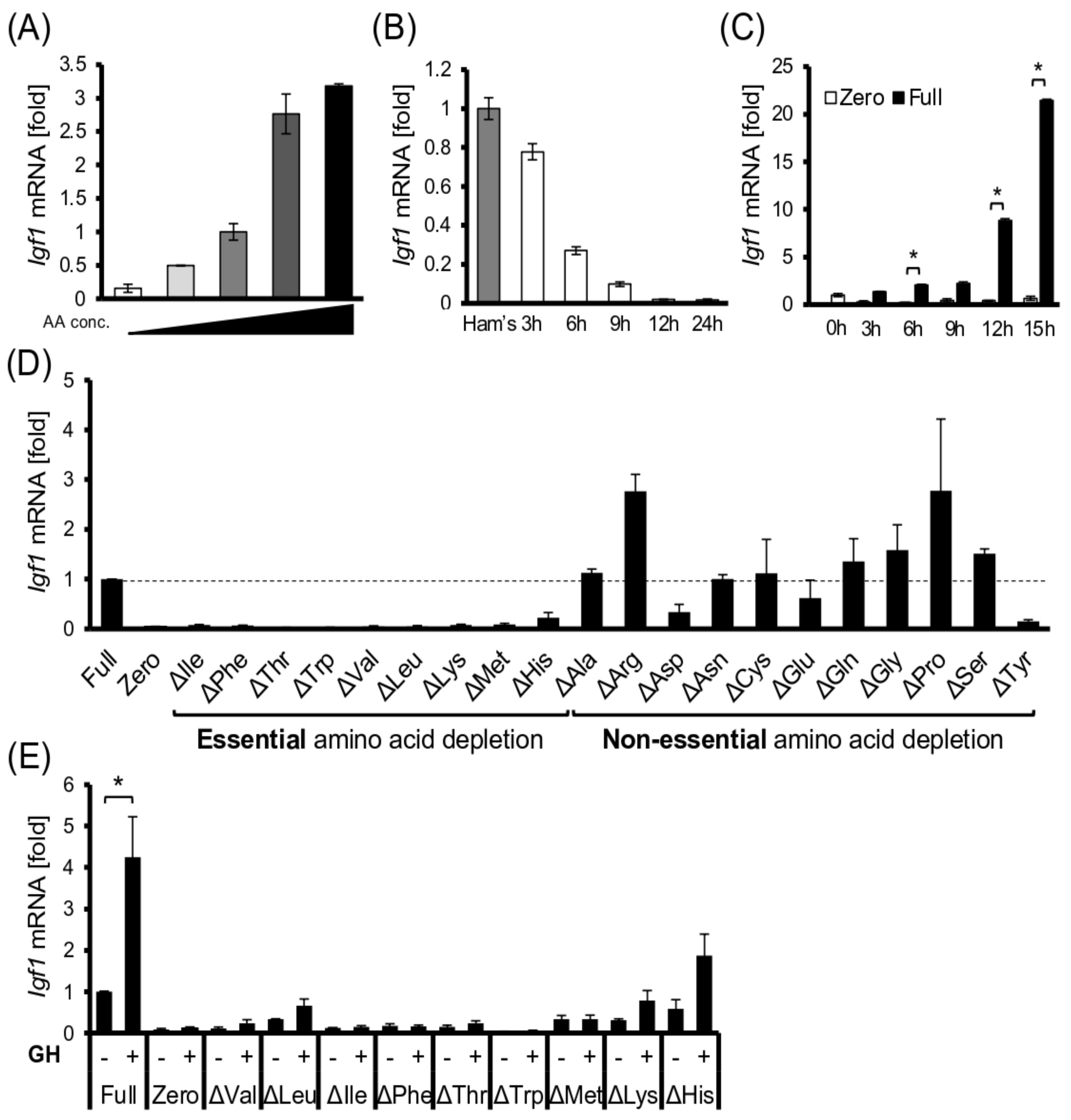
3.3. EAAs Are Required for Igf1 Transcription in Hepatocyte Models
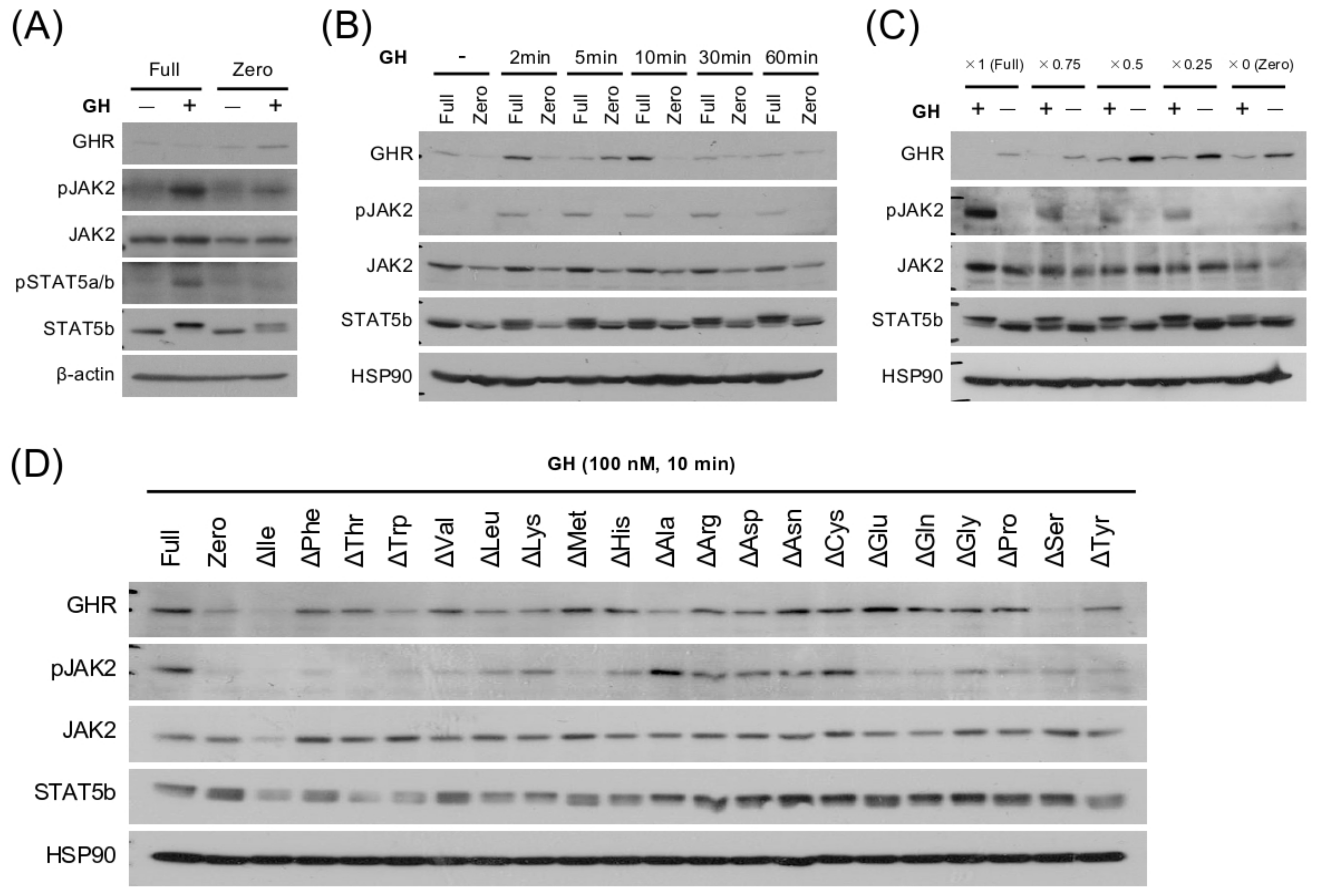
3.4. EAA Deprivation Induces GH Resistance in the Hepatocyte Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simoni, R.D.; Hill, R.L.; Vaughan, M. The discovery of the amino acid threonine: The work of William C. Rose. J. Biol. Chem. 2002, 277, 56–58. [Google Scholar] [CrossRef]
- Pannemans, D.L.; Halliday, D.; Westerterp, K.R.; Kester, A.D. Effect of variable protein intake on whole-body protein turnover in young men and women. Am. J. Clin. Nutr. 1995, 61, 69–74. [Google Scholar] [CrossRef]
- Garlick, P.J.; Millward, D.J.; James, W.P.T.; Waterlow, J.C. The effect of protein deprivation and starvation on the rate of protein synthesis in tissues of the rat. Biochim. Biophys. Acta (BBA)-Nucleic Acids Protein Synth. 1975, 414, 71–84. [Google Scholar] [CrossRef]
- Laurent, B.C.; Moldawer, L.L.; Young, V.R.; Bistrian, B.R.; Blackburn, G.L. Whole-body leucine and muscle protein kinetics in rats fed varying protein intakes. Am. J. Physiol.-Endocrinol. Metab. 1984, 246, E444–E451. [Google Scholar] [CrossRef] [PubMed]
- Forbes, R.M.; Cooper, A.R.; Mitchell, H.H. The composition of the adult human body as determined by chemical analysis. J. Biol. Chem. 1953, 203, 359–366. Available online: http://www.jbc.org/content/203/1/359.short (accessed on 19 June 2016). [CrossRef]
- Yakar, S.; Liu, J.-L.; Le Roith, D. The growth hormone/insulin-like growth factor-I system: Implications for organ growth and development. Pediatr. Nephrol. 2000, 14, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Herrington, J.; Smit, L.S.; Schwartz, J.; Carter-Su, C. The role of STAT proteins in growth hormone signaling. Oncogene 2000, 19, 2585–2597. [Google Scholar] [CrossRef]
- Lupu, F.; Terwilliger, J.D.; Lee, K.; Segre, G.V.; Efstratiadis, A. Roles of Growth Hormone and Insulin-like Growth Factor 1 in Mouse Postnatal Growth. Dev. Biol. 2001, 229, 141–162. [Google Scholar] [CrossRef]
- Zhang, M.; Xuan, S.; Bouxsein, M.L.; von Stechow, D.; Akeno, N.; Faugere, M.C.; Malluche, H.; Zhao, G.; Rosen, C.J.; Efstratiadis, A.; et al. Osteoblast-specific Knockout of the Insulin-like Growth Factor (IGF) Receptor Gene Reveals an Essential Role of IGF Signaling in Bone Matrix Mineralization. J. Biol. Chem. 2002, 277, 44005–44012. [Google Scholar] [CrossRef]
- Takahashi, S.-I.; Takenaka, A.; Noguchi, T. Relationship between Nutrient Intake and Insulin-like Growth Factor I (IGF-I). Clin. Pediatr. Endocrinol. 1994, 3, 111–114. [Google Scholar] [CrossRef]
- Harel, Z.; Tannenbaum, G.S. Dietary protein restriction impairs both spontaneous and growth hormone-releasing factor-stimulated growth hormone release in the rat. Endocrinology 1993, 133, 1035–1043. [Google Scholar] [CrossRef]
- Thissen, J.P.; Triest, S.; Underwood, L.E.; Maes, M.; Ketelslegers, J.M. Divergent Responses of Serum Insulin-Like Growth Factor-I and Liver Growth Hormone (GH) Receptors to Exogenous GH in Protein-Restricted Rats. Endocrinology 1990, 126, 908–913. [Google Scholar] [CrossRef]
- Nagasawa, T.; Sakai, T.; Onodera, R. Simple and sensitive determination of plasma Nτ-methylhistidine by high-performance liquid chromatography using pre-column derivative formation with o-phthalaldehyde—2-mercaptoethanol. J. Chromatogr. B Biomed. Sci. Appl. 1991, 566, 223–227. [Google Scholar] [CrossRef]
- Nishi, H.; Yamanaka, D.; Kamei, H.; Goda, Y.; Kumano, M.; Toyoshima, Y.; Takenaka, A.; Masuda, M.; Nakabayashi, Y.; Shioya, R.; et al. Importance of Serum Amino Acid Profile for Induction of Hepatic Steatosis under Protein Malnutrition. Sci. Rep. 2018, 8, 5461. [Google Scholar] [CrossRef]
- Jones, J.I.; Clemmons, D.R. Insulin-Like Growth Factors and Their Binding Proteins: Biological Actions. Endocr. Rev. 1995, 16, 3–34. [Google Scholar] [CrossRef]
- Laeger, T.; Henagan, T.M.; Albarado, D.C.; Redman, L.M.; Bray, G.A.; Noland, R.C.; Münzberg, H.; Hutson, S.M.; Gettys, T.W.; Schwartz, M.W.; et al. FGF21 is an endocrine signal of protein restriction. J. Clin. Investig. 2014, 124, 3913–3922. [Google Scholar] [CrossRef]
- Takenaka, A.; Komori, K.; Morishita, T.; Takahashi, S.; Hidaka, T.; Noguchi, T. Amino acid regulation of gene transcription of rat insulin-like growth factor-binding protein-1. J. Endocrinol. 2000, 164, R11–R16. [Google Scholar] [CrossRef][Green Version]
- Fukushima, S.; Nishi, H.; Kumano, M.; Yamanaka, D.; Kataoka, N.; Hakuno, F.; Takahashi, S.-I. A novel amino acid signaling process governs glucose-6-phosphatase transcription. iScience 2021, 24, 102778. [Google Scholar] [CrossRef]
- The National Academy of Sciences; The National Research Council. Nutrient Requirements of Laboratory Animals, 4th ed.; National Academies Press: Washington, DC, USA, 1995. [Google Scholar]
- Amaya-Farfán, J.; Beaufrère, B.; Butte, N.F.; Cabrera, M.I.Z.; Garlick, P.; Gilani, G.S.; Jackson, A.; Kishi, K.; Kurpad, A.V.; Millward, J.; et al. Protein and Amino Acid Requirements in Human Nutrition: Report of a Joint FAO/WHO/UNU Expert Consultation; WHO Technical Report Series; WHO: Geneva, Switzerland, 2007; p. 935. [Google Scholar]
- Toyoshima, Y.; Tokita, R.; Taguchi, Y.; Akiyama-Akanishi, N.; Takenaka, A.; Kato, H.; Chida, K.; Hakuno, F.; Minami, S.; Takahashi, S.-I. Tissue-specific effects of protein malnutrition on insulin signaling pathway and lipid accumulation in growing rats. Endocr. J. 2014, 61, 499–512. [Google Scholar] [CrossRef]
- Mizushima, N.; Yamamoto, A.; Matsui, M.; Yoshimori, T.; Ohsumi, Y. In Vivo Analysis of Autophagy in Response to Nutrient Starvation Using Transgenic Mice Expressing a Fluorescent Autophagosome Marker. Mol. Biol. Cell 2004, 15, 1101–1111. [Google Scholar] [CrossRef]
- Metges, C.C. Contribution of Microbial Amino Acids to Amino Acid Homeostasis of the Host. J. Nutr. 2000, 130, 1857S–1864S. [Google Scholar] [CrossRef] [PubMed]
- Philipps, A.F.; Persson, B.; Hall, K.; Lake, M.; Skottner, A.; Sanengen, T.; Sara, V.R. The Effects of Biosynthetic Insulin-Like Growth Factor-1 Supplementation on Somatic Growth, Maturation, and Erythropoiesis on the Neonatal Rat. Pediatr. Res. 1988, 23, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Thissen, J.-P.; Underwood, L.E.; Maiter, D.; Maes, M.; Clemmons, D.R.; Ketelslegers, J.-M. Failure of Insulin-Like Growth Factor-I (IGF-I) Infusion to Promote Growth in Protein-Restricted Rats Despite Normalization of Serum IGF-I Concentrations. Endocrinology 1991, 128, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Golde, D.W.; Bailey, R.; Geffner, M.E. Insulin-Like Growth Factor-I Resistance. Endocr. Rev. 1998, 19, 625–646. [Google Scholar] [CrossRef]
- Yakar, S.; Liu, J.-L.; Stannard, B.; Butler, A.; Accili, D.; Sauer, B.; LeRoith, D. Normal growth and development in the absence of hepatic insulin-like growth factor I. Proc. Natl. Acad. Sci. USA 1999, 96, 7324–7329. [Google Scholar] [CrossRef]
- Toyoshima, Y.; Tokita, R.; Ohne, Y.; Hakuno, F.; Noguchi, T.; Minami, S.; Kato, H.; Takahashi, S.-I. Dietary protein deprivation upregulates insulin signaling and inhibits gluconeogenesis in rat liver. J. Mol. Endocrinol. 2010, 45, 329–340. [Google Scholar] [CrossRef]
- Fontana, L.; Partridge, L.; Longo, V.D. Extending Healthy Life Span—From Yeast to Humans. Science 2010, 328, 321–326. [Google Scholar] [CrossRef]
- Hadden, D.R.; Rutishauser, I.H. Effect of human growth hormone in kwashiorkor and marasmus. Arch. Dis. Child. 1967, 42, 29–33. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishi, H.; Uchida, K.; Saito, M.; Yamanaka, D.; Nagata, H.; Tomoshige, H.; Miyata, I.; Ito, K.; Toyoshima, Y.; Takahashi, S.-I.; et al. Essential Amino Acid Intake Is Required for Sustaining Serum Insulin-like Growth Factor-I Levels but Is Not Necessarily Needed for Body Growth. Cells 2022, 11, 1523. https://doi.org/10.3390/cells11091523
Nishi H, Uchida K, Saito M, Yamanaka D, Nagata H, Tomoshige H, Miyata I, Ito K, Toyoshima Y, Takahashi S-I, et al. Essential Amino Acid Intake Is Required for Sustaining Serum Insulin-like Growth Factor-I Levels but Is Not Necessarily Needed for Body Growth. Cells. 2022; 11(9):1523. https://doi.org/10.3390/cells11091523
Chicago/Turabian StyleNishi, Hiroki, Kaito Uchida, Maki Saito, Daisuke Yamanaka, Haruka Nagata, Hinako Tomoshige, Ichiro Miyata, Koichi Ito, Yuka Toyoshima, Shin-Ichiro Takahashi, and et al. 2022. "Essential Amino Acid Intake Is Required for Sustaining Serum Insulin-like Growth Factor-I Levels but Is Not Necessarily Needed for Body Growth" Cells 11, no. 9: 1523. https://doi.org/10.3390/cells11091523
APA StyleNishi, H., Uchida, K., Saito, M., Yamanaka, D., Nagata, H., Tomoshige, H., Miyata, I., Ito, K., Toyoshima, Y., Takahashi, S.-I., Hakuno, F., & Takenaka, A. (2022). Essential Amino Acid Intake Is Required for Sustaining Serum Insulin-like Growth Factor-I Levels but Is Not Necessarily Needed for Body Growth. Cells, 11(9), 1523. https://doi.org/10.3390/cells11091523