
Human Wharton’s Jelly Mesenchymal Stem Cells Secretome Inhibits Human SARS-CoV-2 and Avian Infectious Bronchitis Coronaviruses

 , , , ,
, , , ,  , , ,
, , , 

Abstract
1. Introduction
2. Materials and Method
2.1. Cells and Viruses
2.2. Collection and Processing of Human Umbilical Cords (hUCs)
2.3. Culturing of hUC-Derived Mesenchymal Stem Cells (hUC-MSCs)
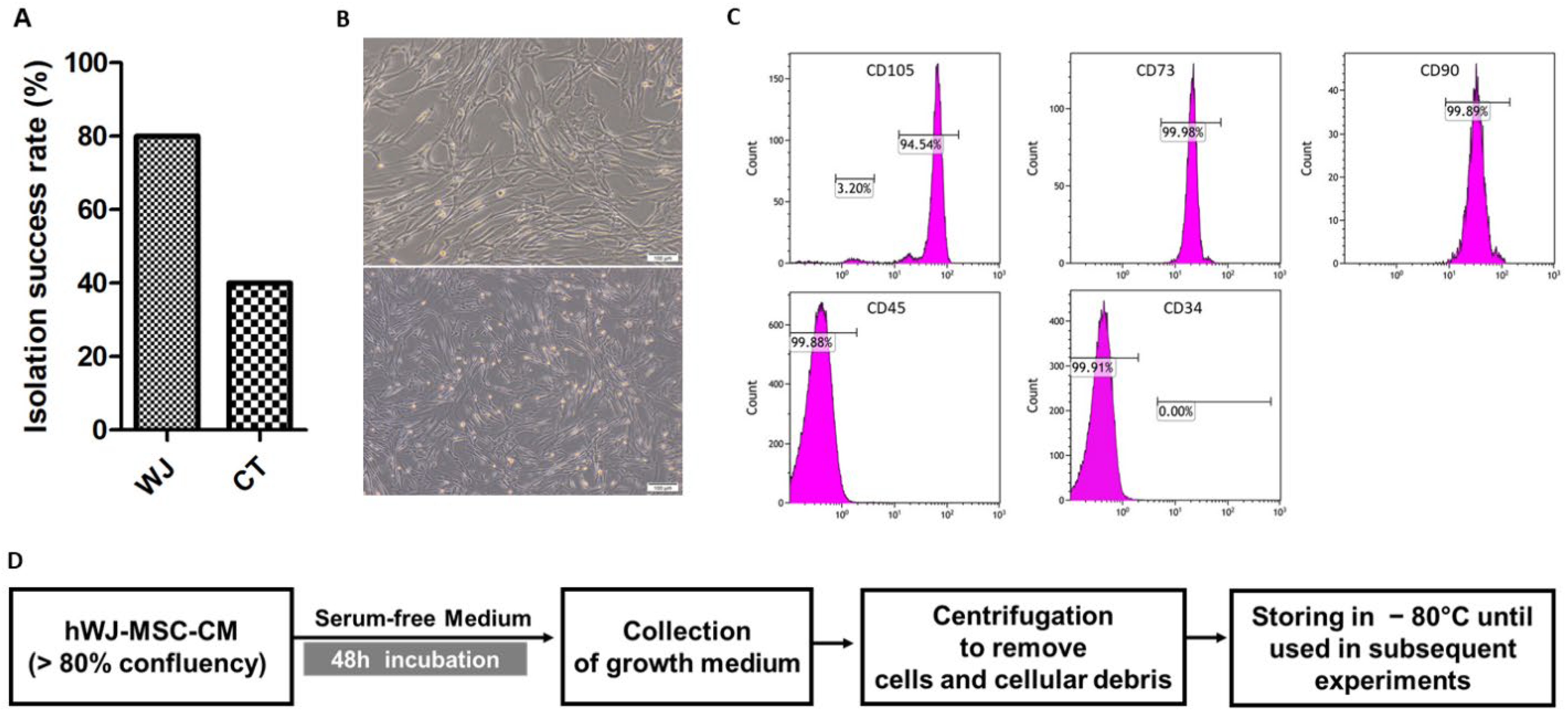
2.4. Flow Cytometry Characterization of hUC-MSCs
2.5. Collection of hWJ-MSC-S
2.6. Cytotoxicity Assay
2.7. Embryotoxicity
2.8. Determination of Viral Inhibitory Concentration 50 (IC50)
2.9. Plaque-Reduction Assay
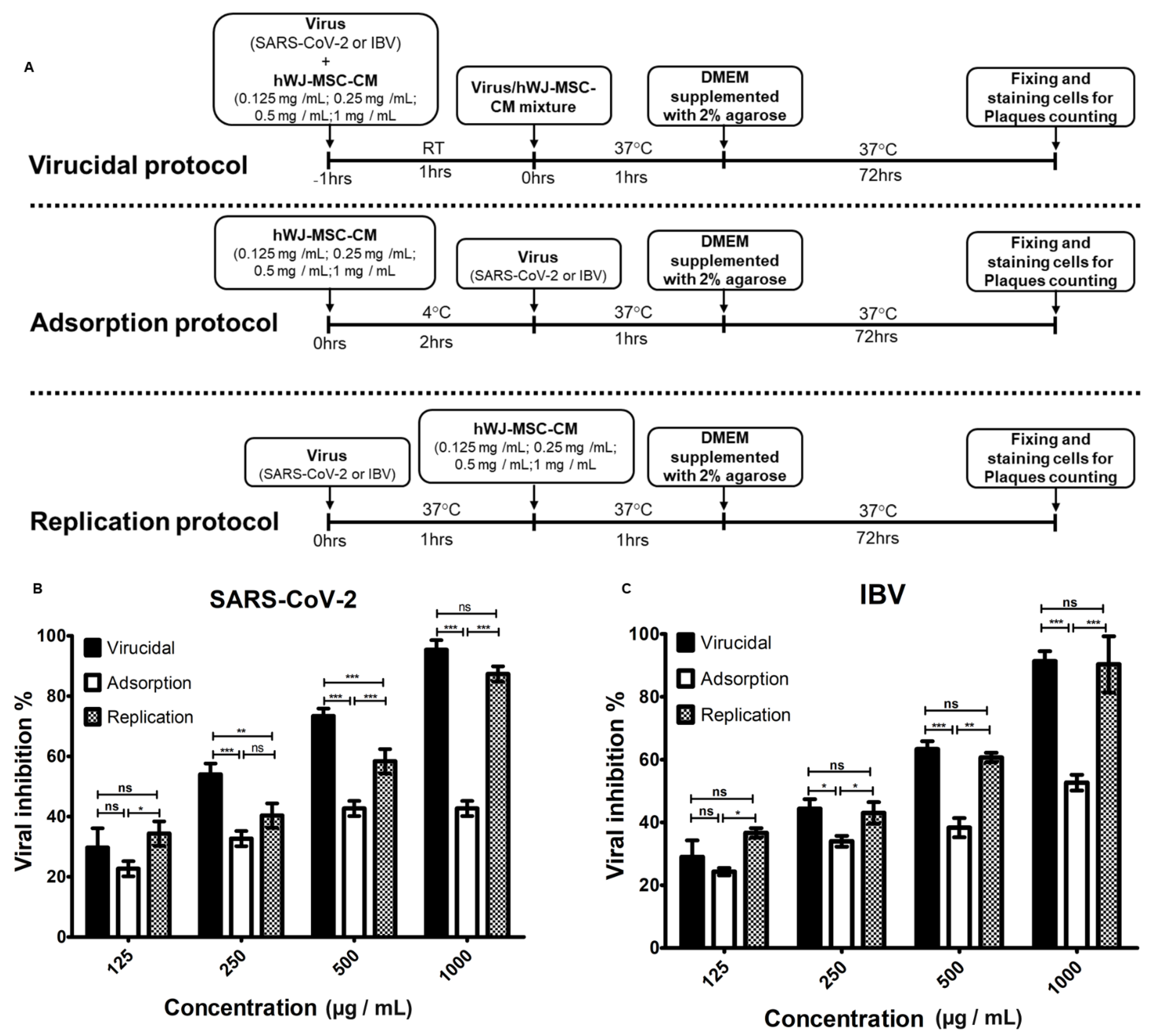
2.10. Determination of the Mode of Action
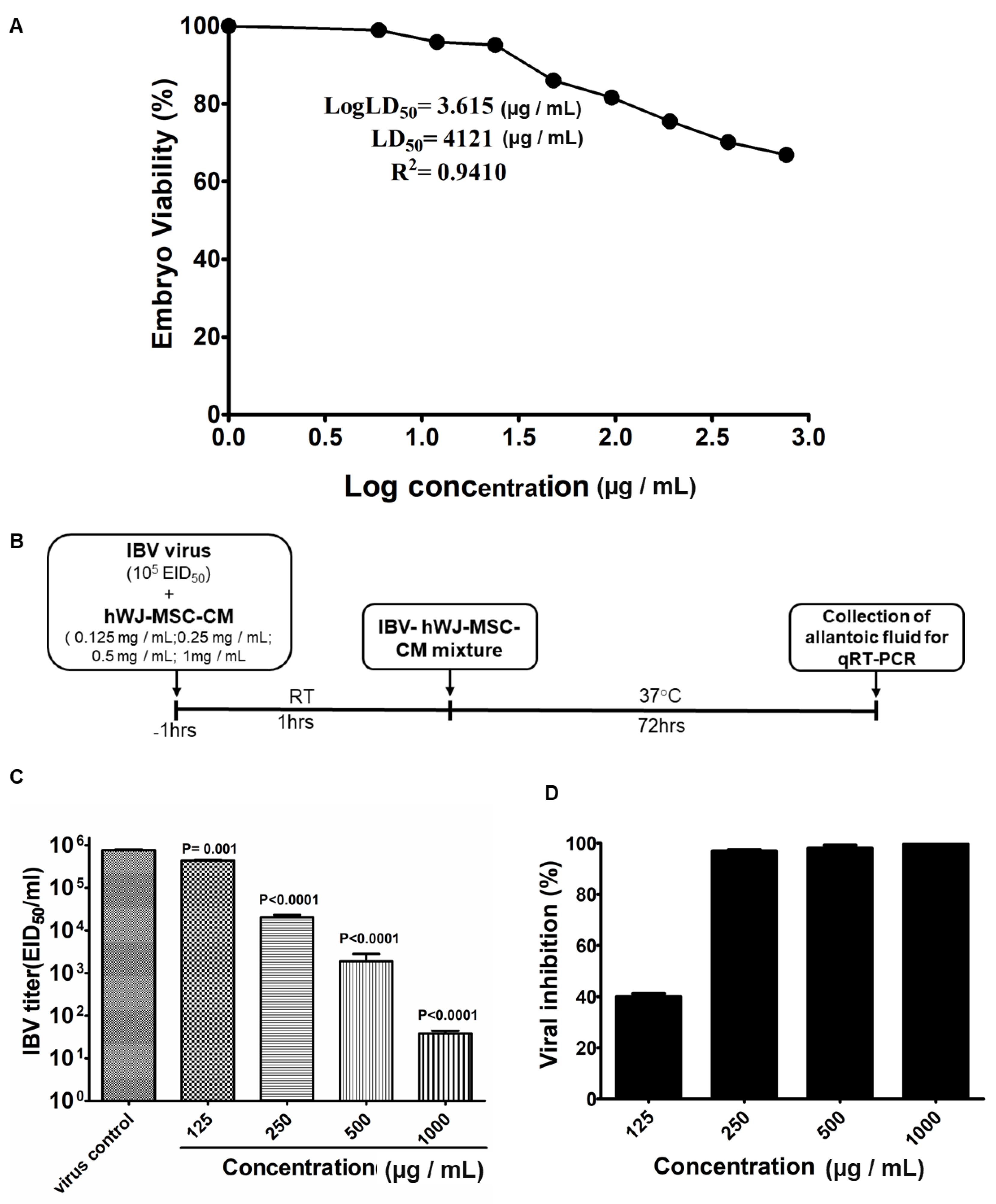
2.11. In Ovo Anti-IBV Activity of hWJ-MSC-S
2.12. RNA Extraction and RT-qPCR to Detect IBV
2.13. Mass Spectrometry Analysis of hWJ-MSC-CM
2.14. Bioinformatics Analysis
2.15. Gene Ontology and Pathway Enrichment Analysis
2.16. Statistical Analysis
3. Results
3.1. Isolation and Characterization of hUCMSC
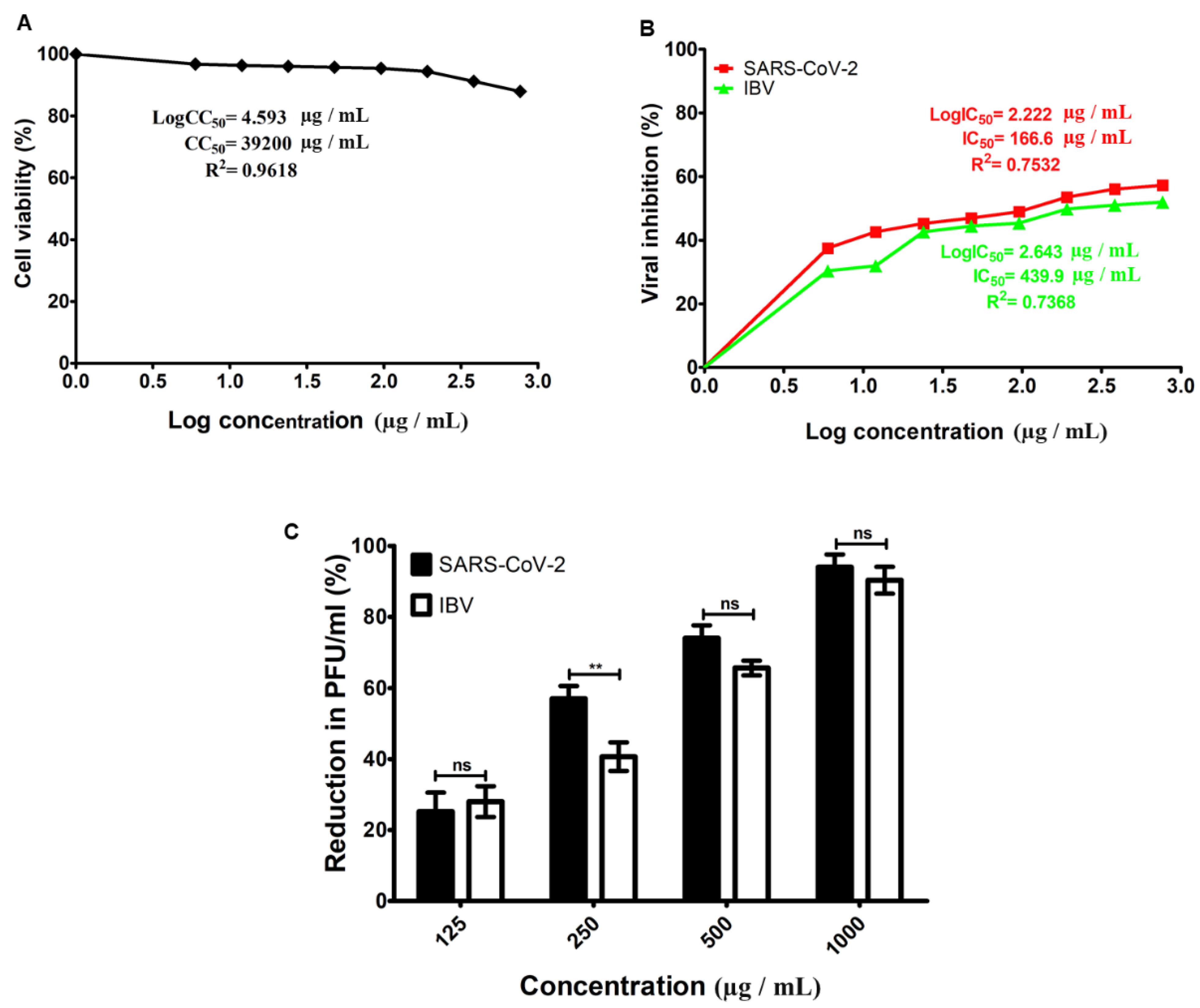
3.2. Cytotoxicity and Antiviral Activity of hWJ-MSC-S
3.3. Determination of the Antiviral Mechanism of hWJ-MSC-S
3.4. Determination of in Ovo Anti-IBV Activity of hWJ-MSC-S
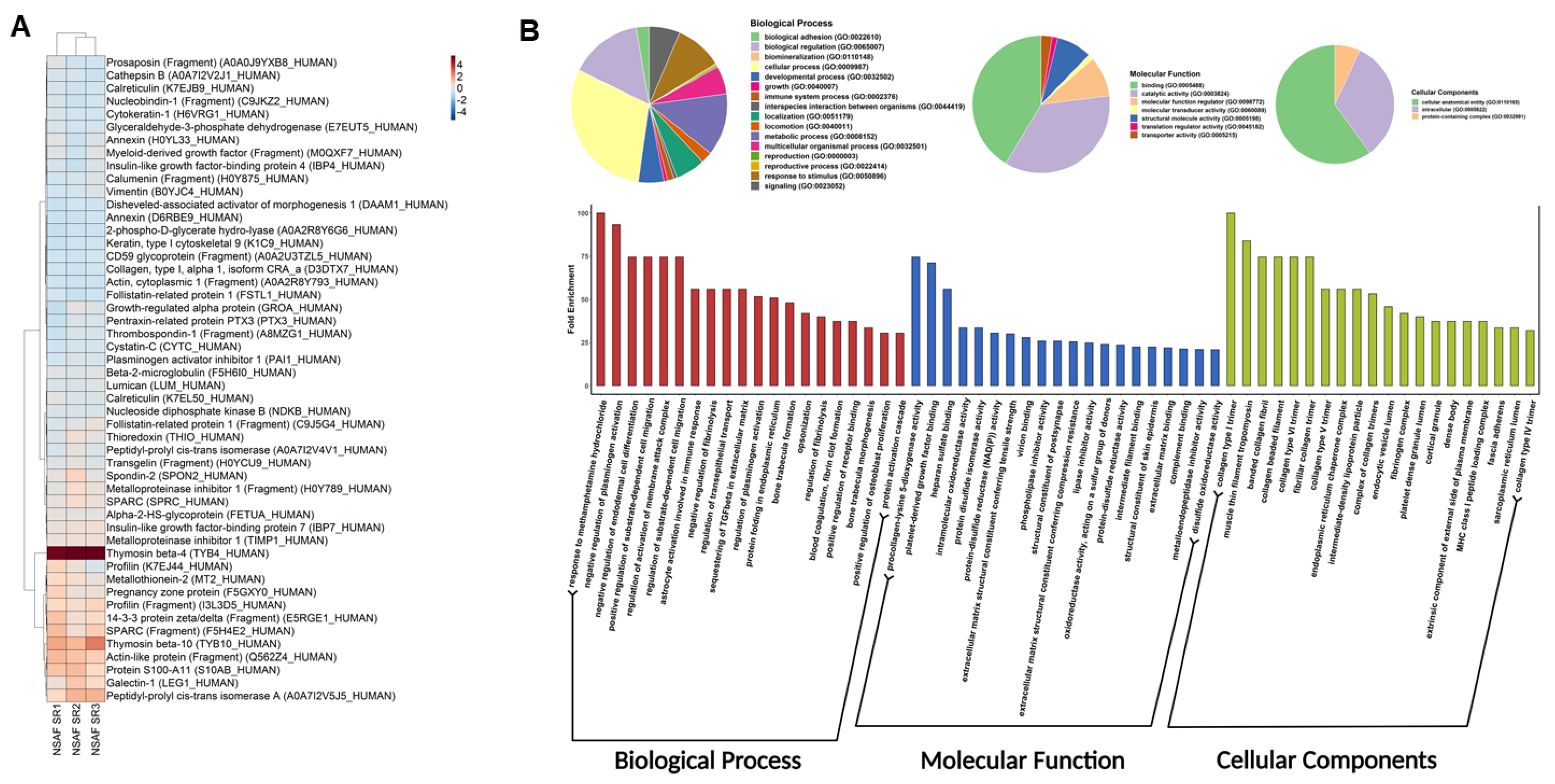
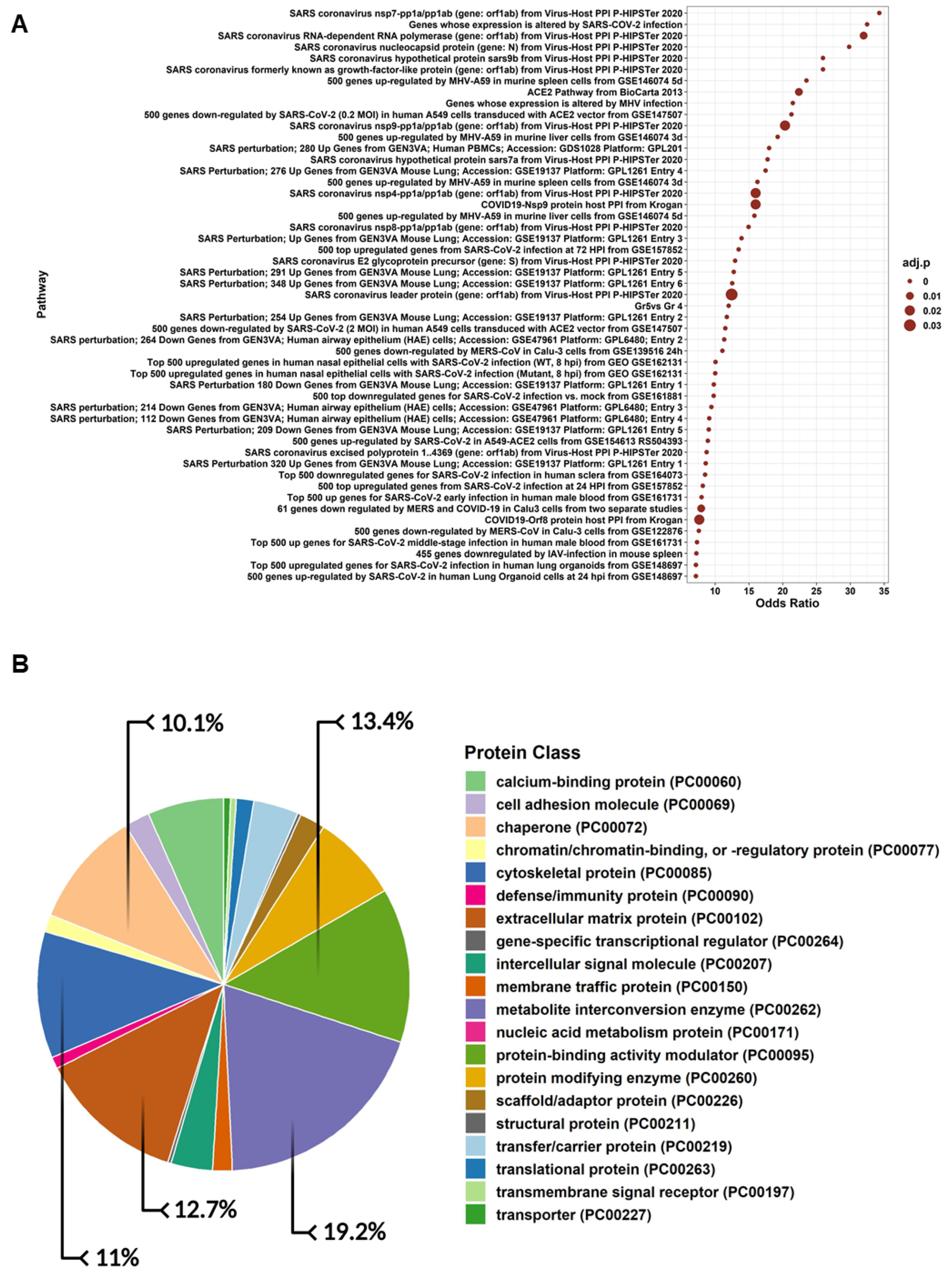
3.5. Proteomics Profiling and Gene Ontology of hWJ-MSC-S
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Bande, F.; Arshad, S.S.; Omar, A.R.; Bejo, M.H.; Abubakar, M.S.; Abba, Y. Pathogenesis and Diagnostic Approaches of Avian Infectious Bronchitis. Adv. Virol. 2016, 2016, 4621659. [Google Scholar] [CrossRef] [PubMed]
- Marty, A.M.; Jones, M.K. The novel Coronavirus (SARS-CoV-2) is a one health issue. One Health 2020, 9, 100123. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.K.; Jackwood, M.; Jones, R.C. The long view: 40 years of infectious bronchitis research. Avian Pathol. 2012, 41, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wang, Z.; Shao, C.; Yu, J.; Liu, H.; Chen, H.; Li, L.; Wang, X.; Ren, Y.; Huang, X.; et al. Analysis of chicken macrophage functions and gene expressions following infectious bronchitis virus M41 infection. Vet. Res. 2021, 52, 14. [Google Scholar] [CrossRef] [PubMed]
- Alhatami, A.O.; Alaraji, F.; Abdulwahab, H.M.; Khudhair, Y.I. Sequencing and phylogenetic analysis of infectious bronchitis virus variant strain from an outbreak in egg-layer flocks in Baghdad, Iraq. Vet. World 2020, 13, 1358–1362. [Google Scholar] [CrossRef] [PubMed]
- Meir, R.; Maharat, O.; Farnushi, Y.; Simanov, L. Development of a real-time TaqMan RT-PCR assay for the detection of infectious bronchitis virus in chickens, and comparison of RT-PCR and virus isolation. J. Virol. Methods 2010, 163, 190–194. [Google Scholar] [CrossRef]
- Kuehnle, I.; Goodell, M.A. The therapeutic potential of stem cells from adults. BMJ 2002, 325, 372–376. [Google Scholar] [CrossRef]
- Zakrzewski, W.; Dobrzyński, M.; Szymonowicz, M.; Rybak, Z. Stem cells: Past, present, and future. Stem. Cell Res. Ther. 2019, 10, 68. [Google Scholar] [CrossRef]
- Hattori, Y.; Kim, H.; Tsuboi, N.; Yamamoto, A.; Akiyama, S.; Shi, Y.; Katsuno, T.; Kosugi, T.; Ueda, M.; Matsuo, S.; et al. Therapeutic Potential of Stem Cells from Human Exfoliated Deciduous Teeth in Models of Acute Kidney Injury. PLoS ONE 2015, 10, e0140121. [Google Scholar]
- Ueda, T.; Inden, M.; Ito, T.; Kurita, H.; Hozumi, I. Characteristics and Therapeutic Potential of Dental Pulp Stem Cells on Neurodegenerative Diseases. Front. Neurosci. 2020, 14, 407. [Google Scholar] [CrossRef] [PubMed]
- Moreira, A.; Kahlenberg, S.; Hornsby, P. Therapeutic potential of mesenchymal stem cells for diabetes. J. Mol. Endocrinol. 2017, 59, R109-r. [Google Scholar] [CrossRef]
- Hatzistergos, K.E.; Blum, A.; Ince, T.; Grichnik, J.M.; Hare, J.M. What is the oncologic risk of stem cell treatment for heart disease? Circ. Res. 2011, 108, 1300–1303. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Lin, T.; Xu, Y. Stem Cell Therapy and Immunological Rejection in Animal Models. Curr. Mol. Pharmacol. 2016, 9, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Tullis, G.E.; Spears, K.; Kirk, M.D. Immunological barriers to stem cell therapy in the central nervous system. Stem Cells Int. 2014, 2014, 507905. [Google Scholar] [CrossRef]
- Young, J.-A.H.; Weisdorf, D.J. Infections in Recipients of Hematopoietic Stem Cell Transplants. Mandell Douglas Bennett’s Princ. Pract. Infect. Dis. 2015, 2015, 3425–3439.e3425. [Google Scholar] [CrossRef]
- Daneshmandi, L.; Shah, S.; Jafari, T.; Bhattacharjee, M.; Momah, D.; Saveh-Shemshaki, N.; Lo, K.W.; Laurencin, C.T. Emergence of the Stem Cell Secretome in Regenerative Engineering. Trends Biotechnol. 2020, 38, 1373–1384. [Google Scholar] [CrossRef]
- Lin, L.; Du, L. The role of secreted factors in stem cells-mediated immune regulation. Cell. Immunol. 2018, 326, 24–32. [Google Scholar] [CrossRef]
- Meldolesi, J. Exosomes and Ectosomes in Intercellular Communication. Curr. Biol. CB 2018, 28, R435–R444. [Google Scholar] [CrossRef]
- Budoni, M.; Fierabracci, A.; Luciano, R.; Petrini, S.; Di Ciommo, V.; Muraca, M. The immunosuppressive effect of mesenchymal stromal cells on B lymphocytes is mediated by membrane vesicles. Cell Transpl. 2013, 22, 369–379. [Google Scholar] [CrossRef]
- Fong, C.Y.; Tam, K.; Cheyyatraivendran, S.; Gan, S.U.; Gauthaman, K.; Armugam, A.; Jeyaseelan, K.; Choolani, M.; Biswas, A.; Bongso, A. Human Wharton’s jelly stem cells and its conditioned medium enhance healing of excisional and diabetic wounds. J. Cell. Biochem. 2014, 115, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Nadesh, R.; Menon, K.N.; Biswas, L.; Mony, U.; Subramania Iyer, K.; Vijayaraghavan, S.; Nambiar, A.; Nair, S. Adipose derived mesenchymal stem cell secretome formulation as a biotherapeutic to inhibit growth of drug resistant triple negative breast cancer. Sci. Rep. 2021, 11, 23435. [Google Scholar] [CrossRef] [PubMed]
- Foo, J.B.; Looi, Q.H.; Chong, P.P.; Hassan, N.H.; Yeo, G.E.C.; Ng, C.Y.; Koh, B.; How, C.W.; Lee, S.H.; Law, J.X. Comparing the Therapeutic Potential of Stem Cells and their Secretory Products in Regenerative Medicine. Stem Cells Int. 2021, 2021, 2616807. [Google Scholar] [CrossRef] [PubMed]
- Hofer, H.R.; Tuan, R.S. Secreted trophic factors of mesenchymal stem cells support neurovascular and musculoskeletal therapies. Stem Cell Res. Ther. 2016, 7, 131. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Su, H.; Li, J.; Lyon, C.; Tang, W.; Wan, M.; Hu, T.Y. Clinical applications of exosome membrane proteins. Precis. Clin. Med. 2020, 3, 54–66. [Google Scholar] [CrossRef] [PubMed]
- González-González, A.; García-Sánchez, D.; Dotta, M.; Rodríguez-Rey, J.C.; Pérez-Campo, F.M. Mesenchymal stem cells secretome: The cornerstone of cell-free regenerative medicine. World J. Stem Cells 2020, 12, 1529–1552. [Google Scholar] [CrossRef]
- Kamal, M.M.; Kassem, D.H. Therapeutic Potential of Wharton’s Jelly Mesenchymal Stem Cells for Diabetes: Achievements and Challenges. Front. Cell Dev. Biol. 2020, 8, 16. [Google Scholar] [CrossRef]
- La Rocca, G.; Anzalone, R.; Corrao, S.; Magno, F.; Loria, T.; Lo Iacono, M.; Di Stefano, A.; Giannuzzi, P.; Marasà, L.; Cappello, F.; et al. Isolation and characterization of Oct-4+/HLA-G+ mesenchymal stem cells from human umbilical cord matrix: Differentiation potential and detection of new markers. Histochem. Cell Biol. 2009, 131, 267–282. [Google Scholar] [CrossRef]
- Gauthaman, K.; Fong, C.Y.; Suganya, C.A.; Subramanian, A.; Biswas, A.; Choolani, M.; Bongso, A. Extra-embryonic human Wharton’s jelly stem cells do not induce tumorigenesis, unlike human embryonic stem cells. Reprod. Biomed. Online 2012, 24, 235–246. [Google Scholar] [CrossRef]
- Barrett, A.N.; Fong, C.Y.; Subramanian, A.; Liu, W.; Feng, Y.; Choolani, M.; Biswas, A.; Rajapakse, J.C.; Bongso, A. Human Wharton’s Jelly Mesenchymal Stem Cells Show Unique Gene Expression Compared with Bone Marrow Mesenchymal Stem Cells Using Single-Cell RNA-Sequencing. Stem Cells Dev. 2019, 28, 196–211. [Google Scholar] [CrossRef]
- Fong, C.Y.; Chak, L.L.; Biswas, A.; Tan, J.H.; Gauthaman, K.; Chan, W.K.; Bongso, A. Human Wharton’s jelly stem cells have unique transcriptome profiles compared to human embryonic stem cells and other mesenchymal stem cells. Stem Cell Rev. Rep. 2011, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kandeil, A.; Mostafa, A.; El-Shesheny, R.; Shehata, M.; Roshdy, W.H.; Ahmed, S.S.; Gomaa, M.; Taweel, A.E.; Kayed, A.E.; Mahmoud, S.H.; et al. Coding-Complete Genome Sequences of Two SARS-CoV-2 Isolates from Egypt. Microbiol. Resour. Announc. 2020, 9, e00489-20. [Google Scholar] [CrossRef] [PubMed]
- Yehia, N.; Said, D.; Zanaty, A.M. Characterization and Analysis of the Major Structural Protein Genes of the Recently Isolated Avian Infectious Bronchitis Virus in Egypt. J. World Poult. Res. 2020, 10, 649–661. [Google Scholar] [CrossRef]
- Zhao, H.; To, K.K.W.; Chu, H.; Ding, Q.; Zhao, X.; Li, C.; Shuai, H.; Yuan, S.; Zhou, J.; Kok, K.H.; et al. Dual-functional peptide with defective interfering genes effectively protects mice against avian and seasonal influenza. Nat. Commun. 2018, 9, 2358. [Google Scholar] [CrossRef] [PubMed]
- Beeravolu, N.; McKee, C.; Alamri, A.; Mikhael, S.; Brown, C.; Perez-Cruet, M.; Chaudhry, G.R. Isolation and Characterization of Mesenchymal Stromal Cells from Human Umbilical Cord and Fetal Placenta. J. Vis. Exp. 2017, 122, e55224. [Google Scholar] [CrossRef]
- Elebeedy, D.; Elkhatib, W.F.; Kandeil, A.; Ghanem, A.; Kutkat, O.; Alnajjar, R.; Saleh, M.A.; Abd El Maksoud, A.I.; Badawy, I.; Al-Karmalawy, A.A. Anti-SARS-CoV-2 activities of tanshinone IIA, carnosic acid, rosmarinic acid, salvianolic acid, baicalein, and glycyrrhetinic acid between computational and in vitro insights. RSC Adv. 2021, 11, 29267–29286. [Google Scholar] [CrossRef]
- Nwodo, U.U.; Ngene, A.A.; Iroegbu, C.U.; Onyedikachi, O.A.; Chigor, V.N.; Okoh, A.I. In vivo evaluation of the antiviral activity of Cajanus cajan on measles virus. Arch. Virol. 2011, 156, 1551–1557. [Google Scholar] [CrossRef]
- Jackwood, M.W.; Rosenbloom, R.; Petteruti, M.; Hilt, D.A.; McCall, A.W.; Williams, S.M. Avian coronavirus infectious bronchitis virus susceptibility to botanical oleoresins and essential oils in vitro and in vivo. Virus Res. 2010, 149, 86–94. [Google Scholar] [CrossRef]
- Feoktistova, M.; Geserick, P.; Leverkus, M. Crystal Violet Assay for Determining Viability of Cultured Cells. Cold Spring Harb. Protoc. 2016, 2016, pdb.prot087379. [Google Scholar] [CrossRef]
- Alnajjar, R.; Mostafa, A.; Kandeil, A.; Al-Karmalawy, A.A. Molecular docking, molecular dynamics, and in vitro studies reveal the potential of angiotensin II receptor blockers to inhibit the COVID-19 main protease. Heliyon 2020, 6, e05641. [Google Scholar] [CrossRef]
- Mostafa, A.; Kandeil, A.; Elshaier, Y.A.M.M.; Kutkat, O.; Moatasim, Y.; Rashad, A.A.; Shehata, M.; Gomaa, M.R.; Mahrous, N.; Mahmoud, S.H.; et al. FDA-Approved Drugs with Potent In Vitro Antiviral Activity against Severe Acute Respiratory Syndrome Coronavirus 2. Pharmaceuticals 2020, 13, 443. [Google Scholar] [CrossRef] [PubMed]
- Roshdy, W.H.; Rashed, H.A.; Kandeil, A.; Mostafa, A.; Moatasim, Y.; Kutkat, O.; Abo Shama, N.M.; Gomaa, M.R.; El-Sayed, I.H.; El Guindy, N.M.; et al. EGYVIR: An immunomodulatory herbal extract with potent antiviral activity against SARS-CoV-2. PLoS ONE 2020, 15, e0241739. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Paintsil, E.; Dutschman, G.E.; Grill, S.P.; Wang, C.J.; Wang, J.; Tanaka, H.; Hamasaki, T.; Baba, M.; Cheng, Y.C. Impact of novel human immunodeficiency virus type 1 reverse transcriptase mutations P119S and T165A on 4’-ethynylthymidine analog resistance profile. Antimicrob. Agents Chemother. 2009, 53, 4640–4646. [Google Scholar] [CrossRef]
- Ghoke, S.S.; Sood, R.; Kumar, N.; Pateriya, A.K.; Bhatia, S.; Mishra, A.; Dixit, R.; Singh, V.K.; Desai, D.N.; Kulkarni, D.D.; et al. Evaluation of antiviral activity of Ocimum sanctum and Acacia arabica leaves extracts against H9N2 virus using embryonated chicken egg model. BMC Complement. Altern. Med. 2018, 18, 174. [Google Scholar] [CrossRef]
- Hussein, H.A.M.; Akula, S.M. Profiling of cellular microRNA responses during the early stages of KSHV infection. Arch. Virol. 2017, 162, 3293–3303. [Google Scholar] [CrossRef] [PubMed]
- Magdeldin, S.; Yamamoto, T.; Tooyama, I.; Abdelalim, E.M. New proteomic insights on the role of NPR-A in regulating self-renewal of embryonic stem cells. Stem Cell Rev. Rep. 2014, 10, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Ezzeldin, S.; El-Wazir, A.; Enany, S.; Muhammad, A.; Johar, D.; Osama, A.; Ahmed, E.; Shikshaky, H.; Magdeldin, S. Current Understanding of Human Metaproteome Association and Modulation. J. Proteome Res. 2019, 18, 3539–3554. [Google Scholar] [CrossRef] [PubMed]
- Neilson, K.A.; Keighley, T.; Pascovici, D.; Cooke, B.; Haynes, P.A. Label-free quantitative shotgun proteomics using normalized spectral abundance factors. Methods Mol. Biol. 2013, 1002, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef]
- R Core Team, R. Language and Environment for Statistical Computing. 2018. Available online: https://www.r-project.org/ (accessed on 15 December 2021).
- Kolde, R. pheatmap: Pretty Heatmaps. 2019. Available online: https://cran.r-project.org/package=pheatmap (accessed on 15 December 2021).
- Li, S.; Chen, W.; Shen, Y.; Xia, J.; Fan, S.; Li, N.; Luo, Y.; Han, X.; Cui, M.; Zhao, Y.; et al. Molecular characterization of infectious bronchitis virus in Southwestern China for the protective efficacy evaluation of four live vaccine strains. Vaccine 2021, 40, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Korneta, P.; Zawiła-Niedźwiecki, J.; Domański, J. Mutual relationships between SARS-CoV-2 test numbers, fatality and morbidity rates. BMC Public Health 2021, 21, 1980. [Google Scholar] [CrossRef] [PubMed]
- Dudley, J.P.; Lee, N.T. Disparities in Age-specific Morbidity and Mortality From SARS-CoV-2 in China and the Republic of Korea. Clin. Infect. Dis. 2020, 71, 863–865. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.M.; Monogue, M.L.; Jodlowski, T.Z.; Cutrell, J.B. Pharmacologic Treatments for Coronavirus Disease 2019 (COVID-19): A Review. JAMA 2020, 323, 1824–1836. [Google Scholar] [CrossRef] [PubMed]
- Drożdżal, S.; Rosik, J.; Lechowicz, K.; Machaj, F.; Szostak, B.; Przybyciński, J.; Lorzadeh, S.; Kotfis, K.; Ghavami, S.; Łos, M.J. An update on drugs with therapeutic potential for SARS-CoV-2 (COVID-19) treatment. Drug Resist. Updates 2021, 100794, 100794. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.W.X.; Zhang, J.; Zhang, K.; Zou, W.; Zhang, K.; Wang, L.; Guo, Z.; Qiu, Z.; Wang, G.; Xin, R.; et al. Combined Effect of Shegandilong Granule and Doxycycline on Immune Responses and Protection Against Avian Infectious Bronchitis Virus in Broilers. Front. Vet. Sci. 2021, 8, 756629. [Google Scholar] [CrossRef] [PubMed]
- Abbas, G.; Zhang, Y.; Sun, X.; Chen, H.; Ren, Y.; Wang, X.; Ahmad, M.Z.; Huang, X.; Li, G. Molecular Characterization of Infectious Bronchitis Virus Strain HH06 Isolated in a Poultry Farm in Northeastern China. Front. Vet. Sci. 2021, 8, 794228. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, C.Y.; Wan, X.-F. Antigenic characterization of influenza and SARS-CoV-2 viruses. Anal. Bioanal. Chem. 2021, 414, 2841–2881. [Google Scholar] [CrossRef]
- Hu, C.; Zhao, L.; Zhang, L.; Bao, Q.; Li, L. Mesenchymal stem cell-based cell-free strategies: Safe and effective treatments for liver injury. Stem Cell Res. Ther. 2020, 11, 377. [Google Scholar] [CrossRef]
- Petrou, P.; Gothelf, Y.; Argov, Z.; Gotkine, M.; Levy, Y.S.; Kassis, I.; Vaknin-Dembinsky, A.; Ben-Hur, T.; Offen, D.; Abramsky, O.; et al. Safety and Clinical Effects of Mesenchymal Stem Cells Secreting Neurotrophic Factor Transplantation in Patients With Amyotrophic Lateral Sclerosis: Results of Phase 1/2 and 2a Clinical Trials. JAMA Neurol. 2016, 73, 337–344. [Google Scholar] [CrossRef]
- Buigues, A.; Marchante, M.; de Miguel-Gómez, L.; Martinez, J.; Cervelló, I.; Pellicer, A.; Herraiz, S. Stem cell-secreted factor therapy regenerates the ovarian niche and rescues follicles. Am. J. Obstet. Gynecol. 2021, 225, e61–e65. [Google Scholar] [CrossRef] [PubMed]
- Basiri, A.; Mansouri, F.; Azari, A.; Ranjbarvan, P.; Zarein, F.; Heidari, A.; Golchin, A. Stem Cell Therapy Potency in Personalizing Severe COVID-19 Treatment. Stem Cell Rev. Rep. 2021, 17, 193–213. [Google Scholar] [CrossRef] [PubMed]
- Amable, P.R.; Teixeira, M.V.; Carias, R.B.; Granjeiro, J.M.; Borojevic, R. Protein synthesis and secretion in human mesenchymal cells derived from bone marrow, adipose tissue and Wharton’s jelly. Stem Cell Res. 2014, 5, 53. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.L.; Medicetty, S.; Bledsoe, A.R.; Rachakatla, R.S.; Choi, M.; Merchav, S.; Luo, Y.; Rao, M.S.; Velagaleti, G.; Troyer, D. Human Umbilical Cord Matrix Stem Cells: Preliminary Characterization and Effect of Transplantation in a Rodent Model of Parkinson’s Disease. Stem Cells 2005, 24, 781–792. [Google Scholar] [CrossRef]
- Chen, M.Y.; Lie, P.C.; Li, Z.L.; Wei, X. Endothelial differentiation of Wharton’s jelly-derived mesenchymal stem cells in comparison with bone marrow-derived mesenchymal stem cells. Exp. Hematol. 2009, 37, 629–640. [Google Scholar] [CrossRef]
- Ranjbaran, H.; Abediankenari, S.; Mohammadi, M.; Jafari, N.; Khalilian, A.; Rahmani, Z.; Momeninezhad Amiri, M.; Ebrahimi, P. Wharton’s Jelly Derived-Mesenchymal Stem Cells: Isolation and Characterization. Acta Med. Iran. 2018, 56, 28–33. [Google Scholar]
- Kint, J.; Dickhout, A.; Kutter, J.; Maier, H.J.; Britton, P.; Koumans, J.; Pijlman, G.P.; Fros, J.J.; Wiegertjes, G.F.; Forlenza, M. Infectious Bronchitis Coronavirus Inhibits STAT1 Signaling and Requires Accessory Proteins for Resistance to Type I Interferon Activity. J. Virol. 2015, 89, 12047–12057. [Google Scholar] [CrossRef]
- Zheng, B.; He, M.L.; Wong, K.L.; Lum, C.T.; Poon, L.L.; Peng, Y.; Guan, Y.; Lin, M.C.; Kung, H.F. Potent inhibition of SARS-associated coronavirus (SCOV) infection and replication by type I interferons (IFN-alpha/beta) but not by type II interferon (IFN-gamma). J. Interferon. Cytokine Res. 2004, 24, 388–390. [Google Scholar] [CrossRef]
- Sze, S.K.; de Kleijn, D.P.; Lai, R.C.; Khia Way Tan, E.; Zhao, H.; Yeo, K.S.; Low, T.Y.; Lian, Q.; Lee, C.N.; Mitchell, W.; et al. Elucidating the secretion proteome of human embryonic stem cell-derived mesenchymal stem cells. Mol. Cell. Proteom. MCP 2007, 6, 1680–1689. [Google Scholar] [CrossRef]
- Joseph, A.; Baiju, I.; Bhat, I.A.; Pandey, S.; Bharti, M.; Verma, M.; Pratap Singh, A.; Ansari, M.M.; Chandra, V.; Saikumar, G.; et al. Mesenchymal stem cell-conditioned media: A novel alternative of stem cell therapy for quality wound healing. J. Cell. Physiol. 2020, 235, 5555–5569. [Google Scholar] [CrossRef]
- Yu, R.; Mao, Y.; Li, K.; Zhai, Y.; Zhang, Y.; Liu, S.; Gao, Y.; Chen, Z.; Liu, Y.; Fang, T.; et al. Recombinant Human Thymosin Beta-4 Protects against Mouse Coronavirus Infection. Mediat. Inflamm. 2021, 2021, 9979032. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Hayano, T.; Suzuki, M. Peptidyl-prolyl cis-trans isomerase is the cyclosporin A-binding protein cyclophilin. Nature 1989, 337, 473–475. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhao, Z.; Xu, C.; Sun, L.; Chen, J.; Zhang, L.; Liu, W. Cyclophilin A restricts influenza A virus replication through degradation of the M1 protein. PLoS ONE 2012, 7, e31063. [Google Scholar] [CrossRef]
- Hu, J.; Van den Steen, P.E.; Sang, Q.-X.A.; Opdenakker, G. Matrix metalloproteinase inhibitors as therapy for inflammatory and vascular diseases. Nat. Rev. Drug Discov. 2007, 6, 480–498. [Google Scholar] [CrossRef] [PubMed]
- Lozito, T.P.; Jackson, W.M.; Nesti, L.J.; Tuan, R.S. Human mesenchymal stem cells generate a distinct pericellular zone of MMP activities via binding of MMPs and secretion of high levels of TIMPs. Matrix Biol. 2014, 34, 132–143. [Google Scholar] [CrossRef]
- Han, T.; Liu, Y.; Liu, H.; Zhu, Z.Y.; Li, Y.; Xiao, S.X.; Guo, Z.; Zhao, Z.G. Serum thymosin beta4 levels in patients with hepatitis B virus-related liver failure. World J. Gastroenterol. 2010, 16, 625–630. [Google Scholar] [CrossRef]
- Zheng, Q.N.; Xu, M.Y.; Gan, F.M.; Ye, S.S.; Zhao, H. Thymosin as a possible therapeutic drug for COVID-19: A case report. World J. Clin. Cases 2021, 9, 4090–4094. [Google Scholar] [CrossRef]
- Towers, G.J.; Hatziioannou, T.; Cowan, S.; Goff, S.P.; Luban, J.; Bieniasz, P.D. Cyclophilin A modulates the sensitivity of HIV-1 to host restriction factors. Nat. Med. 2003, 9, 1138–1143. [Google Scholar] [CrossRef]
- Tian, L.; Liu, W.; Sun, L. Role of cyclophilin A during coronavirus replication and the antiviral activities of its inhibitors. Chin. J. Biotechnol. 2020, 36, 605–611. [Google Scholar] [CrossRef]
- Liu, X.; Sun, L.; Yu, M.; Wang, Z.; Xu, C.; Xue, Q.; Zhang, K.; Ye, X.; Kitamura, Y.; Liu, W. Cyclophilin A interacts with influenza A virus M1 protein and impairs the early stage of the viral replication. Cell. Microbiol. 2009, 11, 730–741. [Google Scholar] [CrossRef]
- Laurie, K.; Holcomb, D.; Kames, J.; Komar, A.A.; DiCuccio, M.; Ibla, J.C.; Kimchi-Sarfaty, C. In Silico Evaluation of Cyclophilin Inhibitors as Potential Treatment for SARS-CoV-2. Open Forum Infect. Dis. 2021, 8, ofab189. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; von Brunn, A.; Zhu, D. Cyclophilin A and CD147: Novel therapeutic targets for the treatment of COVID-19. Med. Drug. Discov. 2020, 7, 100056. [Google Scholar] [CrossRef] [PubMed]
- Guizani, I.; Fourti, N.; Zidi, W.; Feki, M.; Allal-Elasmi, M. SARS-CoV-2 and pathological matrix remodeling mediators. Inflamm. Res. 2021, 70, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Kanbarkar, N.; Mishra, S. Matrix metalloproteinase inhibitors identified from Camellia sinensis for COVID-19 prophylaxis: An in silico approach. Adv. Tradit. Med. 2021, 21, 173–188. [Google Scholar] [CrossRef]
- Solun, B.; Shoenfeld, Y. Inhibition of metalloproteinases in therapy for severe lung injury due to COVID-19. Med. Drug Discov. 2020, 7, 100052. [Google Scholar] [CrossRef]
- Bazzoni, R.; Takam Kamga, P.; Tanasi, I.; Krampera, M. Extracellular Vesicle-Dependent Communication Between Mesenchymal Stromal Cells and Immune Effector Cells. Front. Cell Dev. Biol. 2020, 8, 596079. [Google Scholar] [CrossRef]
- Koch, M.; Lemke, A.; Lange, C. Extracellular Vesicles from MSC Modulate the Immune Response to Renal Allografts in a MHC Disparate Rat Model. Stem Cells Int. 2015, 2015, 486141. [Google Scholar] [CrossRef]
- Morrison, T.J.; Jackson, M.V.; Cunningham, E.K.; Kissenpfennig, A.; McAuley, D.F.; O’Kane, C.M.; Krasnodembskaya, A.D. Mesenchymal Stromal Cells Modulate Macrophages in Clinically Relevant Lung Injury Models by Extracellular Vesicle Mitochondrial Transfer. Am. J. Respir. Crit. Care Med. 2017, 196, 1275–1286. [Google Scholar] [CrossRef]
- Hyvärinen, K.; Holopainen, M.; Skirdenko, V.; Ruhanen, H.; Lehenkari, P.; Korhonen, M.; Käkelä, R.; Laitinen, S.; Kerkelä, E. Mesenchymal Stromal Cells and Their Extracellular Vesicles Enhance the Anti-Inflammatory Phenotype of Regulatory Macrophages by Downregulating the Production of Interleukin (IL)-23 and IL-22. Front. Immunol. 2018, 9, 771. [Google Scholar] [CrossRef]
- Ikhlas, S.; Usman, A.; Kim, D.; Cai, D. Exosomes/microvesicles target SARS-CoV-2 via innate and RNA-induced immunity with PIWI-piRNA system. Life Sci. Alliance 2022, 5, e202101240. [Google Scholar] [CrossRef]
- Yu, B.; Ikhlas, S.; Ruan, C.; Zhong, X.; Cai, D. Innate and Adaptive Immunity of Murine Neural Stem Cell-Derived piRNA Exosomes/Microvesicles against Pseudotyped SARS-CoV-2 and HIV-Based Lentivirus. IScience 2020, 23, 101806. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Xu, F.; Zhang, X.; Mu, J.; Sayed, M.; Hu, X.; Lei, C.; Sriwastva, M.; Kumar, A.; Sundaram, K.; et al. Plant-derived exosomal microRNAs inhibit lung inflammation induced by exosomes SARS-CoV-2 Nsp12. Mol. Ther. 2021, 29, 2424–2440. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Huang, F.; Xia, B.; Yuan, Y.; Yu, F.; Wang, G.; Chen, Q.; Wang, Q.; Li, Y.; Li, R.; et al. The interferon-stimulated exosomal hACE2 potently inhibits SARS-CoV-2 replication through competitively blocking the virus entry. Signal Transduct Target 2021, 6, 189. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Cell | CC50 (µg/mL) | IC50 (µg/mL) | SI (CC50/IC50) |
|---|---|---|---|---|
| SARS-CoV-2 | Vero-E6 | 39,200 | 166.6 | 235.29 |
| IBV | Vero-E6 | 3900 | 439.9 | 8.87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussein, M.A.A.; Hussein, H.A.M.; Thabet, A.A.; Selim, K.M.; Dawood, M.A.; El-Adly, A.M.; Wardany, A.A.; Sobhy, A.; Magdeldin, S.; Osama, A.; et al. Human Wharton’s Jelly Mesenchymal Stem Cells Secretome Inhibits Human SARS-CoV-2 and Avian Infectious Bronchitis Coronaviruses. Cells 2022, 11, 1408. https://doi.org/10.3390/cells11091408
Hussein MAA, Hussein HAM, Thabet AA, Selim KM, Dawood MA, El-Adly AM, Wardany AA, Sobhy A, Magdeldin S, Osama A, et al. Human Wharton’s Jelly Mesenchymal Stem Cells Secretome Inhibits Human SARS-CoV-2 and Avian Infectious Bronchitis Coronaviruses. Cells. 2022; 11(9):1408. https://doi.org/10.3390/cells11091408
Chicago/Turabian StyleHussein, Mohamed A. A., Hosni A. M. Hussein, Ali A. Thabet, Karim M. Selim, Mervat A. Dawood, Ahmed M. El-Adly, Ahmed A. Wardany, Ali Sobhy, Sameh Magdeldin, Aya Osama, and et al. 2022. "Human Wharton’s Jelly Mesenchymal Stem Cells Secretome Inhibits Human SARS-CoV-2 and Avian Infectious Bronchitis Coronaviruses" Cells 11, no. 9: 1408. https://doi.org/10.3390/cells11091408
APA StyleHussein, M. A. A., Hussein, H. A. M., Thabet, A. A., Selim, K. M., Dawood, M. A., El-Adly, A. M., Wardany, A. A., Sobhy, A., Magdeldin, S., Osama, A., Anwar, A. M., Abdel-Wahab, M., Askar, H., Bakhiet, E. K., Sultan, S., Ezzat, A. A., Abdel Raouf, U., & Afifi, M. M. (2022). Human Wharton’s Jelly Mesenchymal Stem Cells Secretome Inhibits Human SARS-CoV-2 and Avian Infectious Bronchitis Coronaviruses. Cells, 11(9), 1408. https://doi.org/10.3390/cells11091408