
Blood-Based Biomarkers for Alzheimer’s Disease Diagnosis and Progression: An Overview

 , , ,
, , ,  , ,
, ,  ,
,  and
and
Abstract

1. Introduction
2. Methods
3. Results
3.1. Long-Studied and Well-Known Biomarkers: Amyloid-β Peptides and Tau
3.2. Plasma Neurofilament Light
3.3. Inflammation
3.3.1. Inflammatory Molecules
3.3.2. Circulating Cytokines
3.4. Metabolism
3.5. Oxidative Stress
3.6. Circulating Non-Coding RNAs
3.6.1. miRNAs—Alzheimer’s Disease
3.6.2. miRNAs—Early Diagnosis
3.6.3. miRNAs—Exosomes
3.6.4. miRNAs—Limitations
3.6.5. Long Non-Coding RNAs
3.7. Lipids
3.8. Vitamins
3.8.1. Water-Soluble Vitamins: Vitamins B and C
3.8.2. Liposoluble Vitamins: Vitamins D, A, and E
3.9. Gut Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hodson, R. Alzheimer’s Disease. Nature 2018, 559, S1. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P. Pathways towards and away from Alzheimer’s Disease. Nature 2004, 430, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Bachurin, S.O.; Bovina, E.V.; Ustyugov, A.A. Drugs in Clinical Trials for Alzheimer’s Disease: The Major Trends. Med. Res. Rev. 2017, 37, 1186–1225. [Google Scholar] [CrossRef] [PubMed]
- Arnerić, S.P.; Batrla-Utermann, R.; Beckett, L.; Bittner, T.; Blennow, K.; Carter, L.; Dean, R.; Engelborghs, S.; Genius, J.; Gordon, M.F.; et al. Cerebrospinal Fluid Biomarkers for Alzheimer’s Disease: A View of the Regulatory Science Qualification Landscape from the Coalition Against Major Diseases CSF Biomarker Team. J. Alzheimer’s Dis. 2016, 55, 19–35. [Google Scholar] [CrossRef]
- Mattsson, N.; Carrillo, M.C.; Dean, R.A.; Devous, M.D.; Nikolcheva, T.; Pesini, P.; Salter, H.; Potter, W.Z.; Sperling, R.S.; Bateman, R.J.; et al. Revolutionizing Alzheimer’s Disease and Clinical Trials through Biomarkers. Alzheimer’s Dement. Diagn. Assess. Dis. Monit. 2015, 1, 412–419. [Google Scholar] [CrossRef]
- Schneider, L.S.; Mangialasche, F.; Andreasen, N.; Feldman, H.; Giacobini, E.; Jones, R.; Mantua, V.; Mecocci, P.; Pani, L.; Winblad, B.; et al. Clinical Trials and Late-Stage Drug Development for Alzheimer’s Disease: An Appraisal from 1984 to 2014. J. Intern. Med. 2014, 275, 251–283. [Google Scholar] [CrossRef]
- Galasko, D.R.; Grill, J.D.; Lingler, J.H.; Heidebrink, J.L. A Blood Test for Alzheimer’s Disease: It’s about Time or Not Ready for Prime Time? J. Alzheimer’s Dis. 2022; in press. [Google Scholar] [CrossRef]
- Molinuevo, J.L.; Ayton, S.; Batrla, R.; Bednar, M.M.; Bittner, T.; Cummings, J.; Fagan, A.M.; Hampel, H.; Mielke, M.M.; Mikulskis, A.; et al. Current State of Alzheimer’s Fluid Biomarkers. Acta Neuropathol. 2018, 136, 821–853. [Google Scholar] [CrossRef]
- Burnham, S.C.; Rowe, C.C.; Baker, D.; Bush, A.I.; Doecke, J.D.; Faux, N.G.; Laws, S.M.; Martins, R.N.; Maruff, P.; Macaulay, S.L.; et al. Predicting Alzheimer Disease from a Blood-Based Biomarker Profile. Neurology 2016, 87, 1093–1101. [Google Scholar] [CrossRef]
- García, S.; Amor-Gutiérrez, O.; Palomares-Albarrán, M.; Toyos-Rodríguez, C.; Cuetos, F.; Martínez, C.; Costa-García, A.; Fernández-Sánchez, M.T.; de la Escosura-Muñiz, A.; Novelli, A. Unfolded P53 as a Marker of Oxidative Stress in Mild Cognitive Impairment, Alzheimer’s and Parkinson’s Disease. Curr. Alzheimer Res. 2021, 18, 695–700. [Google Scholar] [CrossRef]
- Hampel, H.; O’Bryant, S.E.; Molinuevo, J.L.; Zetterberg, H.; Masters, C.L.; Lista, S.; Kiddle, S.J.; Batrla, R.; Blennow, K. Blood-Based Biomarkers for Alzheimer Disease: Mapping the Road to the Clinic. Nat. Rev. Neurol. 2018, 14, 639–652. [Google Scholar] [CrossRef]
- Hampel, H.; O’Bryant, S.E.; Castrillo, J.I.; Ritchie, C.; Rojkova, K.; Broich, K.; Benda, N.; Nisticò, R.; Frank, R.A.; Dubois, B.; et al. Precision medicine—The Golden Gate for Detection, Treatment and Prevention of Alzheimer’s Disease. J. Prev. Alzheimer’s Dis. 2016, 3, 243. [Google Scholar] [CrossRef] [PubMed]
- Hampel, H.; O’Bryant, S.E.; Durrleman, S.; Younesi, E.; Rojkova, K.; Escott-Price, V.; Corvol, J.-C.; Broich, K.; Dubois, B.; Lista, S. A Precision Medicine Initiative for Alzheimer’s Disease: The Road Ahead to Biomarker-Guided Integrative Disease Modeling. Climacteric 2017, 20, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Hampel, H.; Lista, S.; Teipel, S.J.; Garaci, F.; Nisticò, R.; Blennow, K.; Zetterberg, H.; Bertram, L.; Duyckaerts, C.; Bakardjian, H.; et al. Perspective on Future Role of Biological Markers in Clinical Therapy Trials of Alzheimer’s Disease: A Long-Range Point of View beyond 2020. Biochem. Pharmacol. 2014, 88, 426–449. [Google Scholar] [CrossRef] [PubMed]
- Blennow, K.; Zetterberg, H. Biomarkers for Alzheimer’s Disease: Current Status and Prospects for the Future. J. Intern. Med. 2018, 284, 643–663. [Google Scholar] [CrossRef] [PubMed]
- Mahaman, Y.A.R.; Embaye, K.S.; Huang, F.; Li, L.; Zhu, F.; Wang, J.-Z.; Liu, R.; Feng, J.; Wang, X. Biomarkers Used in Alzheimer’s Disease Diagnosis, Treatment, and Prevention. Ageing Res. Rev. 2022, 74, 101544. [Google Scholar] [CrossRef] [PubMed]
- Leidinger, P.; Backes, C.; Deutscher, S.; Schmitt, K.; Mueller, S.C.; Frese, K.; Haas, J.; Ruprecht, K.; Paul, F.; Stähler, C.; et al. A Blood Based 12-MiRNA Signature of Alzheimer Disease Patients. Genome Biol. 2013, 14, R78. [Google Scholar] [CrossRef]
- Olazarán, J.; Gil-de-Gómez, L.; Rodríguez-Martín, A.; Valentí-Soler, M.; Frades-Payo, B.; Marín-Muñoz, J.; Antúnez, C.; Frank-García, A.; Acedo-Jiménez, C.; Morlán-Gracia, L.; et al. A Blood-Based, 7-Metabolite Signature for the Early Diagnosis of Alzheimer’s Disease. J. Alzheimer’s Dis. 2015, 45, 1157–1173. [Google Scholar] [CrossRef]
- Yun, G.; Kim, H.J.; Kim, H.-G.; Lee, K.M.; Hong, I.K.; Kim, S.H.; Rhee, H.Y.; Jahng, G.-H.; Yoon, S.S.; Park, K.-C.; et al. Association Between Plasma Amyloid-β and Neuropsychological Performance in Patients with Cognitive Decline. Front. Aging Neurosci. 2021, 13, 736937. [Google Scholar] [CrossRef]
- Shahpasand-Kroner, H.; Klafki, H.-W.; Bauer, C.; Schuchhardt, J.; Hüttenrauch, M.; Stazi, M.; Bouter, C.; Wirths, O.; Vogelgsang, J.; Wiltfang, J. A Two-Step Immunoassay for the Simultaneous Assessment of Aβ38, Aβ40 and Aβ42 in Human Blood Plasma Supports the Aβ42/Aβ40 Ratio as a Promising Biomarker Candidate of Alzheimer’s Disease. Alzheimer’s Res. Ther. 2018, 10, 121. [Google Scholar] [CrossRef]
- Chen, T.-B.; Lin, K.-J.; Lin, S.-Y.; Lee, Y.-J.; Lin, Y.-C.; Wang, C.-Y.; Chen, J.-P.; Wang, P.-N. Prediction of Cerebral Amyloid Pathology Based on Plasma Amyloid and Tau Related Markers. Front. Neurol. 2021, 12, 619388. [Google Scholar] [CrossRef]
- Rembach, A.; Faux, N.G.; Watt, A.D.; Pertile, K.K.; Rumble, R.L.; Trounson, B.O.; Fowler, C.J.; Roberts, B.R.; Perez, K.A.; Li, Q.; et al. Changes in Plasma Amyloid Beta in a Longitudinal Study of Aging and Alzheimer’s Disease. Alzheimer’s Dement. 2014, 10, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.J.; Yi, S.; Han, J.; Park, S.Y.; Jang, J.-W.; Chun, I.K.; Kim, S.E.; Lee, B.S.; Kim, G.J.; Yu, J.S.; et al. Oligomeric Forms of Amyloid-β Protein in Plasma as a Potential Blood-Based Biomarker for Alzheimer’s Disease. Alzheimer’s Res. Ther. 2017, 9, 98. [Google Scholar] [CrossRef] [PubMed]
- Lui, J.K.; Laws, S.M.; Li, Q.-X.; Villemagne, V.L.; Ames, D.; Brown, B.; Bush, A.I.; de Ruyck, K.; Dromey, J.; Ellis, K.A.; et al. Plasma Amyloid-β as a Biomarker in Alzheimer’s Disease: The AIBL Study of Aging. J. Alzheimer’s Dis. 2010, 20, 1233–1242. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.R.; Ashton, N.J.; Karikari, T.K.; Tanaka, T.; Saridin, F.N.; Reilhac, A.; Robins, E.G.; Nai, Y.; Vrooman, H.; Hilal, S.; et al. Plasma P-tau181 to Aβ42 Ratio Is Associated with Brain Amyloid Burden and Hippocampal Atrophy in an Asian Cohort of Alzheimer’s Disease Patients with Concomitant Cerebrovascular Disease. Alzheimer’s Dement. 2021, 17, 1649–1662. [Google Scholar] [CrossRef]
- Chouraki, V.; Beiser, A.; Younkin, L.; Preis, S.R.; Weinstein, G.; Hansson, O.; Skoog, I.; Lambert, J.; Au, R.; Launer, L.; et al. Plasma Amyloid-β and Risk of Alzheimer’s Disease in the Framingham Heart Study. Alzheimer’s Dement. 2015, 11, 249. [Google Scholar] [CrossRef]
- Meng, X.; Li, T.; Wang, X.; Lv, X.; Sun, Z.; Zhang, J.; Su, F.; Kang, S.; Kim, S.; An, S.S.A.; et al. Association between Increased Levels of Amyloid-β Oligomers in Plasma and Episodic Memory Loss in Alzheimer’s Disease. Alzheimer’s Res. Ther. 2019, 11, 89. [Google Scholar] [CrossRef]
- Chatterjee, P.; Elmi, M.; Goozee, K.; Shah, T.; Sohrabi, H.R.; Dias, C.B.; Pedrini, S.; Shen, K.; Asih, P.R.; Dave, P.; et al. Ultrasensitive Detection of Plasma Amyloid-β as a Biomarker for Cognitively Normal Elderly Individuals at Risk of Alzheimer’s Disease. J. Alzheimer’s Dis. 2019, 71, 775–783. [Google Scholar] [CrossRef]
- Jiao, B.; Liu, H.; Guo, L.; Liao, X.; Zhou, Y.; Weng, L.; Xiao, X.; Zhou, L.; Wang, X.; Jiang, Y.; et al. Performance of Plasma Amyloid β, Total Tau, and Neurofilament Light Chain in the Identification of Probable Alzheimer’s Disease in South China. Front. Aging Neurosci. 2021, 13, 749649. [Google Scholar] [CrossRef]
- Shi, Y.; Lu, X.; Zhang, L.; Shu, H.; Gu, L.; Wang, Z.; Gao, L.; Zhu, J.; Zhang, H.; Zhou, D.; et al. Potential Value of Plasma Amyloid-β, Total Tau, and Neurofilament Light for Identification of Early Alzheimer’s Disease. ACS Chem. Neurosci. 2019, 10, 3479–3485. [Google Scholar] [CrossRef]
- Stockmann, J.; Verberk, I.M.W.; Timmesfeld, N.; Denz, R.; Budde, B.; Lange-Leifhelm, J.; Scheltens, P.; van der Flier, W.M.; Nabers, A.; Teunissen, C.E.; et al. Amyloid-β Misfolding as a Plasma Biomarker Indicates Risk for Future Clinical Alzheimer’s Disease in Individuals with Subjective Cognitive Decline. Alzheimer’s Res. Ther. 2020, 12, 169. [Google Scholar] [CrossRef]
- Shen, X.; Li, J.; Wang, H.; Li, H.; Huang, Y.; Yang, Y.; Tan, L.; Dong, Q.; Yu, J. Plasma Amyloid, Tau, and Neurodegeneration Biomarker Profiles Predict Alzheimer’s Disease Pathology and Clinical Progression in Older Adults without Dementia. Alzheimer’s Dement. Diagn. Assess. Dis. Monit. 2020, 12, e12104. [Google Scholar] [CrossRef] [PubMed]
- de Wolf, F.; Ghanbari, M.; Licher, S.; McRae-McKee, K.; Gras, L.; Weverling, G.J.; Wermeling, P.; Sedaghat, S.; Ikram, M.K.; Waziry, R.; et al. Plasma Tau, Neurofilament Light Chain and Amyloid-β Levels and Risk of Dementia; a Population-Based Cohort Study. Brain 2020, 143, 1220–1232. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.Y.; Maruff, P.; Kaneko, N.; Doecke, J.; Fowler, C.; Villemagne, V.L.; Kato, T.; Rowe, C.C.; Arahata, Y.; Iwamoto, S.; et al. Plasma Amyloid-β Biomarker Associated with Cognitive Decline in Preclinical Alzheimer’s Disease. J. Alzheimer’s Dis. 2020, 77, 1057–1065. [Google Scholar] [CrossRef] [PubMed]
- Nabers, A.; Perna, L.; Lange, J.; Mons, U.; Schartner, J.; Güldenhaupt, J.; Saum, K.; Janelidze, S.; Holleczek, B.; Rujescu, D.; et al. Amyloid Blood Biomarker Detects Alzheimer’s Disease. EMBO Mol. Med. 2018, 10, e8763. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Kaneko, N.; Villemagne, V.L.; Kato, T.; Doecke, J.; Doré, V.; Fowler, C.; Li, Q.-X.; Martins, R.; Rowe, C.; et al. High Performance Plasma Amyloid-β Biomarkers for Alzheimer’s Disease. Nature 2018, 554, 249–254. [Google Scholar] [CrossRef]
- Villemagne, V.L.; Burnham, S.; Bourgeat, P.; Brown, B.; Ellis, K.A.; Salvado, O.; Szoeke, C.; Macaulay, S.L.; Martins, R.; Maruff, P.; et al. Amyloid β Deposition, Neurodegeneration, and Cognitive Decline in Sporadic Alzheimer’s Disease: A Prospective Cohort Study. Lancet Neurol. 2013, 12, 357–367. [Google Scholar] [CrossRef]
- Vos, S.J.; Xiong, C.; Visser, P.J.; Jasielec, M.S.; Hassenstab, J.; Grant, E.A.; Cairns, N.J.; Morris, J.C.; Holtzman, D.M.; Fagan, A.M. Preclinical Alzheimer’s Disease and Its Outcome: A Longitudinal Cohort Study. Lancet Neurol. 2013, 12, 957–965. [Google Scholar] [CrossRef]
- Janelidze, S.; Stomrud, E.; Palmqvist, S.; Zetterberg, H.; van Westen, D.; Jeromin, A.; Song, L.; Hanlon, D.; Tan Hehir, C.A.; Baker, D.; et al. Plasma β-Amyloid in Alzheimer’s Disease and Vascular Disease. Sci. Rep. 2016, 6, 26801. [Google Scholar] [CrossRef]
- Ashton, N.J.; Schöll, M.; Heurling, K.; Gkanatsiou, E.; Portelius, E.; Höglund, K.; Brinkmalm, G.; Hye, A.; Blennow, K.; Zetterberg, H. Update on Biomarkers for Amyloid Pathology in Alzheimer’s Disease. Biomark. Med. 2018, 12, 799–812. [Google Scholar] [CrossRef]
- Palmqvist, S.; Janelidze, S.; Stomrud, E.; Zetterberg, H.; Karl, J.; Zink, K.; Bittner, T.; Mattsson, N.; Eichenlaub, U.; Blennow, K.; et al. Performance of Fully Automated Plasma Assays as Screening Tests for Alzheimer Disease–Related β-Amyloid Status. JAMA Neurol. 2019, 76, 1060. [Google Scholar] [CrossRef]
- Guo, Y.; Huang, Y.-Y.; Shen, X.-N.; Chen, S.-D.; Hu, H.; Wang, Z.-T.; Tan, L.; Yu, J.-T. Characterization of Alzheimer’s Tau Biomarker Discordance Using Plasma, CSF, and PET. Alzheimer’s Res. Ther. 2021, 13, 93. [Google Scholar] [CrossRef] [PubMed]
- McGrowder, D.A.; Miller, F.; Vaz, K.; Nwokocha, C.; Wilson-Clarke, C.; Anderson-Cross, M.; Brown, J.; Anderson-Jackson, L.; Williams, L.; Latore, L.; et al. Cerebrospinal Fluid Biomarkers of Alzheimer’s Disease: Current Evidence and Future Perspectives. Brain Sci. 2021, 11, 215. [Google Scholar] [CrossRef] [PubMed]
- Chandra, A.; Valkimadi, P.; Pagano, G.; Cousins, O.; Dervenoulas, G.; Politis, M. Applications of Amyloid, Tau, and Neuroinflammation PET Imaging to Alzheimer’s Disease and Mild Cognitive Impairment. Hum. Brain Mapp. 2019, 40, 5424–5442. [Google Scholar] [CrossRef]
- Milà-Alomà, M.; Salvadó, G.; Gispert, J.D.; Vilor-Tejedor, N.; Grau-Rivera, O.; Sala-Vila, A.; Sánchez-Benavides, G.; Arenaza-Urquijo, E.M.; Crous-Bou, M.; González-de-Echávarri, J.M.; et al. Amyloid Beta, Tau, Synaptic, Neurodegeneration, and Glial Biomarkers in the Preclinical Stage of the Alzheimer’s Continuum. Alzheimer’s Dement. 2020, 16, 1358–1371. [Google Scholar] [CrossRef] [PubMed]
- Mielke, M.M.; Hagen, C.E.; Xu, J.; Chai, X.; Vemuri, P.; Lowe, V.J.; Airey, D.C.; Knopman, D.S.; Roberts, R.O.; Machulda, M.M.; et al. Plasma Phospho-tau181 Increases with Alzheimer’s Disease Clinical Severity and Is Associated with Tau- and Amyloid-positron Emission Tomography. Alzheimer’s Dement. 2018, 14, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Janelidze, S.; Mattsson, N.; Palmqvist, S.; Smith, R.; Beach, T.G.; Serrano, G.E.; Chai, X.; Proctor, N.K.; Eichenlaub, U.; Zetterberg, H.; et al. Plasma P-Tau181 in Alzheimer’s Disease: Relationship to Other Biomarkers, Differential Diagnosis, Neuropathology and Longitudinal Progression to Alzheimer’s Dementia. Nat. Med. 2020, 26, 379–386. [Google Scholar] [CrossRef]
- Karikari, T.K.; Pascoal, T.A.; Ashton, N.J.; Janelidze, S.; Benedet, A.L.; Rodriguez, J.L.; Chamoun, M.; Savard, M.; Kang, M.S.; Therriault, J.; et al. Blood Phosphorylated Tau 181 as a Biomarker for Alzheimer’s Disease: A Diagnostic Performance and Prediction Modelling Study Using Data from Four Prospective Cohorts. Lancet Neurol. 2020, 19, 422–433. [Google Scholar] [CrossRef]
- Thijssen, E.H.; la Joie, R.; Wolf, A.; Strom, A.; Wang, P.; Iaccarino, L.; Bourakova, V.; Cobigo, Y.; Heuer, H.; Spina, S.; et al. Diagnostic Value of Plasma Phosphorylated Tau181 in Alzheimer’s Disease and Frontotemporal Lobar Degeneration. Nat. Med. 2020, 26, 387–397. [Google Scholar] [CrossRef]
- Moscoso, A.; Grothe, M.J.; Ashton, N.J.; Karikari, T.K.; Rodriguez, J.L.; Snellman, A.; Suárez-Calvet, M.; Zetterberg, H.; Blennow, K.; Schöll, M. Time Course of Phosphorylated-Tau181 in Blood across the Alzheimer’s Disease Spectrum. Brain 2021, 144, 325–339. [Google Scholar] [CrossRef]
- Karikari, T.K.; Benedet, A.L.; Ashton, N.J.; Lantero Rodriguez, J.; Snellman, A.; Suárez-Calvet, M.; Saha-Chaudhuri, P.; Lussier, F.; Kvartsberg, H.; Rial, A.M.; et al. Diagnostic Performance and Prediction of Clinical Progression of Plasma Phospho-Tau181 in the Alzheimer’s Disease Neuroimaging Initiative. Mol. Psychiatry 2021, 26, 429–442. [Google Scholar] [CrossRef]
- Janelidze, S.; Berron, D.; Smith, R.; Strandberg, O.; Proctor, N.K.; Dage, J.L.; Stomrud, E.; Palmqvist, S.; Mattsson-Carlgren, N.; Hansson, O. Associations of Plasma Phospho-Tau217 Levels with Tau Positron Emission Tomography in Early Alzheimer Disease. JAMA Neurol. 2021, 78, 149. [Google Scholar] [CrossRef] [PubMed]
- Janelidze, S.; Palmqvist, S.; Leuzy, A.; Stomrud, E.; Verberk, I.M.W.; Zetterberg, H.; Ashton, N.J.; Pesini, P.; Sarasa, L.; Allué, J.A.; et al. Detecting Amyloid Positivity in Early Alzheimer’s Disease Using Combinations of Plasma Aβ42/Aβ40 and P-tau. Alzheimer’s Dement. 2022, 18, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Stotani, S.; Giordanetto, F.; Medda, F. DYRK1A Inhibition as Potential Treatment for Alzheimer’s Disease. Future Med. Chem. 2016, 8, 681–696. [Google Scholar] [CrossRef] [PubMed]
- Delabar, J.M.; Ortner, M.; Simon, S.; Wijkhuisen, A.; Feraudet-Tarisse, C.; Pegon, J.; Vidal, E.; Hirschberg, Y.; Dubois, B.; Potier, M. Altered Age-linked Regulation of Plasma DYRK1A in Elderly Cognitive Complainers (INSIGHT-preAD Study) with High Brain Amyloid Load. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2020, 6, e12046. [Google Scholar] [CrossRef]
- Olsson, B.; Lautner, R.; Andreasson, U.; Öhrfelt, A.; Portelius, E.; Bjerke, M.; Hölttä, M.; Rosén, C.; Olsson, C.; Strobel, G.; et al. CSF and Blood Biomarkers for the Diagnosis of Alzheimer’s Disease: A Systematic Review and Meta-Analysis. Lancet Neurol. 2016, 15, 673–684. [Google Scholar] [CrossRef]
- Khalil, M.; Teunissen, C.E.; Otto, M.; Piehl, F.; Sormani, M.P.; Gattringer, T.; Barro, C.; Kappos, L.; Comabella, M.; Fazekas, F.; et al. Neurofilaments as Biomarkers in Neurological Disorders. Nat. Rev. Neurol. 2018, 14, 577–589. [Google Scholar] [CrossRef]
- Gisslén, M.; Price, R.W.; Andreasson, U.; Norgren, N.; Nilsson, S.; Hagberg, L.; Fuchs, D.; Spudich, S.; Blennow, K.; Zetterberg, H. Plasma Concentration of the Neurofilament Light Protein (NFL) Is a Biomarker of CNS Injury in HIV Infection: A Cross-Sectional Study. EBioMedicine 2016, 3, 135–140. [Google Scholar] [CrossRef]
- Kuhle, J.; Barro, C.; Andreasson, U.; Derfuss, T.; Lindberg, R.; Sandelius, Å.; Liman, V.; Norgren, N.; Blennow, K.; Zetterberg, H. Comparison of Three Analytical Platforms for Quantification of the Neurofilament Light Chain in Blood Samples: ELISA, Electrochemiluminescence Immunoassay and Simoa. Clin. Chem. Lab. Med. 2016, 54, 1655–1661. [Google Scholar] [CrossRef]
- Mattsson, N.; Andreasson, U.; Zetterberg, H.; Blennow, K. Association of Plasma Neurofilament Light With Neurodegeneration in Patients with Alzheimer Disease. JAMA Neurol. 2017, 74, 557. [Google Scholar] [CrossRef]
- Weston, P.S.J.; Poole, T.; Ryan, N.S.; Nair, A.; Liang, Y.; Macpherson, K.; Druyeh, R.; Malone, I.B.; Ahsan, R.L.; Pemberton, H.; et al. Serum Neurofilament Light in Familial Alzheimer Disease. Neurology 2017, 89, 2167–2175. [Google Scholar] [CrossRef]
- Rohrer, J.D.; Woollacott, I.O.C.; Dick, K.M.; Brotherhood, E.; Gordon, E.; Fellows, A.; Toombs, J.; Druyeh, R.; Cardoso, M.J.; Ourselin, S.; et al. Serum Neurofilament Light Chain Protein Is a Measure of Disease Intensity in Frontotemporal Dementia. Neurology 2016, 87, 1329–1336. [Google Scholar] [CrossRef] [PubMed]
- Rojas, J.C.; Karydas, A.; Bang, J.; Tsai, R.M.; Blennow, K.; Liman, V.; Kramer, J.H.; Rosen, H.; Miller, B.L.; Zetterberg, H.; et al. Plasma Neurofilament Light Chain Predicts Progression in Progressive Supranuclear Palsy. Ann. Clin. Transl. Neurol. 2016, 3, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Calsolaro, V.; Edison, P. Neuroinflammation in Alzheimer’s Disease: Current Evidence and Future Directions. Alzheimer’s Dement. 2016, 12, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Byrne, L.M.; Rodrigues, F.B.; Blennow, K.; Durr, A.; Leavitt, B.R.; Roos, R.A.C.; Scahill, R.I.; Tabrizi, S.J.; Zetterberg, H.; Langbehn, D.; et al. Neurofilament Light Protein in Blood as a Potential Biomarker of Neurodegeneration in Huntington’s Disease: A Retrospective Cohort Analysis. Lancet Neurol. 2017, 16, 601–609. [Google Scholar] [CrossRef]
- Buchhave, P.; Blennow, K.; Zetterberg, H.; Stomrud, E.; Londos, E.; Andreasen, N.; Minthon, L.; Hansson, O. Longitudinal Study of CSF Biomarkers in Patients with Alzheimer’s Disease. PLoS ONE 2009, 4, e6294. [Google Scholar] [CrossRef]
- Brunden, K.R.; Zhang, B.; Carroll, J.; Yao, Y.; Potuzak, J.S.; Hogan, A.-M.L.; Iba, M.; James, M.J.; Xie, S.X.; Ballatore, C.; et al. Epothilone D Improves Microtubule Density, Axonal Integrity, and Cognition in a Transgenic Mouse Model of Tauopathy. J. Neurosci. 2010, 30, 13861–13866. [Google Scholar] [CrossRef]
- Brosseron, F.; Traschütz, A.; Widmann, C.N.; Kummer, M.P.; Tacik, P.; Santarelli, F.; Jessen, F.; Heneka, M.T. Characterization and Clinical Use of Inflammatory Cerebrospinal Fluid Protein Markers in Alzheimer’s Disease. Alzheimer’s Res. Ther. 2018, 10, 25. [Google Scholar] [CrossRef]
- Bronzuoli, M.R.; Iacomino, A.; Steardo, L.; Scuderi, C. Targeting Neuroinflammation in Alzheimer’s Disease. J. Inflamm. Res. 2016, 9, 199–208. [Google Scholar] [CrossRef]
- Morgan, A.R.; Touchard, S.; Leckey, C.; O’Hagan, C.; Nevado-Holgado, A.J.; Barkhof, F.; Bertram, L.; Blin, O.; Bos, I.; Dobricic, V.; et al. Inflammatory Biomarkers in Alzheimer’s Disease Plasma. Alzheimer’s Dement. 2019, 15, 776–787. [Google Scholar] [CrossRef]
- Ferri, E.; Rossi, P.D.; Geraci, A.; Ciccone, S.; Cesari, M.; Arosio, B. The STREM2 Concentrations in the Blood: A Marker of Neurodegeneration? Front. Mol. Biosci. 2021, 7, 627931. [Google Scholar] [CrossRef]
- Tan, L.; Yu, J.-T.; Liu, Q.-Y.; Tan, M.-S.; Zhang, W.; Hu, N.; Wang, Y.-L.; Sun, L.; Jiang, T.; Tan, L. Circulating MiR-125b as a Biomarker of Alzheimer’s Disease. J. Neurol. Sci. 2014, 336, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Ochi, S.; Iga, J.; Funahashi, Y.; Yoshino, Y.; Yamazaki, K.; Kumon, H.; Mori, H.; Ozaki, Y.; Mori, T.; Ueno, S. Identifying Blood Transcriptome Biomarkers of Alzheimer’s Disease Using Transgenic Mice. Mol. Neurobiol. 2020, 57, 4941–4951. [Google Scholar] [CrossRef] [PubMed]
- Casati, M.; Ferri, E.; Gussago, C.; Mazzola, P.; Abbate, C.; Bellelli, G.; Mari, D.; Cesari, M.; Arosio, B. Increased Expression of TREM2 in Peripheral Cells from Mild Cognitive Impairment Patients Who Progress into Alzheimer’s Disease. Eur. J. Neurol. 2018, 25, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.J.; Ng, A.S.L.; Vipin, A.; Lim, J.K.W.; Chander, R.J.; Ji, F.; Qiu, Y.; Ting, S.K.S.; Hameed, S.; Lee, T.-S.; et al. Higher Peripheral TREM2 MRNA Levels Relate to Cognitive Deficits and Hippocampal Atrophy in Alzheimer’s Disease and Amnestic Mild Cognitive Impairment. J. Alzheimer’s Dis. 2017, 58, 413–423. [Google Scholar] [CrossRef]
- Hu, N.; Tan, M.-S.; Yu, J.-T.; Sun, L.; Tan, L.; Wang, Y.-L.; Jiang, T.; Tan, L. Increased Expression of TREM2 in Peripheral Blood of Alzheimer’s Disease Patients. J. Alzheimer’s Dis. 2013, 38, 497–501. [Google Scholar] [CrossRef]
- Sao, T.; Yoshino, Y.; Yamazaki, K.; Ozaki, Y.; Mori, Y.; Ochi, S.; Yoshida, T.; Mori, T.; Iga, J.-I.; Ueno, S.-I. TREM1 mRNA Expression in Leukocytes and Cognitive Function in Japanese Patients with Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 64, 1275–1284. [Google Scholar] [CrossRef]
- Wang, Q.; Xu, Y.; Qi, C.; Liu, A.; Zhao, Y. Association Study of Serum Soluble TREM2 with Vascular Dementia in Chinese Han Population. Int. J. Neurosci. 2020, 130, 708–712. [Google Scholar] [CrossRef]
- Jiang, T.; Gong, P.-Y.; Tan, M.-S.; Xue, X.; Huang, S.; Zhou, J.-S.; Tan, L.; Zhang, Y.-D. Soluble TREM1 Concentrations Are Increased and Positively Correlated with Total Tau Levels in the Plasma of Patients with Alzheimer’s Disease. Aging Clin. Exp. Res. 2019, 31, 1801–1805. [Google Scholar] [CrossRef]
- Liu, D.; Cao, B.; Zhao, Y.; Huang, H.; McIntyre, R.S.; Rosenblat, J.D.; Zhou, H. Soluble TREM2 Changes during the Clinical Course of Alzheimer’s Disease: A Meta-Analysis. Neurosci. Lett. 2018, 686, 10–16. [Google Scholar] [CrossRef]
- Vergallo, A.; Lista, S.; Lemercier, P.; Chiesa, P.A.; Zetterberg, H.; Blennow, K.; Potier, M.-C.; Habert, M.-O.; Baldacci, F.; Cavedo, E.; et al. Association of Plasma YKL-40 with Brain Amyloid-β Levels, Memory Performance, and Sex in Subjective Memory Complainers. Neurobiol. Aging 2020, 96, 22–32. [Google Scholar] [CrossRef]
- Craig-Schapiro, R.; Perrin, R.J.; Roe, C.M.; Xiong, C.; Carter, D.; Cairns, N.J.; Mintun, M.A.; Peskind, E.R.; Li, G.; Galasko, D.R.; et al. YKL-40: A Novel Prognostic Fluid Biomarker for Preclinical Alzheimer’s Disease. Biol. Psychiatry 2010, 68, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Wilczyńska, K.; Maciejczyk, M.; Zalewska, A.; Waszkiewicz, N. Serum Amyloid Biomarkers, Tau Protein and YKL-40 Utility in Detection, Differential Diagnosing, and Monitoring of Dementia. Front. Psychiatry 2021, 12, 725511. [Google Scholar] [CrossRef] [PubMed]
- Wilczyńska, K.; Waszkiewicz, N. Diagnostic Utility of Selected Serum Dementia Biomarkers: Amyloid β-40, Amyloid β-42, Tau Protein, and YKL-40: A Review. J. Clin. Med. 2020, 9, 3452. [Google Scholar] [CrossRef] [PubMed]
- Villar-Piqué, A.; Schmitz, M.; Hermann, P.; Goebel, S.; Bunck, T.; Varges, D.; Ferrer, I.; Riggert, J.; Llorens, F.; Zerr, I. Plasma YKL-40 in the Spectrum of Neurodegenerative Dementia. J. Neuroinflamm. 2019, 16, 145. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Zhao, K.; Xia, H.; Xu, Y. Peripheral Inflammatory Biomarkers in Alzheimer’s Disease and Mild Cognitive Impairment: A Systematic Review and Meta-Analysis. Psychogeriatrics 2019, 19, 300–309. [Google Scholar] [CrossRef]
- Shen, X.-N.; Niu, L.-D.; Wang, Y.-J.; Cao, X.-P.; Liu, Q.; Tan, L.; Zhang, C.; Yu, J.-T. Inflammatory Markers in Alzheimer’s Disease and Mild Cognitive Impairment: A Meta-Analysis and Systematic Review of 170 Studies. J. Neurol. Neurosurg. Psychiatry 2019, 90, 590–598. [Google Scholar] [CrossRef]
- Tao, Q.; Alvin Ang, T.F.; Akhter-Khan, S.C.; Itchapurapu, I.S.; Killiany, R.; Zhang, X.; Budson, A.E.; Turk, K.W.; Goldstein, L.; Mez, J.; et al. Impact of C-Reactive Protein on Cognition and Alzheimer Disease Biomarkers in Homozygous APOE Ɛ4 Carriers. Neurology 2021, 97, e1243–e1252. [Google Scholar] [CrossRef]
- Hao, J.; Qiao, Y.; Li, T.; Yang, J.; Song, Y.; Jia, L.; Jia, J. Investigating Changes in the Serum Inflammatory Factors in Alzheimer’s Disease and Their Correlation with Cognitive Function. J. Alzheimer’s Dis. 2021, 84, 835–842. [Google Scholar] [CrossRef]
- Yarchoan, M.; Louneva, N.; Xie, S.X.; Swenson, F.J.; Hu, W.; Soares, H.; Trojanowski, J.Q.; Lee, V.M.-Y.; Kling, M.A.; Shaw, L.M.; et al. Association of Plasma C-Reactive Protein Levels with the Diagnosis of Alzheimer’s Disease. J. Neurol. Sci. 2013, 333, 9–12. [Google Scholar] [CrossRef][Green Version]
- Song, I.-U.; Chung, S.-W.; Kim, Y.-D.; Maeng, L.-S. Relationship between the Hs-CRP as Non-Specific Biomarker and Alzheimer’s Disease According to Aging Process. Int. J. Med. Sci. 2015, 12, 613–617. [Google Scholar] [CrossRef]
- Balkwill, F.R. The Chemokine System and Cancer. J. Pathol. 2012, 226, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Kulczyńska-Przybik, A.; Słowik, A.; Mroczko, P.; Borawski, B.; Groblewska, M.; Borawska, R.; Mroczko, B. Cerebrospinal Fluid and Blood CX3CL1 as a Potential Biomarker in Early Diagnosis and Prognosis of Dementia. Curr. Alzheimer Res. 2020, 17, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Faura, J.; Bustamante, A.; Penalba, A.; Giralt, D.; Simats, A.; Martínez-Sáez, E.; Alcolea, D.; Fortea, J.; Lleó, A.; Teunissen, C.E.; et al. CCL23: A Chemokine Associated with Progression from Mild Cognitive Impairment to Alzheimer’s Disease. J. Alzheimer’s Dis. 2020, 73, 1585–1595. [Google Scholar] [CrossRef] [PubMed]
- Vacinova, G.; Vejražkova, D.; Rusina, R.; Holmerová, I.; Vaňková, H.; Jarolímová, E.; Včelák, J.; Bendlová, B.; Vaňková, M. Regulated upon Activation, Normal T Cell Expressed and Secreted (RANTES) Levels in the Peripheral Blood of Patients with Alzheimer’s Disease. Neural Regen. Res. 2021, 16, 796. [Google Scholar] [CrossRef]
- Reale, M.; Kamal, M.A.; Velluto, L.; Gambi, D.; di Nicola, M.; Greig, N.H. Relationship between Inflammatory Mediators, Aβ Levels and ApoE Genotype in Alzheimer Disease. Curr. Alzheimer Res. 2012, 9, 447–457. [Google Scholar] [CrossRef]
- Marksteiner, J.; Kemmler, G.; Weiss, E.M.; Knaus, G.; Ullrich, C.; Mechtcheriakov, S.; Oberbauer, H.; Auffinger, S.; Hinterhölzl, J.; Hinterhuber, H.; et al. Five out of 16 Plasma Signaling Proteins Are Enhanced in Plasma of Patients with Mild Cognitive Impairment and Alzheimer’s Disease. Neurobiol. Aging 2011, 32, 539–540. [Google Scholar] [CrossRef]
- Tang, P.; Chong, L.; Li, X.; Liu, Y.; Liu, P.; Hou, C.; Li, R. Correlation between Serum RANTES Levels and the Severity of Parkinson’s Disease. Oxidative Med. Cell. Longev. 2014, 2014, 208408. [Google Scholar] [CrossRef]
- Mori, F.; Nisticò, R.; Nicoletti, C.G.; Zagaglia, S.; Mandolesi, G.; Piccinin, S.; Martino, G.; Finardi, A.; Rossini, P.M.; Marfia, G.A.; et al. RANTES Correlates with Inflammatory Activity and Synaptic Excitability in Multiple Sclerosis. Mult. Scler. J. 2016, 22, 1405–1412. [Google Scholar] [CrossRef]
- Markiewski, M.M.; Lambris, J.D. The Role of Complement in Inflammatory Diseases from Behind the Scenes into the Spotlight. Am. J. Pathol. 2007, 171, 715–727. [Google Scholar] [CrossRef]
- Morgan, A.R.; Touchard, S.; O’Hagan, C.; Sims, R.; Majounie, E.; Escott-Price, V.; Jones, L.; Williams, J.; Morgan, B.P. The Correlation between Inflammatory Biomarkers and Polygenic Risk Score in Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 56, 25–36. [Google Scholar] [CrossRef]
- Hakobyan, S.; Harding, K.; Aiyaz, M.; Hye, A.; Dobson, R.; Baird, A.; Liu, B.; Harris, C.L.; Lovestone, S.; Morgan, B.P. Complement Biomarkers as Predictors of Disease Progression in Alzheimer’s Disease. J. Alzheimer’s Dis. 2016, 54, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Suidan, G.L.; Singh, P.K.; Patel-Hett, S.; Chen, Z.-L.; Volfson, D.; Yamamoto-Imoto, H.; Norris, E.H.; Bell, R.D.; Strickland, S. Abnormal Clotting of the Intrinsic/Contact Pathway in Alzheimer Disease Patients Is Related to Cognitive Ability. Blood Adv. 2018, 2, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Guo, S.; Hibbert, J.M.; Jain, V.; Singh, N.; Wilson, N.O.; Stiles, J.K. CXCL10/IP-10 in Infectious Diseases Pathogenesis and Potential Therapeutic Implications. Cytokine Growth Factor Rev. 2011, 22, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Iturria-Medina, Y.; Sotero, R.C.; Toussaint, P.J.; Mateos-Pérez, J.M.; Evans, A.C. Early Role of Vascular Dysregulation on Late-Onset Alzheimer’s Disease Based on Multifactorial Data-Driven Analysis. Nat. Commun. 2016, 7, 11934. [Google Scholar] [CrossRef]
- Galimberti, D.; Venturelli, E.; Fenoglio, C.; Lovati, C.; Guidi, I.; Scalabrini, D.; Mariani, C.; Bresolin, N.; Scarpini, E. IP-10 Serum Levels Are Not Increased in Mild Cognitive Impairment and Alzheimer’s Disease. Eur. J. Neurol. 2007, 14, e3–e4. [Google Scholar] [CrossRef]
- Galimberti, D.; Schoonenboom, N.; Scheltens, P.; Fenoglio, C.; Bouwman, F.; Venturelli, E.; Guidi, I.; Blankenstein, M.A.; Bresolin, N.; Scarpini, E. Intrathecal Chemokine Synthesis in Mild Cognitive Impairment and Alzheimer Disease. Arch. Neurol. 2006, 63, 538. [Google Scholar] [CrossRef]
- Popp, J.; Oikonomidi, A.; Tautvydaitė, D.; Dayon, L.; Bacher, M.; Migliavacca, E.; Henry, H.; Kirkland, R.; Severin, I.; Wojcik, J.; et al. Markers of Neuroinflammation Associated with Alzheimer’s Disease Pathology in Older Adults. Brain Behav. Immun. 2017, 62, 203–211. [Google Scholar] [CrossRef]
- Delaby, C.; Gabelle, A.; Blum, D.; Schraen-Maschke, S.; Moulinier, A.; Boulanghien, J.; Séverac, D.; Buée, L.; Rème, T.; Lehmann, S. Central Nervous System and Peripheral Inflammatory Processes in Alzheimer’s Disease: Biomarker Profiling Approach. Front. Neurol. 2015, 6, 181. [Google Scholar] [CrossRef]
- Abe, K.; Shang, J.; Shi, X.; Yamashita, T.; Hishikawa, N.; Takemoto, M.; Morihara, R.; Nakano, Y.; Ohta, Y.; Deguchi, K.; et al. A New Serum Biomarker Set to Detect Mild Cognitive Impairment and Alzheimer’s Disease by Peptidome Technology. J. Alzheimer’s Dis. 2020, 73, 217–227. [Google Scholar] [CrossRef]
- Cacabelos, R.; Alvarez, X.A.; Fernández-Novoa, L.; Franco, A.; Mangues, R.; Pellicer, A.; Nishimura, T. Brain Interleukin-1 Beta in Alzheimer’s Disease and Vascular Dementia. Methods Find. Exp. Clin. Pharmacol. 1994, 16, 141–151. [Google Scholar]
- Bradburn, S.; Sarginson, J.; Murgatroyd, C.A. Association of Peripheral Interleukin-6 with Global Cognitive Decline in Non-Demented Adults: A Meta-Analysis of Prospective Studies. Front. Aging Neurosci. 2018, 9, 438. [Google Scholar] [CrossRef] [PubMed]
- la Rosa, F.; Saresella, M.; Baglio, F.; Piancone, F.; Marventano, I.; Calabrese, E.; Nemni, R.; Ripamonti, E.; Cabinio, M.; Clerici, M. Immune and Imaging Correlates of Mild Cognitive Impairment Conversion to Alzheimer’s Disease. Sci. Rep. 2017, 7, 16760. [Google Scholar] [CrossRef] [PubMed]
- Swardfager, W.; Lanctôt, K.; Rothenburg, L.; Wong, A.; Cappell, J.; Herrmann, N. A Meta-Analysis of Cytokines in Alzheimer’s Disease. Biol. Psychiatry 2010, 68, 930–941. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.S.P.; Liu, C.S.; Rau, A.; Lanctôt, K.L.; Köhler, C.A.; Pakosh, M.; Carvalho, A.F.; Herrmann, N. Peripheral Inflammatory Markers in Alzheimer’s Disease: A Systematic Review and Meta-Analysis of 175 Studies. J. Neurol. Neurosurg. Psychiatry 2017, 88, 876–882. [Google Scholar] [CrossRef] [PubMed]
- Ng, A.; Tam, W.W.; Zhang, M.W.; Ho, C.S.; Husain, S.F.; McIntyre, R.S.; Ho, R.C. IL-1β, IL-6, TNF- α and CRP in Elderly Patients with Depression or Alzheimer’s Disease: Systematic Review and Meta-Analysis. Sci. Rep. 2018, 8, 12050. [Google Scholar] [CrossRef]
- Neshan, M.; Malakouti, S.K.; Kamalzadeh, L.; Makvand, M.; Campbell, A.; Ahangari, G. Alterations in T-Cell Transcription Factors and Cytokine Gene Expression in Late-Onset Alzheimer’s Disease. J. Alzheimer’s Dis. 2022, 85, 645–665. [Google Scholar] [CrossRef]
- Liang, C.-S.; Su, K.-P.; Tsai, C.-L.; Lee, J.-T.; Chu, C.-S.; Yeh, T.-C.; Su, M.-W.; Lin, G.-Y.; Lin, Y.-K.; Chu, H.-T.; et al. The Role of Interleukin-33 in Patients with Mild Cognitive Impairment and Alzheimer’s Disease. Alzheimer’s Res. Ther. 2020, 12, 86. [Google Scholar] [CrossRef]
- Sun, Y.; Yin, X.S.; Guo, H.; Han, R.K.; He, R.D.; Chi, L.J. Elevated Osteopontin Levels in Mild Cognitive Impairment and Alzheimer’s Disease. Mediat. Inflamm. 2013, 2013, 615745. [Google Scholar] [CrossRef]
- Liang, C.-S.; Tsai, C.-L.; Lin, G.-Y.; Lee, J.-T.; Lin, Y.-K.; Chu, C.-S.; Sung, Y.-F.; Tsai, C.-K.; Yeh, T.-C.; Chu, H.-T.; et al. Better Identification of Cognitive Decline with Interleukin-2 Than with Amyloid and Tau Protein Biomarkers in Amnestic Mild Cognitive Impairment. Front. Aging Neurosci. 2021, 13, 670115. [Google Scholar] [CrossRef]
- Mamas, M.; Dunn, W.B.; Neyses, L.; Goodacre, R. The Role of Metabolites and Metabolomics in Clinically Applicable Biomarkers of Disease. Arch. Toxicol. 2011, 85, 5–17. [Google Scholar] [CrossRef]
- de la Monte, S.M.; Tong, M. Brain Metabolic Dysfunction at the Core of Alzheimer’s Disease. Biochem. Pharmacol. 2014, 88, 548–559. [Google Scholar] [CrossRef] [PubMed]
- Hassan-Smith, G.; Wallace, G.R.; Douglas, M.R.; Sinclair, A.J. The Role of Metabolomics in Neurological Disease. J. Neuroimmunol. 2012, 248, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Y.; Sun, Y.-P.; Luo, Y.-M.; Peng, D.-H.; Li, X.; Yang, B.-Y.; Wang, Q.-H.; Kuang, H.-X. Biomarkers for the Clinical Diagnosis of Alzheimer’s Disease: Metabolomics Analysis of Brain Tissue and Blood. Front. Pharmacol. 2021, 12, 700587. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.J. Early-Stage and Preclinical Alzheimer Disease. Alzheimer Dis. Assoc. Disord. 2005, 19, 163–165. [Google Scholar] [CrossRef] [PubMed]
- Bateman, R.J.; Xiong, C.; Benzinger, T.L.S.; Fagan, A.M.; Goate, A.; Fox, N.C.; Marcus, D.S.; Cairns, N.J.; Xie, X.; Blazey, T.M.; et al. Clinical and Biomarker Changes in Dominantly Inherited Alzheimer’s Disease. N. Engl. J. Med. 2012, 367, 795–804. [Google Scholar] [CrossRef]
- Hane, F.T.; Robinson, M.; Lee, B.Y.; Bai, O.; Leonenko, Z.; Albert, M.S. Recent Progress in Alzheimer’s Disease Research, Part 3: Diagnosis and Treatment. J. Alzheimer’s Dis. 2017, 57, 645–665. [Google Scholar] [CrossRef]
- Yang, J.; Chen, T.; Sun, L.; Zhao, Z.; Qi, X.; Zhou, K.; Cao, Y.; Wang, X.; Qiu, Y.; Su, M.; et al. Potential Metabolite Markers of Schizophrenia. Mol. Psychiatry 2013, 18, 67–78. [Google Scholar] [CrossRef]
- Sun, C.; Gao, M.; Wang, F.; Yun, Y.; Sun, Q.; Guo, R.; Yan, C.; Sun, X.; Li, Y. Serum Metabolomic Profiling in Patients with Alzheimer Disease and Amnestic Mild Cognitive Impairment by GC/MS. Biomed. Chromatogr. 2020, 34, e4875. [Google Scholar] [CrossRef]
- Piubelli, L.; Pollegioni, L.; Rabattoni, V.; Mauri, M.; Princiotta Cariddi, L.; Versino, M.; Sacchi, S. Serum D-Serine Levels Are Altered in Early Phases of Alzheimer’s Disease: Towards a Precocious Biomarker. Transl. Psychiatry 2021, 11, 77. [Google Scholar] [CrossRef]
- Hartley, D.; Blumenthal, T.; Carrillo, M.; DiPaolo, G.; Esralew, L.; Gardiner, K.; Granholm, A.-C.; Iqbal, K.; Krams, M.; Lemere, C.; et al. Down Syndrome and Alzheimer’s Disease: Common Pathways, Common Goals. Alzheimer’s Dement. 2015, 11, 700–709. [Google Scholar] [CrossRef]
- Fortea, J.; Vilaplana, E.; Carmona-Iragui, M.; Benejam, B.; Videla, L.; Barroeta, I.; Fernández, S.; Altuna, M.; Pegueroles, J.; Montal, V.; et al. Clinical and Biomarker Changes of Alzheimer’s Disease in Adults with Down Syndrome: A Cross-Sectional Study. Lancet 2020, 395, 1988–1997. [Google Scholar] [CrossRef]
- Gross, T.J.; Doran, E.; Cheema, A.K.; Head, E.; Lott, I.T.; Mapstone, M. Plasma Metabolites Related to Cellular Energy Metabolism Are Altered in Adults with Down Syndrome and Alzheimer’s Disease. Dev. Neurobiol. 2019, 79, 622–638. [Google Scholar] [CrossRef] [PubMed]
- Gomez, W.; Morales, R.; Maracaja-Coutinho, V.; Parra, V.; Nassif, M. Down Syndrome and Alzheimer’s Disease: Common Molecular Traits beyond the Amyloid Precursor Protein. Aging 2020, 12, 1011–1033. [Google Scholar] [CrossRef] [PubMed]
- Mosconi, L.; de Santi, S.; Li, J.; Tsui, W.H.; Li, Y.; Boppana, M.; Laska, E.; Rusinek, H.; de Leon, M.J. Hippocampal Hypometabolism Predicts Cognitive Decline from Normal Aging. Neurobiol. Aging 2008, 29, 676–692. [Google Scholar] [CrossRef]
- Baik, S.H.; Kang, S.; Lee, W.; Choi, H.; Chung, S.; Kim, J.-I.; Mook-Jung, I. A Breakdown in Metabolic Reprogramming Causes Microglia Dysfunction in Alzheimer’s Disease. Cell Metab. 2019, 30, 493–507.e6. [Google Scholar] [CrossRef]
- Yao, J.; Rettberg, J.R.; Klosinski, L.P.; Cadenas, E.; Brinton, R.D. Shift in Brain Metabolism in Late Onset Alzheimer’s Disease: Implications for Biomarkers and Therapeutic Interventions. Mol. Asp. Med. 2011, 32, 247–257. [Google Scholar] [CrossRef]
- Vlassenko, A.G.; Gordon, B.A.; Goyal, M.S.; Su, Y.; Blazey, T.M.; Durbin, T.J.; Couture, L.E.; Christensen, J.J.; Jafri, H.; Morris, J.C.; et al. Aerobic Glycolysis and Tau Deposition in Preclinical Alzheimer’s Disease. Neurobiol. Aging 2018, 67, 95–98. [Google Scholar] [CrossRef]
- Marksteiner, J.; Blasko, I.; Kemmler, G.; Koal, T.; Humpel, C. Bile Acid Quantification of 20 Plasma Metabolites Identifies Lithocholic Acid as a Putative Biomarker in Alzheimer’s Disease. Metabolomics 2018, 14, 1. [Google Scholar] [CrossRef]
- Voyle, N.; Kim, M.; Proitsi, P.; Ashton, N.J.; Baird, A.L.; Bazenet, C.; Hye, A.; Westwood, S.; Chung, R.; Ward, M.; et al. Blood Metabolite Markers of Neocortical Amyloid-β Burden: Discovery and Enrichment Using Candidate Proteins. Transl. Psychiatry 2016, 6, e719. [Google Scholar] [CrossRef]
- Aizenstein, H.J.; Nebes, R.D.; Saxton, J.A.; Price, J.C.; Mathis, C.A.; Tsopelas, N.D.; Ziolko, S.K.; James, J.A.; Snitz, B.E.; Houck, P.R.; et al. Frequent Amyloid Deposition Without Significant Cognitive Impairment Among the Elderly. Arch. Neurol. 2008, 65, 1509. [Google Scholar] [CrossRef]
- Zolochevska, O.; Taglialatela, G. Non-Demented Individuals with Alzheimer’s Disease Neuropathology: Resistance to Cognitive Decline May Reveal New Treatment Strategies. Curr. Pharm. Des. 2016, 22, 4063–4068. [Google Scholar] [CrossRef] [PubMed]
- Giannakopoulos, P.; Herrmann, F.R.; Bussiere, T.; Bouras, C.; Kovari, E.; Perl, D.P.; Morrison, J.H.; Gold, G.; Hof, P.R. Tangle and Neuron Numbers, but Not Amyloid Load, Predict Cognitive Status in Alzheimer’s Disease. Neurology 2003, 60, 1495–1500. [Google Scholar] [CrossRef] [PubMed]
- Kamogawa, K.; Kohara, K.; Tabara, Y.; Takita, R.; Miki, T.; Konno, T.; Hata, S.; Suzuki, T. Potential Utility of Soluble P3-Alcadeinα Plasma Levels as a Biomarker for Sporadic Alzheimer’s Disease. J. Alzheimer’s Dis. 2012, 31, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Piao, Y.; Kimura, A.; Urano, S.; Saito, Y.; Taru, H.; Yamamoto, T.; Hata, S.; Suzuki, T. Mechanism of Intramembrane Cleavage of Alcadeins by γ-Secretase. PLoS ONE 2013, 8, e62431. [Google Scholar] [CrossRef]
- Jones, L.L.; McDonald, D.A.; Borum, P.R. Acylcarnitines: Role in Brain. Prog. Lipid Res. 2010, 49, 61–75. [Google Scholar] [CrossRef] [PubMed]
- McCoin, C.S.; Knotts, T.A.; Adams, S.H. Acylcarnitines—Old Actors Auditioning for New Roles in Metabolic Physiology. Nat. Rev. Endocrinol. 2015, 11, 617–625. [Google Scholar] [CrossRef]
- Mihalik, S.J.; Goodpaster, B.H.; Kelley, D.E.; Chace, D.H.; Vockley, J.; Toledo, F.G.S.; DeLany, J.P. Increased Levels of Plasma Acylcarnitines in Obesity and Type 2 Diabetes and Identification of a Marker of Glucolipotoxicity. Obesity 2010, 18, 1695–1700. [Google Scholar] [CrossRef]
- Schooneman, M.G.; Vaz, F.M.; Houten, S.M.; Soeters, M.R. Acylcarnitines: Reflecting or Inflicting Insulin Resistance? Diabetes 2013, 62, 1–8. [Google Scholar] [CrossRef]
- Huo, Z.; Yu, L.; Yang, J.; Zhu, Y.; Bennett, D.A.; Zhao, J. Brain and Blood Metabolome for Alzheimer’s Dementia: Findings from a Targeted Metabolomics Analysis. Neurobiol. Aging 2020, 86, 123–133. [Google Scholar] [CrossRef]
- Harrigan, G.; Goodacre, R. Metabolic Profiling: Its Role in Biomarker Discovery and Gene Function Analysis; Harrigan, G.G., Goodacre, R., Eds.; Springer: Boston, MA, USA, 2003; ISBN 978-1-4613-5025-5. [Google Scholar]
- Wang, G.; Zhou, Y.; Huang, F.-J.; Tang, H.-D.; Xu, X.-H.; Liu, J.-J.; Wang, Y.; Deng, Y.-L.; Ren, R.-J.; Xu, W.; et al. Plasma Metabolite Profiles of Alzheimer’s Disease and Mild Cognitive Impairment. J. Proteome Res. 2014, 13, 2649–2658. [Google Scholar] [CrossRef]
- Varma, V.R.; Oommen, A.M.; Varma, S.; Casanova, R.; An, Y.; Andrews, R.M.; O’Brien, R.; Pletnikova, O.; Troncoso, J.C.; Toledo, J.; et al. Brain and Blood Metabolite Signatures of Pathology and Progression in Alzheimer Disease: A Targeted Metabolomics Study. PLoS Med. 2018, 15, e1002482. [Google Scholar] [CrossRef] [PubMed]
- Xicota, L.; Ichou, F.; Lejeune, F.-X.; Colsch, B.; Tenenhaus, A.; Leroy, I.; Fontaine, G.; Lhomme, M.; Bertin, H.; Habert, M.-O.; et al. Multi-Omics Signature of Brain Amyloid Deposition in Asymptomatic Individuals at-Risk for Alzheimer’s Disease: The INSIGHT-PreAD Study. EBioMedicine 2019, 47, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Teunissen, C.E.; Pool, R.; Shipley, M.J.; Teumer, A.; Chouraki, V.; Melo van Lent, D.; Tynkkynen, J.; Fischer, K.; Hernesniemi, J.; et al. Circulating Metabolites and General Cognitive Ability and Dementia: Evidence from 11 Cohort Studies. Alzheimer’s Dement. 2018, 14, 707–722. [Google Scholar] [CrossRef] [PubMed]
- Gella, A.; Durany, N. Oxidative Stress in Alzheimer Disease. Cell Adhes. Migr. 2009, 3, 88–93. [Google Scholar] [CrossRef]
- Huang, W.-J.; Zhang, X.; Chen, W.-W. Role of Oxidative Stress in Alzheimer’s Disease. Biomed. Rep. 2016, 4, 519–522. [Google Scholar] [CrossRef]
- Praticò, D.; Uryu, K.; Leight, S.; Trojanoswki, J.Q.; Lee, V.M.-Y. Increased Lipid Peroxidation Precedes Amyloid Plaque Formation in an Animal Model of Alzheimer Amyloidosis. J. Neurosci. 2001, 21, 4183–4187. [Google Scholar] [CrossRef]
- Markesbery, W.R. Oxidative Stress Hypothesis in Alzheimer’s Disease. Free Radic. Biol. Med. 1997, 23, 134–147. [Google Scholar] [CrossRef]
- Smith, M.A.; Rottkamp, C.A.; Nunomura, A.; Raina, A.K.; Perry, G. Oxidative Stress in Alzheimer’s Disease. Biochim. Biophys. Acta-Mol. Basis Dis. 2000, 1502, 139–144. [Google Scholar] [CrossRef]
- Buccellato, F.R.; D’Anca, M.; Fenoglio, C.; Scarpini, E.; Galimberti, D. Role of Oxidative Damage in Alzheimer’s Disease and Neurodegeneration: From Pathogenic Mechanisms to Biomarker Discovery. Antioxidants 2021, 10, 1353. [Google Scholar] [CrossRef]
- Martins, R.N.; Villemagne, V.; Sohrabi, H.R.; Chatterjee, P.; Shah, T.M.; Verdile, G.; Fraser, P.; Taddei, K.; Gupta, V.B.; Rainey-Smith, S.R.; et al. Alzheimer’s Disease: A Journey from Amyloid Peptides and Oxidative Stress, to Biomarker Technologies and Disease Prevention Strategies—Gains from AIBL and DIAN Cohort Studies. J. Alzheimer’s Dis. 2018, 62, 965–992. [Google Scholar] [CrossRef]
- Lin, C.-H.; Lane, H.-Y. Plasma Glutathione Levels Decreased with Cognitive Decline among People with Mild Cognitive Impairment (MCI): A Two-Year Prospective Study. Antioxidants 2021, 10, 1839. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Wang, F.; Chen, J.; Li, X.; Fu, G.; Zhou, J.; Zhou, D.; Wu, W.; Chen, H. Changes in Biothiol Levels Are Closely Associated with Alzheimer’s Disease. J. Alzheimer’s Dis. 2021, 82, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Evlice, A.; Ulusu, N.N. Glucose-6-Phosphate Dehydrogenase a Novel Hope on a Blood-Based Diagnosis of Alzheimer’s Disease. Acta Neurol. Belg. 2017, 117, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Peña-Bautista, C.; Álvarez-Sánchez, L.; Ferrer, I.; López-Nogueroles, M.; Cañada-Martínez, A.J.; Oger, C.; Galano, J.-M.; Durand, T.; Baquero, M.; Cháfer-Pericás, C. Lipid Peroxidation Assessment in Preclinical Alzheimer Disease Diagnosis. Antioxidants 2021, 10, 1043. [Google Scholar] [CrossRef]
- Peña-Bautista, C.; Álvarez, L.; Baquero, M.; Ferrer, I.; García, L.; Hervás-Marín, D.; Cháfer-Pericás, C. Plasma Isoprostanoids Assessment as Alzheimer’s Disease Progression Biomarkers. J. Neurochem. 2021, 157, 2187–2194. [Google Scholar] [CrossRef]
- Zengi, O.; Karakas, A.; Ergun, U.; Senes, M.; Inan, L.; Yucel, D. Urinary 8-Hydroxy-2′-Deoxyguanosine Level and Plasma Paraoxonase 1 Activity with Alzheimer’s Disease. Clin. Chem. Lab. Med. 2012, 50, 529–534. [Google Scholar] [CrossRef]
- Picco, A.; Polidori, M.C.; Ferrara, M.; Cecchetti, R.; Arnaldi, D.; Baglioni, M.; Morbelli, S.; Bastiani, P.; Bossert, I.; Fiorucci, G.; et al. Plasma Antioxidants and Brain Glucose Metabolism in Elderly Subjects with Cognitive Complaints. Eur. J. Nucl. Med. Mol. Imaging 2014, 41, 764–775. [Google Scholar] [CrossRef]
- Rita Cardoso, B.; Silva Bandeira, V.; Jacob-Filho, W.; Franciscato Cozzolino, S.M. Selenium Status in Elderly: Relation to Cognitive Decline. J. Trace Elem. Med. Biol. 2014, 28, 422–426. [Google Scholar] [CrossRef]
- Wu, L.-Y.; Cheah, I.K.; Chong, J.R.; Chai, Y.L.; Tan, J.Y.; Hilal, S.; Vrooman, H.; Chen, C.P.; Halliwell, B.; Lai, M.K.P. Low Plasma Ergothioneine Levels Are Associated with Neurodegeneration and Cerebrovascular Disease in Dementia. Free Radic. Biol. Med. 2021, 177, 201–211. [Google Scholar] [CrossRef]
- Du, L.; Ma, J.; He, D.; Zhang, X. Serum Ischaemia-modified Albumin Might Be a Potential Biomarker for Oxidative Stress in Amnestic Mild Cognitive Impairment. Psychogeriatrics 2019, 19, 150–156. [Google Scholar] [CrossRef]
- Li, L.-L.; Ma, Y.-H.; Bi, Y.-L.; Sun, F.-R.; Hu, H.; Hou, X.-H.; Xu, W.; Shen, X.-N.; Dong, Q.; Tan, L.; et al. Serum Uric Acid May Aggravate Alzheimer’s Disease Risk by Affecting Amyloidosis in Cognitively Intact Older Adults: The CABLE Study. J. Alzheimer’s Dis. 2021, 81, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Mcintyre, J.A.; Wagenknecht, D.R.; Ramsey, C.J. Redox-Reactive Antiphospholipid Antibody Differences between Serum from Alzheimer’s Patients and Age-Matched Controls. Autoimmunity 2009, 42, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, R.; Singh, A.K.; Kumar, P.; Bajpai, S.; Pathak, M.; Chatterjee, P.; Dwivedi, S.; Dey, A.B.; Dey, S. Blood Circulatory Level of Seven Sirtuins in Alzheimer’s Disease: Potent Biomarker Based on Translational Research. Mol. Neurobiol. 2022, 59, 1440–1451. [Google Scholar] [CrossRef]
- López, N.; Tormo, C.; de Blas, I.; Llinares, I.; Alom, J. Oxidative Stress in Alzheimer’s Disease and Mild Cognitive Impairment with High Sensitivity and Specificity. J. Alzheimer’s Dis. 2013, 33, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.M.; Wilson-Delfosse, A.L.; Mieyal, J.J. Dysregulation of Glutathione Homeostasis in Neurodegenerative Diseases. Nutrients 2012, 4, 1399–1440. [Google Scholar] [CrossRef] [PubMed]
- Haddad, M.; Hervé, V.; ben Khedher, M.R.; Rabanel, J.-M.; Ramassamy, C. Glutathione: An Old and Small Molecule with Great Functions and New Applications in the Brain and in Alzheimer’s Disease. Antioxid. Redox Signal. 2021, 35, 270–292. [Google Scholar] [CrossRef] [PubMed]
- Zheng, T.; Liu, H.; Qin, L.; Chen, B.; Zhang, X.; Hu, X.; Xiao, L.; Qin, S. Oxidative Stress-Mediated Influence of Plasma DPP4 Activity to BDNF Ratio on Mild Cognitive Impairment in Elderly Type 2 Diabetic Patients: Results from the GDMD Study in China. Metabolism 2018, 87, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Zheng, T.; Qin, L.; Hu, X.; Zhang, X.; Liu, Y.; Liu, H.; Qin, S.; Li, G.; Li, Q. Strong Association between Plasma Dipeptidyl Peptidase-4 Activity and Impaired Cognitive Function in Elderly Population with Normal Glucose Tolerance. Front. Aging Neurosci. 2017, 9, 247. [Google Scholar] [CrossRef]
- Signorini, C.; de Felice, C.; Galano, J.-M.; Oger, C.; Leoncini, S.; Cortelazzo, A.; Ciccoli, L.; Durand, T.; Hayek, J.; Lee, J. Isoprostanoids in Clinical and Experimental Neurological Disease Models. Antioxidants 2018, 7, 88. [Google Scholar] [CrossRef]
- Mufson, E.J.; Leurgans, S. Inability of Plasma and Urine F2A-Isoprostane Levels to Differentiate Mild Cognitive Impairment from Alzheimer’s Disease. Neurodegener. Dis. 2010, 7, 139–142. [Google Scholar] [CrossRef]
- Evans, J.R.; Lawrenson, J.G. Antioxidant Vitamin and Mineral Supplements for Preventing Age-Related Macular Degeneration. Cochrane Database Syst. Rev. 2017, 2017, CD000253. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, B.R.; Hare, D.J.; Bush, A.I.; Li, Q.-X.; Fowler, C.J.; Masters, C.L.; Martins, R.N.; Ganio, K.; Lothian, A.; Mukherjee, S.; et al. Selenium Levels in Serum, Red Blood Cells, and Cerebrospinal Fluid of Alzheimer’s Disease Patients: A Report from the Australian Imaging, Biomarker & Lifestyle Flagship Study of Ageing (AIBL). J. Alzheimer’s Dis. 2017, 57, 183–193. [Google Scholar] [CrossRef]
- Ficiarà, E.; Munir, Z.; Boschi, S.; Caligiuri, M.E.; Guiot, C. Alteration of Iron Concentration in Alzheimer’s Disease as a Possible Diagnostic Biomarker Unveiling Ferroptosis. Int. J. Mol. Sci. 2021, 22, 4479. [Google Scholar] [CrossRef] [PubMed]
- Arnal, N.; Morel, G.R.; de Alaniz, M.J.T.; Castillo, O.; Marra, C.A. Role of Copper and Cholesterol Association in the Neurodegenerative Process. Int. J. Alzheimer’s Dis. 2013, 2013, 414817. [Google Scholar] [CrossRef]
- Mueller, C.; Schrag, M.; Crofton, A.; Stolte, J.; Muckenthaler, M.U.; Magaki, S.; Kirsch, W. Altered Serum Iron and Copper Homeostasis Predicts Cognitive Decline in Mild Cognitive Impairment. J. Alzheimer’s Dis. 2012, 29, 341–350. [Google Scholar] [CrossRef]
- de Leeuw, F.A.; van der Flier, W.M.; Tijms, B.M.; Scheltens, P.; Mendes, V.M.; Manadas, B.; Bierau, J.; van Wijk, N.; van den Heuvel, E.G.H.M.; Mohajeri, M.H.; et al. Specific Nutritional Biomarker Profiles in Mild Cognitive Impairment and Subjective Cognitive Decline Are Associated with Clinical Progression: The NUDAD Project. J. Am. Med. Dir. Assoc. 2020, 21, 1513.e1–1513.e17. [Google Scholar] [CrossRef]
- Gündüztepe, Y.; Bukan, N.; Zorlu, E.; Karaman, Y.; Andaç Topkan, T.; Gurbuz, N.; Neşelioğlu, S.; Erel, Ö. The Evaluation of Thiol-Disulfıte Balance, Ischemıa Albumın Modıfıcation and Seruloplazmine as a New Oxidatıve Stress in Mild Cognitive Impairment and Early Stage Alzheimer’s Disease Patients. J. Clin. Neurosci. 2020, 75, 188–194. [Google Scholar] [CrossRef]
- Yadav, J.; Verma, A.K.; Garg, R.K.; Ahmad, K.; Shiuli; Mahdi, A.A.; Srivastava, S. Sialic Acid Associated with Oxidative Stress and Total Antioxidant Capacity (TAC) Expression Level as a Predictive Indicator in Moderate to Severe Alzheimer’s Disease. Exp. Gerontol. 2020, 141, 111092. [Google Scholar] [CrossRef]
- Işık, M.; Beydemir, Ş. AChE MRNA Expression as a Possible Novel Biomarker for the Diagnosis of Coronary Artery Disease and Alzheimer’s Disease, and Its Association with Oxidative Stress. Arch. Physiol. Biochem. 2019, 128, 352–359. [Google Scholar] [CrossRef]
- Engel, B.; Gomm, W.; Broich, K.; Maier, W.; Weckbecker, K.; Haenisch, B. Hyperuricemia and Dementia—A Case-Control Study. BMC Neurol. 2018, 18, 131. [Google Scholar] [CrossRef]
- Yamagishi, K.; Ikeda, A.; Moriyama, Y.; Chei, C.-L.; Noda, H.; Umesawa, M.; Cui, R.; Nagao, M.; Kitamura, A.; Yamamoto, Y.; et al. Serum Coenzyme Q10 and Risk of Disabling Dementia: The Circulatory Risk in Communities Study (CIRCS). Atherosclerosis 2014, 237, 400–403. [Google Scholar] [CrossRef] [PubMed]
- de Bustos, F.; Molina, J.A.; Jiménez-Jiménez, F.J.; García-Redondo, A.; Gómez-Escalonilla, C.; Porta-Etessam, J.; Berbel, A.; Zurdo, M.; Barcenilla, B.; Parrilla, G.; et al. Serum Levels of Coenzyme Q 10 in Patients with Alzheimer’s Disease. J. Neural Transm. 2000, 107, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, A.; Lopez, D.; Khoo, P.; Kalaria, R.N.; Mukaetova-Ladinska, E.B. Exploring Erythrocytes as Blood Biomarkers for Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 60, 845–857. [Google Scholar] [CrossRef]
- Cheng, L.; Quek, C.Y.J.; Sun, X.; Bellingham, S.A.; Hill, A.F. The Detection of MicroRNA Associated with Alzheimer’s Disease in Biological Fluids Using next-Generation Sequencing Technologies. Front. Genet. 2013, 4, 150. [Google Scholar] [CrossRef] [PubMed]
- Fabian, M.R.; Sonenberg, N. The Mechanics of MiRNA-Mediated Gene Silencing: A Look under the Hood of MiRISC. Nat. Struct. Mol. Biol. 2012, 19, 586–593. [Google Scholar] [CrossRef]
- Krol, J.; Loedige, I.; Filipowicz, W. The Widespread Regulation of MicroRNA Biogenesis, Function and Decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Kumar, S.; Reddy, P.H. Are Circulating MicroRNAs Peripheral Biomarkers for Alzheimer’s Disease? Biochim. Biophys. Acta-Mol. Basis Dis. 2016, 1862, 1617–1627. [Google Scholar] [CrossRef]
- Kumar, S.; Vijayan, M.; Bhatti, J.S.; Reddy, P.H. MicroRNAs as Peripheral Biomarkers in Aging and Age-Related Diseases. Prog. Mol. Biol. Transl. Sci. 2017, 146, 47–94. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, Z.; Le, W. Tiny but Mighty: Promising Roles of MicroRNAs in the Diagnosis and Treatment of Parkinson’s Disease. Neurosci. Bull. 2017, 33, 543–551. [Google Scholar] [CrossRef]
- Kume, K.; Iwama, H.; Deguchi, K.; Ikeda, K.; Takata, T.; Kokudo, Y.; Kamada, M.; Fujikawa, K.; Hirose, K.; Masugata, H.; et al. Serum MicroRNA Expression Profiling in Patients with Multiple System Atrophy. Mol. Med. Rep. 2017, 17, 852–860. [Google Scholar] [CrossRef]
- Patil, K.S.; Basak, I.; Dalen, I.; Hoedt, E.; Lange, J.; Lunde, K.A.; Liu, Y.; Tysnes, O.-B.; Forsgren, L.; Aarsland, D.; et al. Combinatory MicroRNA Serum Signatures as Classifiers of Parkinson’s Disease. Parkinsonism Relat. Disord. 2019, 64, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Regev, K.; Paul, A.; Healy, B.; von Glenn, F.; Diaz-Cruz, C.; Gholipour, T.; Mazzola, M.A.; Raheja, R.; Nejad, P.; Glanz, B.I.; et al. Comprehensive Evaluation of Serum MicroRNAs as Biomarkers in Multiple Sclerosis. Neurol.-Neuroimmunol. Neuroinflammation 2016, 3, e267. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Jäkel, L.; Bruinsma, I.B.; Claassen, J.A.; Kuiperij, H.B.; Verbeek, M.M. MicroRNA-29a Is a Candidate Biomarker for Alzheimer’s Disease in Cell-Free Cerebrospinal Fluid. Mol. Neurobiol. 2016, 53, 2894–2899. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Fan, G.; Zhang, J.; Wu, C.; Du, Y.; Ye, H.; Li, Z.; Wang, L.; Zhang, Z.; Zhang, L.; et al. A 9-MicroRNA Signature in Serum Serves as a Noninvasive Biomarker in Early Diagnosis of Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 60, 1365–1377. [Google Scholar] [CrossRef]
- Takousis, P.; Sadlon, A.; Schulz, J.; Wohlers, I.; Dobricic, V.; Middleton, L.; Lill, C.M.; Perneczky, R.; Bertram, L. Differential Expression of MicroRNAs in Alzheimer’s Disease Brain, Blood, and Cerebrospinal Fluid. Alzheimer’s Dement. 2019, 15, 1468–1477. [Google Scholar] [CrossRef]
- Sheinerman, K.S.; Umansky, S.R. Circulating Cell-Free MicroRNA as Biomarkers for Screening, Diagnosis and Monitoring of Neurodegenerative Diseases and Other Neurologic Pathologies. Front. Cell. Neurosci. 2013, 7, 150. [Google Scholar] [CrossRef]
- Nguyen, L.D.; Chau, R.K.; Krichevsky, A.M. Small Molecule Drugs Targeting Non-Coding RNAs as Treatments for Alzheimer’s Disease and Related Dementias. Genes 2021, 12, 2005. [Google Scholar] [CrossRef]
- Kumar, P.; Dezso, Z.; MacKenzie, C.; Oestreicher, J.; Agoulnik, S.; Byrne, M.; Bernier, F.; Yanagimachi, M.; Aoshima, K.; Oda, Y. Circulating MiRNA Biomarkers for Alzheimer’s Disease. PLoS ONE 2013, 8, e69807. [Google Scholar] [CrossRef]
- Kiko, T.; Nakagawa, K.; Tsuduki, T.; Furukawa, K.; Arai, H.; Miyazawa, T. MicroRNAs in Plasma and Cerebrospinal Fluid as Potential Markers for Alzheimer’s Disease. J. Alzheimer’s Dis. 2014, 39, 253–259. [Google Scholar] [CrossRef]
- Sørensen, S.S.; Nygaard, A.-B.; Christensen, T. MiRNA Expression Profiles in Cerebrospinal Fluid and Blood of Patients with Alzheimer’s Disease and Other Types of Dementia—An Exploratory Study. Transl. Neurodegener. 2016, 5, 6. [Google Scholar] [CrossRef]
- Poursaei, E.; Abolghasemi, M.; Bornehdeli, S.; Shanehbandi, D.; Asadi, M.; Sadeghzadeh, M.; Rahmanpour, D.; Sadeh, R.N. Evaluation of Hsa-Let-7d-5p, Hsa-Let-7g-5p and Hsa-MiR-15b-5p Plasma Levels in Patients with Alzheimer’s Disease. Psychiatr. Genet. 2022, 32, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Schipper, H.M.; Maes, O.C.; Chertkow, H.M.; Wang, E. MicroRNA Expression in Alzheimer Blood Mononuclear Cells. Gene Regul. Syst. Biol. 2007, 1, GRSB.S361. [Google Scholar] [CrossRef] [PubMed]
- Villa, C.; Ridolfi, E.; Fenoglio, C.; Ghezzi, L.; Vimercati, R.; Clerici, F.; Marcone, A.; Gallone, S.; Serpente, M.; Cantoni, C.; et al. Expression of the Transcription Factor Sp1 and Its Regulatory Hsa-MiR-29b in Peripheral Blood Mononuclear Cells from Patients with Alzheimer’s Disease. J. Alzheimer’s Dis. 2013, 35, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Reddy, P.H. A New Discovery of MicroRNA-455-3p in Alzheimer’s Disease. J. Alzheimer’s Dis. 2019, 72, S117–S130. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, S.; Want, A.; Laskowska-Kaszub, K.; Fesiuk, A.; Vaz, S.; Logarinho, E.; Wojda, U. Candidate Alzheimer’s Disease Biomarker MiR-483-5p Lowers TAU Phosphorylation by Direct ERK1/2 Repression. Int. J. Mol. Sci. 2021, 22, 3653. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, J.; Wang, Q.; Jiang, H.; Zeng, L.; Li, Z.; Liu, R. MicroRNA-200a-3p Mediates Neuroprotection in Alzheimer-Related Deficits and Attenuates Amyloid-Beta Overproduction and Tau Hyperphosphorylation via Coregulating BACE1 and PRKACB. Front. Pharmacol. 2019, 10, 806. [Google Scholar] [CrossRef]
- Bekris, L.M.; Lutz, F.; Montine, T.J.; Yu, C.E.; Tsuang, D.; Peskind, E.R.; Leverenz, J.B. MicroRNA in Alzheimer’s Disease: An Exploratory Study in Brain, Cerebrospinal Fluid and Plasma. Biomarkers 2013, 18, 455–466. [Google Scholar] [CrossRef]
- Geekiyanage, H.; Jicha, G.A.; Nelson, P.T.; Chan, C. Blood Serum MiRNA: Non-Invasive Biomarkers for Alzheimer’s Disease. Exp. Neurol. 2012, 235, 491–496. [Google Scholar] [CrossRef]
- Turk, A.; Kunej, T.; Peterlin, B. MicroRNA-Target Interaction Regulatory Network in Alzheimer’s Disease. J. Pers. Med. 2021, 11, 1275. [Google Scholar] [CrossRef]
- Yang, G.; Song, Y.; Zhou, X.; Deng, Y.; Liu, T.; Weng, G.; Yu, D.; Pan, S. MicroRNA-29c Targets β-Site Amyloid Precursor Protein-Cleaving Enzyme 1 and Has a Neuroprotective Role in Vitro and in Vivo. Mol. Med. Rep. 2015, 12, 3081–3088. [Google Scholar] [CrossRef]
- Yılmaz, Ş.G.; Erdal, M.E.; Özge, A.A.; Sungur, M.A. Can Peripheral MicroRNA Expression Data Serve as Epigenomic (Upstream) Biomarkers of Alzheimer’s Disease? OMICS: A J. Integr. Biol. 2016, 20, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, X.; Chen, Y.-H.; Zhang, K. Identification of Circular RNA Hsa_Circ_0003391 in Peripheral Blood Is Potentially Associated with Alzheimer’s Disease. Front. Aging Neurosci. 2020, 12, 601965. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, N.; Fehlmann, T.; Kern, F.; Gogol, M.; Maetzler, W.; Deutscher, S.; Gurlit, S.; Schulte, C.; von Thaler, A.-K.; Deuschle, C.; et al. Machine Learning to Detect Alzheimer’s Disease from Circulating Non-Coding RNAs. Genom. Proteom. Bioinform. 2019, 17, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Guo, S.; Zhang, Y.; Zheng, Z.; Wang, H. Upregulation of MicroRNA-206 Enhances Lipopolysaccharide-Induced Inflammation and Release of Amyloid-β by Targeting Insulin-like Growth Factor 1 in Microglia. Mol. Med. Rep. 2016, 14, 1357–1364. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.Z.Y.; Thalamuthu, A.; Cheng, L.; Fowler, C.; Masters, C.L.; Sachdev, P.; Mather, K.A. Differential Blood MiRNA Expression in Brain Amyloid Imaging-Defined Alzheimer’s Disease and Controls. Alzheimer’s Res. Ther. 2020, 12, 59. [Google Scholar] [CrossRef] [PubMed]
- Karaglani, M.; Gourlia, K.; Tsamardinos, I.; Chatzaki, E. Accurate Blood-Based Diagnostic Biosignatures for Alzheimer’s Disease via Automated Machine Learning. J. Clin. Med. 2020, 9, 3016. [Google Scholar] [CrossRef]
- Gullett, J.M.; Chen, Z.; O’Shea, A.; Akbar, M.; Bian, J.; Rani, A.; Porges, E.C.; Foster, T.C.; Woods, A.J.; Modave, F.; et al. MicroRNA Predicts Cognitive Performance in Healthy Older Adults. Neurobiol. Aging 2020, 95, 186–194. [Google Scholar] [CrossRef]
- Yuen, S.C.; Liang, X.; Zhu, H.; Jia, Y.; Leung, S. Prediction of Differentially Expressed MicroRNAs in Blood as Potential Biomarkers for Alzheimer’s Disease by Meta-Analysis and Adaptive Boosting Ensemble Learning. Alzheimer’s Res. Ther. 2021, 13, 126. [Google Scholar] [CrossRef]
- Jia, L.-H.; Liu, Y.-N. Downregulated Serum MiR-223 Servers as Biomarker in Alzheimer’s Disease. Cell Biochem. Funct. 2016, 34, 233–237. [Google Scholar] [CrossRef]
- Galimberti, D.; Villa, C.; Fenoglio, C.; Serpente, M.; Ghezzi, L.; Cioffi, S.M.G.; Arighi, A.; Fumagalli, G.; Scarpini, E. Circulating MiRNAs as Potential Biomarkers in Alzheimer’s Disease. J. Alzheimer’s Dis. 2014, 42, 1261–1267. [Google Scholar] [CrossRef]
- Bhatnagar, S.; Chertkow, H.; Schipper, H.M.; Yuan, Z.; Shetty, V.; Jenkins, S.; Jones, T.; Wang, E. Increased MicroRNA-34c Abundance in Alzheimer’s Disease Circulating Blood Plasma. Front. Mol. Neurosci. 2014, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Hara, N.; Kikuchi, M.; Miyashita, A.; Hatsuta, H.; Saito, Y.; Kasuga, K.; Murayama, S.; Ikeuchi, T.; Kuwano, R. Serum MicroRNA MiR-501-3p as a Potential Biomarker Related to the Progression of Alzheimer’s Disease. Acta Neuropathol. Commun. 2017, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Vijayan, M.; Reddy, P.H. MicroRNA-455-3p as a Potential Peripheral Biomarker for Alzheimer’s Disease. Hum. Mol. Genet. 2017, 26, 3808–3822. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.; Backes, C.; Haas, J.; Leidinger, P.; Maetzler, W.; Deuschle, C.; Berg, D.; Ruschil, C.; Galata, V.; Ruprecht, K.; et al. Validating Alzheimer’s Disease Micro RNAs Using Next-generation Sequencing. Alzheimer’s Dement. 2016, 12, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Yu, J.-T.; Tan, M.-S.; Liu, Q.-Y.; Wang, H.-F.; Zhang, W.; Jiang, T.; Tan, L. Genome-Wide Serum MicroRNA Expression Profiling Identifies Serum Biomarkers for Alzheimer’s Disease. J. Alzheimer’s Dis. 2014, 40, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Shen, L.; Wang, Y.; Huang, S. Integrated Analysis of MiRNA and MRNA Expression in the Blood of Patients with Alzheimer’s Disease. Mol. Med. Rep. 2020, 22, 1053–1062. [Google Scholar] [CrossRef]
- Xu, A.; Kouznetsova, V.L.; Tsigelny, I.F. Alzheimer’s Disease Diagnostics Using MiRNA Biomarkers and Machine Learning. J. Alzheimer’s Dis. 2022, 86, 841–859. [Google Scholar] [CrossRef]
- Denk, J.; Oberhauser, F.; Kornhuber, J.; Wiltfang, J.; Fassbender, K.; Schroeter, M.L.; Volk, A.E.; Diehl-Schmid, J.; Prudlo, J.; Danek, A.; et al. Specific Serum and CSF MicroRNA Profiles Distinguish Sporadic Behavioural Variant of Frontotemporal Dementia Compared with Alzheimer Patients and Cognitively Healthy Controls. PLoS ONE 2018, 13, e0197329. [Google Scholar] [CrossRef]
- Martinez, B.; Peplow, P. MicroRNA Biomarkers in Frontotemporal Dementia and to Distinguish from Alzheimer’s Disease and Amyotrophic Lateral Sclerosis. Neural Regen. Res. 2022, 17, 1412. [Google Scholar] [CrossRef]
- Dong, H.; Li, J.; Huang, L.; Chen, X.; Li, D.; Wang, T.; Hu, C.; Xu, J.; Zhang, C.; Zen, K.; et al. Serum MicroRNA Profiles Serve as Novel Biomarkers for the Diagnosis of Alzheimer’s Disease. Dis. Markers 2015, 2015, 625659. [Google Scholar] [CrossRef]
- Sheinerman, K.S.; Toledo, J.B.; Tsivinsky, V.G.; Irwin, D.; Grossman, M.; Weintraub, D.; Hurtig, H.I.; Chen-Plotkin, A.; Wolk, D.A.; McCluskey, L.F.; et al. Circulating Brain-Enriched MicroRNAs as Novel Biomarkers for Detection and Differentiation of Neurodegenerative Diseases. Alzheimer’s Res. Ther. 2017, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, C.; Zhang, Y. An Investigation of MicroRNA-103 and MicroRNA-107 as Potential Blood-based Biomarkers for Disease Risk and Progression of Alzheimer’s Disease. J. Clin. Lab. Anal. 2020, 34, e23006. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.; Chen, Z.; Cai, P.; Wang, R.; Yang, Q.; Li, L.; Yang, H.; Zhu, R. Targeting MicroRNA-125b Promotes Neurite Outgrowth but Represses Cell Apoptosis and Inflammation via Blocking PTGS2 and CDK5 in a FOXQ1-Dependent Way in Alzheimer Disease. Front. Cell. Neurosci. 2020, 14, 587747. [Google Scholar] [CrossRef]
- Sabry, R.; el Sharkawy, R.E.; Gad, N.M. MiRNA -483-5p as a Potential Noninvasive Biomarker for Early Detection of Alzheimer’s Disease. Egypt. J. Immunol. 2020, 27, 59–72. [Google Scholar] [PubMed]
- Liu, C.-G.; Wang, J.-L.; Li, L.; Wang, P.-C. MicroRNA-384 Regulates Both Amyloid Precursor Protein and β-Secretase Expression and Is a Potential Biomarker for Alzheimer’s Disease. Int. J. Mol. Med. 2014, 34, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, S.; Laskowska-Kaszub, K.; Dębski, K.J.; Wojsiat, J.; Dąbrowski, M.; Gabryelewicz, T.; Kuźnicki, J.; Wojda, U. Profile of 6 MicroRNA in Blood Plasma Distinguish Early Stage Alzheimer’s Disease Patients from Non-Demented Subjects. Oncotarget 2017, 8, 16122–16143. [Google Scholar] [CrossRef] [PubMed]
- Cosín-Tomás, M.; Antonell, A.; Lladó, A.; Alcolea, D.; Fortea, J.; Ezquerra, M.; Lleó, A.; Martí, M.J.; Pallàs, M.; Sanchez-Valle, R.; et al. Plasma MiR-34a-5p and MiR-545-3p as Early Biomarkers of Alzheimer’s Disease: Potential and Limitations. Mol. Neurobiol. 2017, 54, 5550–5562. [Google Scholar] [CrossRef]
- He, H.; Liu, A.; Zhang, W.; Yang, H.; Zhang, M.; Xu, H.; Liu, Y.; Hong, B.; Yan, F.; Yue, L.; et al. Novel Plasma MiRNAs as Biomarkers and Therapeutic Targets of Alzheimer’s Disease at the Prodromal Stage. J. Alzheimer’s Dis. 2021, 83, 779–790. [Google Scholar] [CrossRef]
- Siedlecki-Wullich, D.; Català-Solsona, J.; Fábregas, C.; Hernández, I.; Clarimon, J.; Lleó, A.; Boada, M.; Saura, C.A.; Rodríguez-Álvarez, J.; Miñano-Molina, A.J. Altered MicroRNAs Related to Synaptic Function as Potential Plasma Biomarkers for Alzheimer’s Disease. Alzheimer’s Res. Ther. 2019, 11, 46. [Google Scholar] [CrossRef]
- Dakterzada, F.; David Benítez, I.; Targa, A.; Lladó, A.; Torres, G.; Romero, L.; de Gonzalo-Calvo, D.; Moncusí-Moix, A.; Tort-Merino, A.; Huerto, R.; et al. Reduced Levels of MiR-342-5p in Plasma Are Associated with Worse Cognitive Evolution in Patients with Mild Alzheimer’s Disease. Front. Aging Neurosci. 2021, 13, 705989. [Google Scholar] [CrossRef]
- Shi, Z.; Zhang, K.; Zhou, H.; Jiang, L.; Xie, B.; Wang, R.; Xia, W.; Yin, Y.; Gao, Z.; Cui, D.; et al. Increased MiR-34c Mediates Synaptic Deficits by Targeting Synaptotagmin 1 through ROS-JNK-p53 Pathway in Alzheimer’s Disease. Aging Cell 2020, 19, e13125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Y.; Liu, W.; Wang, T.; Wang, L.; Hao, L.; Ju, M.; Xiao, R. Diet Quality, Gut Microbiota, and MicroRNAs Associated with Mild Cognitive Impairment in Middle-Aged and Elderly Chinese Population. Am. J. Clin. Nutr. 2021, 114, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Chen, K.; Li, H.; Dong, S.; Su, N.; Liu, Y.; Cheng, Y.; Dai, J.; Yang, C.; Xiao, S. The Feasibility of Utilizing Plasma MiRNA107 and BACE1 Messenger RNA Gene Expression for Clinical Diagnosis of Amnestic Mild Cognitive Impairment. J. Clin. Psychiatry 2015, 76, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Li, C.; Sun, A.; Wang, Y.; Zhou, S. Quantification of MicroRNA-210 in the Cerebrospinal Fluid and Serum: Implications for Alzheimer’s Disease. Exp. Ther. Med. 2015, 9, 1013–1017. [Google Scholar] [CrossRef]
- Feng, L.; Liao, Y.-T.; He, J.-C.; Xie, C.-L.; Chen, S.-Y.; Fan, H.-H.; Su, Z.-P.; Wang, Z. Plasma Long Non-Coding RNA BACE1 as a Novel Biomarker for Diagnosis of Alzheimer Disease. BMC Neurol. 2018, 18, 4. [Google Scholar] [CrossRef]
- Fotuhi, S.N.; Khalaj-Kondori, M.; Hoseinpour Feizi, M.A.; Talebi, M. Long Non-Coding RNA BACE1-AS May Serve as an Alzheimer’s Disease Blood-Based Biomarker. J. Mol. Neurosci. 2019, 69, 351–359. [Google Scholar] [CrossRef]
- Barbagallo, C.; Mostile, G.; Baglieri, G.; Giunta, F.; Luca, A.; Raciti, L.; Zappia, M.; Purrello, M.; Ragusa, M.; Nicoletti, A. Specific Signatures of Serum MiRNAs as Potential Biomarkers to Discriminate Clinically Similar Neurodegenerative and Vascular-Related Diseases. Cell. Mol. Neurobiol. 2020, 40, 531–546. [Google Scholar] [CrossRef]
- Sheinerman, K.S.; Tsivinsky, V.G.; Crawford, F.; Mullan, M.J.; Abdullah, L.; Umansky, S.R. Plasma MicroRNA Biomarkers for Detection of Mild Cognitive Impairment. Aging 2012, 4, 590–605. [Google Scholar] [CrossRef]
- Sheinerman, K.S.; Tsivinsky, V.G.; Abdullah, L.; Crawford, F.; Umansky, S.R. Plasma MicroRNA Biomarkers for Detection of Mild Cognitive Impairment: Biomarker Validation Study. Aging 2013, 5, 925–938. [Google Scholar] [CrossRef]
- Kayano, M.; Higaki, S.; Satoh, J.; Matsumoto, K.; Matsubara, E.; Takikawa, O.; Niida, S. Plasma MicroRNA Biomarker Detection for Mild Cognitive Impairment Using Differential Correlation Analysis. Biomark. Res. 2016, 4, 22. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, L. Circulating Exosomal MiRNA as Diagnostic Biomarkers of Neurodegenerative Diseases. Front. Mol. Neurosci. 2020, 13, 53. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Zheng, D.; Nao, J. Circulating Exosome MicroRNAs as Diagnostic Biomarkers of Dementia. Front. Aging Neurosci. 2020, 12, 580199. [Google Scholar] [CrossRef] [PubMed]
- Hornung, S.; Dutta, S.; Bitan, G. CNS-Derived Blood Exosomes as a Promising Source of Biomarkers: Opportunities and Challenges. Front. Mol. Neurosci. 2020, 13, 38. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.-Y.; Lu, J.-M.; Zhao, Z.-Q.; Li, M.-C.; Lu, T.; An, X.-S.; Xue, L.-J. MicroRNA Biomarkers of Parkinson’s Disease in Serum Exosome-like Microvesicles. Neurosci. Lett. 2017, 644, 94–99. [Google Scholar] [CrossRef]
- Lugli, G.; Cohen, A.M.; Bennett, D.A.; Shah, R.C.; Fields, C.J.; Hernandez, A.G.; Smalheiser, N.R. Plasma Exosomal MiRNAs in Persons with and without Alzheimer Disease: Altered Expression and Prospects for Biomarkers. PLoS ONE 2015, 10, e0139233. [Google Scholar] [CrossRef]
- Cha, D.J.; Mengel, D.; Mustapic, M.; Liu, W.; Selkoe, D.J.; Kapogiannis, D.; Galasko, D.; Rissman, R.A.; Bennett, D.A.; Walsh, D.M. MiR-212 and MiR-132 Are Downregulated in Neurally Derived Plasma Exosomes of Alzheimer’s Patients. Front. Neurosci. 2019, 13, 1208. [Google Scholar] [CrossRef]
- Rani, A.; O’Shea, A.; Ianov, L.; Cohen, R.A.; Woods, A.J.; Foster, T.C. MiRNA in Circulating Microvesicles as Biomarkers for Age-Related Cognitive Decline. Front. Aging Neurosci. 2017, 9, 323. [Google Scholar] [CrossRef]
- Gámez-Valero, A.; Campdelacreu, J.; Vilas, D.; Ispierto, L.; Reñé, R.; Álvarez, R.; Armengol, M.P.; Borràs, F.E.; Beyer, K. Exploratory Study on MicroRNA Profiles from Plasma-Derived Extracellular Vesicles in Alzheimer’s Disease and Dementia with Lewy Bodies. Transl. Neurodegener. 2019, 8, 31. [Google Scholar] [CrossRef]
- Sproviero, D.; Gagliardi, S.; Zucca, S.; Arigoni, M.; Giannini, M.; Garofalo, M.; Olivero, M.; Dell’Orco, M.; Pansarasa, O.; Bernuzzi, S.; et al. Different MiRNA Profiles in Plasma Derived Small and Large Extracellular Vesicles from Patients with Neurodegenerative Diseases. Int. J. Mol. Sci. 2021, 22, 2737. [Google Scholar] [CrossRef]
- Cheng, L.; Doecke, J.D.; Sharples, R.A.; Villemagne, V.L.; Fowler, C.J.; Rembach, A.; Martins, R.N.; Rowe, C.C.; Macaulay, S.L.; Masters, C.L.; et al. Prognostic Serum MiRNA Biomarkers Associated with Alzheimer’s Disease Shows Concordance with Neuropsychological and Neuroimaging Assessment. Mol. Psychiatry 2015, 20, 1188–1196. [Google Scholar] [CrossRef]
- Wei, H.; Xu, Y.; Xu, W.; Zhou, Q.; Chen, Q.; Yang, M.; Feng, F.; Liu, Y.; Zhu, X.; Yu, M.; et al. Serum Exosomal MiR-223 Serves as a Potential Diagnostic and Prognostic Biomarker for Dementia. Neuroscience 2018, 379, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.G.; Meng, S.; Li, Y.; Lu, Y.; Zhao, Y.; Wang, P.C. MicroRNA-135a in ABCA1-Labeled Exosome Is a Serum Biomarker Candidate for Alzheimer’s Disease. Biomed. Environ. Sci. 2021, 34, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.T.; Liu, C.G.; Gao, S.C.; Zhang, Y.; Wang, P.C. The Serum Exosome Derived MicroRNA-135a, -193b, and -384 Were Potential Alzheimer’s Disease Biomarkers. Biomed. Environ. Sci. 2018, 31, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Fransquet, P.D.; Ryan, J. Micro RNA as a Potential Blood-Based Epigenetic Biomarker for Alzheimer’s Disease. Clin. Biochem. 2018, 58, 5–14. [Google Scholar] [CrossRef]
- Goh, S.Y.; Chao, Y.X.; Dheen, S.T.; Tan, E.-K.; Tay, S.S.-W. Role of MicroRNAs in Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 5649. [Google Scholar] [CrossRef]
- Wu, Y.-Y.; Kuo, H.-C. Functional Roles and Networks of Non-Coding RNAs in the Pathogenesis of Neurodegenerative Diseases. J. Biomed. Sci. 2020, 27, 49. [Google Scholar] [CrossRef]
- Görücü Yilmaz, Ş.; Erdal, M.E.; Avci Özge, A.; Sungur, M.A. SNP Variation in MicroRNA Biogenesis Pathway Genes as a New Innovation Strategy for Alzheimer Disease Diagnostics. Alzheimer Dis. Assoc. Disord. 2016, 30, 203–209. [Google Scholar] [CrossRef]
- Takousis, P. Analysis of Micro-RNA Expression by QPCR on a Microfluidics Platform for Alzheimer’s Disease Biomarker Discovery. Methods Mol. Biol. 2018, 1750, 283–292. [Google Scholar] [CrossRef]
- Nie, C.; Sun, Y.; Zhen, H.; Guo, M.; Ye, J.; Liu, Z.; Yang, Y.; Zhang, X. Differential Expression of Plasma Exo-MiRNA in Neurodegenerative Diseases by Next-Generation Sequencing. Front. Neurosci. 2020, 14, 438. [Google Scholar] [CrossRef]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long Non-Coding RNAs: Insights into Functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef]
- Wang, M.-Q.; Zhu, W.-J.; Gao, P. New Insights into Long Non-Coding RNAs in Breast Cancer: Biological Functions and Therapeutic Prospects. Exp. Mol. Pathol. 2021, 120, 104640. [Google Scholar] [CrossRef] [PubMed]
- Lan, Z.; Chen, Y.; Jin, J.; Xu, Y.; Zhu, X. Long Non-Coding RNA: Insight into Mechanisms of Alzheimer’s Disease. Front. Mol. Neurosci. 2022, 14, 821002. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-Z.; Cheng, M.-G.; Wang, X.; Hu, Y. The Emerging Role of Non-Coding RNAs from Extracellular Vesicles in Alzheimer’s Disease. J. Integr. Neurosci. 2021, 20, 239. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Xue, W.; Tao, L.; Zhu, F. Identification of Key Long Non-Coding RNAs in the Pathology of Alzheimer’s Disease and Their Functions Based on Genome-Wide Associations Study, Microarray, and RNA-Seq Data. J. Alzheimer’s Dis. 2019, 68, 339–355. [Google Scholar] [CrossRef]
- Massone, S.; Ciarlo, E.; Vella, S.; Nizzari, M.; Florio, T.; Russo, C.; Cancedda, R.; Pagano, A. NDM29, a RNA Polymerase III-Dependent Non Coding RNA, Promotes Amyloidogenic Processing of APP and Amyloid β Secretion. Biochim. Biophys. Acta-Mol. Cell Res. 2012, 1823, 1170–1177. [Google Scholar] [CrossRef]
- Faghihi, M.A.; Modarresi, F.; Khalil, A.M.; Wood, D.E.; Sahagan, B.G.; Morgan, T.E.; Finch, C.E.; St. Laurent, G., III; Kenny, P.J.; Wahlestedt, C. Expression of a Noncoding RNA Is Elevated in Alzheimer’s Disease and Drives Rapid Feed-Forward Regulation of β-Secretase. Nat. Med. 2008, 14, 723–730. [Google Scholar] [CrossRef]
- Ciarlo, E.; Massone, S.; Penna, I.; Nizzari, M.; Gigoni, A.; Dieci, G.; Russo, C.; Florio, T.; Cancedda, R.; Pagano, A. An Intronic NcRNA-Dependent Regulation of SORL1 Expression Affecting Aβ Formation Is Upregulated in Post-Mortem Alzheimer’s Disease Brain Samples. Dis. Models Mech. 2012, 6, 424–433. [Google Scholar] [CrossRef]
- Yan, Y.; Yan, H.; Teng, Y.; Wang, Q.; Yang, P.; Zhang, L.; Cheng, H.; Fu, S. Long Non-coding RNA 00507/MiRNA-181c-5p/TTBK1/MAPT Axis Regulates Tau Hyperphosphorylation in Alzheimer’s Disease. J. Gene Med. 2020, 22, 3268. [Google Scholar] [CrossRef]
- Massone, S.; Vassallo, I.; Fiorino, G.; Castelnuovo, M.; Barbieri, F.; Borghi, R.; Tabaton, M.; Robello, M.; Gatta, E.; Russo, C.; et al. 17A, a Novel Non-Coding RNA, Regulates GABA B Alternative Splicing and Signaling in Response to Inflammatory Stimuli and in Alzheimer Disease. Neurobiol. Dis. 2011, 41, 308–317. [Google Scholar] [CrossRef]
- Aliperti, V.; Skonieczna, J.; Cerase, A. Long Non-Coding RNA (LncRNA) Roles in Cell Biology, Neurodevelopment and Neurological Disorders. Non-Coding RNA 2021, 7, 36. [Google Scholar] [CrossRef]
- Wang, D.; Wang, P.; Bian, X.; Xu, S.; Zhou, Q.; Zhang, Y.; Ding, M.; Han, M.; Huang, L.; Bi, J.; et al. Elevated Plasma Levels of Exosomal BACE1-AS Combined with the Volume and Thickness of the Right Entorhinal Cortex May Serve as a Biomarker for the Detection of Alzheimer’s Disease. Mol. Med. Rep. 2020, 22, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Kao, Y.-C.; Ho, P.-C.; Tu, Y.-K.; Jou, I.-M.; Tsai, K.-J. Lipids and Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 1505. [Google Scholar] [CrossRef] [PubMed]
- Chew, H.; Solomon, V.A.; Fonteh, A.N. Involvement of Lipids in Alzheimer’s Disease Pathology and Potential Therapies. Front. Physiol. 2020, 11, 598. [Google Scholar] [CrossRef] [PubMed]
- Casas-Fernández, E.; Peña-Bautista, C.; Baquero, M.; Cháfer-Pericás, C. Lipids as Early and Minimally Invasive Biomarkers for Alzheimer Disease. Curr. Neuropharmacol. 2021, 19. [Google Scholar] [CrossRef] [PubMed]
- Blasko, I.; Defrancesco, M.; Oberacher, H.; Loacker, L.; Kemmler, G.; Marksteiner, J.; Humpel, C. Plasma Phosphatidylcholines and Vitamin B12/Folate Levels Are Possible Prognostic Biomarkers for Progression of Alzheimer’s Disease. Exp. Gerontol. 2021, 147, 111264. [Google Scholar] [CrossRef] [PubMed]
- Fonteh, A.N.; Chiang, A.J.; Arakaki, X.; Edminster, S.P.; Harrington, M.G. Accumulation of Cerebrospinal Fluid Glycerophospholipids and Sphingolipids in Cognitively Healthy Participants with Alzheimer’s Biomarkers Precedes Lipolysis in the Dementia Stage. Front. Neurosci. 2020, 14, 611393. [Google Scholar] [CrossRef]
- Benussi, L.; Ghidoni, R.; Dal Piaz, F.; Binetti, G.; di Iorio, G.; Abrescia, P. The Level of 24-Hydroxycholesteryl Esters Is an Early Marker of Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 56, 825–833. [Google Scholar] [CrossRef]
- Roy, D.; Chakrabarti, S.S.; Banerjee, A.; Sharma, P.; Biswas, A.; Chakrabarti, S. Serum 24-hydroxycholesterol in Probable Alzheimer’s Dementia: Reexploring the Significance of a Tentative Alzheimer’s Disease Biomarker. Aging Med. 2019, 2, 74–81. [Google Scholar] [CrossRef]
- Wellington, C.L.; Frikke-Schmidt, R. Relation between Plasma and Brain Lipids. Curr. Opin. Lipidol. 2016, 27, 225–232. [Google Scholar] [CrossRef]
- Sakr, F.; Dyrba, M.; Bräuer, A.; Teipel, S. Association of Lipidomics Signatures in Blood with Clinical Progression in Preclinical and Prodromal Alzheimer’s Disease. J. Alzheimer’s Dis. 2022, 85, 1115–1127. [Google Scholar] [CrossRef]
- Tokuoka, S.M.; Kita, Y.; Shimizu, T.; Oda, Y. Isobaric Mass Tagging and Triple Quadrupole Mass Spectrometry to Determine Lipid Biomarker Candidates for Alzheimer’s Disease. PLoS ONE 2019, 14, e0226073. [Google Scholar] [CrossRef] [PubMed]
- Briganti, S.; Truglio, M.; Angiolillo, A.; Lombardo, S.; Leccese, D.; Camera, E.; Picardo, M.; di Costanzo, A. Application of Serum Lipidomics to Biomarkers Discovery in Neurodegenerative Diseases. Metabolites 2021, 11, 819. [Google Scholar] [CrossRef] [PubMed]
- Praticò, D.; Clark, C.M.; Lee, V.M.; Trojanowski, J.Q.; Rokach, J.; FitzGerald, G.A. Increased 8,12-Iso-IPF2alpha-VI in Alzheimer’s Disease: Correlation of a Noninvasive Index of Lipid Peroxidation with Disease Severity. Ann. Neurol. 2000, 48, 809–812. [Google Scholar] [CrossRef]
- Skoumalová, A.; Mádlová, P.; Topinková, E. End Products of Lipid Peroxidation in Erythrocyte Membranes in Alzheimer’s Disease. Cell Biochem. Funct. 2012, 30, 205–210. [Google Scholar] [CrossRef]
- Peña-Bautista, C.; Vigor, C.; Galano, J.-M.; Oger, C.; Durand, T.; Ferrer, I.; Cuevas, A.; López-Cuevas, R.; Baquero, M.; López-Nogueroles, M.; et al. Plasma Lipid Peroxidation Biomarkers for Early and Non-Invasive Alzheimer Disease Detection. Free Radic. Biol. Med. 2018, 124, 388–394. [Google Scholar] [CrossRef]
- Zarrouk, A.; Riedinger, J.-M.; Ahmed, S.H.; Hammami, S.; Chaabane, W.; Debbabi, M.; ben Ammou, S.; Rouaud, O.; Frih, M.; Lizard, G.; et al. Fatty Acid Profiles in Demented Patients: Identification of Hexacosanoic Acid (C26:0) as a Blood Lipid Biomarker of Dementia. J. Alzheimer’s Dis. 2015, 44, 1349–1359. [Google Scholar] [CrossRef]
- Peña-Bautista, C.; Álvarez-Sánchez, L.; Cañada-Martínez, A.J.; Baquero, M.; Cháfer-Pericás, C. Epigenomics and Lipidomics Integration in Alzheimer Disease: Pathways Involved in Early Stages. Biomedicines 2021, 9, 1812. [Google Scholar] [CrossRef]
- Kim, M.; Snowden, S.; Suvitaival, T.; Ali, A.; Merkler, D.J.; Ahmad, T.; Westwood, S.; Baird, A.; Proitsi, P.; Nevado-Holgado, A.; et al. Primary Fatty Amides in Plasma Associated with Brain Amyloid Burden, Hippocampal Volume, and Memory in the European Medical Information Framework for Alzheimer’s Disease Biomarker Discovery Cohort. Alzheimer’s Dement. 2019, 15, 817–827. [Google Scholar] [CrossRef]
- Wang, D.-C.; Sun, C.-H.; Liu, L.-Y.; Sun, X.-H.; Jin, X.-W.; Song, W.-L.; Liu, X.-Q.; Wan, X.-L. Serum Fatty Acid Profiles Using GC-MS and Multivariate Statistical Analysis: Potential Biomarkers of Alzheimer’s Disease. Neurobiol. Aging 2012, 33, 1057–1066. [Google Scholar] [CrossRef]
- Sánchez-Campillo, M.; Ruiz-Pastor, M.J.; Gázquez, A.; Marín-Muñoz, J.; Noguera-Perea, F.; Ruiz-Alcaraz, A.J.; Manzanares-Sánchez, S.; Antúnez, C.; Larqué, E. Decreased Blood Level of MFSD2a as a Potential Biomarker of Alzheimer’s Disease. Int. J. Mol. Sci. 2019, 21, 70. [Google Scholar] [CrossRef]
- Mielke, M.M.; Haughey, N.J. Could Plasma Sphingolipids Be Diagnostic or Prognostic Biomarkers for Alzheimer’s Disease? Clin. Lipidol. 2012, 7, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Llano, D.A.; Devanarayan, V. Serum Phosphatidylethanolamine and Lysophosphatidylethanolamine Levels Differentiate Alzheimer’s Disease from Controls and Predict Progression from Mild Cognitive Impairment. J. Alzheimer’s Dis. 2021, 80, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Nevado-Holgado, A.; Whiley, L.; Snowden, S.G.; Soininen, H.; Kloszewska, I.; Mecocci, P.; Tsolaki, M.; Vellas, B.; Thambisetty, M.; et al. Association between Plasma Ceramides and Phosphatidylcholines and Hippocampal Brain Volume in Late Onset Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 60, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Llano, D.; Issa, L.; Devanarayan, P.; Devanarayan, V. Hearing Loss in Alzheimer’s Disease Is Associated with Altered Serum Lipidomic Biomarker Profiles. Cells 2020, 9, 2556. [Google Scholar] [CrossRef] [PubMed]
- Klavins, K.; Koal, T.; Dallmann, G.; Marksteiner, J.; Kemmler, G.; Humpel, C. The Ratio of Phosphatidylcholines to Lysophosphatidylcholines in Plasma Differentiates Healthy Controls from Patients with Alzheimer’s Disease and Mild Cognitive Impairment. Alzheimer’s Dement. Diagn. Assess. Dis. Monit. 2015, 1, 295–302. [Google Scholar] [CrossRef]
- Mapstone, M.; Cheema, A.K.; Fiandaca, M.S.; Zhong, X.; Mhyre, T.R.; MacArthur, L.H.; Hall, W.J.; Fisher, S.G.; Peterson, D.R.; Haley, J.M.; et al. Plasma Phospholipids Identify Antecedent Memory Impairment in Older Adults. Nat. Med. 2014, 20, 415–418. [Google Scholar] [CrossRef]
- Simpson, B.N.; Kim, M.; Chuang, Y.-F.; Beason-Held, L.; Kitner-Triolo, M.; Kraut, M.; Lirette, S.T.; Windham, B.G.; Griswold, M.E.; Legido-Quigley, C.; et al. Blood Metabolite Markers of Cognitive Performance and Brain Function in Aging. J. Cereb. Blood Flow Metab. 2016, 36, 1212–1223. [Google Scholar] [CrossRef]
- Weng, W.-C.; Huang, W.-Y.; Tang, H.-Y.; Cheng, M.-L.; Chen, K.-H. The Differences of Serum Metabolites Between Patients with Early-Stage Alzheimer’s Disease and Mild Cognitive Impairment. Front. Neurol. 2019, 10, 1223. [Google Scholar] [CrossRef]
- Liu, Y.; Zhong, X.; Shen, J.; Jiao, L.; Tong, J.; Zhao, W.; Du, K.; Gong, S.; Liu, M.; Wei, M. Elevated Serum TC and LDL-C Levels in Alzheimer’s Disease and Mild Cognitive Impairment: A Meta-Analysis Study. Brain Res. 2020, 1727, 146554. [Google Scholar] [CrossRef]
- Sato, Y.; Bernier, F.; Yamanaka, Y.; Aoshima, K.; Oda, Y.; Ingelsson, M.; Lannfelt, L.; Miyashita, A.; Kuwano, R.; Ikeuchi, T. Reduced Plasma Desmosterol-to-cholesterol Ratio and Longitudinal Cognitive Decline in Alzheimer’s Disease. Alzheimer’s Dement. Diagn. Assess. Dis. Monit. 2015, 1, 67–74. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Z.; Jia, X.; Zhang, H.; Zhang, H.; Li, J.; Zhang, K. Prediction of Alzheimer’s Disease with Serum Lipid Levels in Asian Individuals: A Meta-Analysis. Biomarkers 2019, 24, 341–351. [Google Scholar] [CrossRef]
- Tang, Q.; Wang, F.; Yang, J.; Peng, H.; Li, Y.; Li, B.; Wang, S. Revealing a Novel Landscape of the Association Between Blood Lipid Levels and Alzheimer’s Disease: A Meta-Analysis of a Case-Control Study. Front. Aging Neurosci. 2020, 11, 370. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Cheng, Y.; Jin, Y.; Unverzagt, F.; Su, L.; Yang, L.; Ma, F.; Hake, A.; Kettler, C.; Chen, C.; et al. The Relationship between Cholesterol and Cognitive Function Is Homocysteine-Dependent. Clin. Interv. Aging 2014, 2014, 1823–1829. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huang, S.; Ma, Y.; Bi, Y.; Shen, X.; Hou, X.; Cao, X.; Ou, Y.; Zhao, B.; Dong, Q.; Tan, L.; et al. Metabolically Healthy Obesity and Lipids May Be Protective Factors for Pathological Changes of Alzheimer’s Disease in Cognitively Normal Adults. J. Neurochem. 2021, 157, 834–845. [Google Scholar] [CrossRef] [PubMed]
- Kjeldsen, E.W.; Thomassen, J.Q.; Juul Rasmussen, I.; Nordestgaard, B.G.; Tybjærg-Hansen, A.; Frikke-Schmidt, R. Plasma High-Density Lipoprotein Cholesterol and Risk of Dementia: Observational and Genetic Studies. Cardiovasc. Res. 2021, 118, 1330–1340. [Google Scholar] [CrossRef]
- Yun, S.-M.; Park, J.-Y.; Seo, S.W.; Song, J. Association of Plasma Endothelial Lipase Levels on Cognitive Impairment. BMC Psychiatry 2019, 19, 187. [Google Scholar] [CrossRef] [PubMed]
- Montagne, A.; Nation, D.A.; Sagare, A.P.; Barisano, G.; Sweeney, M.D.; Chakhoyan, A.; Pachicano, M.; Joe, E.; Nelson, A.R.; D’Orazio, L.M.; et al. APOE4 Leads to Blood–Brain Barrier Dysfunction Predicting Cognitive Decline. Nature 2020, 581, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Lütjohann, D.; von Bergmann, K. 24S-Hydroxycholesterol: A Marker of Brain Cholesterol Metabolism. Pharmacopsychiatry 2003, 36, 102–106. [Google Scholar] [CrossRef]
- di Natale, C.; Monaco, A.; Pedone, C.; Tessitore, A.; de Mase, A.; Tedeschi, G.; Netti, P.A.; Abrescia, P. The Level of 24-Hydroxycholesteryl Esters Decreases in Plasma of Patients with Parkinson’s Disease. Neurosci. Lett. 2018, 672, 108–112. [Google Scholar] [CrossRef]
- Tardy, A.-L.; Pouteau, E.; Marquez, D.; Yilmaz, C.; Scholey, A. Vitamins and Minerals for Energy, Fatigue and Cognition: A Narrative Review of the Biochemical and Clinical Evidence. Nutrients 2020, 12, 228. [Google Scholar] [CrossRef]
- Mora, J.R.; Iwata, M.; von Andrian, U.H. Vitamin Effects on the Immune System: Vitamins A and D Take Centre Stage. Nat. Rev. Immunol. 2008, 8, 685–698. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D. B Vitamins and the Brain: Mechanisms, Dose and Efficacy—A Review. Nutrients 2016, 8, 68. [Google Scholar] [CrossRef] [PubMed]
- Caccamo, D.; Ricca, S.; Currò, M.; Ientile, R. Health Risks of Hypovitaminosis D: A Review of New Molecular Insights. Int. J. Mol. Sci. 2018, 19, 892. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.; Piccio, L.; Cross, A.H. Use of Vitamins and Dietary Supplements by Patients with Multiple Sclerosis. JAMA Neurol. 2018, 75, 1013. [Google Scholar] [CrossRef]
- Ames, B.N. Prolonging Healthy Aging: Longevity Vitamins and Proteins. Proc. Natl. Acad. Sci. USA 2018, 115, 10836–10844. [Google Scholar] [CrossRef]
- Glasø, M.; Nordbø, G.; Diep, L.; Bøhmer, T. Reduced Concentrations of Several Vitamins in Normal Weight Patients with Late-Onset Dementia of the Alzheimer Type without Vascular Disease. J. Nutr. Health Aging 2004, 8, 407–413. [Google Scholar]
- Gold, M.; Chen, M.F.; Johnson, K. Plasma and Red Blood Cell Thiamine Deficiency in Patients with Dementia of the Alzheimer’s Type. Arch. Neurol. 1995, 52, 1081–1086. [Google Scholar] [CrossRef]
- Wang, C.; Fei, G.; Pan, X.; Sang, S.; Wang, L.; Zhong, C.; Jin, L. High Thiamine Diphosphate Level as a Protective Factor for Alzheimer’s Disease. Neurol. Res. 2018, 40, 432–442. [Google Scholar] [CrossRef]
- D’Cunha, N.M.; Georgousopoulou, E.N.; Boyd, L.; Veysey, M.; Sturm, J.; O’Brien, B.; Lucock, M.; McKune, A.J.; Mellor, D.D.; Roach, P.D.; et al. Relationship Between B-Vitamin Biomarkers and Dietary Intake with Apolipoprotein E є4 in Alzheimer’s Disease. J. Nutr. Gerontol. Geriatr. 2019, 38, 173–195. [Google Scholar] [CrossRef]
- An, Y.; Feng, L.; Zhang, X.; Wang, Y.; Wang, Y.; Tao, L.; Qin, Z.; Xiao, R. Dietary Intakes and Biomarker Patterns of Folate, Vitamin B6, and Vitamin B12 Can Be Associated with Cognitive Impairment by Hypermethylation of Redox-Related Genes NUDT15 and TXNRD1. Clin. Epigenet. 2019, 11, 139. [Google Scholar] [CrossRef]
- Murdaca, G.; Banchero, S.; Tonacci, A.; Nencioni, A.; Monacelli, F.; Gangemi, S. Vitamin D and Folate as Predictors of MMSE in Alzheimer’s Disease: A Machine Learning Analysis. Diagnostics 2021, 11, 940. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, G.A.A.; Pardi, P.C. Biomarkers in Alzheimer’s Disease: Evaluation of Platelets, Hemoglobin and Vitamin B12. Dement. Neuropsychol. 2020, 14, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Baldacci, F.; Lista, S.; Manca, M.L.; Chiesa, P.A.; Cavedo, E.; Lemercier, P.; Zetterberg, H.; Blennow, K.; Habert, M.-O.; Potier, M.C.; et al. Age and Sex Impact Plasma NFL and T-Tau Trajectories in Individuals with Subjective Memory Complaints: A 3-Year Follow-up Study. Alzheimer’s Res. Ther. 2020, 12, 147. [Google Scholar] [CrossRef] [PubMed]
- Dursun, E.; Alaylıoğlu, M.; Bilgiç, B.; Hanağası, H.; Lohmann, E.; Atasoy, I.L.; Candaş, E.; Araz, Ö.S.; Önal, B.; Gürvit, H.; et al. Vitamin D Deficiency Might Pose a Greater Risk for ApoEε4 Non-Carrier Alzheimer’s Disease Patients. Neurol. Sci. 2016, 37, 1633–1643. [Google Scholar] [CrossRef]
- Ouma, S.; Suenaga, M.; Bölükbaşı Hatip, F.F.; Hatip-Al-Khatib, I.; Tsuboi, Y.; Matsunaga, Y. Serum Vitamin D in Patients with Mild Cognitive Impairment and Alzheimer’s Disease. Brain Behav. 2018, 8, e00936. [Google Scholar] [CrossRef]
- Hooshmand, B.; Lökk, J.; Solomon, A.; Mangialasche, F.; Miralbell, J.; Spulber, G.; Annerbo, S.; Andreasen, N.; Winblad, B.; Cedazo-Minguez, A.; et al. Vitamin D in Relation to Cognitive Impairment, Cerebrospinal Fluid Biomarkers, and Brain Volumes. J. Gerontol. Ser. A 2014, 69, 1132–1138. [Google Scholar] [CrossRef]
- Al-Amin, M.; Bradford, D.; Sullivan, R.K.P.; Kurniawan, N.D.; Moon, Y.; Han, S.-H.; Zalesky, A.; Burne, T.H.J. Vitamin D Deficiency Is Associated with Reduced Hippocampal Volume and Disrupted Structural Connectivity in Patients with Mild Cognitive Impairment. Hum. Brain Mapp. 2019, 40, 394–406. [Google Scholar] [CrossRef]
- Raszewski, G.; Chwedorowicz, R.; Chwedorowicz, A.; Gustaw Rothenberg, K. Homocysteine, Antioxidant Vitamins and Lipids as Biomarkers of Neurodegeneration in Alzheimer’s Disease versus Non-Alzheimer’s Dementia. Ann. Agric. Environ. Med. 2015, 23, 193–196. [Google Scholar] [CrossRef][Green Version]
- Lanyau-Domínguez, Y.; Macías-Matos, C.; Llibre-Rodríguez, J.d.J.; Pita-Rodríguez, G.M.; Suárez-Medina, R.; Quintero-Alejo, M.E.; Noriega-Fernández, L.; Guerra-Hernández, M.; Calvo-Rodríguez, M.; Sánchez-Gil, Y.; et al. Levels of Vitamins and Homocysteine in Older Adults with Alzheimer Disease or Mild Cognitive Impairment in Cuba. MEDICC Rev. 2020, 22, 40. [Google Scholar] [CrossRef]
- Gibson, G.E.; Blass, J.P. Thiamine-Dependent Processes and Treatment Strategies in Neurodegeneration. Antioxid. Redox Signal. 2007, 9, 1605–1620. [Google Scholar] [CrossRef]
- Cordaro, M.; Siracusa, R.; Fusco, R.; Cuzzocrea, S.; di Paola, R.; Impellizzeri, D. Involvements of Hyperhomocysteinemia in Neurological Disorders. Metabolites 2021, 11, 37. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.; Wengreen, H.J.; Munger, R.G.; Corcoran, C.D. Dietary Folate, Vitamin B-12, Vitamin B-6 and Incident Alzheimer’s Disease: The Cache County Memory, Health, and Aging Study. J. Nutr. Health Aging 2009, 13, 899–905. [Google Scholar] [CrossRef] [PubMed]
- Lue, L.-F.; Sabbagh, M.N.; Chiu, M.-J.; Jing, N.; Snyder, N.L.; Schmitz, C.; Guerra, A.; Belden, C.M.; Chen, T.-F.; Yang, C.-C.; et al. Plasma Levels of Aβ42 and Tau Identified Probable Alzheimer’s Dementia: Findings in Two Cohorts. Front. Aging Neurosci. 2017, 9, 226. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer’s Association 2016 Alzheimer’s Disease Facts and Figures. Alzheimer’s Dement. 2016, 12, 459–509. [CrossRef]
- McLimans, K.E.; Collazo Martinez, A.D.; Mochel, J.P.; Allenspach, K.; Willette, A.A. Serum Vitamin B12 and Related 5-Methyltetrahydrofolate-Homocysteine Methyltransferase Reductase and Cubilin Genotypes Predict Neural Outcomes across the Alzheimer’s Disease Spectrum. Br. J. Nutr. 2020, 124, 135–145. [Google Scholar] [CrossRef]
- Dursun, E.; Gezen-Ak, D. Vitamin D Basis of Alzheimer’s Disease: From Genetics to Biomarkers. Hormones 2019, 18, 7–15. [Google Scholar] [CrossRef]
- Annweiler, C.; Dursun, E.; Féron, F.; Gezen-Ak, D.; Kalueff, A.V.; Littlejohns, T.; Llewellyn, D.; Millet, P.; Scott, T.; Tucker, K.L.; et al. Vitamin D and Cognition in Older Adults: International Consensus Guidelines. Gériatrie Psychol. Neuropsychiatr. Viellissement 2016, 14, 265–273. [Google Scholar] [CrossRef]
- Blennow, K.; Hampel, H.; Weiner, M.; Zetterberg, H. Cerebrospinal Fluid and Plasma Biomarkers in Alzheimer Disease. Nat. Rev. Neurol. 2010, 6, 131–144. [Google Scholar] [CrossRef]
- Soares, J.Z.; Pettersen, R.; Benth, J.Š.; Persson, K.; Strobel, C.; Selbæk, G.; Bogdanovic, N. Vitamin D Levels, APOE Allele, and MRI Volumetry Assessed by NeuroQuant in Norwegian Adults with Cognitive Symptoms. J. Alzheimer’s Dis. 2021, 79, 311–321. [Google Scholar] [CrossRef]
- Huebbe, P.; Nebel, A.; Siegert, S.; Moehring, J.; Boesch-Saadatmandi, C.; Most, E.; Pallauf, J.; Egert, S.; Müller, M.J.; Schreiber, S.; et al. APOEε4 Is Associated with Higher Vitamin D Levels in Targeted Replacement Mice and Humans. FASEB J. 2011, 25, 3262–3270. [Google Scholar] [CrossRef]
- Bivona, G.; lo Sasso, B.; Iacolino, G.; Gambino, C.M.; Scazzone, C.; Agnello, L.; Ciaccio, M. Standardized Measurement of Circulating Vitamin D [25(OH)D] and Its Putative Role as a Serum Biomarker in Alzheimer’s Disease and Parkinson’s Disease. Clin. Chim. Acta 2019, 497, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Varesi, A.; Deumer, U.-S.; Ananth, S.; Ricevuti, G. The Emerging Role of Gut Microbiota in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS): Current Evidence and Potential Therapeutic Applications. J. Clin. Med. 2021, 10, 5077. [Google Scholar] [CrossRef] [PubMed]
- Ling, Z.; Liu, X.; Jia, X.; Cheng, Y.; Luo, Y.; Yuan, L.; Wang, Y.; Zhao, C.; Guo, S.; Li, L.; et al. Impacts of Infection with Different Toxigenic Clostridium Difficile Strains on Faecal Microbiota in Children. Sci. Rep. 2015, 4, 7485. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef] [PubMed]
- Abenavoli, L.; Scarpellini, E.; Colica, C.; Boccuto, L.; Salehi, B.; Sharifi-Rad, J.; Aiello, V.; Romano, B.; de Lorenzo, A.; Izzo, A.A.; et al. Gut Microbiota and Obesity: A Role for Probiotics. Nutrients 2019, 11, 2690. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, R.F.; Jobin, C. The Microbiome and Cancer. Nat. Rev. Cancer 2013, 13, 800–812. [Google Scholar] [CrossRef]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the Immune System. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef]
- Jiang, H.; Ling, Z.; Zhang, Y.; Mao, H.; Ma, Z.; Yin, Y.; Wang, W.; Tang, W.; Tan, Z.; Shi, J.; et al. Altered Fecal Microbiota Composition in Patients with Major Depressive Disorder. Brain Behav. Immun. 2015, 48, 186–194. [Google Scholar] [CrossRef]
- Gerhardt, S.; Mohajeri, M. Changes of Colonic Bacterial Composition in Parkinson’s Disease and Other Neurodegenerative Diseases. Nutrients 2018, 10, 708. [Google Scholar] [CrossRef]
- Sun, M.-F.; Shen, Y.-Q. Dysbiosis of Gut Microbiota and Microbial Metabolites in Parkinson’s Disease. Ageing Res. Rev. 2018, 45, 53–61. [Google Scholar] [CrossRef]
- Gotkine, M.; Kviatcovsky, D.; Elinav, E. Amyotrophic Lateral Sclerosis and Intestinal Microbiota—Toward Establishing Cause and Effect. Gut Microbes 2020, 11, 1833–1841. [Google Scholar] [CrossRef] [PubMed]
- Cenit, M.C.; Sanz, Y.; Codoñer-Franch, P. Influence of Gut Microbiota on Neuropsychiatric Disorders. World J. Gastroenterol. 2017, 23, 5486. [Google Scholar] [CrossRef] [PubMed]
- Bello-Medina, P.C.; Hernández-Quiroz, F.; Pérez-Morales, M.; González-Franco, D.A.; Cruz-Pauseno, G.; García-Mena, J.; Díaz-Cintra, S.; Pacheco-López, G. Spatial Memory and Gut Microbiota Alterations Are Already Present in Early Adulthood in a Pre-Clinical Transgenic Model of Alzheimer’s Disease. Front. Neurosci. 2021, 15, 595583. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, A.; Cattane, N.; Galluzzi, S.; Provasi, S.; Lopizzo, N.; Festari, C.; Ferrari, C.; Guerra, U.P.; Paghera, B.; Muscio, C.; et al. Association of Brain Amyloidosis with Pro-Inflammatory Gut Bacterial Taxa and Peripheral Inflammation Markers in Cognitively Impaired Elderly. Neurobiol. Aging 2017, 49, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Vogt, N.M.; Kerby, R.L.; Dill-McFarland, K.A.; Harding, S.J.; Merluzzi, A.P.; Johnson, S.C.; Carlsson, C.M.; Asthana, S.; Zetterberg, H.; Blennow, K.; et al. Gut Microbiome Alterations in Alzheimer’s Disease. Sci. Rep. 2017, 7, 13537. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.-Q.; Shen, L.-L.; Li, W.-W.; Fu, X.; Zeng, F.; Gui, L.; Lü, Y.; Cai, M.; Zhu, C.; Tan, Y.-L.; et al. Gut Microbiota Is Altered in Patients with Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 63, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Alhasson, F.; Das, S.; Seth, R.; Dattaroy, D.; Chandrashekaran, V.; Ryan, C.N.; Chan, L.S.; Testerman, T.; Burch, J.; Hofseth, L.J.; et al. Altered Gut Microbiome in a Mouse Model of Gulf War Illness Causes Neuroinflammation and Intestinal Injury via Leaky Gut and TLR4 Activation. PLoS ONE 2017, 12, e0172914. [Google Scholar] [CrossRef]
- Varesi, A.; Pierella, E.; Romeo, M.; Piccini, G.B.; Alfano, C.; Bjørklund, G.; Oppong, A.; Ricevuti, G.; Esposito, C.; Chirumbolo, S.; et al. The Potential Role of Gut Microbiota in Alzheimer’s Disease: From Diagnosis to Treatment. Nutrients 2022, 14, 668. [Google Scholar] [CrossRef]
- Vogt, N.M.; Romano, K.A.; Darst, B.F.; Engelman, C.D.; Johnson, S.C.; Carlsson, C.M.; Asthana, S.; Blennow, K.; Zetterberg, H.; Bendlin, B.B.; et al. The Gut Microbiota-Derived Metabolite Trimethylamine N-Oxide Is Elevated in Alzheimer’s Disease. Alzheimer’s Res. Ther. 2018, 10, 124. [Google Scholar] [CrossRef]
- Yan, Y.; Gao, Y.; Fang, Q.; Zhang, N.; Kumar, G.; Yan, H.; Song, L.; Li, J.; Zhang, Y.; Sun, J.; et al. Inhibition of Rho Kinase by Fasudil Ameliorates Cognition Impairment in APP/PS1 Transgenic Mice via Modulation of Gut Microbiota and Metabolites. Front. Aging Neurosci. 2021, 13, 755164. [Google Scholar] [CrossRef]
- Wu, L.; Han, Y.; Zheng, Z.; Peng, G.; Liu, P.; Yue, S.; Zhu, S.; Chen, J.; Lv, H.; Shao, L.; et al. Altered Gut Microbial Metabolites in Amnestic Mild Cognitive Impairment and Alzheimer’s Disease: Signals in Host–Microbe Interplay. Nutrients 2021, 13, 228. [Google Scholar] [CrossRef] [PubMed]
- Valeri, F.; dos Santos Guilherme, M.; He, F.; Stoye, N.M.; Schwiertz, A.; Endres, K. Impact of the Age of Cecal Material Transfer Donors on Alzheimer’s Disease Pathology in 5xFAD Mice. Microorganisms 2021, 9, 2548. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Jaber, V.; Lukiw, W.J. Secretory Products of the Human GI Tract Microbiome and Their Potential Impact on Alzheimer’s Disease (AD): Detection of Lipopolysaccharide (LPS) in AD Hippocampus. Front. Cell. Infect. Microbiol. 2017, 7, 318. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Zhou, J.; Zhou, Y.; Wang, H.; Si, N.; Ren, W.; Zhao, W.; Fan, X.; Gao, W.; Wei, X.; et al. Huanglian Jiedu Decoction Remodels the Periphery Microenvironment to Inhibit Alzheimer’s Disease Progression Based on the “Brain-Gut” Axis through Multiple Integrated Omics. Alzheimer’s Res. Ther. 2021, 13, 44. [Google Scholar] [CrossRef] [PubMed]
- O’Riordan, K.J.; Collins, M.K.; Moloney, G.M.; Knox, E.G.; Aburto, M.R.; Fülling, C.; Morley, S.J.; Clarke, G.; Schellekens, H.; Cryan, J.F. Short Chain Fatty Acids: Microbial Metabolites for Gut-Brain Axis Signalling. Mol. Cell. Endocrinol. 2022, 546, 111572. [Google Scholar] [CrossRef]
- Mullane, K.; Williams, M. Alzheimer’s Disease beyond Amyloid: Can the Repetitive Failures of Amyloid-Targeted Therapeutics Inform Future Approaches to Dementia Drug Discovery? Biochem. Pharmacol. 2020, 177, 113945. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet Rapidly and Reproducibly Alters the Human Gut Microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- Hampel, H.; Cummings, J.; Blennow, K.; Gao, P.; Jack, C.R.; Vergallo, A. Developing the ATX(N) Classification for Use across the Alzheimer Disease Continuum. Nat. Rev. Neurol. 2021, 17, 580–589. [Google Scholar] [CrossRef]
- Jack, C.R.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Feldman, H.H.; Frisoni, G.B.; Hampel, H.; Jagust, W.J.; Johnson, K.A.; Knopman, D.S.; et al. A/T/N: An Unbiased Descriptive Classification Scheme for Alzheimer Disease Biomarkers. Neurology 2016, 87, 539–547. [Google Scholar] [CrossRef]
- Soldan, A.; Pettigrew, C.; Fagan, A.M.; Schindler, S.E.; Moghekar, A.; Fowler, C.; Li, Q.-X.; Collins, S.J.; Carlsson, C.; Asthana, S.; et al. ATN Profiles among Cognitively Normal Individuals and Longitudinal Cognitive Outcomes. Neurology 2019, 92, e1567–e1579. [Google Scholar] [CrossRef]
- Vos, S.J.B.; Duara, R. The Prognostic Value of ATN Alzheimer Biomarker Profiles in Cognitively Normal Individuals. Neurology 2019, 92, 643–644. [Google Scholar] [CrossRef] [PubMed]
- Burnham, S.C.; Coloma, P.M.; Li, Q.-X.; Collins, S.; Savage, G.; Laws, S.; Doecke, J.; Maruff, P.; Martins, R.N.; Ames, D.; et al. Application of the NIA-AA Research Framework: Towards a Biological Definition of Alzheimer’s Disease Using Cerebrospinal Fluid Biomarkers in the AIBL Study. J. Prev. Alzheimer’s Dis. 2019, 6, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Bai, B.; Vanderwall, D.; Li, Y.; Wang, X.; Poudel, S.; Wang, H.; Dey, K.K.; Chen, P.-C.; Yang, K.; Peng, J. Proteomic Landscape of Alzheimer’s Disease: Novel Insights into Pathogenesis and Biomarker Discovery. Mol. Neurodegener. 2021, 16, 55. [Google Scholar] [CrossRef] [PubMed]
- Bagyinszky, E.; van Giau, V.; An, S.A. Transcriptomics in Alzheimer’s Disease: Aspects and Challenges. Int. J. Mol. Sci. 2020, 21, 3517. [Google Scholar] [CrossRef]
- Wang, H.; Dey, K.K.; Chen, P.-C.; Li, Y.; Niu, M.; Cho, J.-H.; Wang, X.; Bai, B.; Jiao, Y.; Chepyala, S.R.; et al. Integrated Analysis of Ultra-Deep Proteomes in Cortex, Cerebrospinal Fluid and Serum Reveals a Mitochondrial Signature in Alzheimer’s Disease. Mol. Neurodegener. 2020, 15, 43. [Google Scholar] [CrossRef]
- van der Schaar, J.; Visser, L.N.C.; Bouwman, F.H.; Ket, J.C.F.; Scheltens, P.; Bredenoord, A.L.; van der Flier, W.M. Considerations Regarding a Diagnosis of Alzheimer’s Disease before Dementia: A Systematic Review. Alzheimer’s Res. Ther. 2022, 14, 31. [Google Scholar] [CrossRef]
- Pernigoni, N.; Zagato, E.; Calcinotto, A.; Troiani, M.; Mestre, R.P.; Calì, B.; Attanasio, G.; Troisi, J.; Minini, M.; Mosole, S.; et al. Commensal Bacteria Promote Endocrine Resistance in Prostate Cancer through Androgen Biosynthesis. Science 2021, 374, 216–224. [Google Scholar] [CrossRef]
- Bian, X.; Liu, R.; Meng, Y.; Xing, D.; Xu, D.; Lu, Z. Lipid Metabolism and Cancer. J. Exp. Med. 2021, 218, e20201606. [Google Scholar] [CrossRef]
- Anastasiadou, E.; Jacob, L.S.; Slack, F.J. Non-Coding RNA Networks in Cancer. Nat. Rev. Cancer 2018, 18, 5–18. [Google Scholar] [CrossRef]
- Liu, N.; Wang, Z.-Z.; Zhao, M.; Zhang, Y.; Chen, N.-H. Role of Non-Coding RNA in the Pathogenesis of Depression. Gene 2020, 735, 144276. [Google Scholar] [CrossRef]
- Yang, L.; Chen, Y.; Liu, Y.; Xing, Y.; Miao, C.; Zhao, Y.; Chang, X.; Zhang, Q. The Role of Oxidative Stress and Natural Antioxidants in Ovarian Aging. Front. Pharmacol. 2021, 11, 617843. [Google Scholar] [CrossRef] [PubMed]
- Salim, S. Oxidative Stress and the Central Nervous System. J. Pharmacol. Exp. Ther. 2017, 360, 201–205. [Google Scholar] [CrossRef] [PubMed]
- D’Amelio, P.; Quacquarelli, L. Hypovitaminosis D and Aging: Is There a Role in Muscle and Brain Health? Nutrients 2020, 12, 628. [Google Scholar] [CrossRef] [PubMed]
- Milà-Alomà, M.; Suárez-Calvet, M.; Molinuevo, J.L. Latest Advances in Cerebrospinal Fluid and Blood Biomarkers of Alzheimer’s Disease. Ther. Adv. Neurol. Disord. 2019, 12, 1756286419888819. [Google Scholar] [CrossRef]
- Chaudhry, A.; Rizig, M. Comparing Fluid Biomarkers of Alzheimer’s Disease between African American or Black African and White Groups: A Systematic Review and Meta-Analysis. J. Neurol. Sci. 2021, 421, 117270. [Google Scholar] [CrossRef]
- Wang, H.-F.; Tan, L.; Hao, X.-K.; Jiang, T.; Tan, M.-S.; Liu, Y.; Zhang, D.-Q.; Yu, J.-T. Effect of EPHA1 Genetic Variation on Cerebrospinal Fluid and Neuroimaging Biomarkers in Healthy, Mild Cognitive Impairment and Alzheimer’s Disease Cohorts. J. Alzheimer’s Dis. 2015, 44, 115–123. [Google Scholar] [CrossRef]
- Cicognola, C.; Chiasserini, D.; Parnetti, L. Preanalytical Confounding Factors in the Analysis of Cerebrospinal Fluid Biomarkers for Alzheimer’s Disease: The Issue of Diurnal Variation. Front. Neurol. 2015, 6, 143. [Google Scholar] [CrossRef][Green Version]
- Mattsson-Carlgren, N.; Palmqvist, S.; Blennow, K.; Hansson, O. Increasing the Reproducibility of Fluid Biomarker Studies in Neurodegenerative Studies. Nat. Commun. 2020, 11, 6252. [Google Scholar] [CrossRef]
- Mattsson, N.; Blennow, K.; Zetterberg, H. Inter-Laboratory Variation in Cerebrospinal Fluid Biomarkers for Alzheimer’s Disease: United We Stand, Divided We Fall. Clin. Chem. Lab. Med. 2010, 48, 131. [Google Scholar] [CrossRef]
- Syaifullah, A.H.; Shiino, A.; Kitahara, H.; Ito, R.; Ishida, M.; Tanigaki, K. Machine Learning for Diagnosis of AD and Prediction of MCI Progression from Brain MRI Using Brain Anatomical Analysis Using Diffeomorphic Deformation. Front. Neurol. 2021, 11, 576029. [Google Scholar] [CrossRef]
- Castellazzi, G.; Cuzzoni, M.G.; Cotta Ramusino, M.; Martinelli, D.; Denaro, F.; Ricciardi, A.; Vitali, P.; Anzalone, N.; Bernini, S.; Palesi, F.; et al. A Machine Learning Approach for the Differential Diagnosis of Alzheimer and Vascular Dementia Fed by MRI Selected Features. Front. Neuroinform. 2020, 14, 25. [Google Scholar] [CrossRef] [PubMed]
- Sala, A.; Caprioglio, C.; Santangelo, R.; Vanoli, E.G.; Iannaccone, S.; Magnani, G.; Perani, D. Brain Metabolic Signatures across the Alzheimer’s Disease Spectrum. Eur. J. Nucl. Med. Mol. Imaging 2020, 47, 256–269. [Google Scholar] [CrossRef] [PubMed]


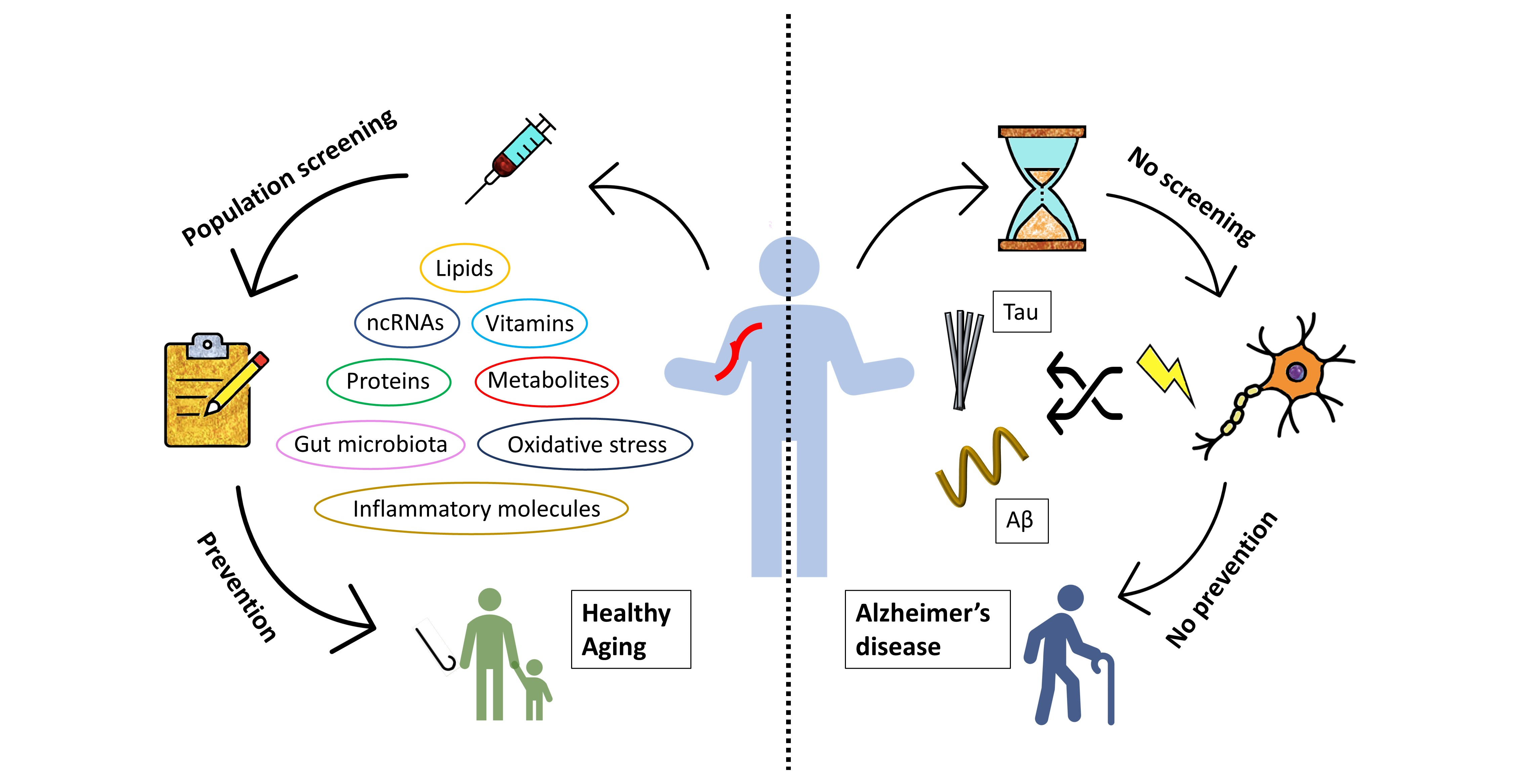{kind=link}
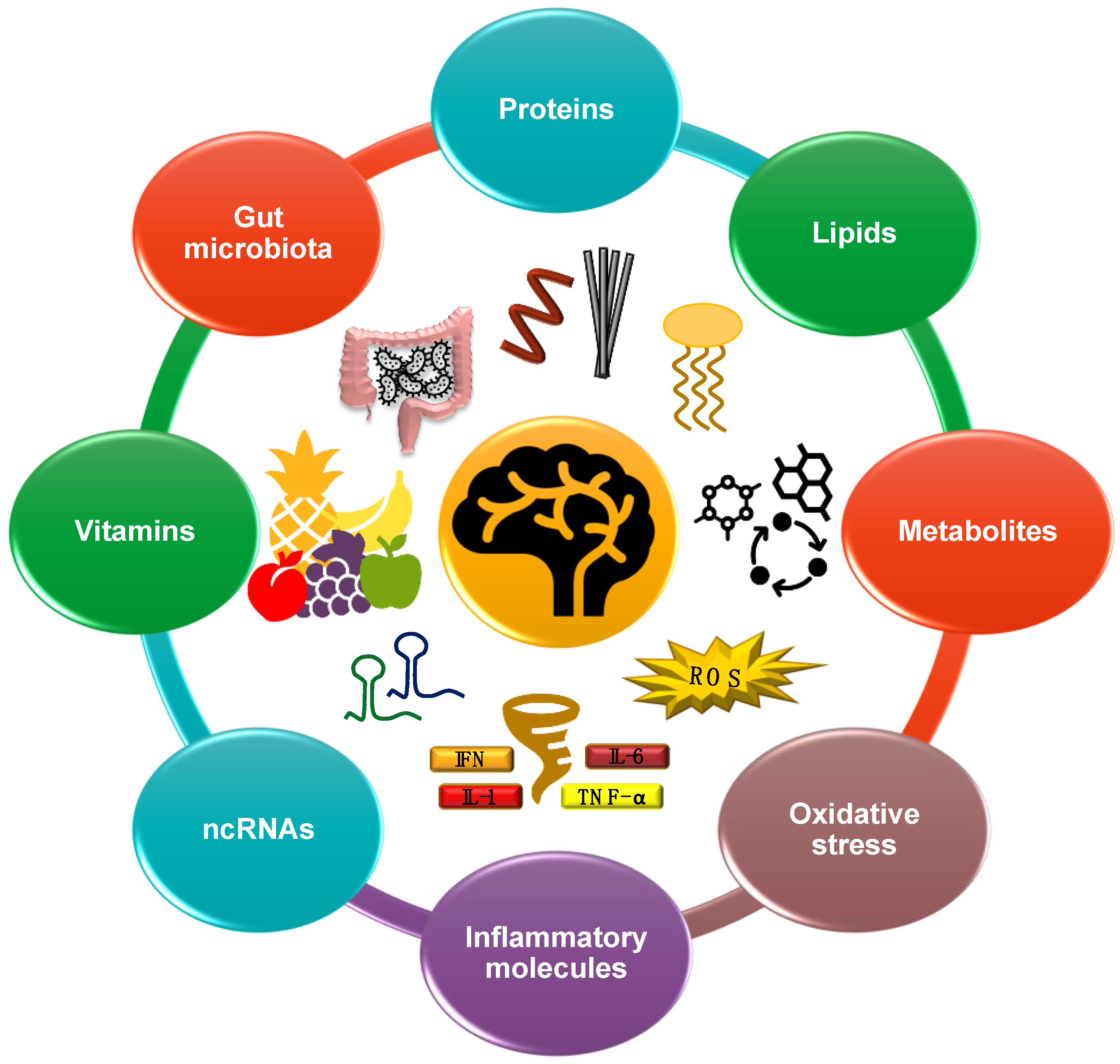{kind=link}
| Ref | Study Cohort and Design | Plasma/Serum | Measurement Methods | Results | Cohort of Variation | Biomarker/s Proposed |
|---|---|---|---|---|---|---|
| Han et al., 2021 [164] | Aβ1–42-treated PC12 cells, brain and hippocampus of APP/PS1 mouse, and the serum of AD patients | Serum | Probe 1 *, ELISA assay or LC–MS | ↑ Hcy ↓ Cys and GSH | AD vs. HC | Hcy, Cys, and GSH changes in the serum |
| Evlice et al., 2017 [165] | 30 AD (15 females and 15 males) and 10 HC (7 males and 3 females) | Serum | Activity and quantitative G6PD kit | ↑ serum G6PD | AD vs. HC | Serum G6PD levels |
| Peña-Bautista et al., 2021 [166] | 12 preclinical AD and 31 HC | Plasma | Chromatography and mass spectrometry | ↓ lipid peroxidation -15-F2t-IsoP correlates with p-tau -15-F2t-IsoP correlates with t-tau | AD vs. HC (non-significant) | Plasma isoprostanoids (combination of 10 biomarkers) |
| Zengi et al., 2012 [168] | 21 AD (10 men and 11 women) and 20 HC (11 men and 9 women) | Serum | PON1 activity absorbance assay | ↓ serum PON1 | AD vs. HC | Serum PON1 activity |
| López et al., 2013 [176] | 36 AD, 18 MCI, and 33 aged HC | Blood | ↑ Copper and MDA | AD and MCI vs. HC | Blood copper, MDA, and SOD | |
| Pradhan et al., 2022 [175] | 47 AD, 43 MCI, and 48 HC | Serum | SPR and Western blot | ↓ SIRT1, SIRT3, and SIRT6 | AD vs. MCI and HC | Serum SIRT1, SIRT3, and SIRT6 concentration |
| Cardoso et al., 2014 [170] | 27 AD, 17 MCI, and 28 HC | Plasma | Hydride generation atomic absorption spectroscopy | ↓ plasma Se ↓ erythrocyte Se | -AD vs. MCI and HC -AD and MCI vs. HC | Plasma Se levels |
| García et al., 2021 [10] | 20 MCI (13 males and 7 females), 20 AD (11 males and 9 females), and 15 PD (12 males and 3 females) and HC (age and sex matched). | Plasma | Electrochemical immunosensor | ↑ Unfolded p53 ↑ Unfolded p53 | -MCI, AD, and PD vs. HC -AD vs. MCI and PD | Plasma unfolded p53 |
| Peña-Bautista et al., 2021 [167] | 6 AD and 13 MCI | Plasma | LC–MS | ↑ dihomo-isoprostanes (17-epi-17-F2t-dihomo-IsoP, 17-F2t-dihomo-IsoP, Ent-7(RS)-7-F2t-dihomo-IsoP) and neuroprostanes (10-epi-10-F4t-NeuroP) | AD vs. MCI | Plasma isoprostanoids levels |
| Picco et al., 2014 [169] | 23 SCI, 28 MCI, and 34 mild AD | Plasma | Spectrophotometric analysis | ↓ eSOD activity ↓ CAT activity = GPx activity | -AD vs. SCI -AD vs. MCI and SCI | Plasma eSOD, CAT, and GPx activity combined with functional neuroimaging |
| Lin et al., 2021 [163] | 49 MCI and 16 HC | Plasma | Commercially available assay kit | ↓ plasma GSH | MCI vs. HC | Plasma GSH levels |
| Li et al., 2021 [173] | 839 HC | Serum | ↑ Serum uric acid = Serum uric acid in healthy individuals with or without tau pathology | Preclinical AD vs. HC | Serum uric acid | |
| Du et al., 2019 [172] | 113 aMCI and 832 HC | Serum | Commercial ELISA kit | ↑ Serum IMA and IMA/albumin | aMCI vs. HC | Serum IMA |
| Wu et al., 2021 [171] | 88 HC, 201 with cognitive impairment and no dementia (CIND) and 207 with dementia (160 AD and 47 vascular dementia) | Plasma | LC–MS/MS | ↓ plasma | Dementia vs. CIND and HC | Plasma ergothioneine levels |
| Ref | Study Cohort | Plasma/ Serum/Blood | Upregulated | Downregulated | Cohort of ncRNA Variation | Method |
|---|---|---|---|---|---|---|
| Dakterzada et al., 2021 [252] | Discovery cohort (n = 19, mild AD) and validation cohort (n = 53, mild AD) | Plasma | miR-342-5p | Severe AD | RT–PCR | |
| Poursaei et al., 2022 [213] | 50 AD and 50 HC | Plasma | hsa-let7d-5p hsa-let7g-5p | AD | RT–PCR | |
| Galimberti et al., 2014 [232] | 22 AD, 18 NINDCs, 8 NIDCs, and 10 FTD | Serum | miR-125b miR-23a miR-26b | AD | RT–PCR | |
| Kumar et al., 2017 [235] | Discovery cohort (10 AD, 6 MCI, and 14 HC) and validation cohort (11 AD, 20 MCI, and 18 HC) | Serum | miR-455-3p miR-4668-5p | AD | RT–PCR | |
| Yilmaz et al., 2016 [223] | 172 AD and 109 HC | Whole blood | hsa-miR-9-5p hsa-miR-106a-5p hsa-miR-106b-5p hsa-miR-107 | AD | RT–PCR | |
| Wang et al., 2020 [244] | 120 AD, 120 PD, and 120 HC | Plasma | miR-107 | miR-103 | AD | RT–PCR |
| Barbagallo et al., 2020 [259] | 30 AD, 30 PD, 24 VD, 25 VP, and 30 HC | Serum | miR-22 miR-23a miR-29a miR-125b | AD | RT–PCR | |
| Fotuhi et al., 2019 [258] | 45 AD and 36 HC | Whole plasma | lncRNA BACE1-AS | AD | RT–PCR | |
| Feng et al., 2018 [257] | 88 AD and 72 HC | Plasma | lncRNA BACE1 | AD | RT–PCR | |
| Yang et al., 2015 [222] | 30 AD and 30 HC | Blood | miR-29c | AD | RT–PCR | |
| Bhatnagar et al., 2014 [233] | 110 AD and 123 HC | Plasma | miR-34c | AD | RT–PCR | |
| Leidinger et al., 2013 [17] | 106 AD, 18 MCI, 16 CIS, 9 PD, 15 DEP, 15 BD, 14 SCHIZ, and 22 HC | Blood | hsa-miR-30d-5p | hsa-miR-144-5p | AD | NGS and RT–PCR |
| Zhu et al., 2015 [256] | 26 AD, 30 MCI, and 42 HC | Serum | miRNA-210 | AD | RT–PCR | |
| Kiko et al., 2014 [211] | Plasma | miR-34a miR-146a | AD | RT–PCR | ||
| Xing et al., 2016 [226] | 30 AD and 30 HC | Blood | miR-206 | AD | RT–PCR | |
| Wu et al., 2020 [227] | 40 AD (amyloid positive) and 31 controls (amyloid negative) | Blood | miR-146b-5p miR-15b-5p | AD | Small RNA sequencing | |
| Kumar et al., 2013 [210] | 11 AD, 9 MCI, and 20 HC | Plasma | hsa-miR-191-5p hsa-miR-15b-5p hsa-let-7d-5p hsa-let-7g-5p hsa-miR-142-3p | AD | nCounter miRNA expression assay v1 and RT–PCR | |
| Geekiyanage et al., 2012 [220] | 7 AD and 7 HC | Serum | miR-137 miR-181c miR-9 miR-29a/b | AD | RT–PCR | |
| Tan et al., 2014 [72] | 105 AD and 150 HC | Serum | miR-9 | miR-125b miR-181c | AD | RT–PCR |
| Sørensen et al., 2016 [212] | 10 AD and 10 VD/FTD or LBD | Plasma | miR-590-5p miR-142-5p | miR-194-5p | AD | RT–PCR |
| Ludwig et al., 2019 [225] | AD, MCI, HC, and ODN (total subjects 465) | Blood | miR-532-5p | AD | RT–PCR | |
| Liu et al., 2014 [247] | 32 MCI, 45 AD, and 50 HC | Serum | miR-384 | AD | RT–PCR | |
| Wang et al., 2019 [218] | 7 AD and 5 HC | Plasma | miR-200a-3p | AD | Microarray miRNA profile | |
| Liu et al., 2020 [224] | 50 AD, 20 VD, and 50 HC | Blood | miR-574-5p | hsa-circ-0003391 | AD | Microarray analysis |
| Hara et al., 2017 [234] | 27 AD and 18 HC | Serum | hsa-miR-501-3p hsa-let-7f-5p hsa-miR-26b-5p | AD | RT–PCR | |
| Jia et al., 2016 [231] | 84 AD and 62 HC | Serum | miR-519 | miR-29, miR-125b, miR-223 | AD | RT–PCR |
| Cosín-Tomás et al., 2017 [249] | HC, AD, PAD (n = 35 per group), and PD (n = 20) | Plasma | miR-34a-5p miR-545-3p | AD | RT–PCR | |
| Nagaraj et al., 2017 [248] | 15 MCI, 20 AD, and 15 HC | Plasma | miR-483-5p miR-486-5p miR-200a-3p miR-142-3P | miR-30b-5p | AD and MCI | RT–PCR |
| Dong et al., 2015 [242] | 127 AD, 30 MCI, and 30 VD | Serum | miR-93 miR-146a | miR-31 miR-93 miR-143 miR-146a | AD and MCI | Solexa sequencing and RT–PCR |
| Siedlecki-Wullich et al., 2019 [251] | 56 AD, 26 MCI, 38 HC, and 27 FTD | Plasma | miR-92a-3p miR-181c-5p miR-210-3p | AD and MCI | RT–PCR | |
| Sabry et al., 2020 [246] | 40 MCI and AD, and 20 HC | Plasma | miRNA-483-5p | AD and MCI | RT–PCR | |
| Zhang et al., 2021 [254] | 75 MCI and 52 HC | Serum | hsa-let-7g-5p hsa-miR-107 hsa-miR-186-3p | MCI | RT–PCR | |
| Shi et al., 2020 [253] | 71 aMCI and 69 HC | Serum | miR-34c | aMCI | RT–PCR | |
| He et al., 2021 [250] | Discovery cohort (n = 10), analysis cohort (n = 30), and validation cohort (n = 80) | Plasma | miR-1185-2-3p miR-1909-3p miR-22-5p miR-134-3p | aMCI | Microarray sequencing | |
| Wang et al., 2015 [255] | 97 AD, 116 aMCI, and 81 HC | Plasma | miR-107 | aMCI | RT–PCR |
| Ref | Study Cohort and Design | Analysis Performed | Results | Cohort of Variation | Biomarker/s Proposed |
|---|---|---|---|---|---|
| Glasø et al., 2004 [338] | AD (n = 20), HC (n = 18) | Analysis on serum and blood | ↓ Blood thiamine ↓ Blood TDP | AD | Vit B1 |
| dos Santos et al., 2020 [344] | AD (n = 60), HC (n = 60) | Complete blood count and Vit B12 levels assessment | ↓ Vit B12 | AD | Vit B12 |
| Lanyau-Domínguez et al., 2020 [351] | AD (n = 43), MCI (n = 131), HC (n = 250) | Spectrophotometry and high-resolution liquid chromatography on plasma | ↓ Vit A and vit C | AD | Combination of vit A and vit C |
| Gold et al., 1995 [339] | AD (n = 17), n-AD (n = 17) | Microbiologic assay on plasma and RBC | ↓ Plasma thiamine -No correlation between RBC thiamine and AD | AD | Vit B1 |
| Wang et al., 2018 [340] | AD (n = 90), HC (n = 90) | HPLC on whole blood samples | ↓ TDP | Female AD vs. male AD | TDP as protective factor for AD |
| D’Cunha et al., 2019 [341] | AD (n = 63), HC (n = 63) | ELISA kit to determine APOE4 on serum | ↓ Vit B2 dietary intake | AD without APOE4 genotype | Vit B2 and folate |
| Dursun et al., 2016 [346] | EOAD (n = 22), LOAD (n = 72), MCI (n = 32), HC (n = 70) | Chemiluminescent immunoassay on serum | ↓ 25(OH)D | LOAD ApoEε4 non-carriers | Vit D (in ApoEε4 allele non-carriers) |
| Ouma et al., 2018 [347] | AD (mild: n = 41, moderate: n = 35, severe: n = 32), MCI (n = 61), HC (n = 61) | Competitive radioimmunoassay on serum | ↓ 25(OH)D3 | MCI and AD | 25(OH)D3 |
| Blasko et al., 2021 [297] | Non-converting HC (n = 13), HC converting to MCI (n = 6), HC converting to AD (n = 6), MCI converting to AD (n = 8), MCI converting to HC (n = 8) and stable MCI (n = 7) | Competitive immunoassay on serum | ↓ Folate | MCI–AD converting pt | Folate |
| An et al., 2019 [342] | 2533 participants followed for an average of 2.3 y | Immunoassay on serum | ↑ Folate, vit B6, and vit B12 intake | Pt with better cognitive reserve | B vitamins and folate |
| Murdaca et al., 2021 [343] | AD (n = 108) | Machine learning approach to correlate blood vitamin levels with MMSE score | ↓ Vit D and folic acid | Pt with lower MMSE score | Combination of vit D and folic acid |
| Baldacci et al., 2020 [345] | SMC (n = 316) | Aβ-PET (n = 316, at baseline and 2 y follow-up). Lumbar puncture (n = 40 at baseline). Immunoassay on plasma (n = 79, at baseline, 1 y and 3 y follow-up) | ↓ Vit B12 | Pt with higher plasma total Tau levels | Vit B12 |
| de Leeuw et al., 2020 [188] | SCD (n = 149), MCI (n = 150). | Analysis on serum and plasma | ↑ 1,25(OH)2D3 | SCD | 1,25(OH)2D3 |
| Hooshmand et al., 2014 [348] | AD (n = 18), MCI (n = 28), SCI (n = 29) | Immunoassay on plasma, ELISA on CSF, MRI scans | ↑ 25(OH)D3 ↑ 25(OH)D3 | -Pt with higher CSF Aβ1–42 levels -Pt with greater brain volumes | Vit D |
| Al-Amin et al., 2019 [349] | MCI (n = 54) | Analysis on serum MRtrix and NBS on MRI scans | ↓ 25(OH)D3 | Pt with reduction in total hippocampal volume and connection deficit | Vit D |
| Raszewski et al., 2015 [350] | AD (n = 33), n-AD (n = 31) | HPLC on serum | ↓ Vit A and vit E | n-AD | Combination of vit A and vit E |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varesi, A.; Carrara, A.; Pires, V.G.; Floris, V.; Pierella, E.; Savioli, G.; Prasad, S.; Esposito, C.; Ricevuti, G.; Chirumbolo, S.; et al. Blood-Based Biomarkers for Alzheimer’s Disease Diagnosis and Progression: An Overview. Cells 2022, 11, 1367. https://doi.org/10.3390/cells11081367
Varesi A, Carrara A, Pires VG, Floris V, Pierella E, Savioli G, Prasad S, Esposito C, Ricevuti G, Chirumbolo S, et al. Blood-Based Biomarkers for Alzheimer’s Disease Diagnosis and Progression: An Overview. Cells. 2022; 11(8):1367. https://doi.org/10.3390/cells11081367
Chicago/Turabian StyleVaresi, Angelica, Adelaide Carrara, Vitor Gomes Pires, Valentina Floris, Elisa Pierella, Gabriele Savioli, Sakshi Prasad, Ciro Esposito, Giovanni Ricevuti, Salvatore Chirumbolo, and et al. 2022. "Blood-Based Biomarkers for Alzheimer’s Disease Diagnosis and Progression: An Overview" Cells 11, no. 8: 1367. https://doi.org/10.3390/cells11081367
APA StyleVaresi, A., Carrara, A., Pires, V. G., Floris, V., Pierella, E., Savioli, G., Prasad, S., Esposito, C., Ricevuti, G., Chirumbolo, S., & Pascale, A. (2022). Blood-Based Biomarkers for Alzheimer’s Disease Diagnosis and Progression: An Overview. Cells, 11(8), 1367. https://doi.org/10.3390/cells11081367