
Effects of Soy-Based Infant Formula on Weight Gain and Neurodevelopment in an Autism Mouse Model

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
3. Results
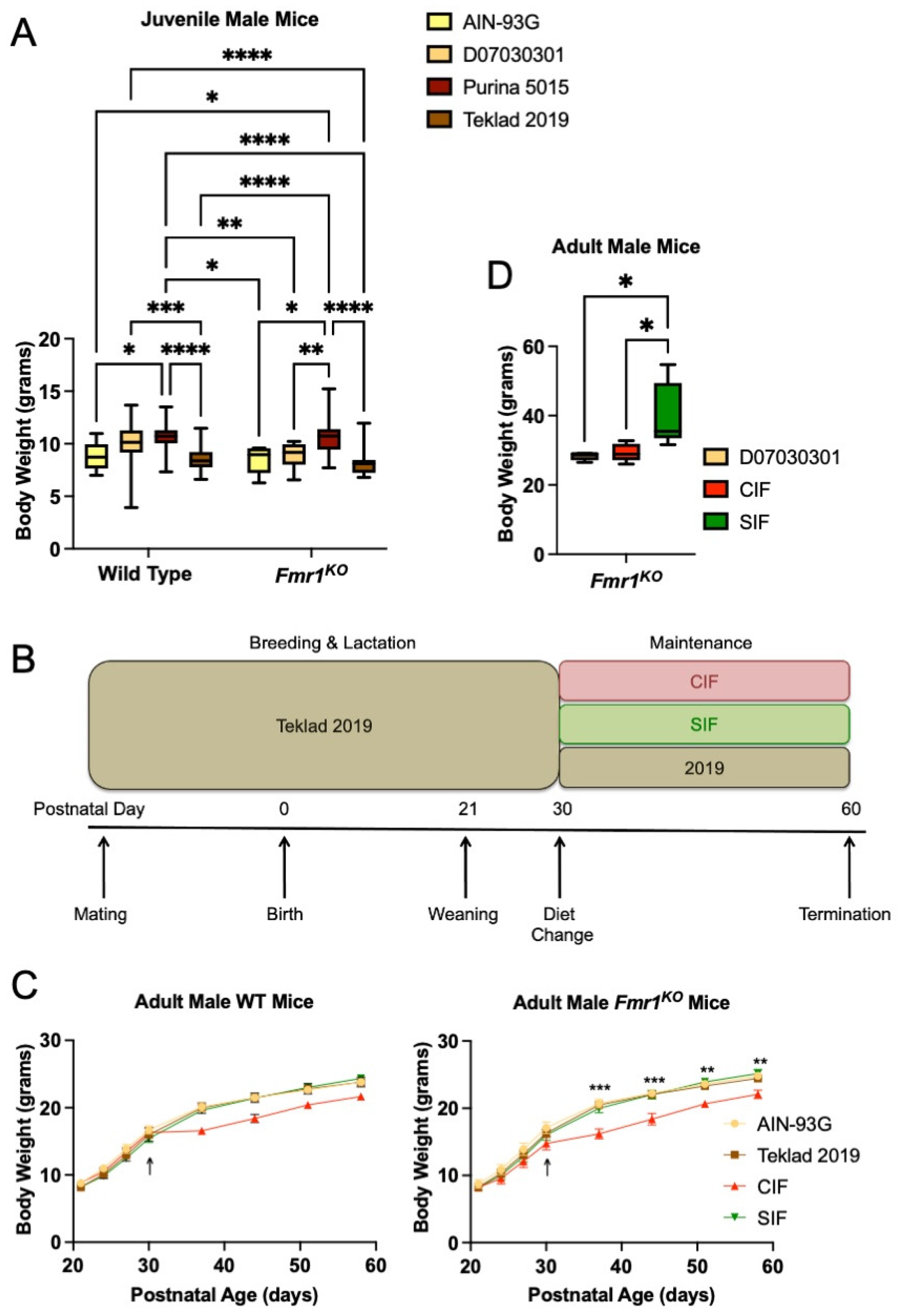
3.1. Soy/Grain-Based Rodent Diet Is Associated with Increased Weight Gain in Juvenile Fmr1KO Mice
3.2. Purina 5015 Does Not Statistically Alter Weight Gain in Adult Animals
3.3. SIF Increases Weight Gain in Adolescent Fmr1KO Mice Compared to CIF
3.4. CIF and SIF Are Suboptimal for Mouse Breeding
3.5. SIF Is Associated with Obesity in Adult Male Fmr1KO Mice
3.6. Consumption of SIF Is Associated with Altered Plasma Biomarker Expression
3.7. SIF Is Associated with Altered Behavior
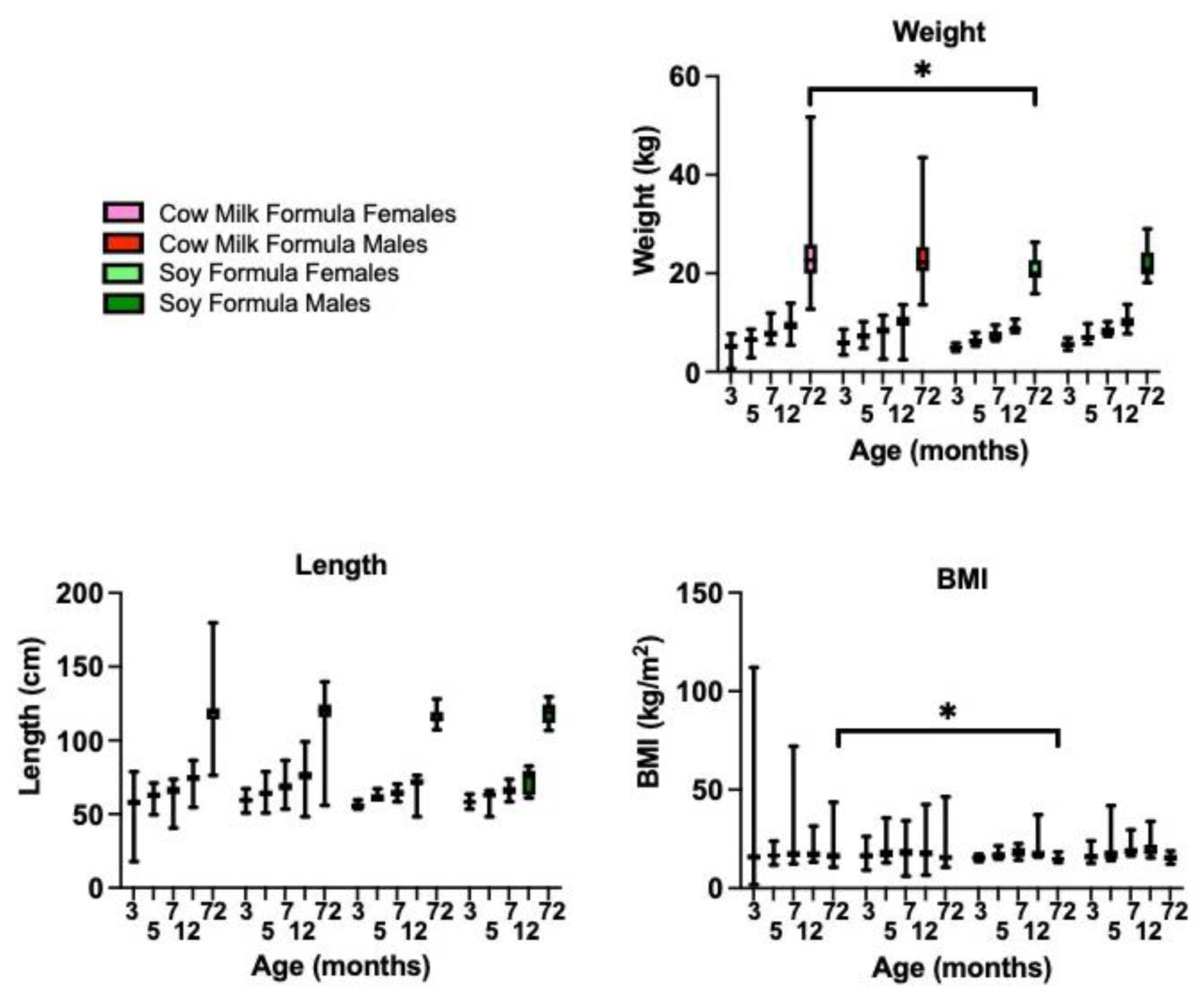
3.8. Soy-Based Infant Formula Is Not Associated with Obesity in Typically Developing Subjects at 6 Years of Age
3.9. Soy-Based Infant Formula Is Associated with an Increased Need for Support in School
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hales, C.M.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Prevalence of Obesity and Severe Obesity Among Adults: United States, 2017–2018; CDC National Center for Health Statistics: Hyattsville, MD, USA, 2020; pp. 1–8.
- Ogden, C.L.; Fryar, C.D.; Martin, C.B.; Freedman, D.S.; Carroll, M.D.; Gu, Q.; Hales, C.M. Trends in Obesity Prevalence by Race and Hispanic Origin—1999–2000 to 2017–2018. JAMA 2020, 324, 1208. [Google Scholar] [CrossRef] [PubMed]
- Ogden, C.L.; Carroll, M.D.; Kit, B.K.; Flegal, K.M. Prevalence of Childhood and Adult Obesity in the United States, 2011–2012. JAMA 2014, 311, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Rutter, M.; Le Couteur, A.; Lord, C. Autism Diagnostic Interview-Revised-WPS; Western Psychological Services: Los Angeles, CA, USA, 2003. [Google Scholar]
- Krakowiak, P.; Walker, C.K.; Bremer, A.A.; Baker, A.S.; Ozonoff, S.; Hansen, R.L.; Hertz-Picciotto, I. Maternal Metabolic Conditions and Risk for Autism and Other Neurodevelopmental Disorders. Pediatrics 2012, 129, e1121–e1128. [Google Scholar] [CrossRef] [PubMed]
- Heindel, J.J.; Saal, F.S.V. Role of nutrition and environmental endocrine disrupting chemicals during the perinatal period on the aetiology of obesity. Mol. Cell. Endocrinol. 2009, 304, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Bazar, K.A.; Yun, A.J.; Lee, P.Y.; Daniel, S.M.; Doux, J.D. Obesity and ADHD may represent different manifestations of a common environmental oversampling syndrome: A model for revealing mechanistic overlap among cognitive, metabolic, and inflammatory disorders. Med. Hypotheses 2006, 66, 263–269. [Google Scholar] [CrossRef]
- Nevison, C.; Parker, W. California Autism Prevalence by County and Race/Ethnicity: Declining Trends Among Wealthy Whites. J. Autism. Dev. Disord. 2020, 50, 4011–4021. [Google Scholar] [CrossRef]
- Hagerman, R.J.; Hagerman, P.J. Physical and Behavioral Phenotype; John Hopkins University Press: Baltimore, MD, USA, 2002. [Google Scholar]
- Verkerk, A.J.; Pieretti, M.; Sutcliffe, J.S.; Fu, Y.-H.; Kuhl, D.P.; Pizzuti, A.; Reiner, O.; Richards, S.; Victoria, M.F.; Zhang, F.; et al. Identification of a gene (FMR1) containing a CGG repeat coincident with a breakpoint cluster region exhibiting length variation in fragile X syndrome. Cell 1991, 65, 905–914. [Google Scholar] [CrossRef]
- Oberlé, I.; Rousseau, F.; Heitz, D.; Kretz, C.; Devys, D.; Hanauer, A.; Boué, J.; Bertheas, M.F.; Mandel, J.L. Instability of a 550-Base Pair DNA Segment and Abnormal Methylation in Fragile X Syndrome. Science 1991, 252, 1097–1102. [Google Scholar] [CrossRef]
- Davis, J.K.; Broadie, K. Multifarious Functions of the Fragile X Mental Retardation Protein. Trends Genet. 2017, 33, 703–714. [Google Scholar] [CrossRef]
- The Dutch-Belgian fragile X consortium; Bakker, C.E.; Verheij, C.; Willemsen, R.; van der Helm, R.; Oerlemans, F.; Vermey, M.; Bygrave, A.; Hoogeveen, A.T.; Oostra, B.A.; et al. Fmr1 knockout mice: A model to study fragile X mental retardation. Cell 1994, 78, 23–33. [Google Scholar]
- Loesch, D.Z.; Huggins, R.M.; Hoang, N.H. Growth in stature in fragile X families: A mixed longitudinal study. Am. J. Med. Genet. 1995, 58, 249–256. [Google Scholar] [CrossRef] [PubMed]
- De Vries, B.B.; Robinson, H.; Stolte-Dijkstra, I.; Tjon Pian Gi, C.V.; Dijkstra, P.F.; van Doorn, J.; Halley, D.J.J.; Oostra, B.A.; Turner, G.; Niermeijer, M.F. General overgrowth in the fragile X syndrome: Variability in the phenotypic expression of the FMR1 gene mutation. J. Med. Genet. 1995, 32, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Raspa, M.; Bailey, D.B.; Bishop, E.; Holiday, D.; Olmsted, M. Obesity, Food Selectivity, and Physical Activity in Individuals With Fragile X Syndrome. Am. J. Intellect. Dev. Disabil. 2010, 115, 482–495. [Google Scholar] [CrossRef] [PubMed]
- Westmark, C.J.; Westmark, P.R.; Malter, J.S. Soy-Based Diet Exacerbates Seizures in Mouse Models of Neurological Disease. J. Alzheimer’s Dis. 2013, 33, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Westmark, C.J. Soy Infant Formula and Seizures in Children with Autism: A Retrospective Study. PLoS ONE 2014, 9, e80488. [Google Scholar] [CrossRef]
- Westmark, C.J. Soy infant formula may be associated with increased autistic behaviors. Autism-Open Access 2013, 3, 20727. [Google Scholar]
- Westmark, C.J.; Kniss, C.; Sampene, E.; Wang, A.; Milunovich, A.; Elver, K.; Hessl, D.; Talboy, A.; Picker, J.; Haas-Givler, B.; et al. Soy-Based Infant Formula is Associated with an Increased Prevalence of Comorbidities in Fragile X Syndrome. Nutrients 2020, 12, 3136. [Google Scholar] [CrossRef]
- Westmark, C.J. Parental Reports on Early Autism Behaviors in Their Children with Fragile X Syndrome as a Function of Infant Feeding. Nutrients 2021, 13, 2888. [Google Scholar] [CrossRef]
- Westmark, C.J.; Malter, J.S. FMRP Mediates mGluR5-Dependent Translation of Amyloid Precursor Protein. PLoS Biol. 2007, 5, e52. [Google Scholar] [CrossRef]
- Jensen, M.N.; Ritskes-Hoitinga, M. How isoflavone levels in common rodent diets can interfere with the value of animal models and with experimental results. Lab. Anim. 2007, 41, 1–18. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 Purified Diets for Laboratory Rodents: Final Report of the American Institute of Nutrition Ad Hoc Writing Committee on the Reformulation of the AIN-76A Rodent Diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Westmark, P.R.; Gutierrez, A.; Gholston, A.K.; Wilmer, T.M.; Westmark, C.J. Preclinical testing of the ketogenic diet in fragile X mice. Neurochem. Int. 2020, 134, 104687. [Google Scholar] [CrossRef] [PubMed]
- Filon, M.J.; Wallace, E.; Wright, S.; Douglas, D.J.; Steinberg, L.I.; Verkuilen, C.L.; Westmark, P.R.; Maganti, R.K.; Westmark, C.J. Sleep and diurnal rest-activity rhythm disturbances in a mouse model of Alzheimer’s disease. Sleep 2020, 43, zsaa087. [Google Scholar] [CrossRef] [PubMed]
- Fein, S.B.; Li, R.; Chen, J.; Scanlon, K.S.; Grummer-Strawn, L.M. Methods for the Year 6 Follow-Up Study of Children in the Infant Feeding Practices Study II. Pediatrics 2014, 134 (Suppl. 1), S4–S12. [Google Scholar] [CrossRef]
- Grummer-Strawn, L.M.; Li, R.; Perrine, C.G.; Scanlon, K.S.; Fein, S.B. Infant Feeding and Long-Term Outcomes: Results from the Year 6 Follow-Up of Children in the Infant Feeding Practices Study II. Pediatrics 2014, 134 (Suppl. 1), S1–S3. [Google Scholar] [CrossRef]
- Menzies, C.; Naz, S.; Patten, D.; Alquier, T.; Bennett, B.M.; Lacoste, B. Distinct Basal Metabolism in Three Mouse Models of Neurodevelopmental Disorders. eNeuro 2021, 8, 1–14. [Google Scholar] [CrossRef]
- Sobik, S.; Sims, C.R.; McCorkle, G.; Bellando, J.; Sorensen, S.T.; Badger, T.M.; Casey, P.H.; Williams, D.K.; Andres, A. Early infant feeding effect on growth and body composition during the first 6 years and neurodevelopment at age 72 months. Pediatr. Res. 2021, 90, 140–147. [Google Scholar] [CrossRef]
- Gittner, L.S.; Ludington-Hoe, S.M.; Haller, H.S. Utilising infant growth to predict obesity status at 5 years. J. Paediatr. Child. Health 2013, 49, 564–574. [Google Scholar] [CrossRef]
- Gittner, L.S.; Ludington-Hoe, S.M.; Haller, H.S. Infant Obesity and Severe Obesity Growth Patterns in the First Two Years of Life. Matern. Child Health J. 2014, 18, 613–624. [Google Scholar] [CrossRef]
- Maenner, M.J.; Shaw, K.A.; Baio, J.; Washington, A.; Patrick, M.; DiRienzo, M.; Christensen, D.L.; Wiggins, D.L.; Pettygrove, S.; Andrews, J.G.; et al. Prevalence of autism spectrum disorder among children aged 8 years—autism and developmental disabilities monitoring network, 11 sites, United States. MMWR Surveill. Summ. 2020, 69, 1–12. [Google Scholar] [CrossRef]
- Blaxill, M.; Rogers, T.; Nevison, C. Autism Tsunami: The Impact of Rising Prevalence on the Societal Cost of Autism in the United States. J. Autism Dev. Disord. 2021, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.A.; Walker, M.; Morris, K.; Greig, I.; Mason, J.I.; Sharpe, R.M. Infant feeding with soy formula milk: Effects on puberty progression, reproductive function and testicular cell numbers in marmoset monkeys in adulthood. Hum. Reprod. 2006, 21, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, R.M.; Martin, B.; Morris, K.; Greig, I.; McKinnell, C.; McNeilly, A.S.; Walker, M. Infant feeding with soy formula milk: Effects on the testis and on blood testosterone levels in marmoset monkeys during the period of neonatal testicular activity. Hum. Reprod. 2002, 17, 1692–1703. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.; Karanam, M.; Robinson, E. Prevention of Cyclophosphamide-induced Accelerated Diabetes in the NOD Mouse by Nicotinamide or a Soy Protein-based Infant Formula. Int. J. Exp. Diabetes Res. 2001, 1, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Doerge, D.R.; Woodling, K.A.; Churchwell, M.I.; Fleck, S.C.; Helferich, W.G. Pharmacokinetics of isoflavones from soy infant formula in neonatal and adult rhesus monkeys. Food Chem. Toxicol. 2016, 92, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Westmark, C. Consumption of Breast Milk Is Associated with Decreased Prevalence of Autism in Fragile X Syndrome. Nutrients 2021, 13, 1785. [Google Scholar] [CrossRef]
- Ha, E.K.; Lee, S.W.; Kim, J.H.; Shim, S.; Kim, Y.H.; Song, J.Y.; Koh, H.Y.; Shin, Y.H.; Han, M.Y. Neurodevelopmental Outcomes in Infants Fed with Soy Formula: A Retrospective, National Population-Based Observational Cohort Study. J. Nutr. 2021, 151, 3045–3052. [Google Scholar] [CrossRef]
- Westmark, C.J. Increased incidence of epilepsy in response to soy-based infant formula in a national Korean cohort study. J. Nutr 2021, in press. [Google Scholar]
- Martin, C.R.; Ling, P.-R.; Blackburn, G.L. Review of infant feeding: Key features of breast milk and infant formula. Nutrients 2016, 8, 279. [Google Scholar] [CrossRef]
- Grummer-Strawn, L.M.; Scanlon, K.S.; Fein, S.B. Infant Feeding and Feeding Transitions During the First Year of Life. Pediatrics 2008, 122 (Suppl. 2), S36–S42. [Google Scholar] [CrossRef]
- Barrett, J.R. The science of soy: What do we really know? Environ. Health Perspect. 2006, 114, A352–A358. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, J.; Greer, F. American Academy of Pediatrics Committee on Nutrition. Use of soy protein-based formulas in infant feeding. Pediatrics 2008, 121, 1062–1068. [Google Scholar] [CrossRef] [PubMed]
- Merritt, R.J.; Jenks, B.H. Safety of Soy-Based Infant Formulas Containing Isoflavones: The Clinical Evidence. J. Nutr. 2004, 134, 1220S–1224S. [Google Scholar] [CrossRef] [PubMed]
- Rossen, L.M.; Simon, A.E.; Herrick, K.A. Types of Infant Formulas Consumed in the United States. Clin. Pediatr. 2015, 55, 278–285. [Google Scholar] [CrossRef]
- Westmark, C.J. Soy-Based Therapeutic Baby Formulas: Testable Hypotheses Regarding the Pros and Cons. Front. Nutr. 2017, 3, 59. [Google Scholar] [CrossRef] [PubMed]
- Andres, A.; Casey, P.H.; Cleves, M.A.; Badger, T.M. Body Fat and Bone Mineral Content of Infants Fed Breast Milk, Cow’s Milk Formula, or Soy Formula during the First Year of Life. J. Pediatr. 2013, 163, 49–54. [Google Scholar] [CrossRef]
- Andres, A.; Cleves, M.A.; Bellando, J.B.; Pivik, R.T.; Casey, P.H.; Badger, T.M. Developmental Status of 1-Year-Old Infants Fed Breast Milk, Cow’s Milk Formula, or Soy Formula. Pediatrics 2012, 129, 1134–1140. [Google Scholar] [CrossRef]
- Bellando, J.; McCorkle, G.; Spray, B.; Sims, C.R.; Badger, T.M.; Casey, P.H.; Scott, H.; Beall, S.R.; Sorensen, S.T.; Andres, A. Developmental assessments during the first 5 years of life in infants fed breast milk, cow’s milk formula, or soy formula. Food Sci. Nutr. 2020, 8, 3469–3478. [Google Scholar] [CrossRef]
- Strom, B.L.; Schinnar, R.; Ziegler, E.E.; Barnhart, K.T.; Sammel, M.D.; Macones, G.A.; Stallings, V.A.; Drulis, J.M.; Nelson, S.E.; Hanson, S.A. Exposure to Soy-Based Formula in Infancy and Endocrinological and Reproductive Outcomes in Young Adulthood. JAMA 2001, 286, 807–814. [Google Scholar] [CrossRef]
- Li, T.; Badger, T.; Bellando, B.; Sorensen, S.; Lou, X.; Ou, X. Brain Cortical Structure and Executive Function in Children May Be Influenced by Parental Choices of Infant Diets. Am. J. Neuroradiol. 2020, 41, 1302–1308. [Google Scholar] [CrossRef]
- Young, B. Variation in Infant Formula Macronutrient Ingredients Is Associated with Infant Anthropometrics. Nutrients 2020, 12, 3465. [Google Scholar] [CrossRef] [PubMed]
- Berry-Kravis, E.; Hessl, D.; Abbeduto, L.; Reiss, A.L.; Beckel-Mitchener, A.; Urv, T.K. Outcome Measures for Clinical Trials in Fragile X Syndrome. J. Dev. Behav. Pediatr. 2013, 34, 508–522. [Google Scholar] [CrossRef] [PubMed]
- Westmark, C.J. The quest for fragile X biomarkers. Mol. Cell. Pediatr. 2014, 1, 1. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Folch, J.; Patraca, I.; Martinez, N.; Pedros, I.; Petrov, D.; Ettcheto, M.; Abad, S.; Marin, M.; Beas-Zarate, C.; Camins, A. The role of leptin in the sporadic form of Alzheimer’s disease. interactions with the adipokines amylin, ghrelin and the pituitary hormone prolactin. Life Sci. 2015, 140, 19–28. [Google Scholar] [CrossRef]
- Maffei, M.; Halaas, J.; Ravussin, E.; Pratley, R.E.; Lee, G.H.; Zhang, Y.; Fei, H.; Kim, S.; Lallone, R.; Ranganathan, S.; et al. Leptin levels in human and rodent: Measurement of plasma leptin and ob RNA in obese and weight-reduced subjects. Nat. Med. 1995, 1, 1155–1161. [Google Scholar] [CrossRef]
- Friedman, J.M.; Halaas, J.L. Leptin and the regulation of body weight in mammals. Nature 1998, 395, 763–770. [Google Scholar] [CrossRef]
- Giordano, E.; Dávalos, A.; Crespo, M.C.; Tomé-Carneiro, J.; Gómez-Coronado, D.; Visioli, F. Soy Isoflavones in Nutritionally Relevant Amounts Have Varied Nutrigenomic Effects on Adipose Tissue. Molecules 2015, 20, 2310–2322. [Google Scholar] [CrossRef]
- Lephart, E.D.; Setchell, K.D.R.; Handa, R.J.; Lund, T.D. Behavioral effects of endocrine-disrupting substances: Phytoestrogens. ILAR J. 2004, 45, 443–454. [Google Scholar] [CrossRef]
- Leboucher, A.; Pisani, D.F.; Martinez-Gili, L.; Chilloux, J.; Bermudez-Martin, P.; Van Dijck, A.; Ganief, T.; Macek, B.; Becker, J.A.; Le Merrer, J.; et al. The translational regulator FMRP controls lipid and glucose metabolism in mice and humans. Mol. Metab. 2019, 21, 22–35. [Google Scholar] [CrossRef]
- Lumaban, J.G.; Nelson, D.L. The Fragile X proteins Fmrp and Fxr2p cooperate to regulate glucose metabolism in mice. Hum. Mol. Genet. 2015, 24, 2175–2184. [Google Scholar] [CrossRef]
- Lisik, M.Z.; Gutmajster, E.; Sieroń, A.L. Plasma Levels of Leptin and Adiponectin in Fragile X Syndrome. Neuroimmunomodulation 2016, 23, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Tanida, M.; Ono, Y.; Kasahara, R.; Fujii, Y.; Ohora, K.; Suzuki, K.; Sobue, K. Leptin inhibits amyloid beta-protein degradation through decrease of neprilysin expression in primary cultured astrocytes. Biochem. Biophys. Res. Commun. 2014, 445, 214–217. [Google Scholar] [CrossRef] [PubMed]
- Nalivaeva, N.N.; Zhuravin, I.A.; Turner, A.J. Neprilysin expression and functions in development, ageing and disease. Mech. Ageing Dev. 2020, 192, 111363. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.; Siems, W.-E.; Kluge, R.; Gembardt, F.; Schultheiss, H.-P.; Schirner, M.; Walther, T. New Function for an Old Enzyme: NEP Deficient Mice Develop Late-Onset Obesity. PLoS ONE 2010, 5, e12793. [Google Scholar] [CrossRef] [PubMed]
- Standeven, K.F.; Hess, K.; Carter, A.M.; Rice, G.I.; Cordell, P.A.; Balmforth, A.J.; Scott, D.J.; Turner, A.J.; Hooper, N.M.; Grant, P.J.; et al. Neprilysin, obesity and the metabolic syndrome. Int. J. Obes. 2011, 35, 1031–1040. [Google Scholar] [CrossRef]
- Czeczor, J.K.; McGee, S.L. Emerging roles for the amyloid precursor protein and derived peptides in the regulation of cellular and systemic metabolism. J. Neuroendocr. 2017, 29, 1–8. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, Q.; Chen, S.; Xu, C. Functions of amyloid precursor protein in metabolic diseases. Metabolism 2021, 115, 154454. [Google Scholar] [CrossRef]
- Clarke, J.R.; Lyra, E.; Silva, N.M.; Figueiredo, C.P.; Frozza, R.L.; Ledo, J.H.; Beckman, D.; Katashima, C.K.; Razolli, D.; Carvalho, B.M.; et al. Alzheimer-associated Abeta oligomers impact the central nervous system to induce peripheral metabolic deregulation. EMBO Mol. Med. 2015, 7, 190–210. [Google Scholar] [CrossRef]
- Westmark, C.J.; Sokol, D.K.; Maloney, B.; Lahiri, D.K. Novel roles of amyloid-beta precursor protein metabolites in fragile X syndrome and autism. Mol. Psychiatry 2016, 21, 1333–1341. [Google Scholar] [CrossRef]
- Westmark, C.J. Fragile X and APP: A Decade in Review, a Vision for the Future. Mol. Neurobiol. 2019, 56, 3904–3921. [Google Scholar] [CrossRef]
- Çaku, A.; Seidah, N.G.; Lortie, A.; Gagné, N.; Perron, P.; Dubé, J.; Corbin, F. New insights of altered lipid profile in Fragile X Syndrome. PLoS ONE 2017, 12, e0174301. [Google Scholar] [CrossRef]
- Westmark, C.J. A Hypothesis Regarding the Molecular Mechanism Underlying Dietary Soy-Induced Effects on Seizure Propensity. Front. Neurol. 2014, 5, 169. [Google Scholar] [CrossRef] [PubMed]
- Marwarha, G.; Raza, S.; Meiers, C.; Ghribi, O. Leptin attenuates BACE1 expression and amyloid-beta genesis via the activation of SIRT1 signaling pathway. Biochim. Biophys. Acta 2014, 1842, 1587–1595. [Google Scholar] [CrossRef] [PubMed]
- Meakin, P.J.; Jalicy, S.M.; Montagut, G.; Allsop, D.J.P.; Cavellini, D.L.; Irvine, S.W.; McGinley, C.; Liddell, M.K.; McNeilly, A.; Parmionova, K.; et al. Bace1-dependent amyloid processing regulates hypothalamic leptin sensitivity in obese mice. Sci. Rep. 2018, 8, 55. [Google Scholar] [CrossRef]
- Kim, J.; Han, D.; Byun, S.-H.; Kwon, M.; Cho, S.-J.; Koh, Y.H.; Yoon, K. Neprilysin facilitates adipogenesis through potentiation of the phosphatidylinositol 3-kinase (PI3K) signaling pathway. Mol. Cell. Biochem. 2017, 430, 1–9. [Google Scholar] [CrossRef]
- Liu, J.Y.; Sun, M.Y.; Sommerville, N.; Ngan, M.P.; Ponomarev, E.; Lin, G.; Rudd, J.A. Soy flavonoids prevent cognitive deficits induced by intra-gastrointestinal administration of beta-amyloid. Food Chem. Toxicol. 2020, 141, 111396. [Google Scholar] [CrossRef]
- Tong, J.Q.; Zhang, J.; Hao, M.; Yang, J.; Han, Y.F.; Liu, X.J.; Shi, H.; Wu, M.-n.; Liu, Q.-s.; Qi, J.-s. Leptin attenuates the detrimental effects of beta-amyloid on spatial memory and hippocampal later-phase long term potentiation in rats. Horm. Behav. 2015, 73, 125–130. [Google Scholar] [CrossRef]
- Cirillo, D.; Agostini, F.; Klus, P.; Marchese, D.; Rodriguez, S.; Bolognesi, B.; Tartaglia, G.G. Neurodegenerative diseases: Quantitative predictions of protein–RNA interactions. RNA 2012, 19, 129–140. [Google Scholar] [CrossRef]
- Riley, C.; Wheeler, A. Assessing the Fragile X Syndrome Newborn Screening Landscape. Pediatrics 2017, 139, S207–S215. [Google Scholar] [CrossRef]
- Marahatta, A.; Bhandary, B.; Jeong, S.-K.; Kim, H.-R.; Chae, H.-J. Soybean greatly reduces valproic acid plasma concentrations: A food–drug interaction study. Sci. Rep. 2014, 4, 4362. [Google Scholar] [CrossRef]
- Sankhe, R.; Rathi, E.; Manandhar, S.; Kumar, A.; Pai, S.R.K.; Kini, S.G.; Kishore, A. Repurposing of existing FDA approved drugs for Neprilysin inhibition: An in-silico study. J. Mol. Struct. 2021, 1224, 129073. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Target | Fold Change | Post Hoc t-Test (p) |
|---|---|---|
| 6Ckine | 0.45 1 | ≤0.05 |
| Axl | 2.02 1 | ≤0.02 |
| 1.76 2 | ≤0.05 | |
| B7-1 | 1.48 1 | ≤0.05 |
| 2.04 2 | ≤0.00 | |
| 0.73 3 | ≤0.05 | |
| CD30 | 1.35 1 | ≤0.02 |
| CD36 | 2.40 1 | ≤0.00 |
| 2.37 3 | ≤0.05 | |
| Epiregulin | 1.54 1 | ≤0.05 |
| Galactin-7 | 0.44 1 | ≤0.04 |
| ICAM-1 | 6.08 1 | ≤0.04 |
| 3.68 3 | ≤0.05 | |
| IL-1 R4 | 0.12 2 | ≤0.01 |
| IL-2 Ra | 1.43 1 | ≤0.00 |
| Leptin | 5.68 2 | ≤0.02 |
| 0.11 3 | ≤0.02 | |
| MFG-E8 | 4.34 3 | ≤0.00 |
| MMP-10 | 4.23 1 | ≤0.01 |
| 2.07 2 | ≤0.00 | |
| 2.04 3 | ≤0.04 | |
| Neprilysin | 4.38 2 | ≤0.03 |
| 0.25 3 | ≤0.05 |
| Protein Target | Fold Change | Post Hoc t-Test (p) |
|---|---|---|
| B7-1 | 2.18 2 | ≤0.01 |
| CXCL 16 | 0.77 2 | ≤0.04 |
| Epiregulin | 1.56 1 | ≤0.04 |
| IGFBP-5 | 0.63 1 | ≤0.04 |
| 0.73 3 | ≤0.05 | |
| IL-1 R4 | 0.17 1 | ≤0.02 |
| Lipocalin-2 | 8.73 2 | ≤0.01 |
| Neprilysin | 2.45 2 | ≤0.05 |
| P-Cadherin | 2.11 2 | ≤0.00 |
| Prolactin | 3.60 1 | ≤0.00 |
| Renin 1 | 2.28 1 | ≤0.03 |
| VEGF-B | 4.42 1 | ≤0.05 |
| Protein Target | Fold Change | Post Hoc t-Test (p) |
|---|---|---|
| 6Ckine | 3.27 1 | ≤0.02 |
| B7-1 | 1.87 1 | ≤0.02 |
| 1.75 3 | ≤0.01 | |
| BLC | 2.08 1 | ≤0.03 |
| Galactin-7 | 2.96 1 | ≤0.00 |
| ICAM-1 | 3.71 2 | ≤0.05 |
| MFG-E8 | 4.77 2 | ≤0.00 |
| Prolactin | 0.34 2 | ≤0.02 |
| TRANCE | 2.58 1 | ≤0.02 |
| VEGF-B | 4.12 1 | ≤0.02 |
| Metric | Breast | Cow Milk Formula | Soy Formula | p1 |
|---|---|---|---|---|
| N | 419 | 455 | 36 | |
| IEP | 10% | 15% | 17% | 0.093 |
| Speech Therapy | 9.3% | 15% | 17% | 0.037 2 |
| Occupational Therapy | 3.3% | 4.8% | 8.3% | 0.26 |
| Help in School | 5.5% | 11% | 17% | 0.0035 3 |
| Support in Classroom | 2.6% | 2.2% | 14% | 0.0003 4 |
| Hay Fever or Respiratory Allergy | 18% | 25% | 31% | 0.024 5 |
| Asthma | 8.1% | 13% | 14% | 0.07 |
| ADD or ADHD | 1.2% | 5.1% | 0% | 0.0024 6 |
| Autism or Developmental Delay | 1.9% | 3.5% | 5.6% | 0.22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Westmark, C.J.; Filon, M.J.; Maina, P.; Steinberg, L.I.; Ikonomidou, C.; Westmark, P.R. Effects of Soy-Based Infant Formula on Weight Gain and Neurodevelopment in an Autism Mouse Model. Cells 2022, 11, 1350. https://doi.org/10.3390/cells11081350
Westmark CJ, Filon MJ, Maina P, Steinberg LI, Ikonomidou C, Westmark PR. Effects of Soy-Based Infant Formula on Weight Gain and Neurodevelopment in an Autism Mouse Model. Cells. 2022; 11(8):1350. https://doi.org/10.3390/cells11081350
Chicago/Turabian StyleWestmark, Cara J., Mikolaj J. Filon, Patricia Maina, Lauren I. Steinberg, Chrysanthy Ikonomidou, and Pamela R. Westmark. 2022. "Effects of Soy-Based Infant Formula on Weight Gain and Neurodevelopment in an Autism Mouse Model" Cells 11, no. 8: 1350. https://doi.org/10.3390/cells11081350
APA StyleWestmark, C. J., Filon, M. J., Maina, P., Steinberg, L. I., Ikonomidou, C., & Westmark, P. R. (2022). Effects of Soy-Based Infant Formula on Weight Gain and Neurodevelopment in an Autism Mouse Model. Cells, 11(8), 1350. https://doi.org/10.3390/cells11081350