
Ubiquitin-Proteasome System–Regulated Protein Degradation in Spermatogenesis


Abstract
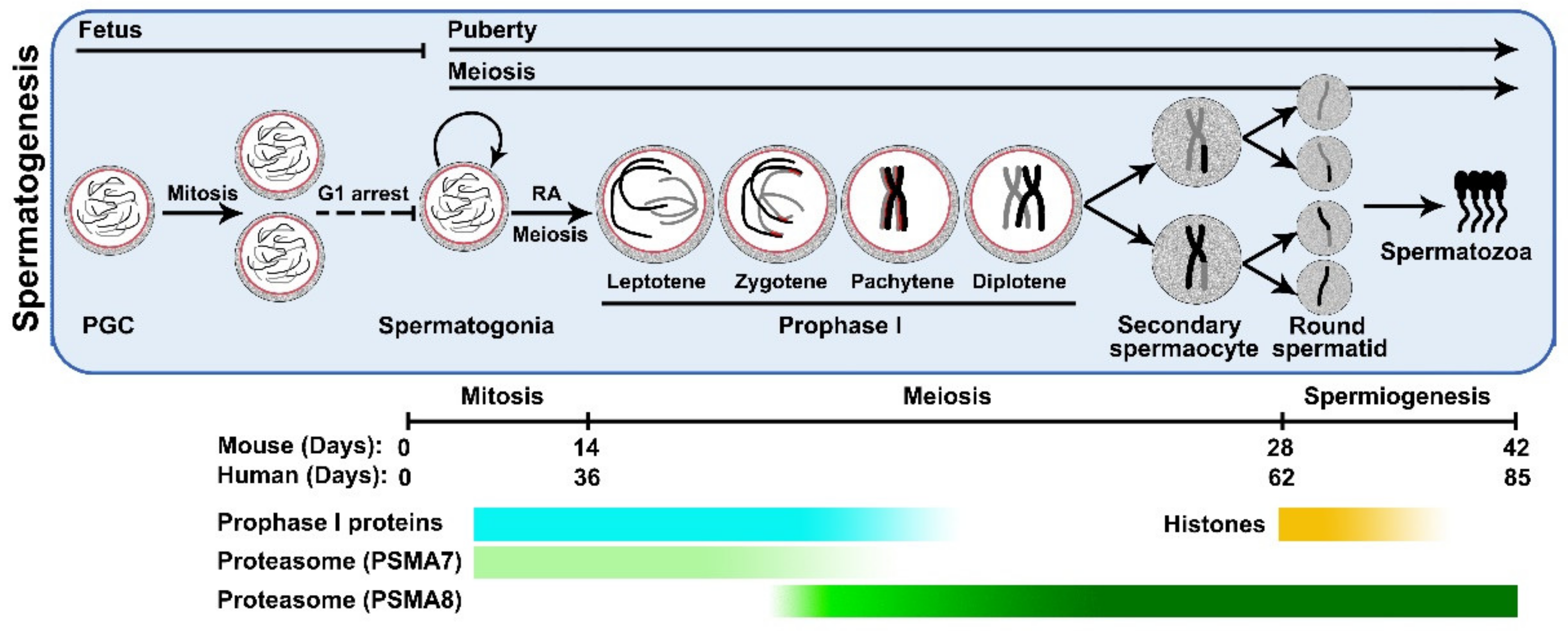
1. Introduction
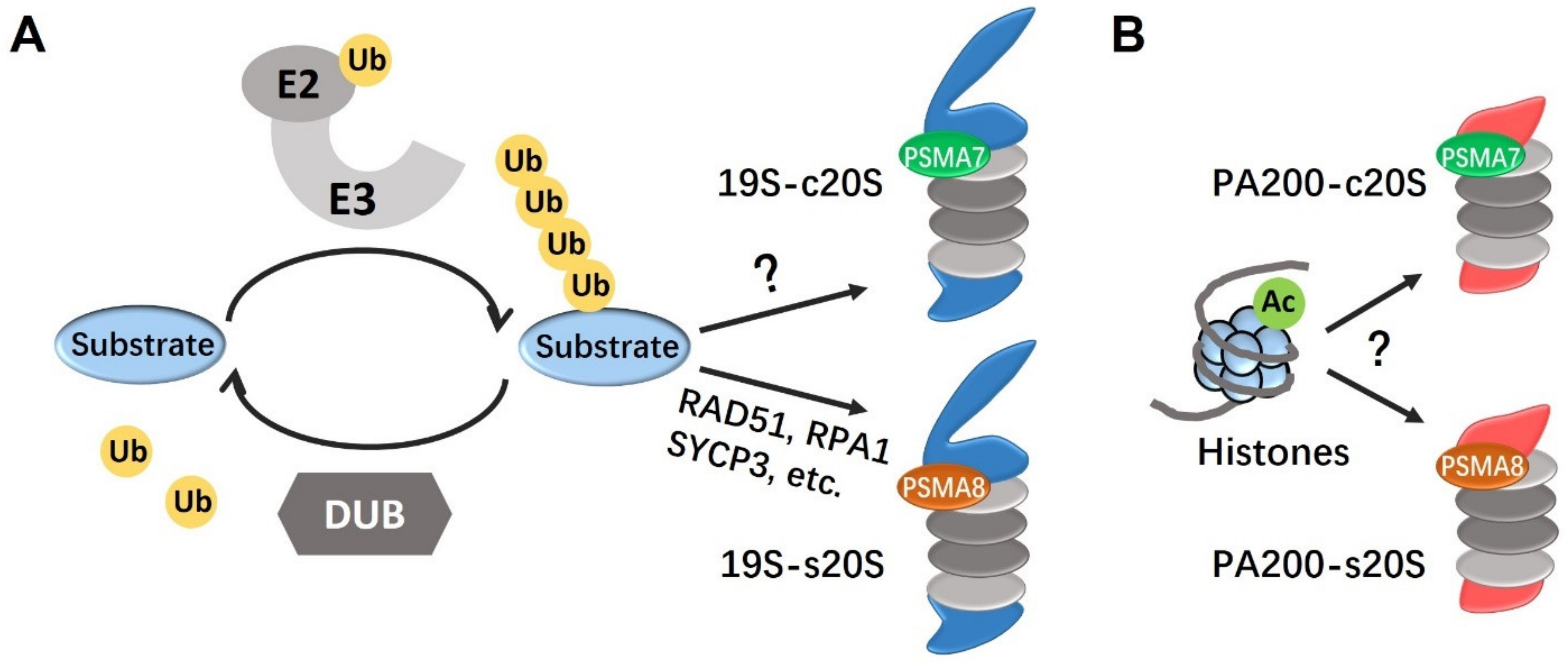
2. The Ubiquitination-Proteasome System
3. Proteasomes in Male Germ Cells
3.1. PSMA8
3.2. PA200
4. E3 Ubiquitin Ligases in Spermatogenesis
4.1. Cullin-Ring Ligase Family
4.2. Other E3 Ubiquitin Ligases
5. Protein Deubiquitination in Spermatogenesis
6. Conclusions and Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hess, R.A.; de Franca, L.R. Spermatogenesis and cycle of the seminiferous epithelium. Adv. Exp. Med. Biol. 2008, 636, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Oatley, J.M.; Brinster, R.L. The germline stem cell niche unit in mammalian testes. Physiol. Rev. 2012, 92, 577–595. [Google Scholar] [CrossRef] [PubMed]
- Hunter, N. Meiotic recombination: The essence of heredity. Cold Spring Harb. Perspect. Biol. 2015, 7, a016618. [Google Scholar] [CrossRef] [PubMed]
- Adler, I.D. Comparison of the duration of spermatogenesis between male rodents and humans. Mutat. Res. 1996, 352, 169–172. [Google Scholar] [CrossRef]
- Marchetti, F.; Wyrobek, A.J. Mechanisms and consequences of paternally-transmitted chromosomal abnormalities. Birth Defects Res. Part. C Embryo Today Rev. 2005, 75, 112–129. [Google Scholar] [CrossRef]
- Griswold, M.D. Spermatogenesis: The commitment to meiosis. Physiol. Rev. 2016, 96, 1–17. [Google Scholar] [CrossRef]
- Chen, Y.; Zheng, Y.; Gao, Y.; Lin, Z.; Yang, S.; Wang, T.; Wang, Q.; Xie, N.; Hua, R.; Liu, M.; et al. Single-cell RNA-seq uncovers dynamic processes and critical regulators in mouse spermatogenesis. Cell Res. 2018, 28, 879–896. [Google Scholar] [CrossRef]
- Soumillon, M.; Necsulea, A.; Weier, M.; Brawand, D.; Zhang, X.; Gu, H.; Barthès, P.; Kokkinaki, M.; Nef, S.; Gnirke, A.; et al. Cellular source and mechanisms of high transcriptome complexity in the mammalian testis. Cell Rep. 2013, 3, 2179–2190. [Google Scholar] [CrossRef]
- Glickman, M.H.; Ciechanover, A. The ubiquitin-proteasome proteolytic pathway: Destruction for the sake of construction. Physiol. Rev. 2002, 82, 373–428. [Google Scholar] [CrossRef]
- Sadowski, M.; Suryadinata, R.; Tan, A.R.; Roesley, S.N.A.; Sarcevic, B. Protein monoubiquitination and polyubiquitination generate structural diversity to control distinct biological processes. IUBMB Life 2012, 64, 136–142. [Google Scholar] [CrossRef]
- Hochstrasser, M. Ubiquitin-dependent protein degradation. Annu. Rev. Genet. 1996, 30, 405–439. [Google Scholar] [CrossRef]
- Nijman, S.M.B.; Luna-Vargas, M.P.A.; Velds, A.; Brummelkamp, T.R.; Dirac, A.M.G.; Sixma, T.K.; Bernards, R. A genomic and functional inventory of deubiquitinating enzymes. Cell 2005, 123, 773–786. [Google Scholar] [CrossRef]
- Tanaka, K. The proteasome: Overview of structure and functions. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 12–36. [Google Scholar] [CrossRef]
- Huang, X.; Luan, B.; Wu, J.; Shi, Y. An atomic structure of the human 26S proteasome. Nat. Struct. Mol. Biol. 2016, 23, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Groll, M.; Ditzel, L.; Löwe, J.; Stock, D.; Bochtler, M.; Bartunik, H.D.; Huber, R. Structure of 20S proteasome from yeast at 2.4 Å resolution. Nature 1997, 386, 463–471. [Google Scholar] [CrossRef]
- Tanaka, K.; Kasahara, M. The MHC class I ligand-generating system: Roles of immunoproteasomes and the interferon-gamma-inducible proteasome activator PA28. Immunol. Rev. 1998, 163, 161–176. [Google Scholar] [CrossRef]
- Zhang, Q.; Ji, S.-Y.; Busayavalasa, K.; Shao, J.; Yu, C. Meiosis I progression in spermatogenesis requires a type of testis-specific 20S core proteasome. Nat. Commun. 2019, 10, 3387. [Google Scholar] [CrossRef]
- Gómez-H, L.; Felipe-Medina, N.; Condezo, Y.B.; Garcia-Valiente, R.; Ramos, I.; Suja, J.A.; Barbero, J.L.; Roig, I.; Sánchez-Martín, M.; de Rooij, D.G.; et al. The PSMA8 subunit of the spermatoproteasome is essential for proper meiotic exit and mouse fertility. PLoS Genet. 2019, 15, e1008316. [Google Scholar] [CrossRef]
- Zhang, Z.-H.; Jiang, T.-X.; Chen, L.-B.; Zhou, W.; Liu, Y.; Gao, F.; Qiu, X.-B. Proteasome subunit alpha4s is essential for formation of spermatoproteasomes and histone degradation during meiotic DNA repair in spermatocytes. J. Biol. Chem. 2021, 296, 100130. [Google Scholar] [CrossRef]
- Uechi, H.; Hamazaki, J.; Murata, S. Characterization of the testis-specific proteasome subunit alpha4s in mammals. J. Biol. Chem. 2014, 289, 12365–12374. [Google Scholar] [CrossRef]
- Asher, G.; Reuven, N.; Shaul, Y. 20S proteasomes and protein degradation “by default”. Bioessays 2006, 28, 844–849. [Google Scholar] [CrossRef]
- Tenzer, S.; Stoltze, L.; Schönfisch, B.; Dengjel, J.; Müller, M.; Stevanović, S.; Rammensee, H.-G.; Schild, H. Quantitative analysis of prion-protein degradation by constitutive and immuno-20S proteasomes indicates differences correlated with disease susceptibility. J. Immunol. 2004, 172, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Coux, O.; Zieba, B.A.; Meiners, S. The proteasome system in health and disease. Adv. Exp. Med. Biol. 2020, 1233, 55–100. [Google Scholar] [CrossRef] [PubMed]
- Preckel, T.; Fung-Leung, W.P.; Cai, Z.; Vitiello, A.; Salter-Cid, L.; Winqvist, O.; Wolfe, T.G.; Von Herrath, M.; Angulo, A.; Ghazal, P.; et al. Impaired immunoproteasome assembly and immune responses in PA28-/- mice. Science 1999, 286, 2162–2165. [Google Scholar] [CrossRef] [PubMed]
- Khor, B.; Bredemeyer, A.L.; Huang, C.-Y.; Turnbull, I.R.; Evans, R.; Maggi, L.B., Jr.; White, J.M.; Walker, L.M.; Carnes, K.; Hess, R.A.; et al. Proteasome activator PA200 is required for normal spermatogenesis. Mol. Cell. Biol. 2006, 26, 2999–3007. [Google Scholar] [CrossRef] [PubMed]
- Sutovsky, P. Ubiquitin-dependent proteolysis in mammalian spermatogenesis, fertilization, and sperm quality control: Killing three birds with one stone. Microsc. Res. Tech. 2003, 61, 88–102. [Google Scholar] [CrossRef] [PubMed]
- Robzyk, K.; Recht, L.; Osley, M.A. Rad6-dependent ubiquitination of histone H2B in yeast. Science 2000, 287, 501–504. [Google Scholar] [CrossRef]
- Lewis, J.D.; Abbott, D.W.; Ausió, J. A haploid affair: Core histone transitions during spermatogenesis. Biochem. Cell Biol. 2003, 81, 131–140. [Google Scholar] [CrossRef]
- Hecht, N.B. The making of a spermatozoon: A molecular perspective. Dev. Genet. 1995, 16, 95–103. [Google Scholar] [CrossRef]
- Karbowski, M.; Youle, R.J. Regulating mitochondrial outer membrane proteins by ubiquitination and proteasomal degradation. Curr. Opin. Cell Biol. 2011, 23, 476–482. [Google Scholar] [CrossRef]
- Ziviani, E.; Tao, R.N.; Whitworth, A.J. Drosophila parkin requires PINK1 for mitochondrial translocation and ubiquitinates mitofusin. Proc. Natl. Acad. Sci. USA 2010, 107, 5018–5023. [Google Scholar] [CrossRef]
- Ma, J.; Katz, E.; Belote, J.M. Expression of proteasome subunit isoforms during spermatogenesis in Drosophila melanogaster. Insect Mol. Biol. 2002, 11, 627–639. [Google Scholar] [CrossRef]
- Zhong, L.; Belote, J.M. The testis-specific proteasome subunit Prosalpha6T of D. melanogaster is required for individualization and nuclear maturation during spermatogenesis. Development 2007, 134, 3517–3525. [Google Scholar] [CrossRef]
- Yuan, X.; Miller, M.; Belote, J.M. Duplicated proteasome subunit genes in Drosophila melanogaster encoding testes-specific isoforms. Genetics 1996, 144, 147–157. [Google Scholar] [CrossRef]
- Zivkovic, D.; Dafun, A.S.; Menneteau, T.; Schahl, A.; Lise, S.; Kervarrec, C.; Rêgo, A.T.; da Fonseca, P.C.A.; Chavent, M.; Pineau, C.; et al. Proteasome complexes experience profound structural and functional rearrangements throughout mammalian spermatogenesis. Biorxiv Prepr. Serv. Biol. 2021. preprint. [Google Scholar] [CrossRef]
- Rao, H.B.D.P.; Qiao, H.; Bhatt, S.K.; Bailey, L.R.; Tran, H.D.; Bourne, S.L.; Qiu, W.; Deshpande, A.; Sharma, A.N.; Beebout, C.J.; et al. A sumo-ubiquitin relay recruits proteasomes to chromosome axes to regulate meiotic recombination. Science 2017, 355, 403–407. [Google Scholar] [CrossRef]
- Ustrell, V.; Hoffman, L.; Pratt, G.; Rechsteiner, M. PA200, a nuclear proteasome activator involved in DNA repair. EMBO J. 2002, 21, 3516–3525. [Google Scholar] [CrossRef]
- Blickwedehl, J.; McEvoy, S.; Wong, I.; Kousis, P.; Clements, J.; Elliott, R.; Cresswell, P.; Liang, P.; Bangia, N. Proteasomes and proteasome activator 200 kda (PA200) accumulate on chromatin in response to ionizing radiation. Radiat. Res. 2007, 167, 663–674. [Google Scholar] [CrossRef]
- Qian, M.-X.; Pang, Y.; Liu, C.H.; Haratake, K.; Du, B.-Y.; Ji, D.-Y.; Wang, G.-F.; Zhu, Q.-Q.; Song, W.; Yu, Y.; et al. Acetylation-mediated proteasomal degradation of core histones during DNA repair and spermatogenesis. Cell 2013, 153, 1012–1024. [Google Scholar] [CrossRef]
- Braun, R.E. Packaging paternal chromosomes with protamine. Nat. Genet. 2001, 28, 10–12. [Google Scholar] [CrossRef]
- Savulescu, A.F.; Glickman, M.H. Proteasome activator 200: The heat is on. Mol. Cell Proteom. 2011, 10, R110.006890. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Wang, Y.; Yu, T.; Huang, Y.; Li, M.; Saeed, A.F.U.H.; Perčulija, V.; Li, D.; Xiao, J.; Wang, D.; et al. Cryo-EM structures of the human PA200 and PA200-20S complex reveal regulation of proteasome gate opening and two PA200 apertures. PLoS Biol. 2020, 18, e3000654. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.-C.; Yang, W.-X. New insights to the ubiquitin-proteasome pathway (UPP) mechanism during spermatogenesis. Mol. Biol. Rep. 2013, 40, 3213–3230. [Google Scholar] [CrossRef]
- Richburg, J.H.; Myers, J.L.; Bratton, S.B. The role of E3 ligases in the ubiquitin-dependent regulation of spermatogenesis. Semin. Cell Dev. Biol. 2014, 30, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Lin, C.; Kim, S.T.; Roig, I.; Chen, H.; Liu, L.; Veith, G.M.; Jin, R.U.; Keeney, S.; Jasin, M.; et al. The E3 ubiquitin ligase cullin 4A regulates meiotic progression in mouse spermatogenesis. Dev. Biol. 2011, 356, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Kopanja, D.; Roy, N.; Stoyanova, T.; Hess, R.A.; Bagchi, S.; Raychaudhuri, P. Cul4A is essential for spermatogenesis and male fertility. Dev. Biol. 2011, 352, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Lidak, T.; Baloghova, N.; Korinek, V.; Sedlacek, R.; Balounova, J.; Kasparek, P.; Cermak, L. CRL4-DCAF12 ubiquitin ligase controls MOV10 RNA helicase during spermatogenesis and T cell activation. Int. J. Mol. Sci. 2021, 22, 5394. [Google Scholar] [CrossRef]
- Yin, Y.; Liu, L.; Yang, C.; Lin, C.; Veith, G.M.; Wang, C.; Sutovsky, P.; Zhou, P.; Ma, L. Cell autonomous and nonautonomous function of CUL4B in mouse spermatogenesis. J. Biol. Chem. 2016, 291, 6923–6935. [Google Scholar] [CrossRef]
- Arama, E.; Bader, M.; Rieckhof, G.E.; Steller, H. A ubiquitin ligase complex regulates caspase activation during sperm differentiation in Drosophila. PLoS Biol. 2007, 5, e251. [Google Scholar] [CrossRef]
- Wang, S.H.; Zheng, H.L.; Esaki, Y.; Kelly, F.; Yan, W. Cullin3 is a KLHL10-interacting protein preferentially expressed during late spermiogenesis. Biol. Reprod. 2006, 74, 102–108. [Google Scholar] [CrossRef]
- Singer, J.D.; Gurian-West, M.; Clurman, B.; Roberts, J.M. Cullin-3 targets cyclin E for ubiquitination and controls S phase in mammalian cells. Genes Dev. 1999, 13, 2375–2387. [Google Scholar] [CrossRef] [PubMed]
- Burger, J.; Merlet, J.; Tavernier, N.; Richaudeau, B.; Arnold, A.; Ciosk, R.; Bowerman, B.; Pintard, L. CRL2(LRR-1) E3-ligase regulates proliferation and progression through meiosis in the Caenorhabditis elegans germline. PLoS Genet. 2013, 9, e1003375. [Google Scholar] [CrossRef]
- Merlet, J.; Burger, J.; Tavernier, N.; Richaudeau, B.; Gomes, J.-E.; Pintard, L. The CRL2LRR-1ubiquitin ligase regulates cell cycle progression during C. elegans development. Development 2010, 137, 3857–3866. [Google Scholar] [CrossRef]
- Huen, M.S.Y.; Grant, R.; Manke, I.; Minn, K.; Yu, X.; Yaffe, M.B.; Chen, J. RNF8 transduces the DNA-damage signal via histone ubiquitylation and checkpoint protein assembly. Cell 2007, 131, 901–914. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.-Y.; Wu, J.; Ye, L.; Gavrilina, G.B.; Saunders, T.L.; Yu, X. RNF8-dependent histone modifications regulate nucleosome removal during spermatogenesis. Dev. Cell 2010, 18, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Halaby, M.-J.; Hakem, A.; Cardoso, R.; El Ghamrasni, S.; Harding, S.; Chan, N.; Bristow, R.; Sanchez, O.; Durocher, D.; et al. Rnf8 deficiency impairs class switch recombination, spermatogenesis, and genomic integrity and predisposes for cancer. J. Exp. Med. 2010, 207, 983–997. [Google Scholar] [CrossRef]
- Ma, T.; Keller, J.A.; Yu, X.C. RNF8-dependent histone ubiquitination during DNA damage response and spermatogenesis. Acta Bioch. Bioph. Sin. 2011, 43, 339–345. [Google Scholar] [CrossRef]
- Sin, H.S.; Barski, A.; Zhang, F.; Kartashov, A.V.; Nussenzweig, A.; Chen, J.; Andreassen, P.R.; Namekawa, S.H. RNF8 regulates active epigenetic modifications and escape gene activation from inactive sex chromosomes in post-meiotic spermatids. Genes Dev. 2012, 26, 2737–2748. [Google Scholar] [CrossRef]
- Ozata, D.M.; Gainetdinov, I.; Zoch, A.; O’Carroll, D.; Zamore, P.D. PIWI-interacting RNAs: Small RNAs with big functions. Nat. Rev. Genet. 2019, 20, 89–108. [Google Scholar] [CrossRef]
- Zhao, S.; Gou, L.-T.; Zhang, M.; Zu, L.-D.; Hua, M.-M.; Hua, Y.; Shi, H.-J.; Li, Y.; Li, J.; Li, D.; et al. piRNA-triggered MIWI ubiquitination and removal by APC/C in late spermatogenesis. Dev. Cell 2013, 24, 13–25. [Google Scholar] [CrossRef]
- Kwon, Y.T.; Xia, Z.; An, J.Y.; Tasaki, T.; Davydov, I.V.; Seo, J.W.; Sheng, J.; Xie, Y.; Varshavsky, A. Female lethality and apoptosis of spermatocytes in mice lacking the UBR2 ubiquitin ligase of the N-end rule pathway. Mol. Cell Biol. 2003, 23, 8255–8271. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Y.; Kwon, Y.T.; An, J.Y.; Eller, D.; Tsai, S.-C.; Diaz-Perez, S.; Troke, J.J.; Teitell, M.A.; Marahrens, Y. Loss of Ubr2, an E3 ubiquitin ligase, leads to chromosome fragility and impaired homologous recombinational repair. Mutat. Res. 2006, 596, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Cheng, Y.; An, J.Y.; Kwon, Y.T.; Eckardt, S.; Leu, N.A.; McLaughlin, K.J.; Wang, P.J. The ubiquitin ligase Ubr2, a recognition E3 component of the N-end rule pathway, stabilizes Tex19.1 during spermatogenesis. PLoS ONE 2010, 5, e14017. [Google Scholar] [CrossRef] [PubMed]
- An, J.Y.; Kim, E.-A.; Jiang, Y.; Zakrzewska, A.; Kim, D.E.; Lee, M.J.; Mook-Jung, I.; Zhang, Y.; Kwon, Y.T. UBR2 mediates transcriptional silencing during spermatogenesis via histone ubiquitination. Proc. Natl. Acad. Sci. USA 2010, 107, 1912–1917. [Google Scholar] [CrossRef] [PubMed]
- An, J.Y.; Kim, E.; Zakrzewska, A.; Yoo, Y.D.; Jang, J.M.; Han, D.H.; Lee, M.J.; Seo, J.W.; Lee, Y.J.; Kim, T.Y.; et al. UBR2 of the N-end rule pathway is required for chromosome stability via histone ubiquitylation in spermatocytes and somatic cells. PLoS ONE 2012, 7, e37414. [Google Scholar] [CrossRef] [PubMed]
- Dickins, R.A.; Frew, I.J.; House, C.M.; O’Bryan, M.K.; Holloway, A.J.; Haviv, I.; Traficante, N.; de Kretser, D.M.; Bowtell, D.D.L. The ubiquitin ligase component Siah1a is required for completion of meiosis I in male mice. Mol. Cell Biol. 2002, 22, 2294–2303. [Google Scholar] [CrossRef]
- Frew, I.J.; Hammond, V.E.; Dickins, R.A.; Quinn, J.M.W.; Walkley, C.R.; Sims, N.A.; Schnall, R.; Della, N.G.; Holloway, A.J.; Digby, M.R.; et al. Generation and analysis of Siah2 mutant mice. Mol. Cell Biol. 2003, 23, 9150–9161. [Google Scholar] [CrossRef]
- Ward, J.O.; Reinholdt, L.G.; Motley, W.W.; Niswander, L.M.; Deacon, D.C.; Griffin, L.B.; Langlais, K.K.; Backus, V.L.; Schimenti, K.J.; O’Brien, M.J.; et al. Mutation in mouse hei10, an E3 ubiquitin ligase, disrupts meiotic crossing over. PLoS Genet. 2007, 3, e139. [Google Scholar] [CrossRef]
- Kanarek, N.; Horwitz, E.; Mayan, I.; Leshets, M.; Cojocaru, G.; Davis, M.; Tsuberi, B.Z.; Pikarsky, E.; Pagano, M.; Ben-Neriah, Y. Spermatogenesis rescue in a mouse deficient for the ubiquitin ligase SCFβ-TrCP by single substrate depletion. Genes Dev. 2010, 24, 470–477. [Google Scholar] [CrossRef]
- Guardavaccaro, D.; Kudo, Y.; Boulaire, J.; Barchi, M.; Busino, L.; Donzelli, M.; Margottin-Goguet, F.; Jackson, P.K.; Yamasaki, L.; Pagano, M. Control of meiotic and mitotic progression by the F box protein β-Trcp1 in vivo. Dev. Cell 2003, 4, 799–812. [Google Scholar] [CrossRef]
- van der Laan, R.; Uringa, E.-J.; Wassenaar, E.; Hoogerbrugge, J.W.; Sleddens, E.; Odijk, H.; Roest, H.P.; de Boer, P.; Hoeijmakers, J.H.J.; Grootegoed, J.A.; et al. Ubiquitin ligase Rad18Sc localizes to the XY body and to other chromosomal regions that are unpaired and transcriptionally silenced during male meiotic prophase. J. Cell Sci. 2004, 117, 5023–5033. [Google Scholar] [CrossRef]
- van der Laan, R.; Roest, H.P.; Hoogerbrugge, J.W.; Smit, E.M.; Slater, R.; Baarends, W.M.; Hoeijmakers, J.H.; Grootegoed, J.A. Characterization of mRAD18Sc, a mouse homolog of the yeast postreplication repair gene RAD18. Genomics 2000, 69, 86–94. [Google Scholar] [CrossRef]
- Gillot, I.; Matthews, C.; Puel, D.; Vidal, F.; Lopez, P. Ret finger protein: An E3 ubiquitin ligase juxtaposed to the XY body in meiosis. Int. J. Cell Biol. 2009, 2009, 524858. [Google Scholar] [CrossRef]
- Tezel, G.; Nagasaka, T.; Iwahashi, N.; Asai, N.; Iwashita, T.; Sakata, K.; Takahashi, M. Different nuclear/cytoplasmic distributions of RET finger protein in different cell types. Pathol. Int. 1999, 49, 881–886. [Google Scholar] [CrossRef]
- Riparbelli, M.G.; Callaini, G. The Drosophila parkin homologue is required for normal mitochondrial dynamics during spermlogenesis. Dev. Biol. 2007, 303, 108–120. [Google Scholar] [CrossRef]
- Greene, J.C.; Whitworth, A.J.; Kuo, I.; Andrews, L.A.; Feany, M.B.; Pallanck, L.J. Mitochondrial pathology and apoptotic muscle degeneration in Drosophila parkin mutants. Proc. Natl. Acad. Sci. USA 2003, 100, 4078–4083. [Google Scholar] [CrossRef]
- Smith, C.L.; DeVera, D.G.; Lamb, D.J.; Nawaz, Z.; Jiang, Y.-H.; Beaudet, A.L.; O’Malley, B.W. Genetic ablation of the steroid receptor coactivator-ubiquitin ligase, E6-AP, results in tissue-selective steroid hormone resistance and defects in reproduction. Mol. Cell Biol. 2002, 22, 525–535. [Google Scholar] [CrossRef]
- Liu, Y.-Q.; Bai, G.; Zhang, H.; Su, D.; Tao, D.-C.; Yang, Y.; Ma, Y.-X.; Zhang, S.-Z. Human RING finger protein ZNF645 is a novel testis-specific E3 ubiquitin ligase. Asian J. Androl. 2010, 12, 658–666. [Google Scholar] [CrossRef]
- Dwyer, J.L.; Richburg, J.H. Age-dependent alterations in spermatogenesis in itchy mice. Spermatogenesis 2012, 2, 104–116. [Google Scholar] [CrossRef]
- Lui, W.Y.; Lee, W.M. cAMP perturbs inter-Sertoli tight junction permeability barrier in vitro via its effect on proteasome-sensitive ubiquitination of occludin. J. Cell Physiol. 2005, 203, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Oughtred, R.; Wing, S.S. Characterization of E3Histone, a novel testis ubiquitin protein ligase which ubiquitinates histones. Mol. Cell. Biol. 2005, 25, 2819–2831. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Miao, D.; Xia, Q.; Hermo, L.; Wing, S.S. Regulated expression of the ubiquitin protein ligase, E3(Histone)/LASU1/Mule/ARF-BP1/HUWE1, during spermatogenesis. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2007, 236, 2889–2898. [Google Scholar] [CrossRef]
- Rodriguez, C.I.; Stewart, C.L. Disruption of the ubiquitin ligase HERC4 causes defects in spermatozoon maturation and impaired fertility. Dev. Biol. 2007, 312, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Manku, G.; Wing, S.S.; Culty, M. Expression of the ubiquitin proteasome system in neonatal rat gonocytes and spermatogonia: Role in gonocyte differentiation. Biol. Reprod. 2012, 87, 44. [Google Scholar] [CrossRef]
- Oughtred, R.; Bedard, N.; Adegoke, O.A.J.; Morales, C.R.; Trasler, J.; Rajapurohitam, V.; Wing, S.S. Characterization of rat100, a 300-kilodalton ubiquitin-protein ligase induced in germ cells of the rat testis and similar to the Drosophila hyperplastic discs gene. Endocrinology 2002, 143, 3740–3747. [Google Scholar] [CrossRef]
- Mansfield, E.; Hersperger, E.; Biggs, J.; Shearn, A. Genetic and molecular analysis of hyperplastic disks, a gene whose product is required for regulation of cell-proliferation in Drosophila melanogaster imaginal disks and germ cells. Dev. Biol. 1994, 165, 507–526. [Google Scholar] [CrossRef]
- Pertceva, J.A.; Dorogova, N.V.; Bolobolova, E.U.; Nerusheva, O.O.; Fedorova, S.A.; Omelyanchuk, L.V. The role of Drosophila hyperplastic discs gene in spermatogenesis. Cell Biol. Int. 2010, 34, 991–996. [Google Scholar] [CrossRef]
- Lerer-Goldshtein, T.; Bel, S.; Shpungin, S.; Pery, E.; Motro, B.; Goldstein, R.S.; Bar-Sheshet, S.I.; Breitbart, H.; Nir, U. TMF/ARA160: A key regulator of sperm development. Dev. Biol. 2010, 348, 12–21. [Google Scholar] [CrossRef]
- Vyas, R.; Kumar, R.; Clermont, F.; Helfricht, A.; Kalev, P.; Sotiropoulou, P.; Hendriks, I.A.; Radaelli, E.; Hochepied, T.; Blanpain, C.; et al. RNF4 is required for DNA double-strand break repair in vivo. Cell Death Differ. 2013, 20, 490–502. [Google Scholar] [CrossRef]
- Rivkin, E.; Kierszenbaum, A.L.; Gil, M.; Tres, L.L. Rnf19a, a ubiquitin protein ligase, and Psmc3, a component of the 26S proteasome, tether to the acrosome membranes and the head-tail coupling apparatus during rat spermatid development. Dev. Dyn. 2009, 238, 1851–1861. [Google Scholar] [CrossRef]
- Nian, H.; Zhang, W.; Shi, H.X.; Zhao, Q.Z.; Xie, Q.; Liao, S.Y.; Zhang, Y.; Zhang, Z.Q.; Wang, C.; Han, C.S. Mouse RING finger protein RNF133 is a testis-specific endoplasmic reticulum-associated E3 ubiquitin ligase. Cell Res. 2008, 18, 800–802. [Google Scholar] [CrossRef]
- Nian, H.; Fan, C.; Liao, S.; Shi, Y.; Zhang, K.; Liu, Y.; Han, C. RNF151, a testis-specific RING finger protein, interacts with dysbindin. Arch. Biochem. Biophys. 2007, 465, 157–163. [Google Scholar] [CrossRef]
- Bohgaki, T.; Bohgaki, M.; Cardoso, R.; Panier, S.; Zeegers, D.; Li, L.; Stewart, G.S.; Sanchez, O.; Hande, M.P.; Durocher, D.; et al. Genomic instability, defective spermatogenesis, immunodeficiency, and cancer in a mouse model of the RIDDLE syndrome. PLoS Genet. 2011, 7, e1001381. [Google Scholar] [CrossRef]
- Zhao, B.; Ito, K.; Iyengar, P.V.; Hirose, S.; Nakamura, N. MARCH7 E3 ubiquitin ligase is highly expressed in developing spermatids of rats and its possible involvement in head and tail formation. Histochem. Cell Biol. 2013, 139, 447–460. [Google Scholar] [CrossRef]
- Iyengar, P.V.; Hirota, T.; Hirose, S.; Nakamura, N. Membrane-associated RING-CH 10 (MARCH10 protein) is a microtubule-associated E3 ubiquitin ligase of the spermatid flagella. J. Biol. Chem. 2011, 286, 39082–39090. [Google Scholar] [CrossRef]
- Morokuma, Y.; Nakamura, N.; Kato, A.; Notoya, M.; Yamamoto, Y.; Sakai, Y.; Fukuda, H.; Yamashina, S.; Hirata, Y.; Hirose, S. MARCH-XI, a novel transmembrane ubiquitin ligase implicated in ubiquitin-dependent protein sorting in developing spermatids. J. Biol. Chem. 2007, 282, 24806–24815. [Google Scholar] [CrossRef]
- Zheng, N.; Shabek, N. Ubiquitin ligases: Structure, function, and regulation. Annu. Rev. Biochem. 2017, 86, 129–157. [Google Scholar] [CrossRef]
- Sarikas, A.; Hartmann, T.; Pan, Z.-Q. The cullin protein family. Genome Biol. 2011, 12, 220. [Google Scholar] [CrossRef]
- Hsu, J.Y.; Reimann, J.D.R.; Sørensen, C.S.; Lukas, J.; Jackson, P.K. E2F-dependent accumulation of hEmi1 regulates S phase entry by inhibiting APC(Cdh1). Nat. Cell Biol. 2002, 4, 358–366. [Google Scholar] [CrossRef]
- Zhou, B.P.; Deng, J.; Xia, W.; Xu, J.; Li, Y.M.; Gunduz, M.; Hung, M.-C. Dual regulation of Snail by GSK-3β-mediated phosphorylation in control of epithelial-mesenchymal transition. Nat. Cell Biol. 2004, 6, 931–940. [Google Scholar] [CrossRef]
- Yu, C.; Zhang, Y.-L.; Pan, W.-W.; Li, X.-M.; Wang, Z.-W.; Ge, Z.-J.; Zhou, J.-J.; Cang, Y.; Tong, C.; Sun, Q.-Y.; et al. CRL4 complex regulates mammalian oocyte survival and reprogramming by activation of TET proteins. Science 2013, 342, 1518–1521. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Ji, S.-Y.; Sha, Q.-Q.; Sun, Q.-Y.; Fan, H.-Y. CRL4-DCAF1 ubiquitin E3 ligase directs protein phosphatase 2A degradation to control oocyte meiotic maturation. Nat. Commun. 2015, 6, 8017. [Google Scholar] [CrossRef]
- Yu, J.; Lan, X.; Chen, X.; Yu, C.; Xu, Y.; Liu, Y.; Xu, L.; Fan, H.-Y.; Tong, C. Protein synthesis and degradation are essential to regulate germline stem cell homeostasis in Drosophila testes. Development 2016, 143, 2930–2945. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.E.J.; Ruvkun, G. Caenorhabditis elegans ADAR editing and the ERI-6/7/MOV10 RNAi pathway silence endogenous viral elements and LTR retrotransposons. Proc. Natl. Acad. Sci. USA 2020, 117, 5987–5996. [Google Scholar] [CrossRef]
- Goodyer, W.; Kaitna, S.; Couteau, F.; Ward, J.D.; Boulton, S.J.; Zetka, M. HTP-3 links DSB formation with homolog pairing and crossing over during C. elegans meiosis. Dev. Cell 2008, 14, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.-M. The anaphase promoting complex/cyclosome: A machine designed to destroy. Nat. Rev. Mol. Cell Biol. 2006, 7, 644–656. [Google Scholar] [CrossRef]
- Lara-Gonzalez, P.; Moyle, M.W.; Budrewicz, J.; Mendoza-Lopez, J.; Oegema, K.; Desai, A. The G2-to-M transition is ensured by a dual mechanism that protects cyclin B from degradation by Cdc20-activated APC/C. Dev. Cell 2019, 51, 313–325.e10. [Google Scholar] [CrossRef]
- Cappell, S.D.; Chung, M.; Jaimovich, A.; Spencer, S.L.; Meyer, T. Irreversible APCCdh1 inactivation underlies the point of no return for cell cycle entry. Cell 2016, 166, 167–180. [Google Scholar] [CrossRef]
- Siomi, M.C.; Sato, K.; Pezic, D.; Aravin, A.A. PIWI-interacting small RNAs: The vanguard of genome defence. Nat. Rev. Mol. Cell Biol. 2011, 12, 246–258. [Google Scholar] [CrossRef]
- Liu, J.; Stevens, J.; Rote, C.A.; Yost, H.J.; Hu, Y.X.; Neufeld, K.L.; White, R.L.; Matsunami, N. Siah-1 mediates a novel beta-catenin degradation pathway linking p53 to the adenomatous polyposis coli protein. Mol. Cell 2001, 7, 927–936. [Google Scholar] [CrossRef]
- Singleton, M.R.; Uhlmann, F. Separase-securin complex: A cunning way to control chromosome segregation. Nat. Struct. Mol. Biol. 2017, 24, 337–339. [Google Scholar] [CrossRef]
- Qiao, H.; Prasada Rao, H.B.D.; Yang, Y.; Fong, J.H.; Cloutier, J.M.; Deacon, D.C.; Nagel, K.E.; Swartz, R.K.; Strong, E.; Holloway, J.K.; et al. Antagonistic roles of ubiquitin ligase HEI10 and SUMO ligase RNF212 regulate meiotic recombination. Nat. Genet. 2014, 46, 194–199. [Google Scholar] [CrossRef]
- Turner, J.M.A. Meiotic sex chromosome inactivation. Development 2007, 134, 1823–1831. [Google Scholar] [CrossRef]
- Sonnack, V.; Failing, K.; Bergmann, M.; Steger, K. Expression of hyperacetylated histone H4 during normal and impaired human spermatogenesis. Andrologia 2002, 34, 384–390. [Google Scholar] [CrossRef]
- Suresh, B.; Lee, J.; Hong, S.-H.; Kim, K.-S.; Ramakrishna, S. The role of deubiquitinating enzymes in spermatogenesis. Cell Mol. Life Sci. 2015, 72, 4711–4720. [Google Scholar] [CrossRef]
- Mevissen, T.E.T.; Komander, D. Mechanisms of deubiquitinase specificity and regulation. Annu. Rev. Biochem. 2017, 86, 159–192. [Google Scholar] [CrossRef]
- Lin, H.; Keriel, A.; Morales, C.R.; Bedard, N.; Zhao, Q.; Hingamp, P.; Lefrançois, S.; Combaret, L.; Wing, S.S. Divergent N-terminal sequences target an inducible testis deubiquitinating enzyme to distinct subcellular structures. Mol. Cell Biol. 2000, 20, 6568–6578. [Google Scholar] [CrossRef]
- Bedard, N.; Yang, Y.; Gregory, M.; Cyr, D.G.; Suzuki, J.; Yu, X.; Chian, R.-C.; Hermo, L.; O’Flaherty, C.; Smith, C.E.; et al. Mice lacking the USP2 deubiquitinating enzyme have severe male subfertility associated with defects in fertilization and sperm motility. Biol. Reprod. 2011, 85, 594–604. [Google Scholar] [CrossRef]
- Luo, M.; Zhou, J.; Leu, N.A.; Abreu, C.M.; Wang, J.; Anguera, M.C.; de Rooij, D.G.; Jasin, M.; Wang, P.J. Polycomb protein SCML2 associates with USP7 and counteracts Histone H2A ubiquitination in the XY chromatin during male meiosis. PLoS Genet. 2015, 11, e1004954. [Google Scholar] [CrossRef]
- Berruti, G.; Martegani, E. mUBPy and MSJ-1, a deubiquitinating enzyme and a molecular chaperone specifically expressed in testis, associate with the acrosome and centrosome in mouse germ cells. Ann. N. Y. Acad. Sci. 2002, 973, 5–7. [Google Scholar] [CrossRef]
- Berruti, G.; Martegani, E. The deubiquitinating enzyme mUBPy interacts with the sperm-specific molecular chaperone MSJ-1: The relation with the proteasome, acrosome, and centrosome in mouse male germ cells. Biol. Reprod. 2005, 72, 14–21. [Google Scholar] [CrossRef]
- Lee, K.H.; Song, G.J.; Kang, I.S.; Kim, S.W.; Paick, J.-S.; Chung, C.H.; Rhee, K. Ubiquitin-specific protease activity of USP9Y, a male infertility gene on the Y chromosome. Reprod. Fertil. Dev. 2003, 15, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Skaletsky, H.; Birren, B.; Devon, K.; Tang, Z.; Silber, S.; Oates, R.; Page, D.C. An azoospermic man with a de novo point mutation in the Y-chromosomal gene USP9Y. Nat. Genet. 1999, 23, 429–432. [Google Scholar] [CrossRef] [PubMed]
- Luddi, A.; Margollicci, M.; Gambera, L.; Serafini, F.; Cioni, M.; De Leo, V.; Balestri, P.; Piomboni, P. Spermatogenesis in a man with complete deletion of USP9Y. N. Engl. J. Med. 2009, 360, 881–885. [Google Scholar] [CrossRef] [PubMed]
- Krausz, C.; Degl’Innocenti, S.; Nuti, F.; Morelli, A.; Felici, F.; Sansone, M.; Varriale, G.; Forti, G. Natural transmission of USP9Y gene mutations: A new perspective on the role of AZFa genes in male fertility. Hum. Mol. Genet. 2006, 15, 2673–2681. [Google Scholar] [CrossRef]
- Kishi, K.; Uchida, A.; Takase, H.M.; Suzuki, H.; Kurohmaru, M.; Tsunekawa, N.; Kanai-Azuma, M.; Wood, S.A.; Kanai, Y. Spermatogonial deubiquitinase USP9X is essential for proper spermatogenesis in mice. Reproduction 2017, 154, 135–143. [Google Scholar] [CrossRef]
- Crimmins, S.; Sutovsky, M.; Chen, P.-C.; Huffman, A.; Wheeler, C.; Swing, D.A.; Roth, K.; Wilson, J.; Sutovsky, P.; Wilson, S. Transgenic rescue of ataxia mice reveals a male-specific sterility defect. Dev. Biol. 2009, 325, 33–42. [Google Scholar] [CrossRef]
- Kovács, L.; Nagy, A.; Pál, M.; Deák, P. Usp14 is required for spermatogenesis and ubiquitin stress responses in Drosophila melanogaster. J. Cell Sci. 2020, 133, jcs237511. [Google Scholar] [CrossRef]
- Shi, Y.-C.; Wei, L.; Cui, Y.-X.; Shang, X.-J.; Wang, H.-Y.; Xia, X.-Y.; Zhou, Y.-C.; Li, H.; Jiang, H.-T.; Zhu, W.-M.; et al. Association between ubiquitin-specific protease USP26 polymorphism and male infertility in Chinese men. Clin. Chim. Acta 2011, 412, 545–549. [Google Scholar] [CrossRef]
- Luddi, A.; Crifasi, L.; Quagliarello, A.; Governini, L.; De Leo, V.; Piomboni, P. Single nucleotide polymorphisms of USP26 in azoospermic men. Syst. Biol. Reprod. Med. 2016, 62, 372–378. [Google Scholar] [CrossRef][Green Version]
- Zhang, J.; Qiu, S.-D.; Li, S.-B.; Zhou, D.-X.; Tian, H.; Huo, Y.-W.; Ge, L.; Zhang, Q.-Y. Novel mutations in ubiquitin-specific protease 26 gene might cause spermatogenesis impairment and male infertility. Asian J. Androl. 2007, 9, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Stouffs, K.; Lissens, W.; Tournaye, H.; Van Steirteghem, A.; Liebaers, I. Possible role of USP26 in patients with severely impaired spermatogenesis. Eur. J. Hum. Genet. 2005, 13, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shao, X.-G.; Shi, Y.-B.; Yan, L.; Wang, L.; Tian, H.; Qiu, S.-D. Polymorphism of Usp26 correlates with idiopathic male infertility. Zhonghua Nan Ke Xue 2012, 18, 105–108. [Google Scholar]
- Stouffs, K.; Lissens, W.; Tournaye, H.; Van Steirteghem, A.; Liebaers, I. Alterations of the USP26 gene in Caucasian men. Int. J. Androl. 2006, 29, 614–617. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-K.; Kim, Y.-S.; Yoo, K.-J.; Lee, H.-J.; Lee, D.-R.; Yeo, C.Y.; Baek, K.-H. The expression of Usp42 during embryogenesis and spermatogenesis in mouse. Gene Expr. Patterns 2007, 7, 143–148. [Google Scholar] [CrossRef]
- Devi, L.; Pawar, R.M.; Makala, H.; Goel, S. Conserved expression of ubiquitin carboxyl-terminal esterase L1 (UCHL1) in mammalian testes. Indian. J. Exp. Biol. 2015, 53, 305–312. [Google Scholar] [PubMed]
- Toliczenko-Bernatowicz, D.; Matuszczak, E.; Tylicka, M.; Szymańska, B.; Komarowska, M.; Gorodkiewicz, E.; Debek, W.; Hermanowicz, A. Overexpression of ubiquitin carboxyl-terminal hydrolase 1 (UCHL1) in boys with cryptorchidism. PLoS ONE 2018, 13, e0191806. [Google Scholar] [CrossRef]
- Wang, Y.-L.; Liu, W.Z.; Sun, Y.-J.; Kwon, J.; Setsuie, R.; Osaka, H.; Noda, M.; Aoki, S.; Yoshikawa, Y.; Wada, K. Overexpression of ubiquitin carboxyl-terminal hydrolase L1 arrests spermatogenesis in transgenic mice. Mol. Reprod. Dev. 2006, 73, 40–49. [Google Scholar] [CrossRef]
- Kwon, J.; Mochida, K.; Wang, Y.-L.; Sekiguchi, S.; Sankai, T.; Aoki, S.; Ogura, A.; Yoshikawa, Y.; Wada, K. Ubiquitin C-terminal hydrolase L-1 is essential for the early apoptotic wave of germinal cells and for sperm quality control during spermatogenesis. Biol. Reprod. 2005, 73, 29–35. [Google Scholar] [CrossRef]
- Kwon, J. The new function of two ubiquitin C-terminal hydrolase isozymes as reciprocal modulators of germ cell apoptosis. Exp. Anim. 2007, 56, 71–77. [Google Scholar] [CrossRef]
- Kwon, J.; Wang, Y.-L.; Setsuie, R.; Sekiguchi, S.; Sakurai, M.; Sato, Y.; Lee, W.-W.; Ishii, Y.; Kyuwa, S.; Noda, M.; et al. Developmental regulation of ubiquitin C-terminal hydrolase isozyme expression during spermatogenesis in mice. Biol. Reprod. 2004, 71, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, L.J.; Semenova, E.; Levorse, J.M.; Tilghman, S.M. Expression and functional analysis of Uch-L3 during mouse development. Mol. Cell Biol. 2000, 20, 2498–2504. [Google Scholar] [CrossRef]
- Osawa, Y.; Wang, Y.L.; Osaka, H.; Aoki, S.; Wada, K. Cloning, expression, and mapping of a mouse gene, Uchl4, highly homologous to human and mouse Uchl3. Biochem. Biophys. Res. Commun. 2001, 283, 627–633. [Google Scholar] [CrossRef]
- Wright, A.; Reiley, W.W.; Chang, M.; Jin, W.; Lee, A.J.; Zhang, M.; Sun, S.-C. Regulation of early wave of germ cell apoptosis and spermatogenesis by deubiquitinating enzyme CYLD. Dev. Cell. 2007, 13, 705–716. [Google Scholar] [CrossRef]
- Berruti, G.; Paiardi, C. USP8/UBPy-regulated sorting and the development of sperm acrosome: The recruitment of MET. Reproduction 2015, 149, 633–644. [Google Scholar] [CrossRef][Green Version]
- Berruti, G.; Ripolone, M.; Ceriani, M. USP8, a regulator of endosomal sorting, is involved in mouse acrosome biogenesis through interaction with the spermatid ESCRT-0 complex and microtubules. Biol. Reprod. 2010, 82, 930–939. [Google Scholar] [CrossRef]
- Quenneville, N.R.; Chao, T.-Y.; McCaffery, J.M.; Conibear, E. Domains within the GARP subunit Vps54 confer separate functions in complex assembly and early endosome recognition. Mol. Biol. Cell 2006, 17, 1859–1870. [Google Scholar] [CrossRef]
- Lasko, P. The DEAD-box helicase Vasa: Evidence for a multiplicity of functions in RNA processes and developmental biology. Biochim. Biophys. Acta. 2013, 1829, 810–816. [Google Scholar] [CrossRef]
- Wosnitzer, M.S.; Mielnik, A.; Dabaja, A.; Robinson, B.; Schlegel, P.N.; Paduch, D.A. Ubiquitin specific protease 26 (USP26) expression analysis in human testicular and extragonadal tissues indicates diverse action of USP26 in cell differentiation and tumorigenesis. PLoS ONE 2014, 9, e98638. [Google Scholar] [CrossRef]
- Wang, P.J.; Page, D.C.; McCarrey, J.R. Differential expression of sex-linked and autosomal germ-cell-specific genes during spermatogenesis in the mouse. Hum. Mol. Genet. 2005, 14, 2911–2918. [Google Scholar] [CrossRef]
- Nishimune, Y.; Tanaka, H. Infertility caused by polymorphisms or mutations in spermatogenesis-specific genes. J. Androl. 2006, 27, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-L.; Zheng, J.; Mi, Y.-J.; Zhao, J.; Tian, Q.-B. The impacts of nineteen mutations on the enzymatic activity of USP26. Gene 2018, 641, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Felipe-Medina, N.; Gómez-H, L.; Condezo, Y.B.; Sanchez-Martín, M.; Barbero, J.L.; Ramos, I.; Llano, E.; Pendás, A.M. Ubiquitin-specific protease 26 (USP26) is not essential for mouse gametogenesis and fertility. Chromosoma 2019, 128, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Ito, C.; Wakabayashi, M.; Kanzaki, S.; Ito, T.; Takada, S.; Toshimori, K.; Sekita, Y.; Kimura, T. Usp26 mutation in mice leads to defective spermatogenesis depending on genetic background. Sci. Rep. 2019, 9, 13757. [Google Scholar] [CrossRef]
- Wang, M.; Yu, T.; Hu, L.; Cheng, Z.; Li, M. Ubiquitin carboxy-terminal hydrolaseL3 correlates with human sperm count, motility and fertilization. PLoS ONE 2016, 11, e0165198. [Google Scholar] [CrossRef]
- Luo, J.; Megee, S.; Dobrinski, I. Asymmetric distribution of UCH-L1 in spermatogonia is associated with maintenance and differentiation of spermatogonial stem cells. J. Cell Physiol. 2009, 220, 460–468. [Google Scholar] [CrossRef]
- Rodriguez, I.; Ody, C.; Araki, K.; Garcia, I.; Vassalli, P. An early and massive wave of germinal cell apoptosis is required for the development of functional spermatogenesis. EMBO J. 1997, 16, 2262–2270. [Google Scholar] [CrossRef]
- Hogarth, C.A.; Evanoff, R.; Mitchell, D.; Kent, T.; Small, C.; Amory, J.K.; Griswold, M.D. Turning a spermatogenic wave into a tsunami: Synchronizing murine spermatogenesis using WIN 18,446. Biol. Reprod. 2013, 88, 1–9. [Google Scholar] [CrossRef]
- Krausz, C.; Riera-Escamilla, A. Genetics of male infertility. Nat. Rev. Urol. 2018, 15, 369–384. [Google Scholar] [CrossRef]
- Toste Rêgo, A.; da Fonseca, P.C.A. Characterization of fully recombinant human 20S and 20S-PA200 proteasome complexes. Mol. Cell 2019, 76, 138–147.e135. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Names | Expression Pattern (In Testis) | Substrates | Mutant Phenotypes | Species | References |
|---|---|---|---|---|---|
| CUL4A | Sg, Sc, St (low) | Histone H3, H4; MOV10 | Infertility; apoptosis of Sc; decreased St and Sz; abnormal Sz with low motility and defective acrosome formation | M. musculus | [45,46,47] |
| CUL4B | Sg, St | Unknown | Sz with low or no motility | M. musculus | [48] |
| CUL3 | St | dBruce | Reduced or eliminated effector caspase activation in St | D. melanogaster | [49] |
| CUL3 | St | Unknown | Early embryonic death | M. musculus | [50,51] |
| CUL2 | Male germ cells (high in Sc) | HTP-3 | Infertility; cell cycle arrest; premature meiotic entry | C. elegans | [52,53] |
| RNF8 | Undefined | Histones H2A, H2AX, H2B | Infertility; defective nucleosome removal in St | M. musculus | [54,55,56,57,58] |
| APC/C | Sc, St | PIWI/MIWI | Defective maturation of St | M. musculus | [59,60] |
| UBR2 | Testis | Histones H2A, H2B | Infertility; meiotic arrest; defective homologous recombinational repair, MSUC, and synapsis | M. musculus | [61,62,63,64,65] |
| SIAH1A | Testis | Unknown | Infertility; metaphase to telophase arrest during meiosis I; bi- or multi-nucleated anaphase cells | M. musculus | [66,67] |
| Mei4/HEI10 | Undefined | CCNB3 (?) | Infertility; failure of crossover formation; meiotic arrest; apoptosis of Sc | M. musculus | [68] |
| SCFβ-TrCP | Sg, Sc | Snail1, Emi1 (?) | β-TrCP1 deficiency: accumulation of metaphase I Sc and multinucleated St; β-TrCP1/2 double deficiency: absence of Sc, St and Sz | M. musculus | [69,70] |
| RAD18/18sc | Testis (high in Sc) | Unknown | Subfertility; defective meiotic DSB repair and MSCI | M. musculus | [71,72] |
| RFP | Male germ cells | Unknown | Undefined | M. musculus | [73,74] |
| Parkin | Undefined | Unknown | Infertility; defective mitochondrial organization and St individualization | D. melanogaster | [75,76] |
| E6-AP | Undefined | Unknown | Subfertility; reduced Sz | M. musculus | [77] |
| ZNF645 | Sc, St, Leydig cells | Unknown | Undefined | H. sapiens | [78] |
| ITCH | Testis | Occludin | Subfertility; age-dependent impairment; delayed St development and organization | M. musculus | [79,80] |
| HUWE1/LASU1 | Sg, Sc | Histones (?) | Undefined | R. norvegicus | [81,82] |
| HERC4 | Sg, Sc, St | Unknown | Subfertility; reduced and abnormal Sz | M. musculus | [83] |
| UBR5/HYD/EDD | Sc, St (low) | Unknown | Rat: Undefined Drosophila: Infertility; defects in St elongation | R. norvegicus D. melanogaster | [84,85,86,87] |
| TMF/ARA160 | Sc, St | Unknown | Infertility; reduced motility and malformation of Sz | M. musculus | [88] |
| RNF4 | Undefined | MDC1, BRCA1 | Infertility; age dependent testicular atrophy; germ cell depletion | M. musculus | [89] |
| RNF19A | Pachytene Sc, St, Sz | Unknown | Undefined | R. norvegicus | [90] |
| RNF133 | St | Unknown | Undefined | M. musculus | [91] |
| RNF149 | Male germ cells | Unknown | Undefined | R. norvegicus | [84] |
| RNF151 | St | Dysbindin (?) | Undefined | M. musculus | [92] |
| RNF168 | Undefined | Unknown | Subfertility or infertility; age-dependent impairment; testis atrophy | M. musculus | [93] |
| MARCH7 | Testis (high in St) | Unknown | Undefined | R. norvegicus | [94] |
| MARCH10 | St | Unknown | Undefined | R. norvegicus | [95] |
| MARCH11 | St | Unknown | Undefined | R. norvegicus | [96] |
| Names | Expression Pattern (In Testis) | Substrates | Mutant Phenotypes | Species | References |
|---|---|---|---|---|---|
| USP2 | Ubiquitous (high in elongating St) | Unknown | Subfertility; abnormal aggregation of St | M. musculus | [117,118] |
| USP7 | Sc (Early-pachytene) | RNF2 | Undefined | M. musculus | [119] |
| USP8 | Testis (high in St, Sz) | MSJ1 (?) | Undefined | M. musculus | [120,121] |
| USP9Y | Undefined | Unknown | SNP associated with azoospermia/oligospermia | H. sapiens | [122,123,124,125] |
| USP9X | Sg (high), Sc (low) | Unknown | Infertility; reduced reduced Sc; malformation of St and Sz | M. musculus | [126] |
| USP14 | St and Sz | Unknown | Defective St individualization; reduced and abnormal Sz | D. melanogaster, M. musculus | [127,128] |
| USP26 | Testis | Unknown | Polymorphisms associated with azoospermia/oligozoospermia/asthenozoospermia | H. sapiens | [129,130,131,132,133,134] |
| UPS42 | St | Unknown | Undefined | M. musculus | [135] |
| UCHL1 | Sg, Sertoli cells | Unknown | Increased Sg; reduced motility and malformation of Sz; resistant to early wave of germ cell apoptosis | M. musculus | [136,137,138,139] |
| UCHL3 | Pachytene Sc, St | Unknown | Increased germ cell loss after cryptorchid injury | M. musculus | [140,141,142] |
| UCHL4 | Sg | Unknown | Undefined | M. musculus | [141,143] |
| UCHL5 | Sc, St | Unknown | Undefined | M. musculus | [141] |
| CYLD | Ubiquitous | RIP1 | Infertility; deficiency and malformation of St; attenuates the early wave of germ cell apoptosis | M. musculus | [144] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, Y.; Yu, C.; Zhang, Q. Ubiquitin-Proteasome System–Regulated Protein Degradation in Spermatogenesis. Cells 2022, 11, 1058. https://doi.org/10.3390/cells11061058
Xiong Y, Yu C, Zhang Q. Ubiquitin-Proteasome System–Regulated Protein Degradation in Spermatogenesis. Cells. 2022; 11(6):1058. https://doi.org/10.3390/cells11061058
Chicago/Turabian StyleXiong, Yi, Chao Yu, and Qianting Zhang. 2022. "Ubiquitin-Proteasome System–Regulated Protein Degradation in Spermatogenesis" Cells 11, no. 6: 1058. https://doi.org/10.3390/cells11061058
APA StyleXiong, Y., Yu, C., & Zhang, Q. (2022). Ubiquitin-Proteasome System–Regulated Protein Degradation in Spermatogenesis. Cells, 11(6), 1058. https://doi.org/10.3390/cells11061058