
Review of Mechanisms and Treatment of Cancer-Induced Cardiac Cachexia

 , , , and
, , , and 
Abstract
:1. Introduction
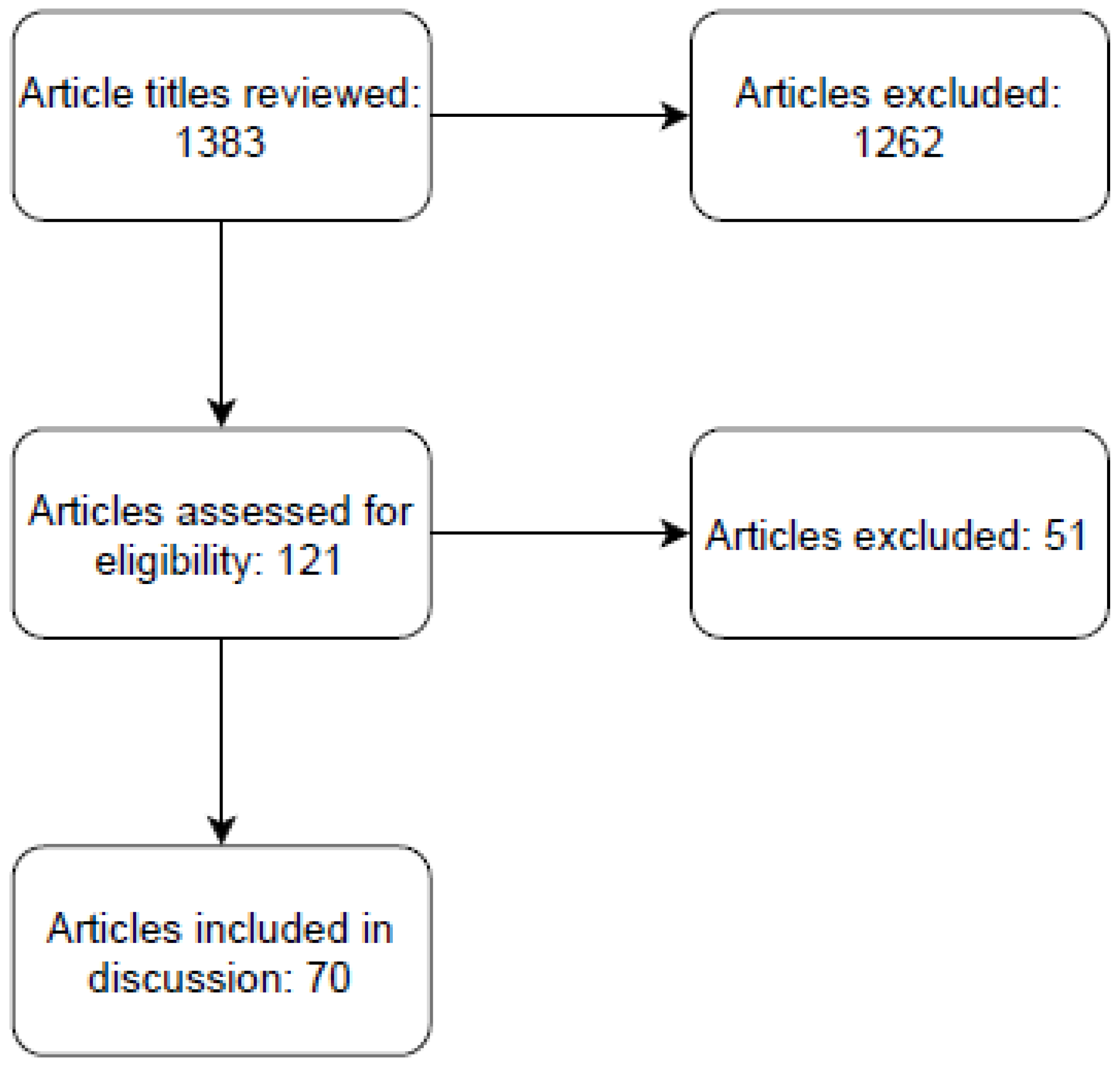
2. Materials and Methods
3. Results
4. Discussion
4.1. Mechanisms
4.1.1. TNFα
4.1.2. Autophagy
4.1.3. Reactive Oxygen Species
4.1.4. Ubiquitination: Atrogin-1/MuRF1
4.1.5. Cancer-Induced Cardiac Cachexia Pathways
4.2. Models to Study Cardiac Cachexia
4.3. Methods to Assess Cardiac Cachexia
4.4. Treatment
4.4.1. Anti-Proteolytics
4.4.2. Statins
4.4.3. STAT3 Inhibition
4.4.4. Heart Failure Medication
4.4.5. Reactive Oxygen Species
4.4.6. NF-κB
4.4.7. ActRIIB
4.4.8. Oxypurinol
4.4.9. Antidepressants
4.4.10. Weight and Muscle Gain
4.4.11. Testosterone
4.4.12. Nutrition
4.4.13. Exercise
4.4.14. Gene Therapy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blum, D.; Omlin, A.; Baracos, V.E.; Solheim, T.S.; Tan, B.H.; Stone, P.; Kaasa, S.; Fearon, K.; Strasser, F. Cancer cachexia: A systematic literature review of items and domains associated with involuntary weight loss in cancer. Crit. Rev. Oncol. Hematol. 2011, 80, 114–144. [Google Scholar] [CrossRef] [PubMed]
- Fearon, K.; Strasser, F.; Anker, S.D.; Bosaeus, I.; Bruera, E.; Fainsinger, R.L.; Jatoi, A.; Loprinzi, C.; MacDonald, N.; Mantovani, G.; et al. Definition and classification of cancer cachexia: An international consensus. Lancet Oncol. 2011, 12, 489–495. [Google Scholar] [CrossRef]
- Herremans, K.M.; Riner, A.N.; Cameron, M.E.; Trevino, J.G. The Microbiota and Cancer Cachexia. Int. J. Mol. Sci. 2019, 20, 6267. [Google Scholar] [CrossRef] [Green Version]
- Ni, J.; Zhang, L. Cancer Cachexia: Definition, Staging, and Emerging Treatments. Cancer Manag. Res. 2020, 12, 5597–5605. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, M.; Keshavarz-Fathi, M.; Baracos, V.; Arends, J.; Mahmoudi, M.; Rezaei, N. Cancer cachexia: Diagnosis, assessment, and treatment. Crit. Rev. Oncol. Hematol. 2018, 127, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, J. Lung Cancer Cachexia: Can Molecular Understanding Guide Clinical Management? Integr. Cancer Ther. 2018, 17, 1000–1008. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.R.; Yaffee, P.M.; Jamil, L.H.; Lo, S.K.; Nissen, N.; Pandol, S.J.; Tuli, R.; Hendifar, A.E. Pancreatic cancer cachexia: A review of mechanisms and therapeutics. Front. Physiol. 2014, 5, 88. [Google Scholar] [CrossRef] [Green Version]
- Peixoto da Silva, S.; Santos, J.M.O.; Costa, E.S.M.P.; Gil da Costa, R.M.; Medeiros, R. Cancer cachexia and its pathophysiology: Links with sarcopenia, anorexia and asthenia. J. Cachexia Sarcopenia Muscle 2020, 11, 619–635. [Google Scholar] [CrossRef]
- Stein, P.D.; Goldman, J.; Matta, F.; Yaekoub, A.Y. Diabetes mellitus and risk of venous thromboembolism. Am. J. Med. Sci. 2009, 337, 259–264. [Google Scholar] [CrossRef]
- Simonsen, C.; de Heer, P.; Bjerre, E.D.; Suetta, C.; Hojman, P.; Pedersen, B.K.; Svendsen, L.B.; Christensen, J.F. Sarcopenia and Postoperative Complication Risk in Gastrointestinal Surgical Oncology: A Meta-analysis. Ann. Surg. 2018, 268, 58–69. [Google Scholar] [CrossRef]
- Porporato, P.E. Understanding cachexia as a cancer metabolism syndrome. Oncogenesis 2016, 5, e200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skipworth, R.J.; Stewart, G.D.; Dejong, C.H.; Preston, T.; Fearon, K.C. Pathophysiology of cancer cachexia: Much more than host-tumour interaction? Clin. Nutr. 2007, 26, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Kazemi-Bajestani, S.M.; Becher, H.; Fassbender, K.; Chu, Q.; Baracos, V.E. Concurrent evolution of cancer cachexia and heart failure: Bilateral effects exist. J. Cachexia Sarcopenia Muscle 2014, 5, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.T. The pathogenesis and treatment of cardiac atrophy in cancer cachexia. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H466–H477. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Nishijima, Y.; Asp, M.L.; Stout, M.B.; Reiser, P.J.; Belury, M.A. Cardiac alterations in cancer-induced cachexia in mice. Int. J. Oncol. 2010, 37, 347–353. [Google Scholar] [PubMed] [Green Version]
- Muhlfeld, C.; Das, S.K.; Heinzel, F.R.; Schmidt, A.; Post, H.; Schauer, S.; Papadakis, T.; Kummer, W.; Hoefler, G. Cancer induces cardiomyocyte remodeling and hypoinnervation in the left ventricle of the mouse heart. PLoS ONE 2011, 6, e20424. [Google Scholar] [CrossRef] [PubMed]
- Schafer, M.; Oeing, C.U.; Rohm, M.; Baysal-Temel, E.; Lehmann, L.H.; Bauer, R.; Volz, H.C.; Boutros, M.; Sohn, D.; Sticht, C.; et al. Ataxin-10 is part of a cachexokine cocktail triggering cardiac metabolic dysfunction in cancer cachexia. Mol. Metab. 2016, 5, 67–78. [Google Scholar] [CrossRef]
- Costelli, P.; Carbo, N.; Tessitore, L.; Bagby, G.J.; Lopez-Soriano, F.J.; Argiles, J.M.; Baccino, F.M. Tumor necrosis factor-alpha mediates changes in tissue protein turnover in a rat cancer cachexia model. J. Clin. Investig. 1993, 92, 2783–2789. [Google Scholar] [CrossRef] [Green Version]
- Llovera, M.; Lopez-Soriano, F.J.; Argiles, J.M. Effects of tumor necrosis factor-alpha on muscle-protein turnover in female Wistar rats. J. Natl. Cancer Inst. 1993, 85, 1334–1339. [Google Scholar] [CrossRef]
- Pietzsch, S.; Ricke-Hoch, M.; Stapel, B.; Hilfiker-Kleiner, D. Modulation of cardiac AKT and STAT3 signalling in preclinical cancer models and their impact on the heart. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118519. [Google Scholar] [CrossRef]
- Miyagawa, Y.; Nukaga, S.; Mori, T.; Fujiwara-Tani, R.; Fujii, K.; Mori, S.; Goto, K.; Kishi, S.; Sasaki, T.; Nakashima, C.; et al. Evaluation of cancer-derived myocardial impairments using a mouse model. Oncotarget 2020, 11, 3712–3722. [Google Scholar] [CrossRef] [PubMed]
- Cosper, P.F.; Leinwand, L.A. Cancer causes cardiac atrophy and autophagy in a sexually dimorphic manner. Cancer Res. 2011, 71, 1710–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manne, N.D.; Lima, M.; Enos, R.T.; Wehner, P.; Carson, J.A.; Blough, E. Altered cardiac muscle mTOR regulation during the progression of cancer cachexia in the ApcMin/+ mouse. Int. J. Oncol. 2013, 42, 2134–2140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, F.H.; Marinello, P.C.; Cecchini, A.L.; Blegniski, F.P.; Guarnier, F.A.; Cecchini, R. Oxidative and proteolytic profiles of the right and left heart in a model of cancer-induced cardiac cachexia. Pathophysiology 2014, 21, 257–265. [Google Scholar] [CrossRef]
- Hinch, E.C.; Sullivan-Gunn, M.J.; Vaughan, V.C.; McGlynn, M.A.; Lewandowski, P.A. Disruption of pro-oxidant and antioxidant systems with elevated expression of the ubiquitin proteosome system in the cachectic heart muscle of nude mice. J. Cachexia Sarcopenia Muscle 2013, 4, 287–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.E.; Brown, J.L.; Rosa-Caldwell, M.E.; Perry, R.A.; Brown, L.A.; Haynie, W.S.; Washington, T.A.; Wiggs, M.P.; Rajaram, N.; Greene, N.P. Cancer-induced Cardiac Atrophy Adversely Affects Myocardial Redox State and Mitochondrial Oxidative Characteristics. JCSM Rapid Commun. 2021, 4, 3–15. [Google Scholar] [CrossRef]
- Wysong, A.; Couch, M.; Shadfar, S.; Li, L.; Rodriguez, J.E.; Asher, S.; Yin, X.; Gore, M.; Baldwin, A.; Patterson, C.; et al. NF-kappaB inhibition protects against tumor-induced cardiac atrophy in vivo. Am. J. Pathol. 2011, 178, 1059–1068. [Google Scholar] [CrossRef] [Green Version]
- Tian, M.; Asp, M.L.; Nishijima, Y.; Belury, M.A. Evidence for cardiac atrophic remodeling in cancer-induced cachexia in mice. Int. J. Oncol. 2011, 39, 1321–1326. [Google Scholar]
- Matsuyama, T.; Ishikawa, T.; Okayama, T.; Oka, K.; Adachi, S.; Mizushima, K.; Kimura, R.; Okajima, M.; Sakai, H.; Sakamoto, N.; et al. Tumor inoculation site affects the development of cancer cachexia and muscle wasting. Int. J. Cancer 2015, 137, 2558–2565. [Google Scholar] [CrossRef] [Green Version]
- Costelli, P.; De Tullio, R.; Baccino, F.M.; Melloni, E. Activation of Ca2+-dependent proteolysis in skeletal muscle and heart in cancer cachexia. Br. J. Cancer 2001, 84, 946–950. [Google Scholar] [CrossRef] [Green Version]
- Pin, F.; Minero, V.G.; Penna, F.; Muscaritoli, M.; De Tullio, R.; Baccino, F.M.; Costelli, P. Interference with Ca2+-Dependent Proteolysis Does Not Alter the Course of Muscle Wasting in Experimental Cancer Cachexia. Front. Physiol. 2017, 8, 213. [Google Scholar] [CrossRef] [PubMed]
- Huot, J.R.; Pin, F.; Narasimhan, A.; Novinger, L.J.; Keith, A.S.; Zimmers, T.A.; Willis, M.S.; Bonetto, A. ACVR2B antagonism as a countermeasure to multi-organ perturbations in metastatic colorectal cancer cachexia. J. Cachexia Sarcopenia Muscle 2020, 11, 1779–1798. [Google Scholar] [CrossRef] [PubMed]
- Samant, S.A.; Kanwal, A.; Pillai, V.B.; Bao, R.; Gupta, M.P. The histone deacetylase SIRT6 blocks myostatin expression and development of muscle atrophy. Sci. Rep. 2017, 7, 11877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devine, R.D.; Bicer, S.; Reiser, P.J.; Velten, M.; Wold, L.E. Metalloproteinase expression is altered in cardiac and skeletal muscle in cancer cachexia. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H685–H691. [Google Scholar] [CrossRef] [Green Version]
- Michaelis, K.A.; Zhu, X.; Burfeind, K.G.; Krasnow, S.M.; Levasseur, P.R.; Morgan, T.K.; Marks, D.L. Establishment and characterization of a novel murine model of pancreatic cancer cachexia. J. Cachexia Sarcopenia Muscle 2017, 8, 824–838. [Google Scholar] [CrossRef]
- Bonetto, A.; Rupert, J.E.; Barreto, R.; Zimmers, T.A. The Colon-26 Carcinoma Tumor-bearing Mouse as a Model for the Study of Cancer Cachexia. J. Vis. Exp. 2016, 117, e54893. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Tamta, A.K.; Sarikhani, M.; Desingu, P.A.; Kizkekra, S.M.; Pandit, A.S.; Kumar, S.; Khan, D.; Raghavan, S.C.; Sundaresan, N.R. Subcutaneous Ehrlich Ascites Carcinoma mice model for studying cancer-induced cardiomyopathy. Sci. Rep. 2018, 8, 5599. [Google Scholar] [CrossRef]
- Barkhudaryan, A.; Scherbakov, N.; Springer, J.; Doehner, W. Cardiac muscle wasting in individuals with cancer cachexia. ESC Heart Fail. 2017, 4, 458–467. [Google Scholar] [CrossRef]
- Xu, H.; Crawford, D.; Hutchinson, K.R.; Youtz, D.J.; Lucchesi, P.A.; Velten, M.; McCarthy, D.O.; Wold, L.E. Myocardial dysfunction in an animal model of cancer cachexia. Life Sci. 2011, 88, 406–410. [Google Scholar] [CrossRef] [Green Version]
- Cramer, L.; Hildebrandt, B.; Kung, T.; Wichmann, K.; Springer, J.; Doehner, W.; Sandek, A.; Valentova, M.; Stojakovic, T.; Scharnagl, H.; et al. Cardiovascular function and predictors of exercise capacity in patients with colorectal cancer. J. Am. Coll. Cardiol. 2014, 64, 1310–1319. [Google Scholar] [CrossRef] [Green Version]
- Florea, V.G.; Moon, J.; Pennell, D.J.; Doehner, W.; Coats, A.J.; Anker, S.D. Wasting of the left ventricle in patients with cardiac cachexia: A cardiovascular magnetic resonance study. Int. J. Cardiol. 2004, 97, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Jafri, S.H.; Previgliano, C.; Khandelwal, K.; Shi, R. Cachexia Index in Advanced Non-Small-Cell Lung Cancer Patients. Clin. Med. Insights Oncol. 2015, 9, 87–93. [Google Scholar] [CrossRef]
- Shum, A.M.Y.; Poljak, A.; Bentley, N.L.; Turner, N.; Tan, T.C.; Polly, P. Proteomic profiling of skeletal and cardiac muscle in cancer cachexia: Alterations in sarcomeric and mitochondrial protein expression. Oncotarget 2018, 9, 22001–22022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Der-Torossian, H.; Gourin, C.G.; Couch, M.E. Translational implications of novel findings in cancer cachexia: The use of metabolomics and the potential of cardiac malfunction. Curr. Opin. Supportive Palliat. Care 2012, 6, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Barker, T.; Fulde, G.; Moulton, B.; Nadauld, L.D.; Rhodes, T. An elevated neutrophil-to-lymphocyte ratio associates with weight loss and cachexia in cancer. Sci. Rep. 2020, 10, 7535. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Song, M.M.; Zhang, X.; Ding, J.S.; Ruan, G.T.; Zhang, X.W.; Liu, T.; Yang, M.; Ge, Y.-Z.; Tang, M.; et al. Association of systemic inflammation with survival in patients with cancer cachexia: Results from a multicentre cohort study. J. Cachexia Sarcopenia Muscle 2021, 12, 1466–1476. [Google Scholar] [CrossRef]
- Kazemi-Bajestani, S.M.R.; Becher, H.; Butts, C.; Basappa, N.S.; Smylie, M.; Joy, A.A.; Sangha, R.; Gallivan, A.; Kavsak, P.; Chu, Q.; et al. Rapid atrophy of cardiac left ventricular mass in patients with non-small cell carcinoma of the lung. J. Cachexia Sarcopenia Muscle 2019, 10, 1070–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Li, B.; Xu, Y.; Meng, S.; Wang, Y.; Jiang, Y. Luteolin reduces cancerinduced skeletal and cardiac muscle atrophy in a Lewis lung cancer mouse model. Oncol. Rep. 2018, 40, 1129–1137. [Google Scholar]
- Aquila, G.; Re Cecconi, A.D.; Forti, M.; Frapolli, R.; Bello, E.; Novelli, D.; Russo, I.; Licandro, S.A.; Staszewsky, L.; Martinelli, G.B.; et al. Trabectedin and Lurbinectedin Extend Survival of Mice Bearing C26 Colon Adenocarcinoma, without Affecting Tumor Growth or Cachexia. Cancers 2020, 12, 2312. [Google Scholar] [CrossRef]
- Shadfar, S.; Couch, M.E.; McKinney, K.A.; Weinstein, L.J.; Yin, X.; Rodriguez, J.E.; Guttridge, D.C.; Willis, M. Oral resveratrol therapy inhibits cancer-induced skeletal muscle and cardiac atrophy in vivo. Nutr. Cancer 2011, 63, 749–762. [Google Scholar] [CrossRef] [Green Version]
- Willis, M.S.; Rojas, M.; Li, L.; Selzman, C.H.; Tang, R.H.; Stansfield, W.E.; Rodriguez, J.E.; Glass, D.J.; Patterson, C. Muscle ring finger 1 mediates cardiac atrophy in vivo. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H997–H1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smuder, A.J.; Roberts, B.M.; Wiggs, M.P.; Kwon, O.S.; Yoo, J.K.; Christou, D.D.; Fuller, D.D.; Szeto, H.H.; Judge, A.R. Pharmacological targeting of mitochondrial function and reactive oxygen species production prevents colon 26 cancer-induced cardiorespiratory muscle weakness. Oncotarget 2020, 11, 3502–3514. [Google Scholar] [CrossRef] [PubMed]
- Clark, Y.Y.; Wold, L.E.; Szalacha, L.A.; McCarthy, D.O. Ubiquinol reduces muscle wasting but not fatigue in tumor-bearing mice. Biol. Res. Nurs. 2015, 17, 321–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenbaum, L.M.; Sutherland, J.H. Host cathepsin D response to tumor in the normal and pepstatin-treated mouse. Cancer Res. 1983, 43, 2584–2587. [Google Scholar]
- Roberg, K. Relocalization of cathepsin D and cytochrome c early in apoptosis revealed by immunoelectron microscopy. Lab Investig. 2001, 81, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Muscaritoli, M.; Costelli, P.; Bossola, M.; Grieco, G.; Bonelli, G.; Bellantone, R.; Doglietto, G.B.; Fanelli, F.R.; Baccino, F.M. Effects of simvastatin administration in an experimental model of cancer cachexia. Nutrition 2003, 19, 936–939. [Google Scholar] [CrossRef]
- Palus, S.; von Haehling, S.; Flach, V.C.; Tschirner, A.; Doehner, W.; Anker, S.D.; Springer, J. Simvastatin reduces wasting and improves cardiac function as well as outcome in experimental cancer cachexia. Int. J. Cardiol. 2013, 168, 3412–3418. [Google Scholar] [CrossRef]
- Stevens, S.C.; Velten, M.; Youtz, D.J.; Clark, Y.; Jing, R.; Reiser, P.J.; Bicer, S.; Devine, R.D.; McCarthy, D.O.; Wold, L.E.; et al. Losartan treatment attenuates tumor-induced myocardial dysfunction. J. Mol. Cell. Cardiol. 2015, 85, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Kelm, N.Q.; Straughn, A.R.; Kakar, S.S. Withaferin A attenuates ovarian cancer-induced cardiac cachexia. PLoS ONE 2020, 15, e0236680. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, J.L.; Lu, J.; Song, Y.; Kwak, K.S.; Jiao, Q.; Rosenfeld, R.; Chen, Q.; Boone, T.; Simonet, W.S.; et al. Reversal of cancer cachexia and muscle wasting by ActRIIB antagonism leads to prolonged survival. Cell 2010, 142, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Trobec, K.; Palus, S.; Tschirner, A.; von Haehling, S.; Doehner, W.; Lainscak, M.; Anker, S.D.; Springer, J. Rosiglitazone reduces body wasting and improves survival in a rat model of cancer cachexia. Nutrition 2014, 30, 1069–1075. [Google Scholar] [CrossRef] [PubMed]
- Toneto, A.T.; Ferreira Ramos, L.A.; Salomao, E.M.; Tomasin, R.; Aereas, M.A.; Gomes-Marcondes, M.C. Nutritional leucine supplementation attenuates cardiac failure in tumour-bearing cachectic animals. J. Cachexia Sarcopenia Muscle 2016, 7, 577–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nukaga, S.; Mori, T.; Miyagawa, Y.; Fujiwara-Tani, R.; Sasaki, T.; Fujii, K.; Mori, S.; Goto, K.; Kishi, S.; Nakashima, C.; et al. Combined administration of lauric acid and glucose improved cancer-derived cardiac atrophy in a mouse cachexia model. Cancer Sci. 2020, 111, 4605–4615. [Google Scholar] [CrossRef]
- Chance, W.T.; Cao, L.; Zhang, F.S.; Fischer, J.E. Clenbuterol plus acivicin decrease tumor growth and increase muscle mass in rats maintained on total parenteral nutrition. Am. J. Surg. 1991, 161, 51–56. [Google Scholar] [CrossRef]
- Saitoh, M.; Hatanaka, M.; Konishi, M.; Ishida, J.; Palus, S.; Ebner, N.; Döhner, W.; Von Haehling, S.; Anker, S.D.; Springer, J. Erythropoietin improves cardiac wasting and outcomes in a rat model of liver cancer cachexia. Int. J. Cardiol. 2016, 218, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Springer, J.; Tschirner, A.; Hartman, K.; von Haehling, S.; Anker, S.D.; Doehner, W. The xanthine oxidase inhibitor oxypurinol reduces cancer cachexia-induced cardiomyopathy. Int. J. Cardiol. 2013, 168, 3527–3531. [Google Scholar] [CrossRef] [PubMed]
- Elkina, Y.; Palus, S.; Tschirner, A.; Hartmann, K.; von Haehling, S.; Doehner, W.; Mayer, U.; Coats, A.J.; Beadle, J.; Anker, S.D.; et al. Tandospirone reduces wasting and improves cardiac function in experimental cancer cachexia. Int. J. Cardiol. 2013, 170, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.R.; da Cunha, T.F.; da Paixao, N.A.; Brum, P.C. Aerobic exercise training as therapy for cardiac and cancer cachexia. Life Sci. 2015, 125, 9–14. [Google Scholar] [CrossRef]
- Parry, T.L.; Hayward, R. Exercise Protects against Cancer-induced Cardiac Cachexia. Med. Sci. Sports Exerc. 2018, 50, 1169–1176. [Google Scholar] [CrossRef]
- Ranjbar, K.; Ballaro, R.; Bover, Q.; Pin, F.; Beltra, M.; Penna, F.; Costelli, P. Combined Exercise Training Positively Affects Muscle Wasting in Tumor-Bearing Mice. Med. Sci. Sports Exerc. 2019, 51, 1387–1395. [Google Scholar] [CrossRef]
- Fernandes, L.G.; Tobias, G.C.; Paixao, A.O.; Dourado, P.M.; Voltarelli, V.A.; Brum, P.C. Exercise training delays cardiac remodeling in a mouse model of cancer cachexia. Life Sci. 2020, 260, 118392. [Google Scholar] [CrossRef]
- Padrao, A.I.; Moreira-Goncalves, D.; Oliveira, P.A.; Teixeira, C.; Faustino-Rocha, A.I.; Helguero, L.; Vitorino, R.; Santos, L.L.; Amado, F.; Duarte, J.A.; et al. Endurance training prevents TWEAK but not myostatin-mediated cardiac remodelling in cancer cachexia. Arch. Biochem. Biophys. 2015, 567, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Winbanks, C.E.; Murphy, K.T.; Bernardo, B.C.; Qian, H.; Liu, Y.; Sepulveda, P.V.; Beyer, C.; Hagg, A.; Thomson, R.E.; Chen, J.L.; et al. Smad7 gene delivery prevents muscle wasting associated with cancer cachexia in mice. Sci. Transl. Med. 2016, 8, 348ra98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Yang, Q.; Zhang, H.; Wan, L.; Xin, B.; Cao, Y.; Zhang, J.; Guo, C. Cryptotanshinone prevents muscle wasting in CT26-induced cancer cachexia through inhibiting STAT3 signaling pathway. J. Ethnopharmacol. 2020, 260, 113066. [Google Scholar] [CrossRef] [PubMed]
- Devine, R.D.; Eichenseer, C.M.; Wold, L.E. Minocycline attenuates cardiac dysfunction in tumor-burdened mice. J. Mol. Cell. Cardiol. 2016, 100, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Springer, J.; Tschirner, A.; Haghikia, A.; von Haehling, S.; Lal, H.; Grzesiak, A.; Kaschina, E.; Palus, S.; Pötsch, M.; von Websky, K.; et al. Prevention of liver cancer cachexia-induced cardiac wasting and heart failure. Eur. Heart J. 2014, 35, 932–941. [Google Scholar] [CrossRef]
- Musolino, V.; Palus, S.; Latouche, C.; Gliozzi, M.; Bosco, F.; Scarano, F.; Nucera, S.; Carresi, C.; Scicchitano, M.; von Haehling, S.; et al. Cardiac expression of neutrophil gelatinase-associated lipocalin in a model of cancer cachexia-induced cardiomyopathy. ESC Heart Fail. 2019, 6, 89–97. [Google Scholar] [CrossRef]
- VanderVeen, B.N.; Hardee, J.P.; Fix, D.K.; Carson, J.A. Skeletal muscle function during the progression of cancer cachexia in the male Apc(Min/+) mouse. J. Appl. Physiol. 2018, 124, 684–695. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.G.; Gomes-Marcondes, M.C. Metformin treatment modulates the tumour-induced wasting effects in muscle protein metabolism minimising the cachexia in tumour-bearing rats. BMC Cancer 2016, 16, 418. [Google Scholar] [CrossRef] [Green Version]
- Bora, V.; Patel, B.M. Investigation into the role of anti-diabetic agents in cachexia associated with metastatic cancer. Life Sci. 2021, 274, 119329. [Google Scholar] [CrossRef]
- Musolino, V.; Palus, S.; Tschirner, A.; Drescher, C.; Gliozzi, M.; Carresi, C.; Vitale, C.; Muscoli, C.; Doehner, W.; Von Haehling, S.; et al. Megestrol acetate improves cardiac function in a model of cancer cachexia-induced cardiomyopathy by autophagic modulation. J. Cachexia Sarcopenia Muscle 2016, 7, 555–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toledo, M.; Springer, J.; Busquets, S.; Tschirner, A.; Lopez-Soriano, F.J.; Anker, S.D.; López-Soriano, F.J.; Anker, S.D.; Argilés, J.M. Formoterol in the treatment of experimental cancer cachexia: Effects on heart function. J. Cachexia Sarcopenia Muscle 2014, 5, 315–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojima, C.; Noguchi, Y.; Miyamoto, T.; Saito, Y.; Orihashi, H.; Yoshimatsu, Y.; Watabe, T.; Takayama, K.; Hayashi, Y.; Itoh, F. Peptide-2 from mouse myostatin precursor protein alleviates muscle wasting in cancer-associated cachexia. Cancer Sci. 2020, 111, 2954–2964. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.M.; Dillon, E.L.; Kinsky, M.; Chamberlain, A.; McCammon, S.; Jupiter, D.; Willis, M.; Hatch, S.; Richardson, G.; Danesi, C.; et al. Effects of adjunct testosterone on cardiac morphology and function in advanced cancers: An ancillary analysis of a randomized controlled trial. BMC Cancer 2019, 19, 778. [Google Scholar] [CrossRef] [PubMed]
- Solheim, T.S.; Fearon, K.C.; Blum, D.; Kaasa, S. Non-steroidal anti-inflammatory treatment in cancer cachexia: A systematic literature review. Acta Oncol. 2013, 52, 6–17. [Google Scholar] [CrossRef] [Green Version]
- Martins, H.A.; Sehaber, C.C.; Hermes-Uliana, C.; Mariani, F.A.; Guarnier, F.A.; Vicentini, G.E.; Bossolani, G.D.P.; Jussani, L.A.; Lima, M.M.; Bazotte, R.B.; et al. Supplementation with L-glutamine prevents tumor growth and cancer-induced cachexia as well as restores cell proliferation of intestinal mucosa of Walker-256 tumor-bearing rats. Amino Acids 2016, 48, 2773–2784. [Google Scholar] [CrossRef]
- Ham, D.J.; Murphy, K.T.; Chee, A.; Lynch, G.S.; Koopman, R. Glycine administration attenuates skeletal muscle wasting in a mouse model of cancer cachexia. Clin. Nutr. 2014, 33, 448–458. [Google Scholar] [CrossRef]
- Togni, V.; Ota, C.C.; Folador, A.; Junior, O.T.; Aikawa, J.; Yamazaki, R.K.; Freitas, F.A.; Longo, R.; Martins, E.F.; Calder, P.C.; et al. Cancer cachexia and tumor growth reduction in Walker 256 tumor-bearing rats supplemented with N-3 polyunsaturated fatty acids for one generation. Nutr. Cancer 2003, 46, 52–58. [Google Scholar] [CrossRef]
- Liu, S.; Wu, H.J.; Zhang, Z.Q.; Chen, Q.; Liu, B.; Wu, J.P.; Zhu, L. L-carnitine ameliorates cancer cachexia in mice by regulating the expression and activity of carnitine palmityl transferase. Cancer Biol. Ther. 2011, 12, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Cella, P.S.; Marinello, P.C.; Borges, F.H.; Ribeiro, D.F.; Chimin, P.; Testa, M.T.J.; Guirro, P.B.; Duarte, J.A.; Cecchini, R.; Guarnier, F.A.; et al. Creatine supplementation in Walker-256 tumor-bearing rats prevents skeletal muscle atrophy by attenuating systemic inflammation and protein degradation signaling. Eur. J. Nutr. 2020, 59, 661–669. [Google Scholar] [CrossRef]

{kind=link}
| Treatment | Mechanism | Benefit | Reference | |
|---|---|---|---|---|
| NF-κB Pathway Inhibitors | Compound A, NF-κB essential modulator, Luteolin | NF-κB inhibiting agents | Preserved cardiac mass and EF and reduction of inflammatory markers | [27,48] |
| Trabectedin, Lurbinectedin | Inhibits cytokine activation of NF-κB | Some survival benefit but no clear impact on NF-κB signaling in cardiac tissue | [49] | |
| Resveratrol | Inhibition of inflammatory pathways and improvement in myocardial calcium handling | Reduction in cardiac weight loss and preservation of anterior wall thickness | [50,51] | |
| ROS Inhibitors | SS-31 | Antioxidant that reduces ROS in mitochondria | Restored LV function, reduced proteolytic Calpain activity in heart | [52] |
| Ubiquinol | Antioxidant involved in ROS modulation | Increased muscle mass; however, did not improve LV diameter or protein degradation | [53] | |
| Pepstatin | Inhibition of lysosomal protease and oxidative stress | Reduced muscle degradation, but no clear impact on myocardial function | [54,55] | |
| Cardiovascular Drugs | Simvastatin | Decreases activity of matrix metalloproteinase-9 and reduces activity of various inflammatory markers | Decreased weight loss, improved LVEF, and increased SV | [56,57] |
| Bisoprolol and spironolactone | Beta receptor blockade and aldosterone inhibitor respectively | Preserved LV mass, body weight; improved LVEF; reduced cardiac fibrosis | [55] | |
| Losartan and Withaferin A | Angiotensin II inhibition | Preserved EF and SV; reduced fibrotic deposition | [58,59] | |
| Formoterol | B2 selective agonist | Non-significant increase in cardiac weight; significant increase in end-diastolic and systolic volumes | [60] | |
| Nutrition and Appetite | Rosiglitazone | Insulin sensitizer | Improved LVEF and cardiac output; decreased muscle wasting | [61] |
| Megestrol acetate | Appetite stimulant | Increased weight gain; improved LVEF | [49] | |
| Leucine | Decreased levels of chymotrypsin, myeloperoxidase, and caspase 3 and 7 | Improved myocardial function | [62] | |
| Lauric acid and Glucose | Reduced mitochondrial dysfunction and oxidative stress | Reduced myocardial atrophy and improved muscle maturity | [63] | |
| Total parenteral nutrition (TPN) | Intravenous nutrition supplementation | Increased cardiac mass | [64] | |
| Other Categories | EPO | Possible decrease in trypsin levels | Increase in cardiac weight, stroke volume, and physical activity | [65] |
| Oxypurinol | Xanthine oxidase inhibitor | Increased LVEF, total cardiac weight, and cardiac output | [66] | |
| Tandospirone | Antidepressant; serotonin receptor agonist | Preserved muscle mass, improvement in LV mass and EF; some survival benefit | [67] | |
| Testosterone | Unclear | Increase in SV and LVEF | [67] | |
| Exercise | Unclear; possible reduction in cardiac autophagy and NF-κB signaling | Impeded tumor growth, delayed onset of anorexia, improved EF | [68,69,70,71,72] | |
| Gene Therapy via Viral Vector | Upregulating SMAD7, known to inhibit overactivation of procachetic factors | Reduced skeletal and cardiac muscle atrophy | [73] | |
| Crytotanshinone | STAT3 inhibition | Decreased myocardial mass loss, body weight loss, and muscle wasting | [74] | |
| Minocycline | Matrix metalloproteinase inhibitor | Improved FS and EF | [75] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vudatha, V.; Devarakonda, T.; Liu, C.; Freudenberger, D.C.; Riner, A.N.; Herremans, K.M.; Trevino, J.G. Review of Mechanisms and Treatment of Cancer-Induced Cardiac Cachexia. Cells 2022, 11, 1040. https://doi.org/10.3390/cells11061040
Vudatha V, Devarakonda T, Liu C, Freudenberger DC, Riner AN, Herremans KM, Trevino JG. Review of Mechanisms and Treatment of Cancer-Induced Cardiac Cachexia. Cells. 2022; 11(6):1040. https://doi.org/10.3390/cells11061040
Chicago/Turabian StyleVudatha, Vignesh, Teja Devarakonda, Christopher Liu, Devon C. Freudenberger, Andrea N. Riner, Kelly M. Herremans, and Jose G. Trevino. 2022. "Review of Mechanisms and Treatment of Cancer-Induced Cardiac Cachexia" Cells 11, no. 6: 1040. https://doi.org/10.3390/cells11061040
APA StyleVudatha, V., Devarakonda, T., Liu, C., Freudenberger, D. C., Riner, A. N., Herremans, K. M., & Trevino, J. G. (2022). Review of Mechanisms and Treatment of Cancer-Induced Cardiac Cachexia. Cells, 11(6), 1040. https://doi.org/10.3390/cells11061040