
Assessment of Experimental Techniques That Facilitate Human Granuloma Formation in an In Vitro System: A Systematic Review



Abstract
1. Introduction
2. Methodology
3. Results
3.1. General Overview of the Included Studies
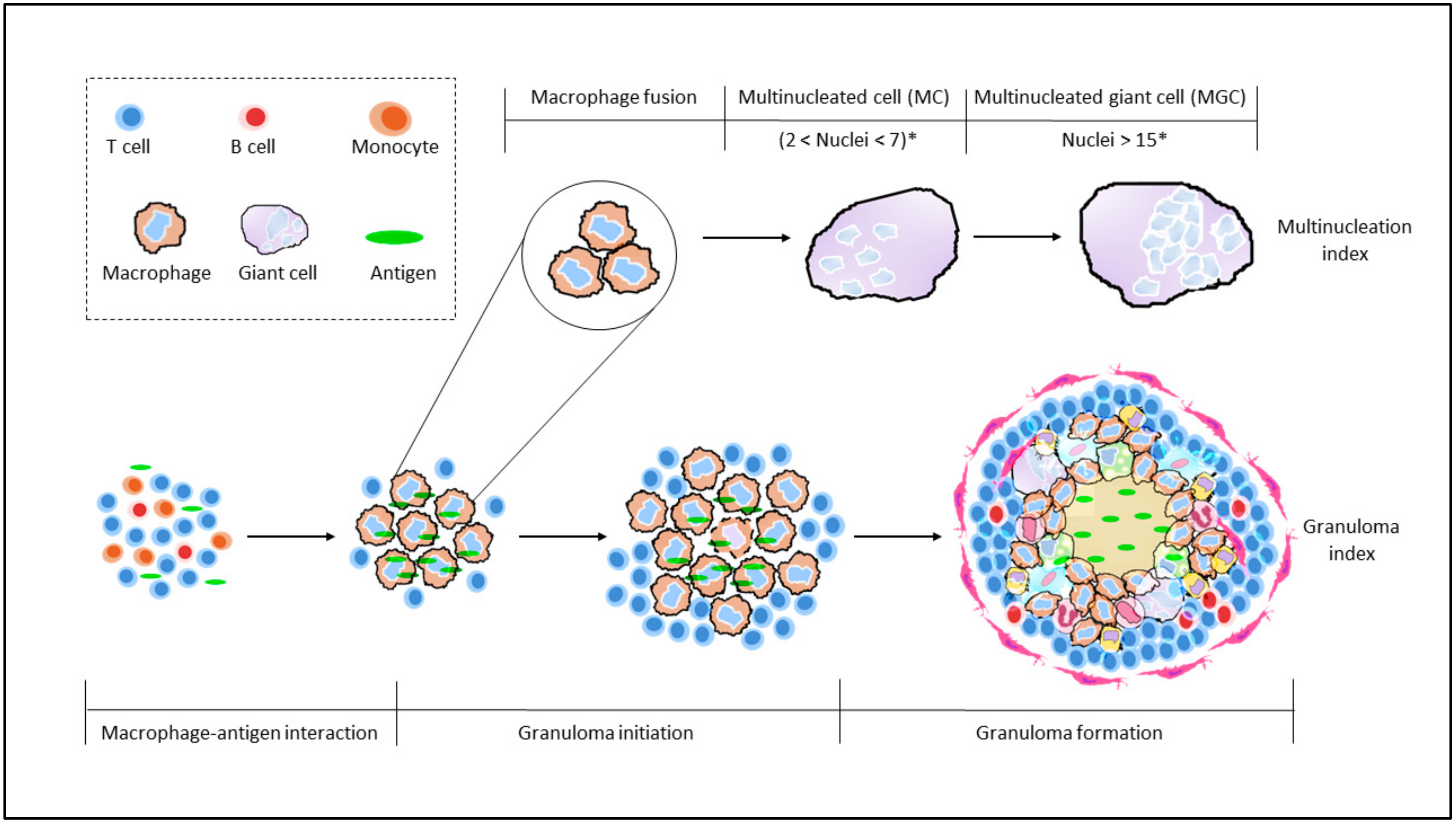
3.2. Granuloma Indices
3.3. Maturation of Granulomas
Lymphocyte Transformation Test (LTT)
3.4. Cell Surface Markers
3.4.1. T Cells
3.4.2. B Cells
3.4.3. Monocytes
3.4.4. Macrophages
3.4.5. M1 Macrophages
3.4.6. M2 Macrophages
- CD209+ [51] expression remained unchanged.
3.4.7. M1/M2 Macrophage Polarization
3.4.8. Remarks Concerning the Use of Cell Surface Markers
- Changes in expression of selected markers was assessed via either microscopy or flow cytometric analysis.
- A considerable variability of similar markers was used based on the focus of each study.
- None of the Schistosoma antigen-focused studies assessed expression of cell surface markers.
3.5. Cytokine Secretion Profile
3.5.1. Multiple Cell Types
3.5.2. Th1
3.5.3. Th2
3.5.4. Th1/Th2 Balance
3.5.5. Th17
3.5.6. Changes over Time
4. Discussion
4.1. Cells/Culture Conditions
4.2. Indexing Granuloma Development
- Across all indices, a specific threshold is lacking to differentiate clusters of cells from the formation of (well-defined) granulomas. Exposing cells of healthy controls, an index range of 2-2.5 was observed with GI-B [35] and GI-S [22], while a score of 3-4 is commonly observed with GI-B in infected persons.
- Taking into consideration that the maximum score of 5 or 6 with respect to granuloma index GI-B and GI-S were rarely observed, it raises the question of whether increasing the length of experiment after exposure might give rise to a more well-defined granuloma.
- Both indices currently lack definition on the number of granulomas to be expected after exposure.
- A dose-dependent increase in the number of granulomas has thus far only been reported with non-infectious antigens [25], independently from GI.
- GI is only focused on the overall structural evolution of granuloma and does not consider, e.g., the maturation of macrophages to MGC.
4.3. Relationship between LTT and Granuloma Index
4.4. Cell-Specific Markers
4.5. Cytokine Secretion Profiles
4.6. Limitations of Our Study
4.7. Future Directions
5. Conclusions
6. Infobox: Important Technical Considerations
6.1. Agents
6.2. Use of Beads
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaye, P. Granulomatous diseases. Int. J. Exp. Pathol. 2004, 81, 289–290. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Rueda, N.; Albassier, M.; Allain, S.; Deknuydt, F.; Altare, F.; Le Pape, P. First Human Model of In Vitro Candida albicans Persistence within Granuloma for the Reliable Study of Host-Fungi Interactions. PLoS ONE 2012, 7, e40185. [Google Scholar] [CrossRef]
- Teskey, G.; Cao, R.; Islamoglu, H.; Medina, A.; Prasad, C.; Prasad, R.; Sathananthan, A.; Fraix, M.; Subbian, S.; Zhong, L.; et al. The Synergistic Effects of the Glutathione Precursor, NAC and First-Line Antibiotics in the Granulomatous Response against Mycobacterium tuberculosis. Front. Immunol. 2018, 9, 2069. [Google Scholar] [CrossRef] [PubMed]
- Saunders, B.M.; Frank, A.; Orme, I.M. Granuloma formation is required to contain bacillus growth and delay mortality in mice chronically infected with Mycobacterium tuberculosis. Immunology 1999, 98, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Mezouar, S.; Diarra, I.; Roudier, J.; Desnues, B.; Mege, J.-L. Tumor Necrosis Factor-Alpha Antagonist Interferes With the Formation of Granulomatous Multinucleated Giant Cells: New Insights into Mycobacterium tuberculosis Infection. Front. Immunol. 2019, 10, 1947. [Google Scholar] [CrossRef]
- Fabrellas, E.F. Epidemiología de la sarcoidosis. Arch. Bronconeumol. 2007, 43, 92–100. [Google Scholar] [CrossRef]
- Sforza, G.G.R.; Marinou, A. Hypersensitivity pneumonitis: A complex lung disease. Clin. Mol. Allergy 2017, 15, 1–8. [Google Scholar] [CrossRef]
- Newman, K.; Newman, L.S. Occupational causes of sarcoidosis. Curr. Opin. Allergy Clin. Immunol. 2012, 12, 145–150. [Google Scholar] [CrossRef]
- Eguirado, E.; Schlesinger, L.S. Modeling the Mycobacterium tuberculosis Granuloma – the Critical Battlefield in Host Immunity and Disease. Front. Immunol. 2013, 4, 98. [Google Scholar] [CrossRef]
- Je, S.; Quan, H.; Na, Y.; Cho, S.-N.; Kim, B.-J.; Seok, S.H. An in vitro model of granuloma-like cell aggregates substantiates early host immune responses against Mycobacterium massiliense infection. Biol. Open 2016, 5, 1118–1127. [Google Scholar] [CrossRef]
- Saunders, B.M.; Cooper, A. Restraining mycobacteria: Role of granulomas in mycobacterial infections. Immunol. Cell Biol. 2000, 78, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Maeda, Y.; Fukutomi, Y.; Makino, M. An in vitro model of Mycobacterium lepraeinduced granuloma formation. BMC Infect. Dis. 2013, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Boros, D.L. Experimental granulomatosis. Clin. Dermatol. 1986, 4, 10–21. [Google Scholar] [CrossRef]
- Franklin, G.; Coghill, G.; McIntosh, L.; Cree, I. Monocyte aggregation around agarose beads in collagen gels: A 3-dimensional model of early granuloma formation? J. Immunol. Methods 1995, 186, 285–291. [Google Scholar] [CrossRef]
- Mathew, D.G.; Sreenivasan, B.; Varghese, S.S.; Sebastian, C.J. Classification of Giant Cell Lesions of the Oral Cavity: A Fresh Perspective. OMPJ 2016, 7. [Google Scholar] [CrossRef]
- Tristão, F.S.M.; Rocha, F.A.; Carlos, D.; Ketelut-Carneiro, N.; Souza, C.; Milanezi, C.M.; Silva, J.S. Th17-Inducing Cytokines IL-6 and IL-23 Are Crucial for Granuloma Formation during Experimental Paracoccidioidomycosis. Front. Immunol. 2017, 8, 949. [Google Scholar] [CrossRef]
- Shah, K.K.; Pritt, B.S.; Alexander, M.P. Histopathologic review of granulomatous inflammation. J. Clin. Tuberc. Other Mycobact. Dis. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Torre, O.; Elia, D.; Caminati, A.; Harari, S. New insights in lymphangioleiomyomatosis and pulmonary Langerhans cell histiocytosis. Eur. Respir. Rev. 2017, 26, 170042. [Google Scholar] [CrossRef]
- Ahmadzai, H.; Huang, S.; Steinfort, C.; Markos, J.; Allen, R.K.; Wakefield, D.; Wilsher, M.; Thomas, P.S. Sarcoidosis: A state of the art review from the Thoracic Society of Australia and New Zealand. Med J. Aust. 2018, 208, 499–504. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.-A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage Plasticity, Polarization, and Function in Health and Disease. J. Cell Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Gupta, U.D.; Singh, A.K. Animal models of tuberculosis: Lesson learnt. Indian J. Med Res. 2018, 147, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Guirado, E.; Mbawuike, U.; Keiser, T.L.; Arcos, J.; Azad, A.K.; Wang, S.-H.; Schlesinger, L.S. Characterization of Host and Microbial Determinants in Individuals with Latent Tuberculosis Infection Using a Human Granuloma Model. MBio. 2015, 6, e02537. [Google Scholar] [CrossRef] [PubMed]
- Crouser, E.D.; White, P.; Caceres, E.G.; Julian, M.W.; Papp, A.C.; Locke, L.W.; Sadee, W.; Schlesinger, L.S. A NovelIn VitroHuman Granuloma Model of Sarcoidosis and Latent Tuberculosis Infection. Am. J. Respir. Cell Mol. Biol. 2017, 57, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Deknuydt, F.; Roquilly, A.; Cinotti, R.; Altare, F.; Asehnoune, K. An In Vitro Model of Mycobacterial Granuloma to Investigate the Immune Response in Brain-Injured Patients. Crit. Care Med. 2013, 41, 245–254. [Google Scholar] [CrossRef] [PubMed]
- De Chambrun, G.P.; Body-Malapel, M.; Frey-Wagner, I.; Djouina, M.; Deknuydt, F.; Atrott, K.; Esquerre, N.; Altare, F.; Neut, C.; Arrieta, M.C.; et al. Aluminum enhances inflammation and decreases mucosal healing in experimental colitis in mice. Mucosal Immunol. 2013, 7, 589–601. [Google Scholar] [CrossRef]
- Heinemann, D.E.H.; Peters, J.H.; Gahr, M. A human in vitro granuloma model using heat killed Candida albicans cells immobilized on plastic culture wells. Scand. J. Immunol. 1997, 45, 596–604. [Google Scholar] [CrossRef]
- Lay, G.; Poquet, Y.; Salek-Peyron, P.; Puissegur, M.-P.; Botanch, C.; Bon, H.; Levillain, F.; Duteyrat, J.-L.; Emile, J.-F.; Altare, F. Langhans giant cells fromM. tuberculosis-induced human granulomas cannot mediate mycobacterial uptake. J. Pathol. 2006, 211, 76–85. [Google Scholar] [CrossRef]
- Puissegur, M.-P.; Lay, G.; Gilleron, M.; Botella, L.; Nigou, J.; Marrakchi, H.; Mari, B.; Duteyrat, J.-L.; Guerardel, Y.; Kremer, L.; et al. Mycobacterial Lipomannan Induces Granuloma Macrophage Fusion via a TLR2-Dependent, ADAM9- and β1Integrin-Mediated Pathway. J. Immunol. 2007, 178, 3161–3169. [Google Scholar] [CrossRef]
- Delcroix, M.; Heydari, K.; Dodge, R.; Riley, L.W. Flow-cytometric analysis of human monocyte subsets targeted byMycobacterium bovisBCG before granuloma formation. Pathog. Dis. 2018, 76, fty080. [Google Scholar] [CrossRef]
- Da Silva, D.A.A.; Da Silva, M.V.; Barros, C.C.O.; Alexandre, P.B.D.; Timóteo, R.P.; Catarino, J.S.; Sales-Campos, H.; Machado, J.R.; Rodrigues, D.B.R.; Oliveira, C.J.; et al. TNF-α blockade impairs in vitro tuberculous granuloma formation and down modulate Th1, Th17 and Treg cytokines. PLoS ONE 2018, 13, e0194430. [Google Scholar] [CrossRef]
- Doughty, B.L.; Goes, A.M.; Parra, J.C.; Rocha, R.S.; Katz, N.; Colley, D.G.; Gazzinelli, G. Granulomatous hypersensitivity to Schistosoma mansoni egg antigens in human Schistosomiasis: I. Granuloma formation and modulation around polyacrylamide antigen-conjugated beads. Mem. Inst. Oswaldo. Cruz. 1987, 82, 47–54. [Google Scholar] [CrossRef]
- Rezende, S.A.; Silva-Teixeira, D.N.; Drummond, S.C.; Goes, A.M. IL-10 Plays a Role in the Modulation of Human Granulomatous Hypersensitivity Against Schistosoma mansoni Eggs Induced by Immune Complexes. Scand. J. Immunol. 1997, 46, 96–102. [Google Scholar] [CrossRef] [PubMed]
- A Rezende, S.; Lambertucci, J.R.; Goes, A.M. Role of Immune Complexes from Pacients with Different Clinical Forms of Schistosomiasis in the Modulation of In Vitro Granuloma Reaction. Mem. Inst. Oswaldo Cruz 1997, 92, 683–687. [Google Scholar] [CrossRef] [PubMed][Green Version]
- A Almeida, C.; Goes, A.M. The Role of Protein Kinases in Antigen-activation of Peripheral Blood Mononuclear Cells of Schistosoma mansoni Infected Individuals. Mem. Inst. Oswaldo Cruz 1997, 92, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Falcão, P.; Malaquias, L.C.; A Martins-Filho, O.; Silveira, A.M.; Passos, V.M.; Prata, A.; Gazzinelli, G.; Coffman, R.L.; Correa-Oliveira, R. Human Schistosomiasis mansoni: IL-10 modulates thein vitrogranuloma formation. Parasite Immunol. 1998, 20, 447–454. [Google Scholar] [CrossRef]
- Oliveira, D.; Silva-Teixeira, D.; Araújo-Filho, R.; Goes, A. Antigenic stimulation is more efficient than LPS in inducing nitric oxide production by human mononuclear cells on the in vitro granuloma reaction in schistosomiasis. Braz. J. Med Biol. Res. 1999, 32, 1437–1445. [Google Scholar] [CrossRef]
- Rezende, C.M.; Goes, T.S.; Goes, V.S.; Azevedo, V.; Leite, M.; Goes, A.M. GM-CSF and TNF-α synergize to increase in vitro granuloma size of PBMC from humans induced by Schistosoma mansoni recombinant 28-kDa GST. Immunol. Lett. 2004, 95, 221–228. [Google Scholar] [CrossRef]
- Wright, C.C.; Hsu, F.F.; Arnett, E.; Dunaj, J.L.; Davidson, P.M.; Pacheco, S.A.; Harriff, M.J.; Lewinsohn, D.M.; Schlesinger, L.S.; Purdy, G.E. The Mycobacterium tuberculosis MmpL11 Cell Wall Lipid Transporter Is Important for Biofilm Formation, Intracellular Growth, and Nonreplicating Persistence. Infect. Immun. 2017, 85, e00131–17. [Google Scholar] [CrossRef]
- Mehta, M.; Singh, A. Mycobacterium tuberculosis WhiB3 maintains redox homeostasis and survival in response to reactive oxygen and nitrogen species. Free Radic. Biol. Med. 2018, 131, 50–58. [Google Scholar] [CrossRef]
- Hanifin, J.M.; Epstein, W.L.; Cline, M.J. In Vitro Studies of Granulomatous Hypersensitivity to Beryllium. J. Investig. Dermatol. 1970, 55, 284–288. [Google Scholar] [CrossRef]
- Soruri, A.; Schweyer, S.; Radzun, H.-J.; Fayyazi, A. Mycobacterial Antigens Induce Apoptosis in Human Purified Protein Derivative-Specific Alphabeta T Lymphocytes in a Concentration-Dependent Manner. Immunology 2002, 105, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Misme-Aucouturier, B.; Albassier, M.; Alvarez-Rueda, N.; Le Pape, P. Specific Human and Candida Cellular Interactions Lead to Controlled or Persistent Infection Outcomes during Granuloma-Like Formation. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef]
- Seitzer, U.; Gerdes, J. Generation and characterization of multicellular heterospheroids formed by human peripheral blood mononuclear cells. Cells Tissues Organs 2003, 174, 110–116. [Google Scholar] [CrossRef]
- Kapoor, N.; Pawar, S.; Sirakova, T.D.; Deb, C.; Warren, W.L.; Kolattukudy, P.E. Human Granuloma In Vitro Model, for TB Dormancy and Resuscitation. PLoS ONE 2013, 8, e53657. [Google Scholar] [CrossRef] [PubMed]
- Yamashiro, L.H.; Eto, C.; Soncini, M.; Horewicz, V.; Garcia, M.; Schlindwein, A.D.; Grisard, E.C.; Rovaris, D.B.; Báfica, A. Isoniazid-induced control ofMycobacterium tuberculosisby primary human cells requires interleukin-1 receptor and tumor necrosis factor. Eur. J. Immunol. 2016, 46, 1936–1947. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.L.; Chan, J.; Lin, P.L. Macrophages and control of granulomatous inflammation in tuberculosis. Mucosal Immunol. 2011, 4, 271–278. [Google Scholar] [CrossRef]
- Locke, L.W.; Kothandaraman, S.; Tweedle, M.; Chaney, S.; Wozniak, D.J.; Schlesinger, L.S. Use of a leukocyte-targeted peptide probe as a potential tracer for imaging the tuberculosis granuloma. Tuberculosis 2018, 108, 201–210. [Google Scholar] [CrossRef]
- Zhang, C.; Chery, S.; Lazerson, A.; Altman, N.H.; Jackson, R.; Holt, G.; Campos, M.; Schally, A.V.; Mirsaeidi, M. Anti-inflammatory effects of α-MSH through p-CREB expression in sarcoidosis like granuloma model. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Huang, Z.; Luo, Q.; Guo, Y.; Chen, J.; Xiong, G.; Peng, Y.; Ye, J.; Li, J. Mycobacterium tuberculosis-Induced Polarization of Human Macrophage Orchestrates the Formation and Development of Tuberculous Granulomas In Vitro. PLoS ONE 2015, 10, e0129744. [Google Scholar] [CrossRef]
- Arbués, A.; Brees, D.; Chibout, S.-D.; Fox, T.; Kammüller, M.; Portevin, D. TNF-α antagonists differentially induce TGF-β1-dependent resuscitation of dormant-like Mycobacterium tuberculosis. PLoS Pathog. 2020, 16, e1008312. [Google Scholar] [CrossRef]
- Tian, W.-W.; Wang, Q.-Q.; Liu, W.-D.; Shen, J.-P.; Wang, H.-S. Mycobacterium marinum: A potential immunotherapy for Mycobacterium tuberculosis infection. Drug Des. Dev. Ther. 2013, 7, 669–680. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Effros, R.B.; Dillard, L.; Zeller, E.; Naeim, F.; Walford, R.L. Strong HLA-DR expression in T cell cultures after activation is necessary for IL-2-dependent proliferation. Hum. Immunol. 1983, 8, 249–254. [Google Scholar] [CrossRef]
- Jeannin, P.; Magistrelli, G.; Aubry, J.-P.; Caron, G.; Gauchat, J.-F.; Renno, T.; Herbault, N.; Goetsch, L.; Blaecke, A.; Dietrich, P.-Y.; et al. Soluble CD86 Is a Costimulatory Molecule for Human T Lymphocytes. Immunity 2000, 13, 303–312. [Google Scholar] [CrossRef]
- Locke, L.W.; Crouser, E.D.; White, P.; Julian, M.W.; Caceres, E.G.; Papp, A.C.; Le, V.T.; Sadee, W.; Schlesinger, L.S. IL-13–regulated Macrophage Polarization during Granuloma Formation in an In Vitro Human Sarcoidosis Model. Am. J. Respir. Cell Mol. Biol. 2019, 60, 84–95. [Google Scholar] [CrossRef]
- A Birkness, K.; Guarner, J.; Sable, S.; Tripp, R.; Kellar, K.L.; Bartlett, J.; Quinn, F.D. An in vitro model of the leukocyte interactions associated with granuloma formation in Mycobacterium tuberculosis infection. Immunol. Cell Biol. 2007, 85, 160–168. [Google Scholar] [CrossRef]
- Alingrin, J.; Coiffard, B.; Textoris, J.; Belenotti, P.; Daumas, A.; Leone, M.; Mege, J.-L. Impaired Granuloma Formation in Sepsis: Impact of Monocytopenia. PLoS ONE 2016, 11, e0158528. [Google Scholar] [CrossRef]
- Daumas, A.; Coiffard, B.; Chartier, C.; Ben Amara, A.; Alingrin, J.; Villani, P.; Mege, J.-L. Defective Granuloma Formation in Elderly Infected Patients. Front. Cell. Infect. Microbiol. 2020, 10, 189. [Google Scholar] [CrossRef]
- Abouel-Nour, M.F.; Lotfy, M.; Attallah, A.M.; Doughty, B.L. Schistosoma mansoni major egg antigen Smp40: Molecular modeling and potential immunoreactivity for anti-pathology vaccine development. Mem. Inst. Oswaldo Cruz 2006, 101, 365–372. [Google Scholar] [CrossRef]
- Crouser, E.D.; Locke, L.W.; Julian, M.W.; Bicer, S.; Sadee, W.; White, P.; Schlesinger, L.S. Phagosome-regulated mTOR signalling during sarcoidosis granuloma biogenesis. Eur. Respir. J. 2020, 57, 2002695. [Google Scholar] [CrossRef]
- Elkington, P.; Lerm, M.; Kapoor, N.; Mahon, R.; Pienaar, E.; Huh, D.; Kaushal, D.; Schlesinger, L.S. In Vitro Granuloma Models of Tuberculosis: Potential and Challenges. J. Infect. Dis. 2019, 219, 1858–1866. [Google Scholar] [CrossRef]
- Cline, M.J.; Swett, V.C. The interaction of human monocytes and lymphocytes. J. Exp. Med. 1968, 128, 1309–1325. [Google Scholar] [CrossRef]
- Taflin, C.; Miyara, M.; Nochy, D.; Valeyre, D.; Naccache, J.-M.; Altare, F.; Salek-Peyron, P.; Badoual, C.; Bruneval, P.; Haroche, J.; et al. FoxP3+ Regulatory T Cells Suppress Early Stages of Granuloma Formation but Have Little Impact on Sarcoidosis Lesions. Am. J. Pathol. 2009, 174, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Cao, R.; Teskey, G.; Islamoglu, H.; Gutierrez, M.; Salaiz, O.; Munjal, S.; Fraix, M.P.; Sathananthan, A.; Nieman, D.C.; Venketaraman, V. Flavonoid Mixture Inhibits Mycobacterium tuberculosis Survival and Infectivity. Molecules 2019, 24, 851. [Google Scholar] [CrossRef] [PubMed]
- Islamoglu, H.; Cao, R.; Teskey, G.; Gyurjian, K.; Lucar, S.; Fraix, M.P.; Sathananthan, A.; Chan, J.K.; Venketaraman, V. Effects of ReadiSorb L-GSH in Altering Granulomatous Responses against Mycobacterium tuberculosis Infection. J. Clin. Med. 2018, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- To, K.; Cao, R.; Yegiazaryan, A.; Owens, J.; Nguyen, T.; Sasaninia, K.; Vaughn, C.; Singh, M.; Truong, E.; Medina, A.; et al. Effects of Oral Liposomal Glutathione in Altering the Immune Responses Against Mycobacterium tuberculosis and the Mycobacterium bovis BCG Strain in Individuals With Type 2 Diabetes. Front. Cell. Infect. Microbiol. 2021, 11, 468. [Google Scholar] [CrossRef]
- Chen, D.-Y.; Chen, Y.-M.; Lin, C.-F.; Lo, C.-M.; Liu, H.-J.; Liao, T.-L. MicroRNA-889 Inhibits Autophagy To Maintain Mycobacterial Survival in Patients with Latent Tuberculosis Infection by Targeting TWEAK. MBio. 2020, 11, e03045–19. [Google Scholar] [CrossRef]
- Marakalala, M.J.; Raju, R.; Sharma, K.; Zhang, Y.J.; Eugenin, E.A.; Prideaux, B.; Daudelin, I.B.; Chen, P.-Y.; Booty, M.G.; Kim, J.H.; et al. Inflammatory signaling in human tuberculosis granulomas is spatially organized. Nat. Med. 2016, 22, 531–538. [Google Scholar] [CrossRef]
- Reyes, N.; Del Valle, U.; Bettin, A.; Reyes, I.; Geliebter, J. MICROARRAY ANALYSIS OF THE IN VITRO GRANULOMATOUS RESPONSE TO MYCOBACTERIUM TUBERCULOSIS H37RA. Colomb. Med. 2015, 46, 26–32. [Google Scholar] [CrossRef]
- Puissegur, M.-P.; Botanch, C.; Duteyrat, J.-L.; Delsol, G.; Caratero, C.; Altare, F. An in vitro dual model of mycobacterial granulomas to investigate the molecular interactions between mycobacteria and human host cells. Cell. Microbiol. 2004, 6, 423–433. [Google Scholar] [CrossRef]
- Terrén, I.; Orrantia, A.; Vitallé, J.; Zenarruzabeitia, O.; Borrego, F. CFSE dilution to study human T and NK cell proliferation in vitro. Methods Enzymol. 2020, 631, 239–255. [Google Scholar] [CrossRef]
- Peyron, P.; Vaubourgeix, J.; Poquet, Y.; Levillain, F.; Botanch, C.; Bardou, F.; Daffé, M.; Emile, J.-F.; Marchou, B.; Cardona, P.-J.; et al. Foamy Macrophages from Tuberculous Patients’ Granulomas Constitute a Nutrient-Rich Reservoir for M. tuberculosis Persistence. PLoS Pathog. 2008, 4, e1000204. [Google Scholar] [CrossRef]
- Dkhar, H.K.; Nanduri, R.; Mahajan, S.; Dave, S.; Saini, A.; Somavarapu, A.K.; Arora, A.; Parkesh, R.; Thakur, K.G.; Mayilraj, S.; et al. Mycobacterium tuberculosis Keto-Mycolic Acid and Macrophage Nuclear Receptor TR4 Modulate Foamy Biogenesis in Granulomas: A Case of a Heterologous and Noncanonical Ligand-Receptor Pair. J. Immunol. 2014, 193, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Silva-Miranda, M.; Ekaza, E.; Breiman, A.; Asehnoune, K.; Barros-Aguirre, D.; Pethe, K.; Ewann, F.; Brodin, P.; Ballell-Pages, L.; Altare, F. High-Content Screening Technology Combined with a Human Granuloma Model as a New Approach To Evaluate the Activities of Drugs against Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2015, 59, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Bielecka, M.K.; Tezera, L.B.; Zmijan, R.; Drobniewski, F.; Zhang, X.; Jayasinghe, S.; Elkington, P. A Bioengineered Three-Dimensional Cell Culture Platform Integrated with Microfluidics To Address Antimicrobial Resistance in Tuberculosis. MBio. 2017, 8, e02073–16. [Google Scholar] [CrossRef] [PubMed]
- Arnett, E.; Weaver, A.M.; Woodyard, K.C.; Montoya, M.J.; Li, M.; Hoang, K.V.; Hayhurst, A.; Azad, A.K.; Schlesinger, L.S. PPARγ is critical for Mycobacterium tuberculosis induction of Mcl-1 and limitation of human macrophage apoptosis. PLoS Pathog. 2018, 14, e1007100. [Google Scholar] [CrossRef] [PubMed]
- Mahatha, A.C.; Mal, S.; Majumder, D.; Saha, S.; Ghosh, A.; Basu, J.; Kundu, M. RegX3 Activates whiB3 Under Acid Stress and Subverts Lysosomal Trafficking of Mycobacterium tuberculosis in a WhiB3-Dependent Manner. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Ashley, D.; Hernandez, J.; Cao, R.; To, K.; Yegiazaryan, A.; Abrahem, R.; Nguyen, T.; Owens, J.; Lambros, M.; Subbian, S.; et al. Antimycobacterial Effects of Everolimus in a Human Granuloma Model. J. Clin. Med. 2020, 9, 2043. [Google Scholar] [CrossRef]
- Hernandez, J.; Ashley, D.; Cao, R.; Abrahem, R.; Nguyen, T.; To, K.; Yegiazaryan, A.; David, A.A.; Tiwari, R.K.; Venketaraman, V. Cyclic Peptide [R4W4] in Improving the Ability of First-Line Antibiotics to Inhibit Mycobacterium tuberculosis Inside in vitro Human Granulomas. Front. Immunol. 2020, 11, 1677. [Google Scholar] [CrossRef]
- Sampaio, E.P.; Caneshi, J.R.; A Nery, J.; Duppre, N.C.; Pereira, G.M.; Vieira, L.M.; Moreira, A.L.; Kaplan, G.; Sarno, E.N. Cellular immune response to Mycobacterium leprae infection in human immunodeficiency virus-infected individuals. Infect. Immun. 1995, 63, 1848–1854. [Google Scholar] [CrossRef]
- Jain, S.; Kaur, I.R.; Das, S.; Bhattacharya, S.N.; Singh, A. T helper 1 to T helper 2 shift in cytokine expression: An autoregulatory process in superantigen-associated psoriasis progression? J. Med Microbiol. 2009, 58, 180–184. [Google Scholar] [CrossRef]
- Vuk-Pavlović, Z.; Rohrbach, M.S. An in vitro model for the induction of angiotensin-converting enzyme in sarcoidosis: Possible parallels to the immune response. Clin. Exp. Immunol. 1988, 72, 499–504. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scoring System | Definition | Reference |
|---|---|---|
| Granuloma Index GI-B (Granuloma index-beads supported) | Granuloma formation is initiated and scored in accompany of polyacrylamide beads Cellular reactivity was determined by morphological observation based on the following criteria: the number of cells binding to polyacrylamide beads, visual evidence of blast transforming cells accompanied by cellular migration and adherent cell layers surrounding the beads. Three hundred separate determinations of cellular reactivity were made for each experimental group. A numerical score equivalent to the following classification was assigned to each cell-bead reaction observed:
 Adapted from Figure 1 of Silva DAAd et al., [30] (A–F refers to score 1 to 6) | [30,31,32,33,34,35,36,37] |
| Granuloma Index GI-S (Granuloma index-spontaneous) |  Adapted from Guirado E et al., [23] Granuloma formation in absence of polyacrylamide beads Phase I (Score 1): No cellular aggregation. Phase II (Score 2): Cellular gathering begins with the recruitment of mostly monocytes and some lymphocytes, approximate diameter: <25/50 μm. Phase III (Score 3): Lymphocyte-like cell recruitment increases, approximate diameter: 25/50–100 µm. Phase IV (Score 4): The recruited cells form multi-layers, comprising macrophages and lymphocytes, approximate diameter: 100–200/300 μm. Phase V (Score 5): The cellular structure shows signs of cellular differentiation (multinucleated giant cells and numerous pseudopodia at the cell surface) mimicking mature in vitro granulomas, with large, differentiated monocyte/macrophage-like cells surrounded by lymphocytes, approximate diameter: >200/500 μm. | [22,23,38] |
| Index of maturation | Granuloma formation is scored from small to large size Deknuydt F et al. [24] Index 1: small and poorly differentiated structures Index 2: larger and slightly differentiated structures Index 3: Standard granulomas with a good differentiation of cells Index 4: Very large and highly differentiated structures Pineton de Chambrun G et al. [25]: results are represented as the total number of granulomas per well and the percentage of small size (index 1) and big size granulomas (indexes 2–4). Mehta M et al., [39]: clusters with granulomas < 100 µm were classified as small and >100 µm were classified as large granulomas. | [25,26,38] |
| Multinucleation Index | A total of 300–500 nuclei per culture were counted. | [26] |
| Strengths | |
|---|---|
| Not infection site specific | Able to generate hepatic Schistosoma granulomas [32,33,37] and pulmonary TB [22,23,38] granulomas in vitro from PBMCs with the same cellular conditions |
| Non-invasive and easily manipulatable | Method of acquiring PBMCs from whole blood is not only non-invasive and readily available but also easily manipulatable. There are no qualms with testing against several antigens [28] simultaneously due to this availability. |
| Longitudinal studies | Allows for the continual assessment of antigen manipulation and tracking of changes in host cells daily [22] |
| Host specificity | Able to distinguish for differences in hosts’ immune responses from unexposed controls [3,22,23,31,35,48,57,61,62,63,64,65,66] as well as predict and mimic granuloma in vivo [25,47] |
| Genetic variability | Molecular patterns are able to distinguish responses to antigen, across cohorts of interest [23] |
| Antigen conjugation | Conjugation of antigens to beads allows for a spectrum of antigens to be tested in vitro [30,37] |
| 2-/3-dimensional organization | Able to replicate spatial organization in three dimensions, as observed in human response [54,67] |
| Cell surface markers and cytokines expression | Observations reported reflect published data in literature by recapitulating development in vitro [49] |
| Reactivation/Resuscitation | Resuscitation of dormant M.tb in granulomas under immunosuppressive conditions can be achieved [44] |
| Knowledge applicability | Can be applied to other endpoints such as lymphocyte transformation assay [33,34,37,40,41] and histological analysis [49] |
| Weaknesses | |
| Host–antigen interaction | Complex interactions observed in humans have yet to be achieved in vitro [60] |
| Collagen-matrix models | Extracellular matrix (ECM) may play a role in host–antigen reaction, and collagen matrices have been included in 16% of highlighted studies [3,29,44,45,50,63,64]. Advantages to granuloma maintenance cannot be assessed. |
| Continual cell recruitment | Lack of continual influx of mononuclear phagocytes and lymphocytes to mimic cell recruitment over extended time of exposure [60] |
| Vascularization | Lack of vascularization observed in vitro with PBMCs due to its cellular makeup and hence has not been addressed thus far [50] |
| Opportunities | |
| Comorbidities | Accounting for comorbidities such as diabetes and smoking can facilitate for differences in cohorts [60] |
| Microstructure | Aggregation of granuloma-like structures can be easily distinguished from unexposed controls since it is antigen dependent [44] |
| Sensitization in healthy cohort | Activation or proliferation in an otherwise healthy individual indicates sensitization, which can be determined in vitro with no direct impact on donor |
| Concerns/Future challenges | |
| Modulating infection dosage or antigen concentrations | Impact of high or low dosage/concentration of pathogens/antigens, respectively, has not been assessed but can provide an insight on acute or chronic exposure that is relevant to sarcoidosis. |
| Viability | Cell viability by the end of experiment needs to be accounted for, to determine if cell–cell interactions are antigen dependent or due to cell death. |
| Apoptosis/Necrosis | Assessment of apoptosis and necrosis was only performed in several studies [10,43,44] and requires added focus. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ganesan, N.; Ronsmans, S.; Vanoirbeek, J.; Hoet, P.H.M. Assessment of Experimental Techniques That Facilitate Human Granuloma Formation in an In Vitro System: A Systematic Review. Cells 2022, 11, 864. https://doi.org/10.3390/cells11050864
Ganesan N, Ronsmans S, Vanoirbeek J, Hoet PHM. Assessment of Experimental Techniques That Facilitate Human Granuloma Formation in an In Vitro System: A Systematic Review. Cells. 2022; 11(5):864. https://doi.org/10.3390/cells11050864
Chicago/Turabian StyleGanesan, Nirosha, Steven Ronsmans, Jeroen Vanoirbeek, and Peter H. M. Hoet. 2022. "Assessment of Experimental Techniques That Facilitate Human Granuloma Formation in an In Vitro System: A Systematic Review" Cells 11, no. 5: 864. https://doi.org/10.3390/cells11050864
APA StyleGanesan, N., Ronsmans, S., Vanoirbeek, J., & Hoet, P. H. M. (2022). Assessment of Experimental Techniques That Facilitate Human Granuloma Formation in an In Vitro System: A Systematic Review. Cells, 11(5), 864. https://doi.org/10.3390/cells11050864